

ABSTRACT
Mutations in RPGRORF15 (retinitis pigmentosa GTPase regulator) are a major cause of inherited retinal degenerative diseases. RPGRORF15 (1152 residues) is a ciliary protein involved in regulating the composition and function of photoreceptor cilia. The mutational hotspot in RPGRORF15 is an unusual C-terminal domain encoded by exon ORF15, which is rich in polyglutamates and glycine residues (Glu-Gly domain) followed by a short stretch of basic amino acid residues (RPGRC2 domain; residues 1072-1152). However, the properties of the ORF15-encoded domain and its involvement in the pathogenesis of the disease are unclear. Here we show that RPGRORF15 is glutamylated at the C-terminus, as determined by binding to GT335, which recognizes glutamylated substrates. This reactivity is lost in two mouse mutants of Rpgr, which do not express RPGRORF15 due to disease-causing mutations in exon ORF15. Our results indicate that RPGRORF15 is posttranslationally glutamylated in the Glu-Gly domain and that the GT335 antibody predominantly recognizes RPGRORF15 in photoreceptor cilia.
KEY WORDS: Retina, RPGR, Cilia, Glutamylation, GT335
Summary: This study shows that the mutational hotspot of ciliary protein RPGRORF15, commonly associated with severe blindness, is posttranslationally glutamylated at its C-terminus and is a target of GT335.
INTRODUCTION
Retinitis pigmentosa (RP) is a group of genetically and clinically heterogeneous disorders of the eye. RP is characterized by night blindness due to the loss of rod photoreceptors, followed by complete blindness due the loss of cones (Bird, 1987; Fishman, 1978). RP is inherited in autosomal dominant, autosomal recessive, as well as X-linked manner, with over 200 causative genes identified to date (https://sph.uth.edu/retnet/). X-linked RP is one of the most severe forms with symptoms starting as early as in the first decade of life, which progress into complete blindness usually by the second decade of life (Fishman et al., 1988; Heckenlively et al., 1988). Mutations in two genes, RPGR and RP2, account for >80% of XLRP cases. Of these, RPGR mutations are found in ∼70% of cases. Moreover, 15-20% of simplex RP patients carry mutations in RPGR. These data make RPGR a common cause of RP, accounting for ∼20% of all RP cases (Churchill et al., 2013; Daiger et al., 2007).
Photoreceptors are polarized neurons with a distinct inner segment (IS) involved in protein synthesis and trafficking and photosensory outer segment (which is loaded with proteins involved in phototransduction). The outer segment extends from the apical region of the inner segment in the form of a narrow bridge-like structure called microtubule-based sensory or connecting cilium (Besharse and Bok, 2011). RPGRORF15 localizes to the connecting cilium and is likely involved in regulating the composition of the outer segment (Anand and Khanna, 2012; Rao et al., 2015).
RPGR is extensively alternatively spliced; however, there are two major RPGR isoforms: constitutive RPGR (RPGRconst) and RPGRORF15. Whereas the RPGRconst isoform encodes exons 1-19 of the RPGR gene (amino acids 1-815), the RPGRORF15 isoform terminates in an alternate exon ORF15, which includes exon 15 and part of intron 15 of RPGR (amino acids 1-1152) (Anand and Khanna, 2012). Both variants share a common N-terminal domain (encoded by exons 1-15) (Murga-Zamalloa et al., 2010). On the other hand, the C-terminal domain of RPGRconst encoded by exons 16-19 carries an isoprenylation motif (residues 812-815) whereas RPGRORF15 terminates in a long intron 15, which is a purine-rich region encoding a glutamic acid-glycine (Glu-Gly)-rich acidic domain (Vervoort et al., 2000). This domain is followed by a short stretch of basic amino acids, termed RPGRC2 domain (residues 1071-1152). Mutation analysis revealed that exon ORF15 is a mutational hotspot, accounting for 50-60% of XLRP cases (Vervoort et al., 2000). The majority of human disease-causing mutations in this exon are frameshift or nonsense variations, which result in a premature stop codon, whereas in-frame deletions or duplications or missense changes are tolerated.
Mouse and canine models of Rpgr have also been reported. An Rpgrnull mouse was generated by interrupting exons 4-6 of the Rpgr gene and was predicted to affect the expression of all RPGR isoforms (Hong et al., 2000). More recently, a naturally occurring Rpgrrd9 mouse model was characterized; this mouse carries a frameshift mutation in exon ORF15 resulting in a premature stop but does not seem to affect the expression of the RPGRconst isoform (Thompson et al., 2012). Two canine models carrying mutations in exon ORF15 have also been reported (Zhang et al., 2002). These models represent considerable phenotypic variability, which is consistent with heterogenic clinical presentation of RPGRORF15 patients.
