

Abstract
It is well-known that the conversion of normal colon epithelium to adenoma and then to carcinoma stems from acquired molecular changes in the genome. The genetic basis of colorectal cancer has been elucidated to a certain extent, and much remains to be known about the identity of specific cancer genes that are associated with the advancement of colorectal cancer from one stage to the next. Here in this study we attempted to identify novel cancer genes that could underlie the stage-specific progression and metastasis of colorectal cancer. We conducted a stage-based meta-analysis of the voluminous tumor genome-sequencing data and mined using multiple approaches for novel genes driving the progression to stage-II, stage-III and stage-IV colorectal cancer. The consensus of these driver genes seeded the construction of stage-specific networks, which were then analyzed for the centrality of genes, clustering of subnetworks, and enrichment of gene-ontology processes. Our study identified three novel driver genes as hubs for stage-II progression: DYNC1H1, GRIN2A, GRM1. Four novel driver genes were identified as hubs for stage-III progression: IGF1R, CPS1, SPTA1, DSP. Three novel driver genes were identified as hubs for stage-IV progression: GSK3B, GGT1, EIF2B5. We also identified several non-driver genes that appeared to underscore the progression of colorectal cancer. Our study yielded potential diagnostic biomarkers for colorectal cancer as well as novel stage-specific drug targets for rational intervention. Our methodology is extendable to the analysis of other types of cancer to fill the gaps in our knowledge.
Introduction
Colorectal cancer is the third most common cancer in men, and the second in women worldwide according to International Agency for Research on Cancer, World Health Organization [1]. In recent years, the incidence of colorectal cancer has significantly increased in newly developed countries where the risk was once low. Despite the existence of screening and preventive strategies, colorectal cancer remains a major public health problem. Cancer mortality is significantly correlated with the stage of development of the cancer and could be reduced if cases are detected and treated early. The 5-year survival rate of colorectal cancer patients declines significantly with the stage of the cancer. Most importantly, the 5-year survival rate for patients with distant metastases is only 10%. Considering the fact that metastases are the cause of 90% of cancer deaths[2], there are no effective drugs available to curtail the process of cancer spreading to different organ systems. Treatment of metastases remains a major challenge, not least because our knowledge of factors responsible for cancer progression and metastases is far from complete. With an increased trend in incidence and death rates of colon cancer, and the unpredictability of factors responsible for the metastatic potential of the localized tumor, we prioritized our efforts towards identifying the genes responsible for colon cancer progression. Increasing our current knowledge of the genes and pathways that play important roles in the progression of cancer from one stage to the next could prove very useful in early-stage diagnosis as well as identifying targets for personalized cancer therapy.
We have earlier published on important roles of two genes on colon cancer, CELF2 a putative tumor suppressor gene [3,4] and RBM3 a proto-oncogene [5]. Here we have attempted to identify more novel and key genes underpinning colon cancer progression using the available data from the TCGA consortium [6]. Mutations in colon cancer are complex and unclear due to the presence of passenger and driver genes even within the same tumor. Much effort has focused towards identifying driver genes. The aim of the current study is to utilize methods of network analysis to identify novel biomarkers responsible for the colorectal cancer progression to each stage. The differential anatomical penetration of the cancer for each stage is shown in Fig 1.
Fig 1. Staging of colon cancer.

The American Joint Committee on Cancer (AJCC) has staged the colorectal cancer based on the anatomical extent of the disease. Stage I: Tumor that is limited to the mucosal layer (T1) or muscularis propria (T2), without involvement of any lymph node or distant metastatic organs. Stage II: Tumor that penetrates the muscularis propria (T3) or invades nearby organs or structures (T4), without involvement of any lymph node or distant metastatic organs. Stage III: Tumor stages with lymph node metastasis but without distant metastasis. Stage IV: Any tumor stage and lymph node status with distant organ metastasis.
Materials and Methods
Dataset
TCGA datasets annotated by the stage of cancer were retrieved from the DriverDB [7] by performing the following meta-analysis. We selected colon adenocarcinoma as the tissue of interest, and specified ‘tumor stage’ as the clinical criteria. We obtained datasets for each stage of colon adenocarcinoma, namely stage I, stage II, stage III, and stage IV of colon adenocarcinoma.
Identification of consensus driver genes
Framing the stage of tumor as the unit of analysis, we used the following tools to identify driver genes: ActiveDriver[8], Dendrix[9], MDPFinder[10], Simon[11], Netbox[12], OncodriveFM[13], MutSigCV [14], and MEMo [15]. To obtain the consensus driver genes, we determined the overlap between the predictions of the various tools for a given stage. The selective advantage conferred by driver genes to the growth of tumor cells could be either gain of function or loss of function events (for e.g., oncogenes are gain-of-function and insensitivity to tumor-suppressor is a loss of function). We filtered for driver genes that were identified by at least three tools and obtained the consensus prediction of driver genes for each stage.
Novel driver genes
To identify novel driver genes, we subtracted the driver genes of stage I from the driver genes of stage II to ensure stage II-specific driver genes in the progression of cancer. In a similar manner, we obtained stage III-specific and stage IV-specific driver genes. To eliminate non-specific driver genes from the analysis, we screened each stage against a background of driver genes obtained from pooling all samples of colon adenocarcinoma regardless of stage of cancer. This set of non-redundant stage-specific driver genes was further screened against the Cancer Gene Census v68[16] to filter out any remaining known cancer genes. Thus we obtained novel and stage-specific driver gene sets for further analysis.
Network analysis
The construction and analysis of stagewise networks were aided by Cytoscape[17]. The driver gene sets identified above were used to seed the construction of the corresponding stage-specific network using the Genemania tool [18]. We searched for the following types of interactions of the stage driver genes: ‘physical’, ‘protein-protein interactions’ and ‘predicted’. This yielded stage-wise networks. To analyze the topological properties of each network, we used NetworkAnalyzer[19]. The degree distribution of each network was calculated and the goodness of fit with a power-law distribution was determined using the coefficient of determination (R2). A high R2 implied the existence of fat tails in the degree distribution, indicating that some genes played the role of hubs. Alteration of function of these genes due to mutation, translocation or copy number variation could result in deleterious genes damaging cellular activity. To analyze the structure of the stage-wise networks, we performed centrality analysis, modularity analysis and Gene Ontology analysis. Centrality analysis identified the central nodes in each stage-specific network by various metrics using Centiscape [20]. Three metrics of centrality were used to rank the genes, viz. the between-ness centrality, closeness centrality and bottleneck centrality. These metrics were chosen for their measurement of complementary properties of node importance. The top 15 genes from each measure were chosen, and their intersection was determined to yield a consensus set of central genes for each stage (Fig 2). These are the “hub” genes identified in our work and discussed individually below. This stage-specific consensus set was compared with the set of driver genes for each stage. A gene common to both sets is a driver and a hub. Such genes are termed ‘hub’ driver genes for each stage. We then analyzed the clustering pattern of the stage-wise networks using the ModuLand algorithm [21]. The clusters obtained are indicative of driver sub-networks for the stage-wise progression of cancer. Finally, we interrogated each of these networks for Gene Ontology (GO) enrichment using BiNGO [22]for three GO terms: biological process, molecular function and cellular component. To correct for the false discovery rate with p-values in multiple hypothesis testing, we applied the Benjamini-Hochberg filter in BiNGO and obtained the q-values[23].
Fig 2. Identification of hub driver genes.

Consensus novel driver genes were identified for each stage from driverDB data. Consensus central genes (‘hubs’) were identified from each stage-specific network. The overlap between these two sets of genes yields ‘hub driver’ genes for each stage.
Results and Discussion
Table 1 shows the number of driver genes at each step of our screening procedure. The final set of novel driver genes for each stage used in the subsequent Genemania search is shown in S1 Table. We analyzed the degree distribution of each of the stage wise networks and found that the node distribution of all the three networks conformed better to a power law distribution than a linear model (S2 Table). A power-law fit implies the presence of a few highly connected nodes (i.e. hubs)in the network. In general, hubs could predispose vulnerability to disease. Mutations in hub genes could lead to functional alterations in the corresponding protein which could lead to changes in its interactions with other proteins. This could lead to a cascading failure in the network and cause disease [24]. In this context, a power law behavior implies that mutations in hub genes could increase susceptibility to the hallmarks of cancer[25]and facilitate the spread of the perturbation in the network. Therefore, identification of the hub nodes could pinpoint the key genes whose failure would underscore the progression of cancer. Though our method of network construction was blind to the number of components, all our resultant network models were single connected giant components (S3 Table).
Table 1. From tumor genome-sequencing data to network reconstruction.
| Stage | No. of Samples | Consensus driver genes | Subtraction of previous stage | Screening Against background | Screening against Cancer Gene Census | Network reconstruction #node; #edge |
|---|---|---|---|---|---|---|
| II | 108 | 52 | 47 | 34 | 27 | 109; 396 |
| III | 75 | 56 | 43 | 35 | 31 | 109; 199 |
| IV | 28 | 49 | 43 | 32 | 30 | 115; 297 |
The number of driver genes at each step leading to the network reconstruction.
