

Abstract
Background:
Curcumin and ellagic are the natural polyphenols having a wide range of pharmacological actions. They have been reported to have their use in various neurological disorders.
Objective:
This study was aimed to evaluate the effect of curcumin and ellagic acid on the activity of monoamine oxidase (MAO), the enzyme responsible for metabolism of monoamine neurotransmitters which are pivotal for neuronal development and function.
Materials and Methods:
The in vitro effects of these selected polyphenols on MAO activities in mitochondria isolated from rat brains were examined. Brain mitochondria were assayed for MAO type-B (MAO-B) using benzylamine as substrates. Rat brain mitochondrial MAO preparation was used to study the kinetics of enzyme inhibition using double reciprocal Lineweaver–Burk plot.
Results:
MAO activity was inhibited by curcumin and ellagic acid; however, higher half maximal inhibitory concentrations of curcumin (500.46 nM) and ellagic acid (412.24 nM) were required compared to the known MAO-B inhibitor selegiline. It is observed that the curcumin and ellagic acid inhibit the MAO activity with both the competitive and noncompetitive type of inhibitions.
Conclusions:
Curcumin and ellagic acid can be considered a possible source of MAO inhibitor used in the treatment of Parkinson's and other neurological disorders.
SUMMARY
Monoamine oxidase (MAO) is involved in a variety of neurological disorders including Parkinson's disease (PD)
Curcumin and ellagic acid inhibit the monoamine oxidase activity
Ellagic acid revealed more potent MAO type-B (MAO-B) inhibitory activity than curcumin
Kinetic studies of MAO inhibition using different concentrations of curcumin and ellagic acid were plotted as double reciprocal Lineweaver–Burk plot
The mode of inhibition of both compounds toward MAO-B is mixed (competitive and uncompetitive) type of inhibition with both the competitive and noncompetitive type of inhibitions.
Abbreviations used: MAO: Monoamine oxidase, IC50: Higher half maximal inhibitory concentrations, PD: Parkinson's disease, LB: Lewy bodies, SNpc: Substantia nigra pars compacta, ROS: Reactive oxygen species, SG: Selegiline, DMC: demethoxycurcumin, BDMC: Bisdemethoxycurcumin.
Keywords: Curcumin, ellagic acid, enzyme, inhibition, kinetics, monoamine oxidase-B
INTRODUCTION
Parkinson's disease (PD) is the most common neurodegenerative disorder, affecting 1% of the population at the age of 65 and 4–5% of the population of the age of 85.[1,2] PD is pathologically characterized by the loss of dopaminergic (DAergic) neurons and presence of intraneuronal cytoplasmic inclusions, termed “Lewy Bodies” in the substantia nigra pars compacta as well as in certain other brain nuclei.[3] The symptoms of the disease are tremor or trembling in hands, arms, legs, jaw, and face; rigidity or stiffness of the limbs and trunk; bradykinesia or slowness of movement; and postural instability or impaired balance and coordination.[4] Despite the known pathology, the etiology of this disease and the mechanisms involved in the neurodegeneration are still elusive.[5]
There are various proposed mechanisms for the appearance and development of PD, including infection and inflammation, mitochondrial or proteasomal dysfunction, abnormal protein structures, dysregulation of heavy metals, and oxidative stress.[6] Generation of reactive oxygen species (ROS) may be a leading factor causing cell death[7] and other pathological aspects of PD,[8] however, whether ROS is the cause of PD or a consequence of the disease process itself, is still unknown.[9] One reason is that free radicals/toxic particles normally deactivated in the body are responsible, which can be controlled by antioxidants as adjuvant with dopamine agonist or monoamine oxidase (MAO) inhibitors.[10]
