

ABSTRACT
The essential micronutrient vitamin B6 is best known in its enzymatic cofactor form, pyridoxal 5′-phosphate (PLP). However, vitamin B6 comprises the amine pyridoxamine 5′-phosphate (PMP) and the alcohol pyridoxine 5′-phosphate (PNP) in addition to PLP, as well as their corresponding non-phosphorylated forms. The different B6 forms (called vitamers) are enzymatically interconverted in a ubiquitous salvage pathway. Recently, we have shown that balancing the ratio of the different B6 vitamers in particular PMP by the PMP/PNP oxidase PDX3 is essential for growth and development in Arabidopsis thaliana. Intriguingly, nitrate to ammonium conversion is impaired in pdx3 mutants, such that the mutants become ammonium-dependent, suggesting an interaction between vitamin B6 and nitrogen metabolism. In addition, we found a strong up-regulation of genes related to plant defense. Here, we further show that pdx3 mutants display a temperature-sensitive phenotype that is typical of autoimmune mutants and is possibly connected to the impaired nitrogen metabolism.
KEYWORDS: Arabidopsis thaliana, autoimmunity, nitrogen metabolism, PDX3, plant defense, pyridoxamine/pyridoxine phosphate oxidase, vitamin B6 salvage, vitamin B6
The essential water-soluble vitamin, B6, comprises 6 different vitamers, the alcohol pyridoxine (PN), the amine pyridoxamine (PM), the aldehyde pyridoxal (PL) and their respective 5′ phosphorylated forms (PNP, PMP, PLP).1 The enzymatic cofactor PLP is essential for more than 140 enzymatic reactions (http://bioinformatics.unipr.it/cgi-bin/bioinformatics/B6db/home.pl). While its biosynthesis de novo exclusively occurs in plants, bacteria and fungi; humans and animals have to take it up from their diet. Independent from biosynthesis de novo, most organisms possess a salvage pathway through which the different B6 vitamers are interconverted enzymatically. In plants, the main salvage pathway enzymes are the kinase SALT OVERLY SENSITIVE 4 (SOS4) 2,3) that phosphorylates all non-phosphorylated vitamers and the oxidase PDX3 that recycles PLP from PMP and PNP.4,5
Recently, we showed that mutants knocked out in PDX3, pdx3-3 (SALK_054167C) and pdx3-4 (GK-260E03), are strongly impaired in vegetative and reproductive development.6 In addition to aberrant leaf growth, the strong allele (pdx3-3) flowers early, is dwarf and has reduced seed yield.6 At the biochemical level, these pdx3 mutants accumulate high amounts of PMP and show a strongly reduced activity for nitrate reductase (NR) compared to the wild-type.6 They thus become dependent on an exogenous supply of ammonium as a nitrogen source, which in turn rescues the developmental phenotype of the mutants. Moreover, we demonstrated that the levels of the vitamer PMP respond (i.e. increase) to feeding with ammonium, the precise molecular nature of which remains to be deciphered.6 Together these results led to the current working model, in which PMP accumulation is suggested to signal ammonium availability to the plant and consequently inhibit NR activity (directly or indirectly) preventing the further reduction of nitrate into ammonium.6 In the pdx3 mutants, the constitutive accumulation of PMP likely inadvertently signals ammonium availability to the plant and thus leads to inappropriate NR inhibition.6
In addition to the described impairment in nitrogen metabolism, we observed an overrepresentation of biotic-stress related genes among the set of upregulated genes in a transcriptome analysis of pdx3-3 and pdx3-4.6 Interestingly, this coincides with an increase in salicylic acid (SA) content implying that these pdx3 mutants are hyperactivated for defense. Indeed, key genes in the SA-mediated defense pathway, such as PATHOGENESIS RELATED-1 (PR1), ENHANCED DISEASE SUSCEPTIBILITY1 (EDS-1) and SUPPRESSOR OF npr1-1, CONSTITUTIVE1 (SNC1) were strongly upregulated (Fig. 1A).6 In further support of defense activation in these pdx3 mutants, there is enrichment for those genes encoding proteins localized to the plasma membrane or extracellular space among the upregulated genes. Plant defense and rearrangements of the cell wall and plasma membrane are commonly associated.7
Figure 1.