Analysis of progression to stage II
The reconstructed stage-II specific network (with node and edge attributes) is given in S1 File. Table 2 shows the central genes identified by centrality analysis for the stage II network. For progression to stage II, three hub driver genes were identified: DYNC1H1, GRIN2A, GRM1. Eight non-driver hub genes were identified: DLG4, SMC2, PLCG1, GRIN2B, RHOG, SMC1A, GRIN1, CAMK2A. Table 3 shows the modules in the network obtained by ModuLand analysis. Seven significant modules were obtained each centered at a different gene (Fig 3). Each module could function as a driver subnetwork for the progression of cancer to stage II. The most interesting module was that centered at DLG4, which also had the driver genes GRIN2A, GRM1, LRRC7 as its members. Of these, GRIN2A and GRM1 are also central genes. A gene ontology analysis for biological process yielded the key pathways in which these central driver genes played a role. The glutamate signaling pathway was identified as a key enriched biological process (q-value < 0.001). GRIN1, GRIN2A and GRIN2B emerged as key hub genes that could modulate this pathway. Among molecular functions, the glutamate receptor activity emerged significant (q-value < 0.001) in which GRM1 was implicated. The cellular component ontology was enriched in ionotropic glutamate receptor complex and the synapse (q-values < 0.001) (Table 4).
Table 2. Centrality analysis of stage-II network.
| S.No. | Betweenness centrality | Closeness centrality | Bottleneck centrality | Consensus Centrality (hubs) | Hub+driver genes |
|---|---|---|---|---|---|
| 1 | DLG4 | DLG4 | DLG4 | DLG4 | DYNC1H1 |
| 2 | SMC2 | GRIN2B | DYNC1H1 | SMC2 | GRIN2A |
| 3 | DYNC1H1 | GRIN2A | SMC2 | DYNC1H1 | GRM1 |
| 4 | PLCG1 | GRIN1 | PLCG1 | PLCG1 | |
| 5 | GRIN2B | CAMK2A | CAMK2A | GRIN2B | |
| 6 | GRIN2A | PLCG1 | RHOG | GRIN2A | |
| 7 | RHOG | DYNC1H1 | NF2 | RHOG | |
| 8 | SMC1A | RHOG | SMC1A | SMC1A | |
| 9 | GRM1 | LRRC7 | GRIN2A | GRM1 | |
| 10 | NF2 | GRIA1 | GRM1 | GRIN1 | |
| 11 | GRIN1 | GRM1 | HRSP12 | CAMK2A | |
| 12 | NCAPH | DLG2 | GRIN2B | ||
| 13 | CAMK2A | DYNLL1 | LRRC7 | ||
| 14 | SOX9 | SMC2 | RAD21 | ||
| 15 | CHEK1 | SMC1A | GRIN1 |
The top 15 genes obtained by applying each centrality metric are given. The genes at the intersection of all the three metrics are designated as ‘hubs’. Genes that are both hub and driver are shown.
Table 3. Moduland decomposition of stage-II network.
| S. No | Module centre | Eff. size of module | Driver gene members | Hub gene members |
|---|---|---|---|---|
| 1 | DLG4 | 43 | GRIN2A, GRM1, LRRC7 | CAMK2A, DLG4, GRIN1. GRIN2A, GRIN2B, GRM1, PLCG1, RHOG |
| 2 | SMC2 | 17 | NCAPD2, STAG1 | SMC1A, SMC2 |
| 3 | DYNC1H1 | 6 | DYNC1H1 | DYNC1H1 |
| 4 | PPP1R12B | 9 | PPP1R12B, KCNQ5 | |
| 5 | UBE4B | 6 | UBE4B | |
| 6 | CLSPN | 4 | CLSPN | |
| 7 | SOX9 | 5 | SOX9 |
Each module is represented by its centre and effective size. Hub genes identified by centrality analysis and driver genes are indicated by their module membership. Modules with the maximum number of hub genes could function as driver subnetworks in the pathogenesis of disease progression.
Fig 3. Network of cluster centres of stage-II network.
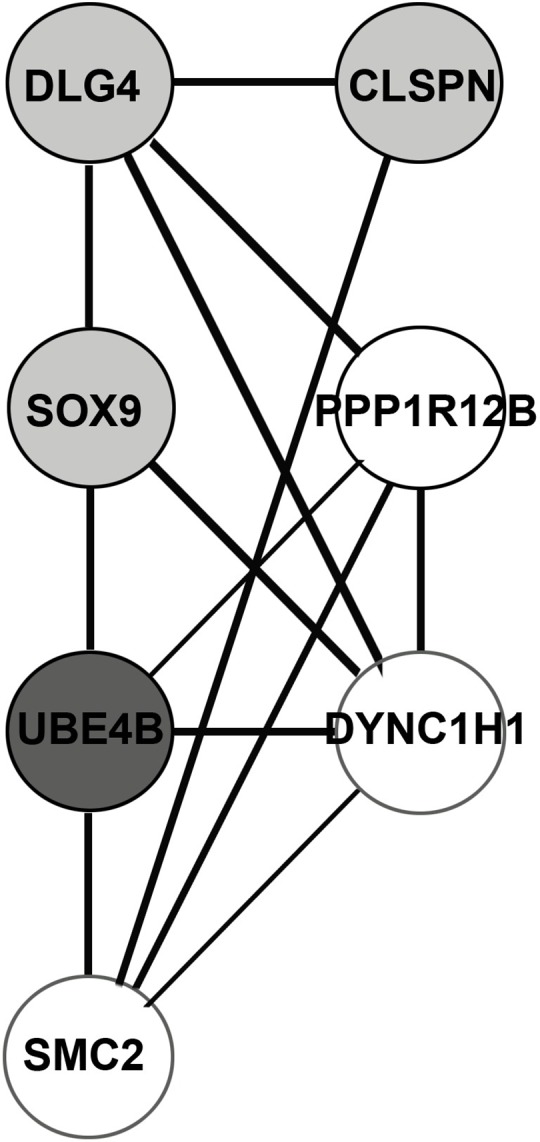
Clustering was done using Moduland.
Table 4. GO enrichment analysis of Stage-II network.
| GO- ID | q-value | %size > 20 | Description | Hub genes | Other genes in the network |
|---|---|---|---|---|---|
| 7059 | 3.10E-12 | N | chromosome segregation | SMC2, SMC1A | PDS5B, SMC3,SMC4,NCAPD2,NCAPH,RAD21, NDEL1, NCAPG, STAG2,NEK6,STAG1 |
| 7215 | 1.62E-10 | Y | glutamate signaling pathway | GRIN1,GRIN2A,GRIN2B | HOMER3, GRIN3B, GRIA4,HOMER1, HOMER2 |
| 45202 | 1.62E-10 | N | Synapse | GRIN1,GRIN2A,GRIN2B, GRM1, DLG4, CAMK2A | GABBR1,NLGN2,GRIN3B,GRIA4,CDH2,HOMER1,ADORA1,HOMER2,SLC17A7,SLC32A1,GRIA1,HOMER3, DLG2 |
| 279 | 5.55E-10 | N | M phase | DYNC1H1, SMC1A, SMC2 | SSSCA1,FZR1,PDS5B,PDS5A,CHEK1,SMC3,SMC4,NCAPD2,NCAPH,RAD21,NCAPG,STAG2,NEK6,NUDC,STAG1 |
| 8328 | 7.06E-09 | Y | ionotropic glutamate receptor complex | GRIN1,GRIN2A,GRIN2B | GRIA1, GRIN3B, GRIA4 |
| 8066 | 2.31E-08 | Y | glutamate receptor activity | GRM1, GRIN1,GRIN2A,GRIN2B | GRIA1, GABBR1, GRIN3B, GRIA4 |
| 7216 | 1.60E-04 | Y | metabotropic glutamate receptor signaling pathway | HOMER3, HOMER1, HOMER2 | |
| 50839 | 1.18E-02 | N | cell adhesion molecule binding | GRIN2A,GRIN2B | PTPRT |
Significant GO terms enriched in stage II network are given. If the ratio of the genes in the network for a given GO term to the total number of genes in that GO term is greater than 20%, ‘Y’ is indicated, otherwise ‘N’ is indicated. Hub genes identified in our analysis are indicated for each GO term.
Analysis of progression to stage III
The reconstructed stage-III specific network (with node and edge attributes) is given in S2 File. Table 5 shows the central genes identified by centrality analysis for the stage III network. For progression to stage III, four hub driver genes were identified: IGF1R, CPS1, SPTA1, DSP. Six non-driver hub genes were identified: HEATR1, MAPK9, ARAF, PRKCE, PLEC, MSN. Table 6 shows the modules in the network identified by the ModuLand algorithm. All the four hub driver genes were classified as core members of the network clusters (Fig 4). The most significant enrichment in the GO analysis of the biological process was the regulation of actin cytoskeleton organization (q-value < 0.001; Table 7). The GO enrichment in the molecular function (“actin binding”) and cellular component (“actin cytoskeleton”, “spectrin”) provided further evidence for the involvement of SPTA1 and PLEC. DSP seemed to modulate the cell adhesion properties in advancing the cancer malignancy.
