

The main task of the mitotic spindle is to generate forces that position the chromosomes at the metaphase plate and subsequently pull them apart toward the opposite spindle poles. These forces in living cells are, unfortunately, not easily accessible by current experimental techniques.1 However, much about the forces can be inferred from the shape of the spindle because the shape is an outcome of forces. K-fibers, which are bundles of microtubules ending at the kinetochore, are typically curved, suggesting that they are under compression. This inference contradicts the fact that sister kinetochores and thus also sister k-fibers are under tension, leaving us with a paradox about the origin of the curved shape of the spindle.
Let us imagine that something connects sister k-fibers and makes them follow a smooth arc. What could this structure be? The type of material that is able to link k-fibers in such a way must be of similar elasticity as the k-fibers. According to the current paradigm, sister k-fibers are connected by chromatin only, which is too flexible, because its Young's modulus is 7 orders of magnitude smaller than that of a microtubule. On the contrary, a microtubule bundle would have adequate elasticity to link sister k-fibers and make them curve into an arc.
To illustrate the origin of the curved shape of the spindle, we have constructed 2 macroscopic models made of wooden rods and a rope (Fig. 1). We chose the rods to represent microtubule bundles because they are stiff, and the rope to symbolize the flexible chromatin. The current paradigm, where sister k-fibers are connected by chromatin, is illustrated by a model in which 2 rods are connected by a rope. However, such a structure cannot take on a curved shape. If, on the other hand, we add a third rod as a link between these 2 rods, the entire structure can curve under compression.
Figure 1.
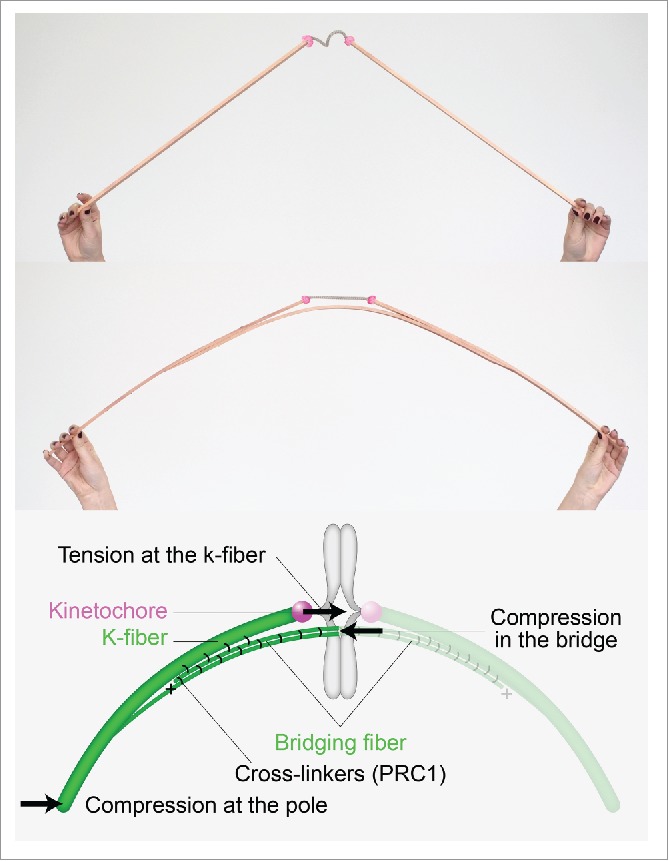
Bridging fiber links sister k-fibers and balances the tension on kinetochores. Macroscopic models made of wooden rods and a rope illustrate that the structure with 2 rods is not curved and the rope is relaxed, whereas the structure with 3 rods curves under compression and the rope gets tensed. The scheme shows the bridging fiber and forces in the spindle.