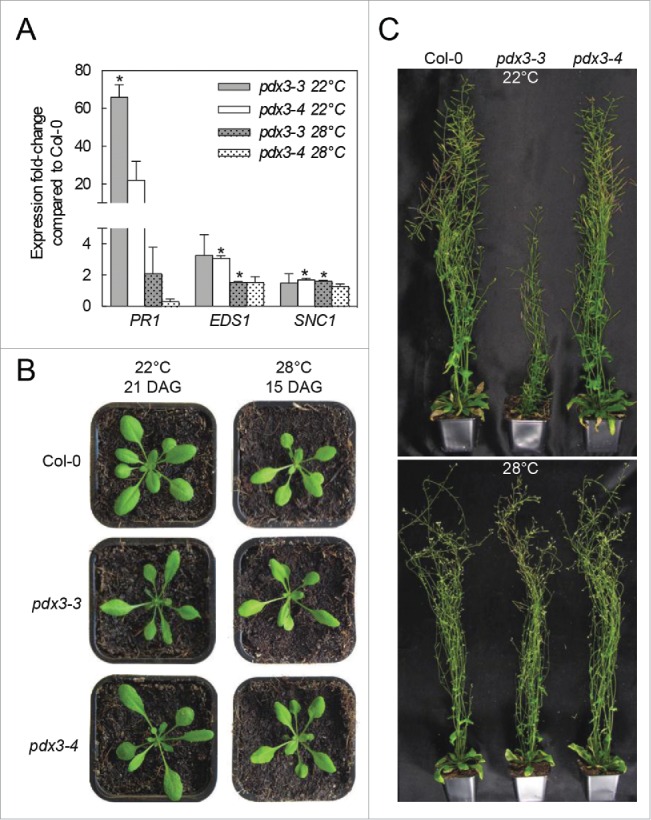
The vegetative and reproductive developmental phenotypes of pdx3 are abrogated by elevated temperature. (A) Expression analysis of selected key genes involved in salicylic acid mediated defense of 21-day-old plants grown on soil at either 22°C or 28°C. The data are presented as means ± SE for 3 biological replicates. Statistically significant changes compared to wild-type under the respective condition were calculated by a 2 tailed Student's t-test for P < 0.05 and are indicated by an asterisk. (B) Vegetative stage phenotype of pdx3-3 and pdx3-4 compared to wild-type grown either at 22°C (21-day-old), or at 28°C (15-day-old). The different ages were chosen to represent equivalent developmental stages under the respective conditions. (C) Reproductive stage phenotype of pdx3-3 and pdx3-4 compared to wild-type grown either at 22°C (upper panel) or at 28°C (lower panel). Pictures were captured of 39-day old plants. In all cases plants were grown on soil under a 16-hour photoperiod (120 μmol photons.m−2 s−1) and 8 hours of darkness either at 22°C or 28°C as indicated. The data at 22°C in (A) and (C) have been reproduced from Colinas, M., Eisenhut, M., Tohge, T., Pesquera, M., Fernie, A.R., Weber, A.P.M., and Fitzpatrick, T.B. (2016). Balancing of B6 vitamers is essential for plant development and metabolism in Arabidopsis. Plant Cell: In press. tpc.15.01033v1-TPC2015-01033; www.plantcell.org; Copyright American Society of Plant Biologists.
The finding that the SA-mediated defense pathway was highly activated at the transcriptional and metabolite level in pdx3-3 and pdx3-4 prompted us to investigate whether the observed vegetative and reproductive phenotype could also be associated with this immune response. Mutants showing an inappropriate activation of defense responses are often referred to as “autoimmune” mutants and generally endure a fitness cost with reduced biomass and seed yield.8,9 Significantly, this autoimmune phenotype can be suppressed by moderately high temperature or high humidity.8 Here, we report a comparison of the performance of pdx3-3 and pdx3-4 when the plants are submitted to growth at 28°C to that previously reported when grown at 22°C (Fig. 1).6 The elevated temperature indeed led to a considerable attenuation, if not complete recovery, of the developmental phenotypes observed at 22°C and was similar to wild-type under the same conditions (Fig. 1B-C). Specifically, leaf shape resembled that of wild-type during the vegetative and reproductive phases at 28°C and is in contrast to the aberrant leaf shape observed in the pdx3-3 and pdx3-4 mutants at 22°C (Fig. 1B). Moreover, the previously observed late flowering phenotype, reduced apical dominance and increased number of primary stems at 22°C was not observed at 28°C (Fig. 1C). Furthermore, the strong upregulation of the defense marker genes PR1 and EDS1 observed at 22°C in these pdx3 mutants, is substantially abrogated in plants grown at 28°C (Fig. 1A). Together these results suggest that pdx3 in addition to being ammonium dependent has characteristics of a temperature-sensitive autoimmune mutant.
The defense response associated with mutation of pdx3 was not anticipated and could suggest that PDX3 contributes to the negative regulation of such responses. However, we have shown in our previous study that pdx3 mutants are ammonium dependent,6 thus the link between nitrogen metabolism and the defense response remains a conundrum. Intriguingly, we could not observe an alteration in the expression of PDX3 upon treatment of plants with SA (data not shown) suggesting that the effects mediated by PDX3 on defense responses are upstream of SA biosynthesis. Nonetheless, it should be noted that autoimmunity and impaired nitrogen metabolism have been associated previously. Interestingly in this context, exogenous ammonium (but not nitrate) has been shown to alleviate autoimmune phenotypes.10,11 Moreover, perturbations in nitrogen metabolism may result in autoimmunity. For instance, a mutant in the ammonium transporter amt1.1 displays an autoimmune phenotype.12 Furthermore, loss of function of a positive regulator of NR, siz1, results in reduced NR activity and ammonium-dependent autoimmunity.10 While the underlying molecular mechanism between autoimmunity and nitrogen metabolism has not been deciphered, it has been suggested that plant pathogens preferentially use ammonium rather than nitrate as a nitrogen source.13 The expected lowering of the ammonium to nitrate ratio may thus signal pathogen invasion to the plant and consequently trigger a defense response. Our previously published study6 and the results presented here reinforce the notion that there is an interaction between nitrogen metabolism and immunity. Indeed, we propose to add the salvage pathway enzyme PDX3 as another component in this interaction. Disruption of PDX3 compromises nitrogen metabolism such that the plant becomes dependent on ammonium. In the absence of the latter nutrient, the plant responds by launching a salicylic acid-mediated defense pathway and suffers strong developmental impairments.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
The Swiss National Science Foundation is gratefully acknowledged for financial support to Teresa B. Fitzpatrick (Grant 31003A-141117/1) as well as the University of Geneva. Additional financial support is gratefully acknowledged from the European Molecular Biology Organization (ASTF 485-2014) to Maite Colinas. We thank the European Arabidopsis Stock Center (NASC) for seeds of SALK_054167C (pdx3-3) and GABI-KAT for GK-260E03 (pdx3-4).