Table 5. Centrality analysis of stage-III network.
| S.No | Betweenness centrality | Closeness centrality | Bottleneck centrality | Consensus Centrality (hubs) | Hub+driver genes |
|---|---|---|---|---|---|
| 1 | HEATR1 | MACF1 | ARAF | HEATR1 | IGF1R |
| 2 | IGF1R | MAPK9 | MAPK9 | IGF1R | CPS1 |
| 3 | MAPK9 | IGF1R | PLEC | MAPK9 | SPTA1 |
| 4 | ARAF | SPTA1 | MSN | ARAF | DSP |
| 5 | XRN1 | PRKCE | HEATR1 | PRKCE | |
| 6 | PRKCE | XRN1 | IGF1R | CPS1 | |
| 7 | CPS1 | DSP | CPS1 | PLEC | |
| 8 | PLEC | ARAF | SPTA1 | MSN | |
| 9 | MSN | PLEC | UTP20 | SPTA1 | |
| 10 | MTOR | HEATR1 | RICTOR | DSP | |
| 11 | MACF1 | CPS1 | MTOR | ||
| 12 | UTP20 | MSN | PIK3R3 | ||
| 13 | SPTA1 | NRXN2 | DSP | ||
| 14 | DSP | COL17A1 | COL17A1 | ||
| 15 | MAPK6 | MAPK6 | PRKCE |
The top 15 genes obtained by applying each centrality metric are given. The genes at the intersection of all the three metrics are designated as ‘hubs’. Genes that are both hub and driver are shown.
Table 6. Moduland decomposition of stage-III network.
| S. No | Module centre | Eff. size of module | Driver gene members | Hub gene members |
|---|---|---|---|---|
| 1 | MTOR | 15 | RICTOR, UTP20, MAGEC1 | HEATR1 |
| 2 | SPTA1 | 11 | SPTA1, MACF1 | SPTA1, PLEC |
| 3 | DSP | 12 | DSP, ADAM28, PKD2L1 | DSP, PRKCE |
| 4 | SPTB | 7 | MACF1 | |
| 5 | NRXN2 | 8 | NRXN3 | |
| 6 | XRN1 | 16 | XRN1, CPS1 | CPS1 |
| 7 | TEK | 7 | TEK | |
| 8 | MAPK9 | 14 | MAPK6, ANK3 | MAPK9, ARAF |
| 9 | IGF1R | 7 | IGF1R | IGF1R, MSN |
| 10 | FBN2 | 6 | FBN2 | |
| 11 | MYLK | 7 | MYLK | |
| 12 | CHRM2 | 4 | CHRM2 | |
| 13 | AIFM1 | 5 | AIFM1 |
Each module is represented by its centre and effective size. Hub genes identified by centrality analysis and driver genes are indicated by their module membership. Modules with the maximum number of hub genes could function as driver subnetworks in the pathogenesis of disease progression.
Fig 4. Network of cluster centres of stage-III network.
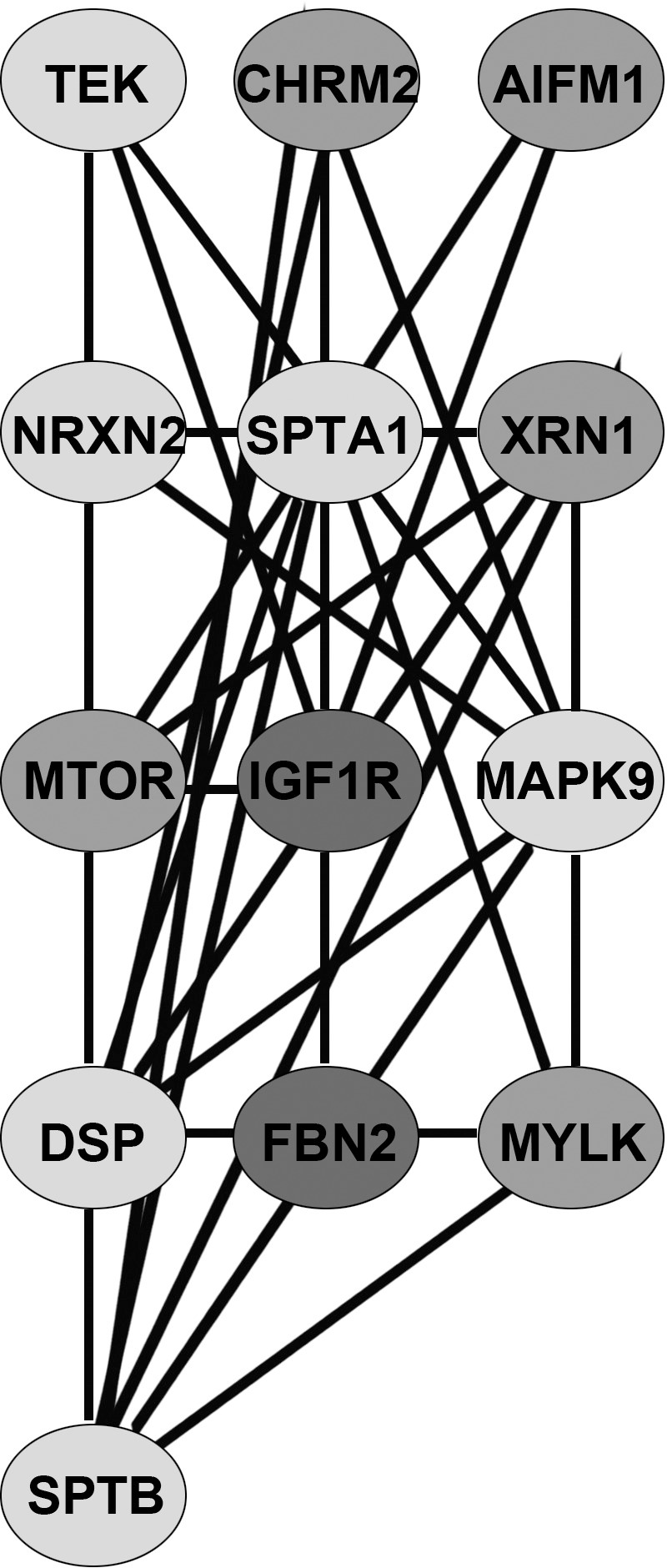
Clustering was done using Moduland.
Table 7. GO enrichment analysis of stage-III network.
| GO- ID | q-value | %size > 20 | Description | Hub genes | Other genes in the network |
|---|---|---|---|---|---|
| 7059 | 3.10E-12 | N | chromosome segregation | SMC2, SMC1A | PDS5B,SMC3,SMC4,NCAPD2,NCAPH,RAD21,NDEL1,NCAPG,STAG2,NEK6,STAG1 |
| 7215 | 1.62E-10 | Y | glutamate signaling pathway | GRIN1,GRIN2A,GRIN2B | HOMER3, GRIN3B,GRIA4,HOMER1, HOMER2 |
| 45202 | 1.62E-10 | N | Synapse | GRIN1,GRIN2A,GRIN2B, GRM1, DLG4,CAMK2A | GABBR1,NLGN2,GRIN3B,GRIA4,CDH2,HOMER1, ADORA1,HOMER2,SLC17A7,SLC32A1, GRIA1,HOMER3, DLG2 |
| 279 | 5.55E-10 | N | M phase | DYNC1H1, SMC1A, SMC2 | SSSCA1,FZR1,PDS5B,PDS5A,CHEK1,SMC3,SMC4,NCAPD2,NCAPH,RAD21,NCAPG,STAG2,NEK6,NUDC,STAG1 |
| 8328 | 7.06E-09 | Y | ionotropic glutamate receptor complex | GRIN1,GRIN2A,GRIN2B | GRIA1, GRIN3B, GRIA4 |
| 8066 | 2.31E-08 | Y | glutamate receptor activity | GRM1,GRIN1,GRIN2A,GRIN2B | GRIA1, GABBR1, GRIN3B, GRIA4 |
| 7216 | 1.60E-04 | Y | metabotropic glutamate receptor signaling pathway | HOMER3, HOMER1, HOMER2 | |
| 50839 | 1.18E-02 | N | cell adhesion molecule binding | GRIN2A,GRIN2B | PTPRT |
Significant GO terms enriched in stage III network are given. If the ratio of the genes in the network for a given GO term to the total number of genes in that GO term is greater than 20%, ‘Y’ is indicated, otherwise ‘N’ is indicated. Hub genes identified in our analysis are indicated for each GO term.
Analysis of progression to stage IV
The reconstructed stage-IV specific network (with node and edge attributes) is given in S3 File. Table 8 shows the central genes identified by centrality analysis for the stage IV network. For progression to stage IV, three hub driver genes were identified: GSK3B, GGT1, EIF2B5. Seven non-driver hub genes were identified: AKT1, PXN, SFN, GNAI2, CHKB, HSPA5, PLCG1. A ModuLand analysis of the network yielded a module centered at EIF2B5, which included all the three hub driver genes noted above (Table 9; Fig 5). A GO biological process enrichment analysis yielded ‘negative regulation of translational initiation in response to stress’ (q-value < 0.001; Table 10). This hit contained the EIF2B5 gene, which was also a member of the enriched cellular component ‘eukaryotic translation initiation factor 2B complex’ (q-value < 0.001). The GO molecular function enrichment analysis yielded the following: adenyl nucleotide binding containing hub genes GSK3B and HSPA5 (q-value = 0.001) and gamma-glutamyl transferase activity containing hub gene GGT1 (q-value ≈ 0.01). A connection with neurogenesis and oligodendrocyte development was seen (q-values < 0.001), which might seem surprising, but recent studies indicated a crucial link of these signaling pathways with the metastasis of colon cancer [26].