Being a mutational hotspot, it is important to evaluate the properties of exon ORF15 of RPGR. In this study, we hypothesized that the polyglutamate rich domain of RPGRORF15 exhibits similar properties as the glutamate-rich regions of α-tubulin, whose C-terminal glutamate residues are posttranslationally glutamylated specifically in cilia.
RESULTS
Posttranslational modification of tubulin in the absence of RPGRORF15
Microtubules are polymers of α/β tubulin heterodimers (Mitchison and Kirschner, 1984). Tubulins undergo diverse posttranslational modifications in an organelle or cellular substructure-specific manner (Verhey and Gaertig, 2007). For example, ciliary microtubules are enriched in acetylation, detyrosination and glutamylation (Yu et al., 2015). These modifications regulate the structure and function of the microtubule cytoskeleton (Mendes Maia et al., 2014; O'hagan and Barr, 2012). Given that RPGR associates with microtubule-based assemblies and that loss of RPGR alters microtubule-based photoreceptor ciliary trafficking (Anand and Khanna, 2012; Rao et al., 2015), we examined whether tubulin modifications are altered in the absence of RPGR. To this end, we used two Rpgr-mutant lines; Rpgrnull and Rpgrrd9. The Rpgrnull mouse does not exhibit expression of RPGRconst and RPGRORF15, whereas the Rpgrrd9 mice only express the RPGRconst (∼90 kDa) isoform (Rao et al., 2015; Thompson et al., 2012). Immunoblot analysis of retinal extracts from wild-type (WT), Rpgrnull, and Rpgrrd9 mouse retinas using antibodies against various post-translationally modified tubulin revealed no changes in the levels of acetylated α-tubulin, detyrosinated tubulin or glutamylated tubulin (B3 and GT335) (Fig. 1A-D).
Fig. 1.

GT335 detects RPGRORF15. (A-C) Immunoblot (IB) analysis of retinal extracts from wild type (WT), Rpgrnull and Rpgrrd9 mice was performed using antibodies against indicated forms of tubulin. An equal amount of protein extract (30 µg) was loaded in each lane. (D-F) Retinal extracts from indicated mouse strains were analyzed by SDS-PAGE and immunoblotting using anti-GT335 (D), anti-RPGR (E) or anti-polyE (F) antibodies. Arrow in D indicates the tubulin-reactive band whereas arrowhead points to the RPGRORF15 band. Three independent replicates were performed for these experiments.
RPGRORF15 is a target of GT335
During our analysis, we found that the GT335 antibody, in addition to detecting the glutamylated tubulin-specific band at ∼50 kDa, recognized a higher molecular weight band (∼200 kDa) in WT mouse retinal extracts (Fig. 1D). This band was of the same molecular weight as the RPGRORF15-immunoreactive band, as determined by western blotting using anti-RPGR antibody (Fig. 1E). We did not detect a similar immunoreactive band (∼200 kDa) using B3 antibody (not shown). Previous studies showed that in addition to tubulins, GT335 recognizes other targets of glutamylation, such as nucleosome assembly proteins, NAP1 and NAP2 (Regnard et al., 2000). However, B3 antibody specifically detects polyglutamylated α-tubulin (Van Dijk et al., 2007).
The GT335 antibody was raised against an octapeptide EGEGE*EEG, which is modified by the addition of two glutamyl subunits on the fifth E (*). The C-terminus of RPGRORF15, on the other hand, predominantly carries GEEEEG and GEEEG repeats. These repeats could potentially be substrates for glutamylation. We thus asked whether the GT335 antibody cross-reacts with an unknown protein of the same molecular weight as RPGRORF15 or specifically recognizes the C-terminal domain of RPGRORF15. We examined the expression of this band in the Rpgrnull and the Rpgrrd9 retinas. Our hypothesis was that if this band were a cross-reacting species, then we would observe it even in the absence of RPGRORF15; however, if this reactivity were specific, then we would not observe this high molecular weight band in Rpgrrd9 (frameshift mutation in exon ORF15) and Rpgrnull retinal extracts. Immunoblot analysis revealed that the GT335-immunoreactive band was undetectable in the Rpgrnull and Rpgrrd9 retinal extracts (Fig. 1D). The GT335 antibody is a well-characterized antibody, which specifically recognizes the first branch point glutamate added to the target residue (Van Dijk et al., 2007; Wolff et al., 1992). Thus, this reactivity does not distinguish between mono- and polyglutamylated RPGRORF15. To clarify this, we performed immunoblot analysis using polyE antibody, which recognizes long polyglutamylated side chains (Magiera and Janke, 2013). As shown in Fig. 1F, polyE antibody did not detect the high molecular band; however, tubulin-specific bands at ∼50 kDa were detected. These results indicate that RPGRORF15 is most likely monoglutamylated.