MAO are responsible for the oxidative deamination of a variety of neurotransmitters (e.g., noradrenaline, dopamine, and serotonin) as well as different exogenous and endogenous amines (i.e., tyramine and benzylamine) to their corresponding aldehydes.[11] Identification of MAO inhibitors is of great interest in drug discovery. Recent efforts toward the development of MAO inhibitors are focused on selective MAO type-A (MAO-A) or MAO type-B (MAO-B) inhibitors.[12] Selective MAO-A inhibitors are effective in the treatment of depression, whereas MAO-B inhibitors are logical application to the DAergic deficit in PD, depression, and Alzheimer's disease.[13,14] Evaluation of naturals, botanicals, and dietary supplements for the MAO inhibitory constituents is considered important for improving their use and supporting their traditional use in treatment of depression, PD, and other neuropsychiatric as well as neurological disorders.[15] The natural polyphenols and their combinations constituted a large group of phytochemicals in herbal beverages, foods, fruits, and vegetables worldwide are suggested to have the optimal neuroprotective effect. However, little information is available on the possible synergistic and/or antagonistic interactions of the various polyphenols and their combinations.[16]
Ellagic acid, 2, 3, 7, 8-tetrahydroxy-chromeno [5, 4, 3-cde] chromene-5, 10-dione, is a naturally occurring powerful bioactive compound found in a range of plant species, especially fruits. It has potential pharmacological and industrial applications.[17] Ellagic acid is present in plants in the form of hydrolyzable tannins called ellagitannins. These are the structural components of the plant cell wall and the cell membrane. Ellagitannins are hydrolysable molecules with hydroxybenzoic acid components.[18] The highest concentration of ellagic acid is found in many berries including strawberries, raspberries, cranberries, and grapes. Other sources of ellagic acid include walnuts, pecans, and beverages. Current research has revealed that ellagic acid possesses wide range of pharmacological properties such as antimutagenic, antioxidant, and anti-inflammatory activity in bacterial and mammalian systems.[19]
Curcumin is the chief, active curcuminoid of the dietary spice found in the rhizomes of Curcuma longa (turmeric), a plant in the ginger family (Zingiberaceae). Turmeric has been consumed for medicinal purposes for thousands of years. Extensive research on curcumin over the past few decades has revealed the health benefits of this ingredient to the modern era. The curcuminoids are a mixture of three principal compounds: Curcumin (curcumin I; 77%), demethoxycurcumin (DMC; curcumin II; 17%), and bis DMC (BDMC; curcumin III; 3%). The chemical name of curcumin is 1, 7-bis-(4-hydroxy-3-methoxyphenyl)-hepta-1,6-diene-3,5-dione; the chemical formula is C21H20O6, and pKa value is 8.54.[20,21]
Selegiline (SG [(R)-(-)-N, α-dimethyl-N-2-propynyl-phenethylamine]) is a potent, irreversible, and selective inhibitor of MAO-B. It has been widely prescribed alone or in combination with levodopa in the treatment of PD.[22]
The present study was intended to investigate effects of the ellagic acid and curcumin on MAO-B and their kinetic inhibition. The in vitro effects of these selected polyphenols on MAO activities in mitochondria isolated from rat brains were examined. Brain mitochondria were assayed for MAO-B using benzylamine as substrates. Finally, we studied changes in kinetic parameters of MAO activity in the presence of different concentrations of ellagic acid and curcumin to determine the mechanism of inhibition.
MATERIALS AND METHODS
Chemicals
Benzylamine was obtained from S.D. Fine Chemicals Ltd. Mumbai; mannitol, ethylenediaminetetraacetic acid disodium salt (EDTA), dimethyl sulfoxide (DMSO), and SG were obtained from Sigma Chemical (St Louis, MO, USA). All other chemicals used were of AR grade and were purchased from commercial sources.
Preparation of rat brain mitochondrial monoamine oxidase
Crude rat brain mitochondria were isolated as per the earlier described method of Gorgun et al.[23] In brief, the brain tissue was homogenized in the buffer of pH 7.4 containing 0.3 M mannitol and 0.1 mM EDTA. Homogenate was centrifuged at 600 g for 10 min at −4°C (R-248Mof CPR-24 plus Instrument, Remi, India). To obtain the brain mitochondria, the supernatant was collected and centrifuged at 10,000 g for 10 min at −4°C. The obtained mitochondrial pellets were washed 3 times with 0.25 M sucrose buffer containing 0.1 mM EDTA, resuspended in 0.25 M sucrose buffer, pH 7.4, and stored at −4°C for further studies.