Table 8. Centrality analysis of stage-IV network.
| S.No. | Betweenness centrality | Closeness centrality | Bottleneck centrality | Consensus Centrality (hubs) | Hub+driver genes |
|---|---|---|---|---|---|
| 1 | GSK3B | GSK3B | GSK3B | GSK3B | GSK3B |
| 2 | AKT1 | PXN | AKT1 | AKT1 | GGT1 |
| 3 | PXN | AKT1 | HSPA5 | PXN | EIF2B5 |
| 4 | GGT1 | VEGFA | CHKB | GGT1 | |
| 5 | SFN | PLCG1 | GGT1 | SFN | |
| 6 | PPIF | EIF2B5 | PXN | GNAI2 | |
| 7 | GNAI2 | CHKB | SFN | CHKB | |
| 8 | CHKB | GNAI2 | PLCG1 | HSPA5 | |
| 9 | HSPA5 | FLT1 | EIF2B5 | EIF2B5 | |
| 10 | EIF2B5 | HSPA5 | QARS | PLCG1 | |
| 11 | PLCG1 | SFN | SSR4 | ||
| 12 | SF3B3 | GGT1 | GNAI2 | ||
| 13 | IARS | SF3B1 | HECW1 | ||
| 14 | NTRK2 | ITGAV | NTRK2 | ||
| 15 | MAGI1 | ITGA9 | SF3B3 |
The top 15 genes obtained by applying each centrality metric are given. The genes at the intersection of all the three metrics are designated as ‘hubs’. Genes that are both hub and driver are shown.
Table 9. Moduland decomposition of stage-IV network.
| S. No | Module centre | Eff. size of module | Driver gene members | Hub gene members |
|---|---|---|---|---|
| 1 | EIF2B5 | 14 | EIF2B5 | EIF2B5, CHKB, GGT1, GSK3B |
| 2 | FLT1 | 29 | FLT1, TNC, ADCY8 | GNAI2, PXN, PLCG |
| 3 | GSK3B | 34 | GSK3B, MAGI1, LCT | GSK3B, AKT1, HSPA5, SFN |
| 4 | NTRK2 | 7 | NTRK2, SLAMF7 | |
| 5 | EPRS | 8 | EPRS | |
| 6 | SF3B3 | 12 | SF3B3, GGT1, SYT14 | GGT1 |
| 7 | HLA-C | 5 | ||
| 8 | ROR2 | 4 | ||
| 9 | FGG | 3 | F13B | |
| 10 | GABRG2 | 4 | GABRG2 |
Each module is represented by its centre and effective size. Hub genes identified by centrality analysis and driver genes are indicated by their module membership. Modules with the maximum number of hub genes could function as driver subnetworks in the pathogenesis of disease progression.
Fig 5. Network of cluster centres of stage-IV network.

Clustering was done using Moduland.
Table 10. GO enrichment analysis of stage-IV network.
| GO- ID | q-value | %size > 20 | Description | Hub genes | Other genes in the network |
|---|---|---|---|---|---|
| 5851 | 7.11E-09 | Y | eukaryotic translation initiation factor 2B complex | EIF2B5 | EIF2B2, EIF2B1,EIF2B, EIF2B4 |
| 22008 | 1.23E-05 | N | Neurogenesis | EIF2B5, AKT1 | WNT5A,NTF3,PTPRZ1,NF1,DLL1,FZD2,EIF2B1,BDNF,DYNLL2,TNR,VEGFA,ROR2,EIF2B2,EIF2B3,EIF2B4,DISC1 |
| 30817 | 3.76E-05 | N | regulation of cAMP biosynthetic process | GNIA2 | ADCY1,ADCY2,ADCY8,DRD5,ADCY5,NF1,NTRK2 |
| 30554 | 1.04E-03 | N | adenyl nucleotide binding | GSK3B, HSPA5 | ADCY1,FLT1,ADCY2,SGK3,MAGI1,TAOK1,ADCY8,ADCY5,CHKB,EPRS,QARS,CLPX,MARK1,IARS,AKT1,MUSK,CBWD1,RARS,NTRK2,ROR2,EIF2B2,DUS2L |
| 43409 | 1.10E-03 | N | negative regulation of MAPKKK cascade | AKT1 | SPRY2, MAGI1,NF1 |
| 3840 | 1.27E-02 | Y | gamma-glutamyl transferase activity | GGT1 | GGT2 |
Significant GO terms enriched in stage IV network are given. If the ratio of the genes in the network for a given GO term to the total number of genes in that GO term is greater than 20%, ‘Y’ is indicated, otherwise ‘N’ is indicated. Hub genes identified in our analysis are indicated for each GO hit.
We performed the literature survey for these novel driver genes, to provide an insight into the possible roles of these genes in cancer progression. To our surprise, most of the genes were under studied in connection with colon cancer.
Stage II hub genes
Dynein, Cytoplasmic 1, Heavy Chain 1(DYNC1H1)
DYNC1H1 has been shown to function in intracellular motility like protein sorting, movement of the organelles and dynamics of the spindles. A recent study has reported that DYNC1H1 is mutated in ovarian cancer, pancreatic neuroendocrine neoplasms, and glioblastoma multiforme (GBM) [27]. In addition, the expression of DYNC1H1 was significantly upregulated in three drug-resistant gastric cancer cell lines (5-fluorouracil (5FU), paclitaxel (TA) and cisplatin (DDP)-resistant gastric cancer cell lines) [28].
Glutamate Receptor, Ionotropic, N-methyl D-aspartate 2A (GRIN2A)
GRIN2A is a member of the glutamate-gated ion channel proteins. It is a subunit of N-methyl-D-aspartate receptor. The activation of these receptors will increase the influx of calcium resulting in the triggering of several signaling cascades. In 2011, Wei et al., has identified that GRIN2A was mutated in 33% of melanoma samples [29]. Furthermore, D’mello et al., has provided evidence that the mutations in this gene are correlative to the progression of melanoma[30].
Glutamate receptor, metabotropic 1 (GRM1)
GRM1 was shown to activate phospholipase C. GRM1 was associated with various diseases such as depression and cancer. GRM1 has been implicated in prostate cancer [31], following identification of novel mutations and single nucleotide polymorphisms. In addition, GRM1 was over expressed in melanoma and ectopic overexpression of this gene in melanocytes resulted in neoplastic transformation [32]. Finally, studies with breast epithelial cells demonstrated that GRM1 cooperates with other factors in the hyperplastic mammary epithelium resulting in progression of breast cancer [33].
Discs, Large Homolog 4 (DLG4)
DLG4(also called as postsynaptic density protein 95) has shown to be expressed in normal cervical keratinocytes. The expression of DLG4 is significantly decreased in cervical cancer cell lines. Also, the tumorigenicity of CaSki cells was repressed following overexpression of DLG4[34]. In addition, Hering et al., has shown that DLG4 cooperates with Frizzled proteins to regulate the WNT signaling pathway, which has been implicated in multiple types of cancer progression, including colon cancer [35]. Table 3 shows that DLG4 is the module center of a key module of the stage II network.
Structural maintenance of chromosomes 2 (SMC2)
SMC2 plays a role in chromosomal stability. It is a subunit of condensing protein complexes that are shown to be involved in the condensation of the chromosomes. Table 3 shows that SMC2 is located at the center of a module of the stage II network. Je et al., has reported that SMC2 is mutated in gastric and colon cancer tissues suggesting its involvement in cancer progression [36]. Interestingly, beta-Catenin a key molecule in colorectal cancer has been shown to directly bind and regulate the transcription of SMC2[37].
Phospholipase C gamma 1 (PLCG1)
PLCG1 has been shown to play a significant role in intracellular signaling pathways as well as an increased apoptotic resistance and invasiveness of the cells [38]. PLCG1 levels were significantly increased in breast cancer tissues compared to normal [39]. Raimondi et al., has demonstrated a link between PLCG1 and phosphoinositide-dependent kinase 1 (PDK1), and their significance in the process of cancer cell invasion[40]. In addition, Park et al., has reported that PLCG1 levels are significantly high in adenomas and carcinomas compared to the normal colonic mucosa suggesting its role in progression of colon cancer[41].
Glutamate receptor, ionotropic, N-methyl D-aspartate 2B (GRIN2B)
GRIN2B is a glutamate-gated ion channel with very high calcium permeability. Park et al., has shown that the GRIN2B promoter region is hypermethylated during breast cancer progression[42]. Similar epigenetic changes at the GRIN2B locus could be involved in driving the progression of colon cancer.
Ras Homolog Family Member G (RHOG)
RHOG is a member of Rho family of GTPases. RHOG is lesser characterized among Rho family members, and its role in cancer progression is unknown. It has been shown to regulate morphological changes in cells. Also, RHOG has been demonstrated to promote cell survival through activation of PI3Kinase and Akt[43]. RHOG has been shown to be upregulated in glioblastoma compared to a non-neoplastic brain. In addition, it has been shown to mediate glioblastoma cell invasion following cMet and EGFR stimulation [44].