RPGRORF15 associates with GT335 in the retina
To further validate the binding of GT335 to RPGRORF15, we performed immunoprecipitation from mouse retina using anti-RPGRORF15 antibody followed by immunoblotting using GT335 antibody or anti-RPGRORF15 antibody. RPGR immunoprecipitation pulled down both RPGRORF15 and RPGRconst isoforms from the wild type retinal extracts but not from Rpgrnull mice; however, the GT335-immunoreactive band coincided with anti-RPGRORF15-reactive band (Fig. 2A, right panel). Furthermore, immunofluorescence analysis of wild-type mouse retina using anti-RPGR and GT335 antibodies showed that they co-localize at the connecting cilium (CC) (Fig. 2B).
Fig. 2.
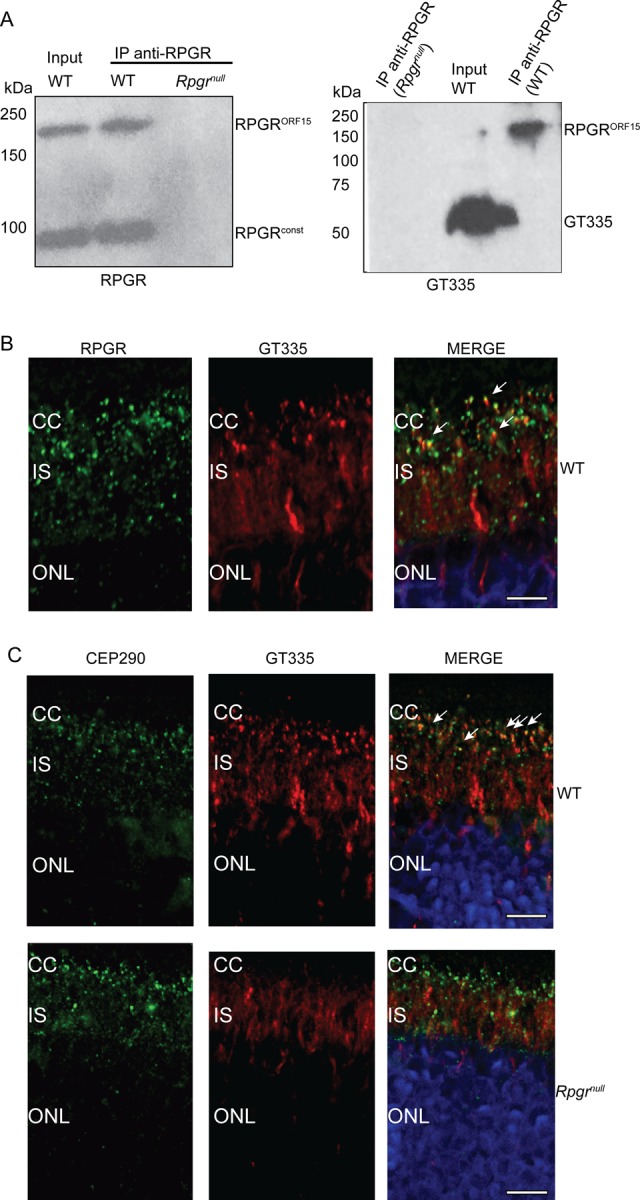
GT335 association and reactivity in Rpgrnull retina. (A) Retinal extracts from wild type (WT) and Rpgrnull mice were subjected to immunoprecipitation (IP) using anti-RPGR antibody followed by SDS-PAGE and immunoblotting using anti-RPGR (left) or GT335 (right) antibody. Input lane shows 10% of the protein extract used in IP. (B,C) Cryosections of wild type (WT; B; upper panel, C) and 2-month-old Rpgrnull (C) mouse retinas were stained with anti-RPGR (B; green) or anti-CEP290 (panel C; green), and anti-GT335 (red; B,C) antibodies. Arrows in merge point to co-localization of RPGR or CEP290 with GT335-specific signal. Scale bar: 2 µm in B; 5 µm in C. Nuclei were stained with Hoechst (blue). CC, connecting cilium; IS, photoreceptor inner segment; ONL, photoreceptor outer nuclear layer. These experiments were repeated three times with independent mouse retinal samples.