Indeed, we have observed a third microtubule bundle, which forms a bridge between sister k-fibers and thus we called it a bridging fiber2 (Fig. 1). A similar conclusion can be drawn from some electron micrographs of spindles such as in plant endosperm.3 Yet, even if microtubule bundles lie close to each other, they do not necessarily interact. By severing a k-fiber with a laser,4 we have shown that the bridging fiber moves together with sister k-fibers, revealing that these 3 fibers are strongly linked and thus able to survive large physical perturbations such as cutting of k-fibers. Severing of a k-fiber at different locations also revealed that the bridging fiber is linked laterally to the k-fibers in the region away from the kinetochore, while these fibers separate from each other close to the kinetochore.
The bridging fiber consists of 10-15 microtubules arranged in an anti-parallel manner, where the anti-parallel overlap, measured by PRC1-GFP, extends over 5 µm: 1 µm between sister kinetochores and 2 µm along each sister k-fiber.2 The observation that bridging fibers contain anti-parallel overlaps suggests that these fibers are a subset of overlap microtubules, also known as interpolar or interdigitating microtubules. These microtubules extend from the opposite spindle poles and interdigitate at the spindle equator. However, the fraction of overlap microtubules that bridge k-fibers, as well as the location of plus and minus ends of microtubules in the bridging fiber, remains to be determined.
The results of our experiments led us to a mathematically exact yet simple model.2 In this model, microtubule fibers in the spindle are elastic rods, which bend under compressive force.5 Their curvature tells us about the forces that bend the k-fibers. At the same time, this model explains how the tension on kinetochores is balanced: Compression in the bridging fiber balances the tension at the kinetochore and the compression at the spindle pole (Fig. 1). Thus, in our model the k-fiber is under tension close to the kinetochore and under compression close to the pole.6 The wooden model with 3 rods provides an intuitive understanding of this force balance.
Our model suggests that the force balance can be established if the bridging fiber splits from the k-fibers roughly 1 µm away from the kinetochore. This means that in experiments, if laser cutting is performed close to the kinetochore, this kinetochore will move toward its sister because the bridging fiber is now disconnected from the severed k-fiber and cannot balance the tension between sister kinetochores. On the contrary, if the cutting is performed far away from the kinetochore, the bridging fiber remains linked with the severed k-fiber and keeps balancing the inter-kinetochore tension and consequently their distance. Indeed, experiments have shown different kinetochore response dependent on the position of the cut.2
In this new picture of microtubule arrangement, linkers between k-fibers and the bridging fiber hold them together. To understand how this structure is maintained, it will be important to identify the key motor proteins and passive cross-linkers,7 as well as to determine their spatial distribution in the spindle. Combining such experiments with new models that include bridging fibers will help to understand how the forces in the spindle are generated and in what way they drive chromosome segregation. Finally, a major challenge will be to uncover how k-fibers, kinetochores and bridging fibers self-assemble into a metaphase spindle.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- [1].Nicklas RB. J Cell Biol 1983; 97:542-8; PMID:6885908; http://dx.doi.org/ 10.1083/jcb.97.2.542 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Kajtez J, et al.. Nat Commun 2016; 7:10298; PMID:26728792; http://dx.doi.org/ 10.1038/ncomms10298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Jensen CG. J Cell Biol 1982; 92:540-58; PMID:7061596; http://dx.doi.org/ 10.1083/jcb.92.2.540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Maiato H, et al.. J Cell Biol 2004; 167:831-40; PMID:15569709; http://dx.doi.org/ 10.1083/jcb.200407090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Rubinstein B, et al.. Phys Biol 2009; 6:016005; PMID:19193975; http://dx.doi.org/ 10.1088/1478-3975/6/1/016005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Dumont S, et al.. Curr Biol 2009; 19:R749-61; PMID:19906577; http://dx.doi.org/ 10.1016/j.cub.2009.07.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Sharp DJ, et al.. Nature 2000; 407:41-7; PMID:10993066; http://dx.doi.org/ 10.1038/35024000 [DOI] [PubMed] [Google Scholar]