Structural maintenance of chromosomes 1A (SMC1A)
SMC protein family contains 6 members from SMC1 to SMC6 with varying functions. SMC1A is mutated in various malignant carcinomas. Down regulation of SMC1A resulted in growth inhibition in lung adenocarcinoma cells [45]. Furthermore, knockdown of SMC1A significantly suppressed the proliferation of the glioblastoma cells [46]. Recently, Wang et al., has published that SMC1A is a predictive factor for poor prognosis of colon cancer[47].
Glutamate Receptor Ionotropic, NMDA 1 (GRIN1)
GRIN1 is the under studied member of the glutamate receptor family with regards to cancer. There is only one study showing the interaction of GRIN1 and GRIN2A and their role in anchorage independent growth of melanoma cells[48].
Calcium/Calmodulin-Dependent Protein Kinase II Alpha (CAMK2A)
CAMK2A is a serine/threonine protein kinase. Yuan et al., has demonstrated that CaMKII played a very important role in osteosarcoma proliferation, and this could be a therapeutic target for osteosarcoma[49].
Stage III hub genes
Insulin-like Growth Factor 1 Receptor (IGF1R)
This tyrosine kinase receptor plays a critical role in cellular transformation events following binding of insulin-like growth factor. It is overexpressed in various malignant tissues, and it enhances cancer cell survival by inhibiting apoptotic cell death. In addition, there was a significant correlation between the expression of IGF1R with colorectal tumor size and depth of tumor invasion [50]. Finally, Kucab et al., has clearly highlighted the role of IGF1R in breast cancer metastasis[51].
Carbamoyl phosphate synthetase 1 (CPS1)
CPS1 is a mitochondrial enzyme involved in the urea cycle. Recent studies have mentioned that the expression of CPS1 is a negative prognostic factor in rectal cancers that receive concurrent chemoradiotherapy[52]. In addition, Li et al., has demonstrated the utilization of the anti-CSP1 for the detection of circulating tumor cells in hepatocellular carcinoma (HCC)[53]. Finally, studies have also shown that CPS1 expression in human HCC cells is silenced by DNA methylation [54].CSP1 could be a potential biomarker for HCC.Milinkovic et al., has identified CPS1 as genetically altered in malignant glioma patient samples [55].
Spectrin, alpha, erythrocytic 1 (SPTA1)
SPTA1 is a scaffold protein that functions in determining cell shape and organization of organelles. These families of proteins are primarily composed of spectrin repeats involved in dimer formation. Recent studies have shown that β2-spectrin is implicated in colorectal and pancreatic cancer, where it regulates the transcriptional activators SMAD to affect transforming growth factor beta (TGFβ) signaling pathway. Dysregulation of TGFβ signaling through loss of β2-spectrin inappropriately activates Wnt signaling and promotes tumorigenesis [56]-[57]. In addition, spectrin family of proteins has been shown to play a role in hepatocellular cancer through regulation of cyclin D1 [58]. Interestingly, they have been shown to contribute to drug resistance in ovarian cancer [59]. Furthermore, the increase in the expression and heterogeneity of the cytoplasmic spectrin is associated with the invasiveness of malignant melanoma and squamous-cell carcinoma [60].
Desmoplakin (DSP)
DSP is an essential component of intercellular junctions called desmosomes. Loss of DSP expression has been shown to play an important role in breast cancer progression and metastasis [61]. Papagerakis et al., has demonstrated the utility of DSP as a marker for evaluating the risk of oropharyngeal cancer metastasis[62].
HEAT Repeat Containing 1 (HEATR1)
HEATR1 is known to be to be involved in the biogenesis of ribosomes. Liu et al., has recently demonstrated that HEATR1 plays a significant role in pancreatic cancer cell drug resistance, and that HEATR1 regulates Akt pathway[63]. In addition, Wu et al., has demonstrated that the overexpression of HEATR1 in glioblastoma cells resulted in the induction of a cytotoxic T lymphocyte response to its epitopes, which allowed for the selective targeting of the glioblastoma cells and glioma stem like cells[64].
Mitogen-activated protein kinase 9 (MAPK9)
MAPK9 also called as c-Jun N-terminal kinases (JNK2), belongs to the family of MAPK kinases, and has shown to regulate multiple cellular processes including proliferation, differentiation, and transcription regulation. Ahmed et al., has demonstrated that JNK2 mediated suppression of JNK1 apoptotic pathway is required for the survival of cancer cells[65]. In addition, blocking the expression of JNK2 has significantly inhibited the migration ability of breast cancer cells [66].
A-Raf serine/threonine kinase (ARAF)
ARAF proto-oncogene is a member of RAF subfamily, and has been implicated in cell growth and development. Mutations in this gene have been shown to transform immortalized human airway epithelial cells [67]. In addition, Mooz et al., has shown that ARAF has an essential part in stimulating MAPK activity and cell migration[68].
Protein Kinase C, Epsilon (PRKCE)
PRKCE is another serine- and threonine-specific protein kinase activated by calcium and diacylglycerol. PRKCE is correlated with cell transformation and tumorigenesis. It has been shown to suppress apoptotic death of cells. The oncogenic potential of PRKEC in thyroid cancer was demonstrated by Zhang et al [69]. PRKCE has been shown to play an important role in promoting an aggressive metastatic breast cancer phenotype [70].
Plectin (PLEC)
PLEC is the cytolinker protein shown to regulate the tissue integrity, actin organization and cell migration. Yoneyama et al., has demonstrated the role of plectin in facilitating cancer cell invasion and metastasis[71]. In addition, plectin has been shown to regulate invasiveness by modulating actin assembly in SW480 colon cancer cells [72].
Moesin (MSN)
MSN is a membrane-organizing extensions spike protein. Moesin is identified to be present in the filopodia and other membranous projections that play a critical role in cell movement and cell signaling. A recent study has demonstrated that microRNA 200b inhibited breast cancer metastasis through regulation of moesin expression. In addition, increased expression of moesin has been shown to have an association with poor relapse-free survival [73]. Interestingly, phosphorylation of moesin by G protein-coupled receptor kinase has been shown to regulate prostate cancer metastasis [74].
Stage IV hub genes
Glycogen synthase kinase-3 (GSK3B)
GSK-3 is a serine-threonine kinase and shown to play a role in a wide range of cellular processes. Inhibition of GSK3B activity by Akt has been shown to influence the cancer progression. Furthermore, studies have shown that inhibition of GSK3 induced invasiveness of the breast cancer. Wnt signaling plays a critical role in colon cancer progression, and GSK3B is known to regulate this pathway[75]. Understanding the mutations in GSK3B will lead to improved therapies for colon cancer.
Gamma-Glutamyl transferase 1 (GGT1)
GGT1 is a membrane-bound enzyme catabolizing reduced glutathione to cysteine and glycine. GGT is a marker of oxidative stress in cells. Increased expression of GGT has shown to elevate the risk of progression of cervical cancer [76]. In addition, elevated levels of GGT are shown to be associated with increased invasion of melanoma cells in both in vitro and in vivo studies [77].
Eukaryotic Translation Initiation Factor 2B, Subunit 5 Epsilon (EIF2B5)
EIF2B5 is a regulator of protein synthesis. It has been associated with ovarian cancer, and angiogenesis[78]. In addition, genome-wide array study has identified EIF2B5 gene copy alteration in esophageal squamous-cell carcinoma patients [79]-[80].
Paxillin (PXN)
PXN is a cytoskeleton protein shown to be involved in cell adhesion. PXN mutations are associated with lung adenocarcinoma and are an independent predictor of survival and relapse of non-small cell lung cancer[81]. In addition, there are studies showing the role of PXN in the metastasis of osteosarcoma and prostate cancer [82,83]. A very recent study reported that PXN regulates tumor invasion, and is responsible for poor patient outcome in colorectal cancer patients [84].
Stratifin (SFN)
SFN is an adapter protein shown to regulate several signaling pathways. The promoter regions of SFN in most of the invasive lung adenocarcinoma samples are methylated to silence SFN expression[85]. In 2015, Shiba et al demonstrated the role of SFN on lung tumor development and progression[86].
Guanine Nucleotide Binding Protein, Alpha Inhibiting Activity Polypeptide 2 (GNAI2)
GNAI2 has shown to be involved as modulators or transducers in several transmembrane signaling pathways. GNAI2 has been implicated in ovarian cancer, where it acts as a driver of cancer progression[87]. Jiang et al., has reported the proto-oncogenic role of GNAI2 in tongue squamous-cell carcinoma initiation and progression[88].
Choline Kinase Beta (CHKB)
CHKB has been shown to be involved in the biosynthesis of phospholipids. There is very little information on the role of CHKB on cancer. However, TP53 and CHKB may regulate CDK4/6 collaboratively to suppress the progression of ovarian cancer[89]. Gallego-Ortega et al., has attempted to determine the involvement of CHKA and CHKB in cancer[90].