Determination of protein, assay, and activity measurements of monoamine oxidase
The protein concentration and assay of MAO activity measurement with different concentrations (10–300 nM) of ellagic acid and curcumin were performed.[4] In brief, the reaction mixture contained 0.1 M phosphate buffer of pH 7.2, 10 mM benzylamine, MAO preparation, and different concentration of curcumin and ellagic acid (10–300 nM) in a total reaction volume of 200 μL in a 96-well plate. After 15 min of incubations at room temperature, readings were taken in at 250 nm.[24]
The effect of the selected constituents on MAO-B was explored by using the benzylamine deamination assay on 96-well plates.[25] A fixed substrate concentration and varying inhibitor concentrations (10–300 nM) were used to determine the higher half maximal inhibitory concentrations (IC50) value at the point where 50% inhibition of the catalytic activity of the enzyme occurred. For MAO-B, the substrate concentration of 10 mM benzylamine was chosen. The assay was performed wi01th the addition of inhibitor under study. Inhibition was calculated as percent of product formation compared to the corresponding control (enzyme-substrate reaction) without the inhibitors. We also performed separate controls in which the enzyme preparation is replaced by substrate, at the concentration used in the assay. The reactions were carried out in 0.1 M potassium phosphate buffer at pH 7.2. Incubations mixtures contained 100 μL of MAO preparation. The inhibitor was dissolved in buffer. The total reaction volume was 200 μL yielding a final DMSO concentration of 1.0% in the reaction mixture. The reaction mixtures were preincubated for 10 min at 37°C followed by the addition of MAO-B to initiate the reactions. Reactions were incubated for 15 min at 37°C and were determined fluorometrically, a flexible monochromator-based multi-mode microplate reader (Synergy HT, Bio-Tek) at 250 nm by using the GraphPad Prism 5.0 Version for Windows, GraphPad Software (San Diego, CA, USA) data analysis program.
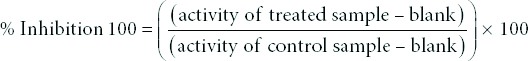
Kinetics of monoamine oxidase inhibition
To understand the nature of MAO inhibition, sets of three concentrations of curcumin and ellagic acid (200, 400, 600 nM) were used and compared with the kinetic behavior of a control set. For determination of Km and Vmax values for inhibition of MAO-B with curcumin and ellagic acid, the assays were carried out at different enzyme concentration and total volume of reaction mixture was kept constant but the substrate (benzylamine) concentration varied (5, 10, and 15 mM). Controls without inhibitor were also run simultaneously. The results are presented as double reciprocal Lineweaver–Burk plots and the kinetic data namely Km and Vmax values.
RESULTS
Determination of monoamine oxidase inhibition and higher half maximal inhibitory concentrations values
Curcumin and ellagic acid were evaluated in vitro for MAO-B inhibitory activity against rat brain MAO-B, whereby the ellagic acid demonstrated more potent MAO-B inhibitory activity than curcumin [Table 1]. The inhibition of MAO-B by curcumin (IC50 500.46 nM) and ellagic acid (IC50 412.24 nM) was found to be 10-fold less potent compared to the inhibition of standard SG (IC50 65.50 nM) [Table 1 and Figure 1].
Table 1.
Inhibition of monoamine oxidase type-B activity in brain crude mitochondrial fraction
Figure 1.
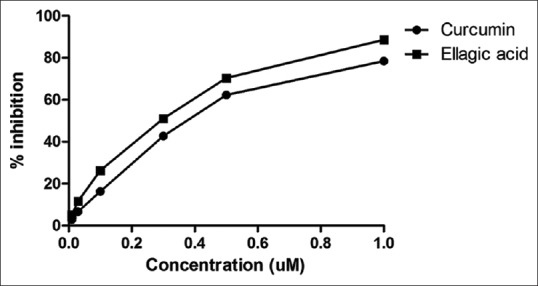
Concentration-dependent inhibition of rat brain monoamine oxidase-B by curcumin and ellagic acid. The activity was expressed as % inhibition. Each point represents mean ± standard deviation of triplicate values
Evaluation of inhibition mechanism and kinetics
Detailed in vitro studies were carried out to understand the kinetics and mechanism of inhibition of rat brain MAO-B by curcumin and ellagic acid. Both curcumin and ellagic acid were tested against MAO-B at varying concentrations of benzylamine, a selective substrate, to investigate the nature of inhibition of the enzymes. Based on dose-response inhibition, two concentrations of the inhibitors were selected, one below and another above IC50 value for the inhibition. For each experiment, three sets of assays were done at varying concentrations of the substrate, one control without inhibitors and with two concentrations of the inhibitor. The enzyme kinetics data are presented as double reciprocal Lineweaver–Burk plots [Figure 2]. Binding of curcumin and ellagic acid with rat brain MAO-B increase the Km value (the Michaelis–Menten constant) with no apparent effect on the Vmax, indicating that the inhibition of MAO-B by curcumin (noncompetitive) and ellagic acid (competitive) is mixed type of inhibition [Figure 2].