Immunofluorescence analysis of GT335 in Rpgrnull mouse retina
GT335 recognizes polyglutamylated tubulin in microtubules, which are abundant in the cilia. To test whether the ciliary staining of GT335 is altered in the absence of RPGR, we performed immunofluorescence analysis of wild-type and Rpgrnull mouse retina. Our analysis revealed reduced staining of GT335 in the Rpgrnull photoreceptor connecting cilium. As shown in Fig. 2C, co-localization of GT335 with anti-CEP290 (a connecting cilium marker; arrows pointing to yellow spots) (Murga-Zamalloa et al., 2011) is observed in wild type retinal sections but not in the Rpgrnull retinas, which predominantly exhibits green signal corresponding to anti-CEP290 antibody, in the connecting cilium. These data indicate that in photoreceptor cilia, the ciliary staining of GT335 is largely mediated by binding to glutamylated RPGR. Such staining is diminished in the Rpgrnull mouse retina.
Human RPGRORF15 is immunoreactive to GT335
The studies described above were carried out using mouse retinal extracts. We also wanted to validate these findings in human RPGR protein. Using immunoreactivity to GT335 as a tractable assay system, we asked whether GT335 also recognizes both major variants of human RPGR: RPGRconst and RPGRORF15 protein. We transiently transfected hTERT-RPE1 cells with mammalian expression constructs encoding Xpress (Xp)-tagged human RPGRconst and RPGRORF15 (Fig. 3A), followed by SDS-PAGE and immunoblotting of protein extracts using anti-Xp or GT335 antibodies. As shown in Fig. 3B, GT335-immunoreactivity was detected only in cells expressing Xp-RPGRORF15. Immunoblotting using anti-Xpress antibody validated the expression of the recombinant human RPGR proteins. These results suggest that the C-terminal glutamic acid rich domain of RPGRORF15 is a likely target for glutamylation.
Fig. 3.
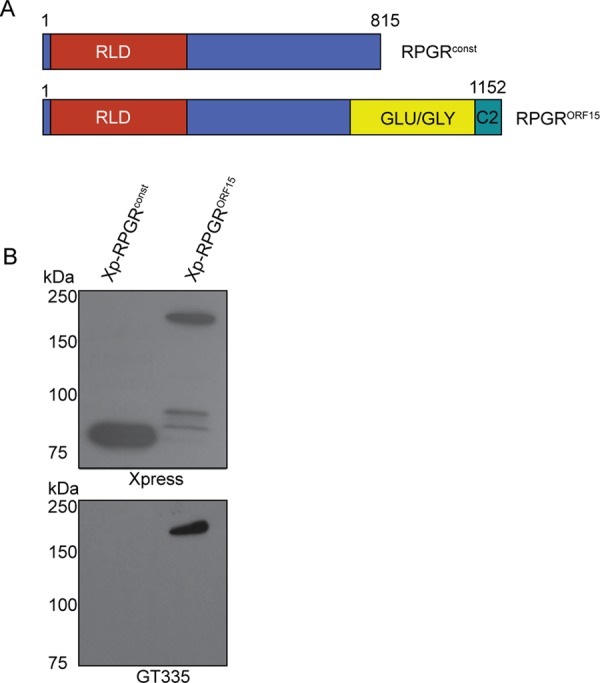
Human RPGRORF15 reacts with GT335. (A) This panel depicts a schematic representation of the human RPGRconst and RPGRORF15 isoforms. Numbers denote the positons of amino acid residues. RLD, RCC1-like domain; Glu-Gly, glutamic acid and glycine rich domain. (B) hTERT-RPE1 cells were transiently transfected with constructs encoding Xp-RPGRconst or Xp-RPGRORF15. An equal amount of cell extract was analyzed by SDS-PAGE and immunoblotting using anti-Xpress or GT335 antibody. All experiments were repeated three independent times.
Disease-causing mutations in RPGRORF15 alter its binding to GT335
We hypothesized that disease-associated mutations in exon ORF15, which alter the polyglutamate repeats affect its glutamylation and thus, reactivity to GT335. To test this, we transiently transfected hTER-RPE1 cells with constructs encoding Xp-tagged RPGRORF15 and truncated versions of RPGRORF15: RPGRORF15-1071X and RPGRORF15-853X (Fig. 4A). Immunoblot analysis of protein extracts using GT335 antibody revealed that whereas full length RPGRORF15 was recognized by GT335, RPGRORF15-Glu1071X exhibited ∼50% reactivity and RPGRORF15-Glu853X exhibited only ∼0.1% immunoreactivity to GT335 (Fig. 4B).