Heat Shock 70kDa Protein 5 (HSPA5)
HSPA5 is involved in the folding and assembly of proteins. It has been shown to regulate the anti-apoptotic unfolded protein response signaling network, which provides a ready mechanism for promoting cancer progression and metastasis [91]. Recent study by Chang et al., has demonstrated the role of HSPA5 on breast cancer cell migration and invasion[92]. Interestingly, Booth et al., 2014, has reported that OSU-03012, a cyclo-oxygenase inhibitor, targets HSPA5 and induces cancer cell death[93].
AKT1
AKT1 is a well-known driver of cancer [94]-[95], hence its appearance in our analysis might seem surprising. Despite the elimination of AKT1 at the screening stage, repopulating the network with interacting partners seemed to have led to the emergence of AKT1 in the network. In fact, it was the only entry in the Cancer Gene Census that re-appeared as a hub. This could be interpreted as additional evidence of the essential role of AKT1 in the metastasis of cancer.
In summary, for each stage of colon cancer, we have identified consensus driver genes, consensus hub genes, and the genes at their intersection referred as ‘hub driver’ genes. Each of these genes is a potential novel diagnostic biomarker of the stage of colorectal cancer. We have discussed the probable association of each biomarker with the progression of cancer, and in each case, we have found ample evidence that the gene in question constituted a missing link in the current understanding of the progression of colorectal cancer. It would appear that some of these biomarkers might even be involved in the progression of multiple unrelated cancers. Each identified biomarker represents a potential target for chemotherapeutic intervention in colorectal cancer. The modularity analysis reaffirmed these key genes, identifying the larger subnetworks of which they are a part. More than individual genes, it could be the subnetworks that are critical to the cancer progression and hence these subnetworks could serve as diagnostic biomarkers for the stage of colorectal cancer as well as provide targets for therapy. The Gene Ontology analysis has shed light on certain novel mechanisms that might underlie the progression of colorectal cancer to its next malignant stage. Our study design has been effective in revealing novel biomarkers and mechanisms that are driving each stage of cancer.
Conclusion
Our study has yielded many novel stage-specific biomarkers which would boost current strategies towards critical early-stage diagnosis of the stage of colorectal cancer as well as target selection for rational and personalized cancer treatment. We have identified DYNC1H1,GRIN2A, and GRM1 as novel hub driver genes for the stage-II progression of colon adenocarcinoma. IGF1R, CPS1, SPTA1 and DSP were identified as novel hub driver genes for the stage-III progression, and GSK3B, GGT1 and EIF2B5 were identified as novel hub driver genes for stage-IV progression. Prognosis is clearly inversely correlated with the stage of cancer and hence the biomarkers discussed above could predict the prognosis based on the stage-specificity. Our results provide multiple concrete directions for the deeper investigation of the biology of colorectal cancer malignancy in the future. Our methodology is extendable to the analysis of multiple types of cancer progression to yield novel useful biomarkers.
Supporting Information
(XGMML)
(XGMML)
(XGMML)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
We would like to thank the faculty members of Allied Health Sciences and the computing staff for their support.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The authors received no specific funding for this work.
References
- 1.Jemal A, Center MM, DeSantis C, Ward EM (2010) Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol Biomarkers Prev 19: 1893–1907. 10.1158/1055-9965.EPI-10-0437 [DOI] [PubMed] [Google Scholar]
- 2.Nagahashi M, Ramachandran S, Rashid OM, Takabe K (2010) Lymphangiogenesis: a new player in cancer progression. World J Gastroenterol 16: 4003–4012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ramalingam S, Natarajan G, Schafer C, Subramaniam D, May R, et al. (2008) Novel intestinal splice variants of RNA-binding protein CUGBP2: isoform-specific effects on mitotic catastrophe. Am J Physiol Gastrointest Liver Physiol 294: G971–981. 10.1152/ajpgi.00540.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ramalingam S, Ramamoorthy P, Subramaniam D, Anant S (2012) Reduced Expression of RNA Binding Protein CELF2, a Putative Tumor Suppressor Gene in Colon Cancer. Immunogastroenterology 1: 27–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sureban SM, Ramalingam S, Natarajan G, May R, Subramaniam D, et al. (2008) Translation regulatory factor RBM3 is a proto-oncogene that prevents mitotic catastrophe. Oncogene 27: 4544–4556. 10.1038/onc.2008.97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.TCGA (2012) Comprehensive molecular characterization of human colon and rectal cancer. Nature 487: 330–337. 10.1038/nature11252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cheng WC, Chung IF, Chen CY, Sun HJ, Fen JJ, et al. (2014) DriverDB: an exome sequencing database for cancer driver gene identification. Nucleic Acids Res 42: D1048–1054. 10.1093/nar/gkt1025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Reimand J, Bader GD (2013) Systematic analysis of somatic mutations in phosphorylation signaling predicts novel cancer drivers. Mol Syst Biol 9: 637 10.1038/msb.2012.68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vandin F, Upfal E, Raphael BJ (2012) De novo discovery of mutated driver pathways in cancer. Genome Res 22: 375–385. 10.1101/gr.120477.111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhao J, Zhang S, Wu LY, Zhang XS (2012) Efficient methods for identifying mutated driver pathways in cancer. Bioinformatics 28: 2940–2947. 10.1093/bioinformatics/bts564 [DOI] [PubMed] [Google Scholar]
- 11.Youn A, Simon R (2011) Identifying cancer driver genes in tumor genome sequencing studies. Bioinformatics 27: 175–181. 10.1093/bioinformatics/btq630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cerami E, Demir E, Schultz N, Taylor BS, Sander C (2010) Automated network analysis identifies core pathways in glioblastoma. PLoS One 5: e8918 10.1371/journal.pone.0008918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gonzalez-Perez A, Lopez-Bigas N (2012) Functional impact bias reveals cancer drivers. Nucleic Acids Res 40: e169 10.1093/nar/gks743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lawrence MS, Stojanov P, Polak P, Kryukov GV, Cibulskis K, et al. (2013) Mutational heterogeneity in cancer and the search for new cancer-associated genes. Nature 499: 214–218. 10.1038/nature12213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ciriello G, Cerami E, Sander C, Schultz N (2012) Mutual exclusivity analysis identifies oncogenic network modules. Genome Res 22: 398–406. 10.1101/gr.125567.111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Futreal PA, Coin L, Marshall M, Down T, Hubbard T, et al. (2004) A census of human cancer genes. Nat Rev Cancer 4: 177–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, et al. (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13: 2498–2504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Montojo J, Zuberi K, Rodriguez H, Kazi F, Wright G, et al. (2010) GeneMANIA Cytoscape plugin: fast gene function predictions on the desktop. Bioinformatics 26: 2927–2928. 10.1093/bioinformatics/btq562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Assenov Y, Ramirez F, Schelhorn SE, Lengauer T, Albrecht M (2008) Computing topological parameters of biological networks. Bioinformatics 24: 282–284. [DOI] [PubMed] [Google Scholar]
- 20.Scardoni G, Petterlini M, Laudanna C (2009) Analyzing biological network parameters with CentiScaPe. Bioinformatics 25: 2857–2859. 10.1093/bioinformatics/btp517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Szalay-Beko M, Palotai R, Szappanos B, Kovacs IA, Papp B, et al. (2012) ModuLand plug-in for Cytoscape: determination of hierarchical layers of overlapping network modules and community centrality. Bioinformatics 28: 2202–2204. 10.1093/bioinformatics/bts352 [DOI] [PubMed] [Google Scholar]
- 22.Maere S, Heymans K, Kuiper M (2005) BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21: 3448–3449. [DOI] [PubMed] [Google Scholar]
- 23.Hochberg Y, Benjamini Y (1990) More powerful procedures for multiple significance testing. Stat Med 9: 811–818. [DOI] [PubMed] [Google Scholar]
- 24.Jeong H, Mason SP, Barabasi AL, Oltvai ZN (2001) Lethality and centrality in protein networks. Nature 411: 41–42. [DOI] [PubMed] [Google Scholar]
- 25.Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144: 646–674. 10.1016/j.cell.2011.02.013 [DOI] [PubMed] [Google Scholar]