Figure 2.

Lineweaver-Burk plots of inhibition of rat brain monoamine oxidase-B with (a) curcumin (200–600 nM) and (b) ellagic acid (200–600 nM); V: nmoles/min/mg protein; S: substrate benzylamine concentration (mM)
DISCUSSION
In recent years, increasing attention has been devoted to the management or prevention of PD by herbal medicines. Recent studies indicated that the active components of herbal medicines, herbal extracts, and herbal formulations have effect on different in vitro and in vivo PD models.[26]
The present advances in pharmacokinetics and new drug development have shown the possibility of selective inhibition of brain MAO as compared to that in periphery, thus increasing the clinical potential of these drugs. SG and rasagiline are propargyl derivatives that have potent neuroprotective action and selective MAO-B inhibitory properties. These drugs proposed to be main discussion about drug treatment of PD.[27]
MAOs are flavoproteins which catalyze the oxidative deamination of primary, secondary, and tertiary amines. The primary functions of MAOs are the metabolism of exogenous amines and the regulation of neurotransmitter level and intracellular amine stores. MAO-B preferentially deaminates phenylethylamine, benzylamine, and dopamine. Thus, inhibition of MAO-B may alleviate symptoms of PD.[15,28]
Curcumin, an active ingredient isolated from Curcuma longa (L.), has been showed to possess extensive pharmacological activities, including anticardiovascular disorders, antitumor activities, cholesterol-lowering, antidiabetic, anti-inflammatory, and myelodysplastic syndromes, and anti-nerve degeneration. However, molecular mechanisms of curcumin-mediated anti-PD are still unknown.[21]
The present study reveals that curcumin and ellagic acid can inhibit MAO activity. The inhibitory potency and mechanism of inhibition of curcumin and ellagic acid toward rat brain MAO-B were assessed. The results indicate that some effects of these selected polyphenols on brain functions could be related to a nonreceptor modulation of monoaminergic neurotransmission.
Inhibitory and kinetic parameters calculated in this article result from total concentrations of curcumin and ellagic acid added to crude brain mitochondrial fraction. The situation in various tissues in vivo may be different from our experimental conditions. This was the reason why the effects of curcumin and ellagic acid on MAO's activity were compared with the effect of a well-known MAO inhibitor such as SG.[12] The significant MAO inhibitory activity of curcumin and ellagic acid as testified herein suggest additional potential utility of these two polyphenols for alleviation of neurological disorders associated with depletion of monoamines, the most important of which are depression and PD.[29]
The results of the kinetic studies of MAO inhibition using different concentrations (200–600 nM) of curcumin and ellagic acid were plotted as double reciprocal Lineweaver–Burk plot [Figure 2]. The mode of inhibition of both compounds toward MAO-B is mixed (competitive and uncompetitive) type of inhibition. The Vmax (8.40 nM/min) remained constant while the Km varied from 16.66 ± 1.2, 28.57 ± 1.1, and 40.0 ± 1.4 mM for 200–600 μg/ml concentration of ellagic acid as compared to control (9.53 ± 2.10 mM) while for curcumin, Km (6.25 mM) remains constant and Vmax varied from 11.76 ± 1.86, 5.26 ± 2.11, and 3.77 ± 1.26 mM for 200–600 μg/ml concentration of curcumin as compared to control (22.22 ± 1.34 mM). The kinetic constant Km which is basically a substrate concentration signifies the extent of affinity of an enzyme with its substrate. The calculated Km values of the present investigation also signify that with increase in the concentration of curcumin and ellagic acid, there is corresponding increase in Km indicating the decrease in affinity of MAO for its substrate. The decrease in Ki in relation to increase in concentration of curcumin and ellagic acid also indicates the effective inactivation of MAO at higher concentrations.