References
- 1.Fitzpatrick TB, Amrhein N, Kappes B, Macheroux P, Tews I, Raschle T. Two independent routes of de novo vitamin B6 biosynthesis: not that different after all. Biochem J 2007; 407:1-13; PMID:17822383; http://dx.doi.org/ 10.1042/BJ20070765 [DOI] [PubMed] [Google Scholar]
- 2.Shi H, Xiong L, Stevenson B, Lu T, Zhu J-K. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell 2002; 14:575-88; PMID:11910005; http://dx.doi.org/ 10.1105/tpc.010417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Shi H, Zhu J-K. SOS4, a pyridoxal kinase gene, is required for root hair development in Arabidopsis. Plant Physiol 2002; 129:585-93; PMID:12068103; http://dx.doi.org/ 10.1104/pp.001982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sang Y, Barbosa JM, Wu H, Locy RD, Singh NK. Identification of a pyridoxine (pyridoxamine) 5′-phosphate oxidase from Arabidopsis thaliana. FEBS Lett 2007; 581:344-8; PMID:17224143; http://dx.doi.org/ 10.1016/j.febslet.2006.12.028 [DOI] [PubMed] [Google Scholar]
- 5.González E, Danehower D, Daub ME. Vitamer levels, stress response, enzyme activity, and gene regulation of Arabidopsis lines mutant in the pyridoxine/pyridoxamine 5′-phosphate oxidase (PDX3) and the pyridoxal kinase (SOS4) genes involved in the vitamin B6 salvage pathway. Plant Physiol 2007;145:985-96; PMID:17873088; http://dx.doi.org/ 10.1104/pp.107.105189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Colinas M, Eisenhut M, Tohge T, Pesquera M, Fernie AR, Weber AP, Fitzpatrick TB. Balancing of B6 vitamers is essential for plant development and metabolism in Arabidopsis. Plant Cell 2016. pii: TPC2015-01033-RA; 28:430-453; PMID:26858304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Malinovsky FG, Fangel JU, Willats WG. The role of the cell wall in plant immunity. Front Plant Sci 2014; 5:178; PMID:24834069; http://dx.doi.org/ 10.3389/fpls.2014.00178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Alcazar R, Parker JE. The impact of temperature on balancing immune responsiveness and growth in Arabidopsis. Trends Plant Sci 2011; 16:666-75; PMID:21963982; http://dx.doi.org/ 10.1016/j.tplants.2011.09.001 [DOI] [PubMed] [Google Scholar]
- 9.Todesco M, Balasubramanian S, Hu TT, Traw MB, Horton M, Epple P, Kuhns C, Sureshkumar S, Schwartz C, Lanz C, et al.. Natural allelic variation underlying a major fitness trade-off in Arabidopsis thaliana. Nature 2010; 465:632-6; PMID:20520716; http://dx.doi.org/ 10.1038/nature09083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Park BS, Song JT, Seo H. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat Commun 2011; 2:400; PMID:21772271; http://dx.doi.org/ 10.1038/ncomms1408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wang H, Lu Y, Liu P, Wen W, Zhang J, Ge X, Xia Y. The ammonium/nitrate ratio is an input signal in the temperature-modulated, SNC1-mediated and EDS1-dependent autoimmunity of nudt6-2 nudt7. Plant J 2013; 73:262-75; PMID:23004358; http://dx.doi.org/ 10.1111/tpj.12032 [DOI] [PubMed] [Google Scholar]
- 12.Pastor V, Gamir J, Camanes G, Cerezo M, Sanchez-Bel P, Flors V. Disruption of the ammonium transporter AMT1.1 alters basal defenses generating resistance against Pseudomonas syringae and Plectosphaerella cucumerina. Front Plant Sci 2014; 5; PMID:24910636; http://dx.doi.org/10.3389/fpls.2014.00231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Marzluf GA. Genetic regulation of nitrogen metabolism in fungi. Microbiol Mol Biol Rev 1997; 61:17-32; PMID:9106362 [DOI] [PMC free article] [PubMed] [Google Scholar]