- 26.Fancy SP, Harrington EP, Baranzini SE, Silbereis JC, Shiow LR, et al. (2014) Parallel states of pathological Wnt signaling in neonatal brain injury and colon cancer. Nat Neurosci 17: 506–512. 10.1038/nn.3676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Huang HL, Yao HS, Wang Y, Wang WJ, Hu ZQ, et al. (2014) Proteomic identification of tumor biomarkers associated with primary gallbladder cancer. World J Gastroenterol 20: 5511–5518. 10.3748/wjg.v20.i18.5511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Huang H, Han Y, Wu J, Tian ZH, Qu LK, et al. (2014) [Establishment of drug resistant cell line of MGC-803 and analysis of differential secretome]. Beijing Da Xue Xue Bao 46: 183–189. [PubMed] [Google Scholar]
- 29.Wei X, Walia V, Lin JC, Teer JK, Prickett TD, et al. (2011) Exome sequencing identifies GRIN2A as frequently mutated in melanoma. Nat Genet 43: 442–446. 10.1038/ng.810 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.D'Mello S A, Flanagan JU, Green TN, Leung EY, Askarian-Amiri ME, et al. (2014) Evidence That GRIN2A Mutations in Melanoma Correlate with Decreased Survival. Front Oncol 3: 333 10.3389/fonc.2013.00333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ali S, Shourideh M, Koochekpour S (2014) Identification of novel GRM1 mutations and single nucleotide polymorphisms in prostate cancer cell lines and tissues. PLoS One 9: e103204 10.1371/journal.pone.0103204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wen Y, Li J, Koo J, Shin SS, Lin Y, et al. (2014) Activation of the glutamate receptor GRM1 enhances angiogenic signaling to drive melanoma progression. Cancer Res 74: 2499–2509. 10.1158/0008-5472.CAN-13-1531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Banda M, Speyer CL, Semma SN, Osuala KO, Kounalakis N, et al. (2014) Metabotropic glutamate receptor-1 contributes to progression in triple negative breast cancer. PLoS One 9: e81126 10.1371/journal.pone.0081126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Handa K, Yugawa T, Narisawa-Saito M, Ohno S, Fujita M, et al. (2007) E6AP-dependent degradation of DLG4/PSD95 by high-risk human papillomavirus type 18 E6 protein. J Virol 81: 1379–1389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hering H, Sheng M (2002) Direct interaction of Frizzled-1, -2, -4, and -7 with PDZ domains of PSD-95. FEBS Lett 521: 185–189. [DOI] [PubMed] [Google Scholar]
- 36.Je EM, Yoo NJ, Lee SH (2014) Mutational and expressional analysis of SMC2 gene in gastric and colorectal cancers with microsatellite instability. APMIS 122: 499–504. 10.1111/apm.12193 [DOI] [PubMed] [Google Scholar]
- 37.Davalos V, Suarez-Lopez L, Castano J, Messent A, Abasolo I, et al. (2012) Human SMC2 protein, a core subunit of human condensin complex, is a novel transcriptional target of the WNT signaling pathway and a new therapeutic target. J Biol Chem 287: 43472–43481. 10.1074/jbc.M112.428466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kunze K, Spieker T, Gamerdinger U, Nau K, Berger J, et al. (2014) A recurrent activating PLCG1 mutation in cardiac angiosarcomas increases apoptosis resistance and invasiveness of endothelial cells. Cancer Res 74: 6173–6183. 10.1158/0008-5472.CAN-14-1162 [DOI] [PubMed] [Google Scholar]
- 39.Arteaga CL, Johnson MD, Todderud G, Coffey RJ, Carpenter G, et al. (1991) Elevated content of the tyrosine kinase substrate phospholipase C-gamma 1 in primary human breast carcinomas. Proc Natl Acad Sci U S A 88: 10435–10439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Raimondi C, Chikh A, Wheeler AP, Maffucci T, Falasca M (2012) A novel regulatory mechanism links PLCgamma1 to PDK1. J Cell Sci 125: 3153–3163. 10.1242/jcs.100511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Park JG, Lee YH, Kim SS, Park KJ, Noh DY, et al. (1994) Overexpression of phospholipase C-gamma 1 in familial adenomatous polyposis. Cancer Res 54: 2240–2244. [PubMed] [Google Scholar]
- 42.Park SY, Kwon HJ, Lee HE, Ryu HS, Kim SW, et al. (2011) Promoter CpG island hypermethylation during breast cancer progression. Virchows Arch 458: 73–84. 10.1007/s00428-010-1013-6 [DOI] [PubMed] [Google Scholar]
- 43.Murga C, Zohar M, Teramoto H, Gutkind JS (2002) Rac1 and RhoG promote cell survival by the activation of PI3K and Akt, independently of their ability to stimulate JNK and NF-kappaB. Oncogene 21: 207–216. [DOI] [PubMed] [Google Scholar]
- 44.Kwiatkowska A, Didier S, Fortin S, Chuang Y, White T, et al. (2012) The small GTPase RhoG mediates glioblastoma cell invasion. Mol Cancer 11: 65 10.1186/1476-4598-11-65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang YF, Jiang R, Li JD, Zhang XY, Zhao P, et al. (2013) SMC1A knockdown induces growth suppression of human lung adenocarcinoma cells through G1/S cell cycle phase arrest and apoptosis pathways in vitro. Oncol Lett 5: 749–755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yang Y, Zhang Z, Wang R, Ma W, Wei J, et al. (2013) siRNA-mediated knockdown of SMC1A expression suppresses the proliferation of glioblastoma cells. Mol Cell Biochem 381: 209–215. 10.1007/s11010-013-1704-9 [DOI] [PubMed] [Google Scholar]
- 47.Wang J, Yu S, Cui L, Wang W, Li J, et al. (2015) Role of SMC1A overexpression as a predictor of poor prognosis in late stage colorectal cancer. BMC Cancer 15: 90 10.1186/s12885-015-1085-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Prickett TD, Zerlanko BJ, Hill VK, Gartner JJ, Qutob N, et al. (2014) Somatic mutation of GRIN2A in malignant melanoma results in loss of tumor suppressor activity via aberrant NMDAR complex formation. J Invest Dermatol 134: 2390–2398. 10.1038/jid.2014.190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yuan K, Chung LW, Siegal GP, Zayzafoon M (2007) alpha-CaMKII controls the growth of human osteosarcoma by regulating cell cycle progression. Lab Invest 87: 938–950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shiratsuchi I, Akagi Y, Kawahara A, Kinugasa T, Romeo K, et al. (2011) Expression of IGF-1 and IGF-1R and their relation to clinicopathological factors in colorectal cancer. Anticancer Res 31: 2541–2545. [PubMed] [Google Scholar]
- 51.Kucab JE, Dunn SE (2003) Role of IGF-1R in mediating breast cancer invasion and metastasis. Breast Dis 17: 41–47. [DOI] [PubMed] [Google Scholar]
- 52.Lee YY, Li CF, Lin CY, Lee SW, Sheu MJ, et al. (2014) Overexpression of CPS1 is an independent negative prognosticator in rectal cancers receiving concurrent chemoradiotherapy. Tumour Biol 35: 11097–11105. 10.1007/s13277-014-2425-8 [DOI] [PubMed] [Google Scholar]
- 53.Li J, Chen L, Zhang X, Zhang Y, Liu H, et al. (2014) Detection of circulating tumor cells in hepatocellular carcinoma using antibodies against asialoglycoprotein receptor, carbamoyl phosphate synthetase 1 and pan-cytokeratin. PLoS One 9: e96185 10.1371/journal.pone.0096185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Liu H, Dong H, Robertson K, Liu C (2011) DNA methylation suppresses expression of the urea cycle enzyme carbamoyl phosphate synthetase 1 (CPS1) in human hepatocellular carcinoma. Am J Pathol 178: 652–661. 10.1016/j.ajpath.2010.10.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Milinkovic V, Bankovic J, Rakic M, Stankovic T, Skender-Gazibara M, et al. (2013) Identification of novel genetic alterations in samples of malignant glioma patients. PLoS One 8: e82108 10.1371/journal.pone.0082108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kitisin K, Ganesan N, Tang Y, Jogunoori W, Volpe EA, et al. (2007) Disruption of transforming growth factor-beta signaling through beta-spectrin ELF leads to hepatocellular cancer through cyclin D1 activation. Oncogene 26: 7103–7110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jiang X, Gillen S, Esposito I, Giese NA, Michalski CW, et al. (2010) Reduced expression of the membrane skeleton protein beta1-spectrin (SPTBN1) is associated with worsened prognosis in pancreatic cancer. Histol Histopathol 25: 1497–1506. [DOI] [PubMed] [Google Scholar]
- 58.Baek HJ, Pishvaian MJ, Tang Y, Kim TH, Yang S, et al. (2011) Transforming growth factor-beta adaptor, beta2-spectrin, modulates cyclin dependent kinase 4 to reduce development of hepatocellular cancer. Hepatology 53: 1676–1684. 10.1002/hep.24128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Maeda O, Shibata K, Hosono S, Fujiwara S, Kajiyama H, et al. (2012) Spectrin alphaII and betaII tetramers contribute to platinum anticancer drug resistance in ovarian serous adenocarcinoma. Int J Cancer 130: 113–121. 10.1002/ijc.25983 [DOI] [PubMed] [Google Scholar]
- 60.Tuominen H, Sormunen R, Kallioinen M (1996) Non-erythroid spectrin (fodrin) in cutaneous tumours: diminished in cell membranes, increased in the cytoplasm. Br J Dermatol 135: 576–580. [PubMed] [Google Scholar]