Inhibitors of MAO-B not only lead to enhanced DAergic neurotransmission but also prevent activation of toxin and free radical formation, and they alleviated the process of neuron denaturalization. The phytochemicals showed that there exists a structural similarity between natural polyphenolic compounds and established cholinesterase inhibitors in terms of molecular weight, phenolic rings, and hydrophobic component are described as competitive inhibitors of MAO.[27,30,31]
CONCLUSIONS
In the present study, we have evaluated kinetics of inhibition of MAO using curcumin and ellagic acid. The results of present reveal that both curcumin and ellagic acid inhibit the MAO activity with both the competitive and noncompetitive type of inhibitions. Curcumin and ellagic acid can be considered as a possible source of MAO inhibitor used in the treatment of Parkinson's and other neurological disorders. Although it is always difficult to extrapolate from in vitro studies to the clinical reality, the present results suggest that the examined polyphenols might contribute to an inhibitory effect on MAO, and a systematic in vivo investigation of the curcumin and ellagic acid effect on MAO activity is needed. The exact physiological role of MAO-B inhibition by curcumin and ellagic acid is not known; however, their effects on MAO activity may contribute to nonreceptor actions of curcumin and ellagic acid participating in modulation of monoamine neurotransmission in the brain.
Financial support and sponsorship
Author would like to acknowledge financial support from University Grant Commission (UGC), New Delhi, India to carry out the research work.
Conflicts of interest
There are no conflicts of interest.
ABOUT AUTHOR

Archana Ramesh Juvekar
Prof. Mrs. Archana Ramesh Juvekar is Professor of Pharmacology and Physiology at the Department of Pharmaceutical Sciences and Technology, Institute of Chemical Technology, Mumbai. Her research interest is in the area of preclinical pharmacodynamics activity evaluation in diseases related to inflammation, immunomodulation, hepatoprotective, central nervous system, cardiovascular system diseases, toxicology - acute, subacute, and chronic toxicity studies. She is a life member of Indian Pharmaceutical Association, Indian Pharmacological Society, and a member of Gesellschaft für Arzneipflanzenforschung Society for Medicinal Plant Research, Germany, Editorial Board of Indian Practitioner, Radiopharmaceuticals Committee under Board of Radiation and Isotope Technology.
REFERENCES
- 1.Khalil WK, Assaf N, ElShebiney SA, Salem NA. Neuroprotective effects of bee venom acupuncture therapy against rotenone-induced oxidative stress and apoptosis. Neurochem Int. 2015;80:79–86. doi: 10.1016/j.neuint.2014.11.008. [DOI] [PubMed] [Google Scholar]
- 2.Patil SP, Jain PD, Sancheti JS, Ghumatkar PJ, Tambe R, Sathaye S. Neuroprotective and neurotrophic effects of apigenin and luteolin in MPTP induced Parkinsonism in mice. Neuropharmacology. 2014;86:192–202. doi: 10.1016/j.neuropharm.2014.07.012. [DOI] [PubMed] [Google Scholar]
- 3.Aluf Y, Vaya J, Khatib S, Loboda Y, Finberg JP. Selective inhibition of monoamine oxidase A or B reduces striatal oxidative stress in rats with partial depletion of the nigro-striatal dopaminergic pathway. Neuropharmacology. 2013;65:48–57. doi: 10.1016/j.neuropharm.2012.08.023. [DOI] [PubMed] [Google Scholar]
- 4.Khatri DK, Juvekar AR. Propensity of Hyoscyamus niger seeds methanolic extract to allay stereotaxically rotenone-induced Parkinson's disease symptoms in rats. Orient Pharm Exp Med. 2015;15:327–39. [Google Scholar]
- 5.Mitra S, Chakrabarti N, Dutta SS, Ray S, Bhattacharya P, Sinha P, et al. Gender-specific brain regional variation of neurons, endogenous estrogen, neuroinflammation and glial cells during rotenone-induced mouse model of Parkinson's disease. Neuroscience. 2015;292:46–70. doi: 10.1016/j.neuroscience.2014.12.052. [DOI] [PubMed] [Google Scholar]
- 6.Nazari QA, Takada-Takatori Y, Hashimoto T, Imaizumi A, Izumi Y, Akaike A, et al. Potential protective effect of highly bioavailable curcumin on an oxidative stress model induced by microinjection of sodium nitroprusside in mice brain. Food Funct. 2014;5:984–9. doi: 10.1039/c4fo00009a. [DOI] [PubMed] [Google Scholar]
- 7.Khatri DK, Juvekar P, Juvekar A. Phytochemical investigation and in vitro antioxidant activities indigofera cordifolia seed extracts. Int J Pharm Pharm Sci. 2013;5:71–5. [Google Scholar]
- 8.Gokul K. Oral supplements of aqueous extract of tomato seeds alleviate motor abnormality, oxidative impairments and neurotoxicity induced by rotenone in mice: Relevance to Parkinson's disease. Neurochem Res. 2014;39:1382–94. doi: 10.1007/s11064-014-1323-1. [DOI] [PubMed] [Google Scholar]
- 9.Beppe GJ, Dongmo AB, Foyet HS, Tsabang N, Olteanu Z, Cioanca O, et al. Memory-enhancing activities of the aqueous extract of Albizia adianthifolia leaves in the 6-hydroxydopamine-lesion rodent model of Parkinson's disease. BMC Complement Altern Med. 2014;14:142. doi: 10.1186/1472-6882-14-142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tarazi FI, Sahli ZT, Wolny M, Mousa SA. Emerging therapies for Parkinson's disease: From bench to bedside. Pharmacol Ther. 2014;144:123–33. doi: 10.1016/j.pharmthera.2014.05.010. [DOI] [PubMed] [Google Scholar]
- 11.Singh R, Ramakrishna R, Bhateria M, Bhatta RS. In vitro evaluation of Bacopa monniera extract and individual constituents on human recombinant monoamine oxidase enzymes. Phytother Res. 2014;28:1419–22. doi: 10.1002/ptr.5116. [DOI] [PubMed] [Google Scholar]
- 12.Fisar Z, Hroudová J, Raboch J. Inhibition of monoamine oxidase activity by antidepressants and mood stabilizers. Neuro Endocrinol Lett. 2010;31:645–56. [PubMed] [Google Scholar]
- 13.Riederer P, Laux G. MAO-inhibitors in Parkinson's disease. Exp Neurobiol. 2011;20:1–17. doi: 10.5607/en.2011.20.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cai Z. Monoamine oxidase inhibitors: Promising therapeutic agents for Alzheimer's disease (Review) Mol Med Rep. 2014;9:1533–41. doi: 10.3892/mmr.2014.2040. [DOI] [PubMed] [Google Scholar]
- 15.Chaurasiya ND, Ibrahim MA, Muhammad I, Walker LA, Tekwani BL. Monoamine oxidase inhibitory constituents of propolis: Kinetics and mechanism of inhibition of recombinant human MAO-A and MAO-B. Molecules. 2014;19:18936–52. doi: 10.3390/molecules191118936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ngoungoure VL, Schluesener J, Moundipa PF, Schluesener H. Natural polyphenols binding to amyloid: A broad class of compounds to treat different human amyloid diseases. Mol Nutr Food Res. 2015;59:8–20. doi: 10.1002/mnfr.201400290. [DOI] [PubMed] [Google Scholar]
- 17.García-Niño WR, Zazueta C. Ellagic acid: Pharmacological activities and molecular mechanisms involved in liver protection. Pharmacol Res. 2015;97:84–103. doi: 10.1016/j.phrs.2015.04.008. [DOI] [PubMed] [Google Scholar]