- 61.Davies EL, Gee JM, Cochrane RA, Jiang WG, Sharma AK, et al. (1999) The immunohistochemical expression of desmoplakin and its role in vivo in the progression and metastasis of breast cancer. Eur J Cancer 35: 902–907. [DOI] [PubMed] [Google Scholar]
- 62.Papagerakis S, Shabana AH, Pollock BH, Papagerakis P, Depondt J, et al. (2009) Altered desmoplakin expression at transcriptional and protein levels provides prognostic information in human oropharyngeal cancer. Hum Pathol 40: 1320–1329. 10.1016/j.humpath.2009.02.002 [DOI] [PubMed] [Google Scholar]
- 63.Liu T, Fang Y, Zhang H, Deng M, Gao B, et al. (2015) HEATR1 negatively regulates Akt to help sensitize pancreatic cancer cells to chemotherapy. Cancer Res. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wu ZB, Qiu C, Zhang AL, Cai L, Lin SJ, et al. (2014) Glioma-associated antigen HEATR1 induces functional cytotoxic T lymphocytes in patients with glioma. J Immunol Res 2014: 131494 10.1155/2014/131494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ahmed SU, Milner J (2009) Basal cancer cell survival involves JNK2 suppression of a novel JNK1/c-Jun/Bcl-3 apoptotic network. PLoS One 4: e7305 10.1371/journal.pone.0007305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kaoud TS, Mitra S, Lee S, Taliaferro J, Cantrell M, et al. (2011) Development of JNK2-selective peptide inhibitors that inhibit breast cancer cell migration. ACS Chem Biol 6: 658–666. 10.1021/cb200017n [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Imielinski M, Greulich H, Kaplan B, Araujo L, Amann J, et al. (2014) Oncogenic and sorafenib-sensitive ARAF mutations in lung adenocarcinoma. J Clin Invest 124: 1582–1586. 10.1172/JCI72763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Mooz J, Oberoi-Khanuja TK, Harms GS, Wang W, Jaiswal BS, et al. (2014) Dimerization of the kinase ARAF promotes MAPK pathway activation and cell migration. Sci Signal 7: ra73 10.1126/scisignal.2005484 [DOI] [PubMed] [Google Scholar]
- 69.Zhang X, Li D, Li M, Ye M, Ding L, et al. (2014) MicroRNA-146a targets PRKCE to modulate papillary thyroid tumor development. Int J Cancer 134: 257–267. 10.1002/ijc.28141 [DOI] [PubMed] [Google Scholar]
- 70.Pan Q, Bao LW, Kleer CG, Sabel MS, Griffith KA, et al. (2005) Protein kinase C epsilon is a predictive biomarker of aggressive breast cancer and a validated target for RNA interference anticancer therapy. Cancer Res 65: 8366–8371. [DOI] [PubMed] [Google Scholar]
- 71.Sutoh Yoneyama M, Hatakeyama S, Habuchi T, Inoue T, Nakamura T, et al. (2014) Vimentin intermediate filament and plectin provide a scaffold for invadopodia, facilitating cancer cell invasion and extravasation for metastasis. Eur J Cell Biol 93: 157–169. 10.1016/j.ejcb.2014.03.002 [DOI] [PubMed] [Google Scholar]
- 72.McInroy L, Maatta A (2011) Plectin regulates invasiveness of SW480 colon carcinoma cells and is targeted to podosome-like adhesions in an isoform-specific manner. Exp Cell Res 317: 2468–2478. 10.1016/j.yexcr.2011.07.013 [DOI] [PubMed] [Google Scholar]
- 73.Li X, Roslan S, Johnstone CN, Wright JA, Bracken CP, et al. (2014) MiR-200 can repress breast cancer metastasis through ZEB1-independent but moesin-dependent pathways. Oncogene 33: 4077–4088. 10.1038/onc.2013.370 [DOI] [PubMed] [Google Scholar]
- 74.Chakraborty PK, Zhang Y, Coomes AS, Kim WJ, Stupay R, et al. (2014) G protein-coupled receptor kinase GRK5 phosphorylates moesin and regulates metastasis in prostate cancer. Cancer Res 74: 3489–3500. 10.1158/0008-5472.CAN-13-2708 [DOI] [PubMed] [Google Scholar]
- 75.McCubrey JA, Steelman LS, Bertrand FE, Davis NM, Sokolosky M, et al. (2014) GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 5: 2881–2911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fentiman IS (2012) Gamma-glutamyl transferase: risk and prognosis of cancer. Br J Cancer 106: 1467–1468. 10.1038/bjc.2012.128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Corti A, Franzini M, Paolicchi A, Pompella A (2010) Gamma-glutamyltransferase of cancer cells at the crossroads of tumor progression, drug resistance and drug targeting. Anticancer Res 30: 1169–1181. [PubMed] [Google Scholar]
- 78.Goode EL, Maurer MJ, Sellers TA, Phelan CM, Kalli KR, et al. (2010) Inherited determinants of ovarian cancer survival. Clin Cancer Res 16: 995–1007. 10.1158/1078-0432.CCR-09-2553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Shi ZZ, Liang JW, Zhan T, Wang BS, Lin DC, et al. (2011) Genomic alterations with impact on survival in esophageal squamous cell carcinoma identified by array comparative genomic hybridization. Genes Chromosomes Cancer 50: 518–526. 10.1002/gcc.20875 [DOI] [PubMed] [Google Scholar]
- 80.Shi ZZ, Shang L, Jiang YY, Hao JJ, Zhang Y, et al. (2013) Consistent and differential genetic aberrations between esophageal dysplasia and squamous cell carcinoma detected by array comparative genomic hybridization. Clin Cancer Res 19: 5867–5878. 10.1158/1078-0432.CCR-12-3753 [DOI] [PubMed] [Google Scholar]
- 81.Wu DW, Cheng YW, Wang J, Chen CY, Lee H (2010) Paxillin predicts survival and relapse in non-small cell lung cancer by microRNA-218 targeting. Cancer Res 70: 10392–10401. 10.1158/0008-5472.CAN-10-2341 [DOI] [PubMed] [Google Scholar]
- 82.Azuma K, Tanaka M, Uekita T, Inoue S, Yokota J, et al. (2005) Tyrosine phosphorylation of paxillin affects the metastatic potential of human osteosarcoma. Oncogene 24: 4754–4764. [DOI] [PubMed] [Google Scholar]
- 83.Ketscher A, Jilg CA, Willmann D, Hummel B, Imhof A, et al. (2014) LSD1 controls metastasis of androgen-independent prostate cancer cells through PXN and LPAR6. Oncogenesis 3: e120 10.1038/oncsis.2014.34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Huang CC, Wu DW, Lin PL, Lee H (2015) Paxillin promotes colorectal tumor invasion and poor patient outcomes via ERK-mediated stabilization of Bcl-2 protein by phosphorylation at Serine 87. Oncotarget 6: 8698–8708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Shiba-Ishii A, Noguchi M (2012) Aberrant stratifin overexpression is regulated by tumor-associated CpG demethylation in lung adenocarcinoma. Am J Pathol 180: 1653–1662. 10.1016/j.ajpath.2011.12.014 [DOI] [PubMed] [Google Scholar]
- 86.Shiba-Ishii A, Kim Y, Shiozawa T, Iyama S, Satomi K, et al. (2015) Stratifin accelerates progression of lung adenocarcinoma at an early stage. Mol Cancer 14: 142 10.1186/s12943-015-0414-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Raymond JR Jr., Appleton KM, Pierce JY, Peterson YK (2014) Suppression of GNAI2 message in ovarian cancer. J Ovarian Res 7: 6 10.1186/1757-2215-7-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Jiang L, Dai Y, Liu X, Wang C, Wang A, et al. (2011) Identification and experimental validation of G protein alpha inhibiting activity polypeptide 2 (GNAI2) as a microRNA-138 target in tongue squamous cell carcinoma. Hum Genet 129: 189–197. 10.1007/s00439-010-0915-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Zhang J, Wu LY, Zhang XS, Zhang S (2014) Discovery of co-occurring driver pathways in cancer. BMC Bioinformatics 15: 271 10.1186/1471-2105-15-271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gallego-Ortega D, Gomez del Pulgar T, Valdes-Mora F, Cebrian A, Lacal JC (2011) Involvement of human choline kinase alpha and beta in carcinogenesis: a different role in lipid metabolism and biological functions. Adv Enzyme Regul 51: 183–194. 10.1016/j.advenzreg.2010.09.010 [DOI] [PubMed] [Google Scholar]
- 91.Wang Q, Shu R, He H, Wang L, Ma Y, et al. (2012) Co-silencing of Birc5 (survivin) and Hspa5 (Grp78) induces apoptosis in hepatoma cells more efficiently than single gene interference. Int J Oncol 41: 652–660. 10.3892/ijo.2012.1471 [DOI] [PubMed] [Google Scholar]
- 92.Chang YW, Tseng CF, Wang MY, Chang WC, Lee CC, et al. (2015) Deacetylation of HSPA5 by HDAC6 leads to GP78-mediated HSPA5 ubiquitination at K447 and suppresses metastasis of breast cancer. Oncogene. [DOI] [PubMed] [Google Scholar]
- 93.Booth L, Roberts JL, Cash DR, Tavallai S, Jean S, et al. (2015) GRP78/BiP/HSPA5/Dna K is a universal therapeutic target for human disease. J Cell Physiol 230: 1661–1676. 10.1002/jcp.24919 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Malinowsky K, Nitsche U, Janssen KP, Bader FG, Spath C, et al. (2014) Activation of the PI3K/AKT pathway correlates with prognosis in stage II colon cancer. Br J Cancer 110: 2081–2089. 10.1038/bjc.2014.100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Cerezo-Guisado MI, Zur R, Lorenzo MJ, Risco A, Martin-Serrano MA, et al. (2015) Implication of Akt, ERK1/2 and alternative p38MAPK signalling pathways in human colon cancer cell apoptosis induced by green tea EGCG. Food Chem Toxicol 84: 125–132. 10.1016/j.fct.2015.08.017 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XGMML)
(XGMML)
(XGMML)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.