- 18.Teixeira Lde L, Bertoldi FC, Lajolo FM, Hassimotto NM. Identification of ellagitannins and flavonoids from Eugenia brasilienses Lam. (Grumixama) by HPLC-ESI-MS/MS. J Agric Food Chem. 2015;63:5417–27. doi: 10.1021/acs.jafc.5b01195. [DOI] [PubMed] [Google Scholar]
- 19.Huerga-González V, Lage-Yusty MA, Lago-Crespo M, López-Hernández J. Comparison of methods for the study of ellagic acid in pomegranate juice beverages. Food Anal Methods. 2015;8:2286–93. [Google Scholar]
- 20.Alexander A, Qureshi A, Kumari L, Vaishnav P, Sharma M, et al. Role of herbal bioactives as a potential bioavailability enhancer for active pharmaceutical ingredients. Fitoterapia. 2014;97:1–14. doi: 10.1016/j.fitote.2014.05.005. [DOI] [PubMed] [Google Scholar]
- 21.Prasad S, Gupta SC, Tyagi AK, Aggarwal BB. Curcumin, a component of golden spice: From bedside to bench and back. Biotechnol Adv. 2014;32:1053–64. doi: 10.1016/j.biotechadv.2014.04.004. [DOI] [PubMed] [Google Scholar]
- 22.Robakis D, Fahn S. Defining the role of the monoamine oxidase-B inhibitors for Parkinson's disease. CNS Drugs. 2015;29:433–41. doi: 10.1007/s40263-015-0249-8. [DOI] [PubMed] [Google Scholar]
- 23.Gorgun FM, Zhuo M, Singh S, Englander EW. Neuroglobin mitigates mitochondrial impairments induced by acute inhalation of combustion smoke in the mouse brain. Inhal Toxicol. 2014;26:361–9. doi: 10.3109/08958378.2014.902147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gacche RN, Shaikh RU, Chapole SM, Jadhav AD, Jadhav SG. Kinetics of inhibition of monoamine oxidase using Cymbopogon martinii (Roxb.) Wats: A potential antidepressant herbal ingredient with antioxidant activity. Indian J Clin Biochem. 2011;26:303–8. doi: 10.1007/s12291-011-0124-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kotelnikova RA, Smolina AV, Grigoryev VV, Faingold II, Mischenko DV, Rybkin AY, et al. Influence of water-soluble derivatives of[60] fullerene on therapeutically important targets related to neurodegenerative diseases. Med Chem Commun. 2014;5:1664–8. [Google Scholar]
- 26.Li XZ, Zhang SN, Liu SM, Lu F. Recent advances in herbal medicines treating Parkinson's disease. Fitoterapia. 2013;84:273–85. doi: 10.1016/j.fitote.2012.12.009. [DOI] [PubMed] [Google Scholar]
- 27.Finberg JP. Update on the pharmacology of selective inhibitors of MAO-A and MAO-B: Focus on modulation of CNS monoamine neurotransmitter release. Pharmacol Ther. 2014;143:133–52. doi: 10.1016/j.pharmthera.2014.02.010. [DOI] [PubMed] [Google Scholar]
- 28.Coelho-Cerqueira E, Netz PA, do Canto VP, Pinto AC, Follmer C. Beyond topoisomerase inhibition: Antitumor 1, 4-naphthoquinones as potential inhibitors of human monoamine oxidase. Chem Biol Drug Des. 2014;83:401–10. doi: 10.1111/cbdd.12255. [DOI] [PubMed] [Google Scholar]
- 29.Yang J, Song S, Li J, Liang T. Neuroprotective effect of curcumin on hippocampal injury in 6-OHDA-induced Parkinson's disease rat. Pathol Res Pract. 2014;210:357–62. doi: 10.1016/j.prp.2014.02.005. [DOI] [PubMed] [Google Scholar]
- 30.Ademosun AO, Oboh G. Comparison of the inhibition of monoamine oxidase and butyrylcholinesterase activities by infusions from green tea and some citrus peels. Int J Alzheimers Dis. 2014;2014:586407. doi: 10.1155/2014/586407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Viña D, Serra S, Lamela M, Delogu G. Herbal natural products as a source of monoamine oxidase inhibitors: A review. Curr Top Med Chem. 2012;12:2131–44. doi: 10.2174/156802612805219996. [DOI] [PubMed] [Google Scholar]