Fig. 4.

Human disease mutations in RPGRORF15 alter binding to GT335. (A) Schematic representation of the location of the human exon ORF15 mutations. RLD, RCC1-like domain; Glu-Gly, glutamic acid and glycine rich domain; C2, RPGRC2 domain. (B) hTERT-RPE1 cells were transiently transfected with constructs encoding Xpress-tagged full-length RPGRORF15 or two disease-causing mutants; RPGRORF15-1071X or RPGRORF15-853X. Equal amount of protein extracts from these cells were analyzed by SDS-PAGE and immunoblotting using anti-Xpress (upper panel) or anti-GT335 (lower panel) antibody. Asterisk indicates degraded protein product. Densitometric analysis was performed to quantify the immunoblot signal. The data are represented as ratio relative to the intensity of GT335-immunoreactivity with full-length RPGRORF15, set as 1. Error bars represent standard deviation. *P<0.001; **P<0.0001. Data are representative of three independent experiments.
DISCUSSION
Our studies reveal two key findings: (i) a novel molecule that is recognized by GT335. In fact, RPGRORF15 is likely the fifth molecule that is recognized by GT335; the first four being α-tubulin, β-tubulin, NAP1 and NAP2 (Regnard et al., 2000); (ii) sequence alterations in the Glu-Gly domain of RPGRORF15 alter its glutamylation. However, the function of glutamylated RPGRORF15 is not known. We propose that glutamylated RPGRORF15 regulates the integrity of the multiprotein complexes at the cilium and modulates their entry or retention inside the cilium. However, human disease mutations in the polyglutamate-rich region appear to alter the length of this region (and the amount of polyglutamates) and are associated with relatively mild retinal dysfunction in patients. Thus, alterations in the glutamylation of RPGRORF15 may compromise but not eliminate its function. Evaluation of complete loss of the ORF15 domain on ciliary function will provide insights into the precise role of glutamylation of RPGRORF15.
What is the mechanism of glutamylation of RPGRORF15? Glutamylation is an evolutionarily conserved and is a widely distributed modification involved in diverse functions (Janke et al., 2008; Van Dijk et al., 2008). Moreover, tubulin glutamylation occurs at the C-terminal glutamate-rich region (Edde et al., 1990) and is carried out by tubulin tyrosine ligase-like (TTLL) family of proteins (Raunser and Gatsogiannis, 2015). Recently, a member of the TTLL family, TTLL5, was reported to be involved in human retinal degeneration (Sergouniotis et al., 2014). We propose that TTLL5 or another TTLL family member is a candidate glutamylase for RPGRORF15. A Ttll5ko mouse model was reported to have sperm defects; however, retinal defects were not detected in this model (Lee et al., 2013). It is possible that the defects could occur at ages older than those tested in that study. Further studies to test these scenarios have the potential to reveal critical insights into the pathogenesis of retinal degeneration due to defective posttranslational modifications of ciliary proteins. Our results showing that RPGRORF15-Glu1071X exhibited ∼50% reactivity and RPGRORF15-Glu853X exhibited only ∼0.1% immunoreactivity to GT335 suggest the involvement of C2 domain in efficient glutamylation of RPGRORF15; however, both the Glu-Gly domain and the C2 domain may be critical for glutamylation of RPGRORF15. We propose that the C2 domain may act as a binding site for the enzymes and that both the Glu-Gly domain and C2 domain work in concert for efficient binding and glutamylation reactions.
Although our data indicate that RPGRORF15 is likely monoglutamylated, we cannot rule out the possibility that RPGRORF15 is polyglutamylated. This is because the C-terminus of RPGRORF15 predominantly carries GEEEEG and GEEEG repeats, which could be glutamylated in tandem. However, identification of the glutamylase will provide further insights into the glutamylation status of RPGRORF15. For example, TTLL4, TTLL5, and TTLL7 are side-chain initiating polyglutamylases, TTLL6, TTLL11 and TTLL13 preferentially elongate the side chains. Nonetheless, glutamylation of RPGRORF15 may alter the function of RPGR by modulating its ability to interact with other proteins at the cilium due to changes in the net charge of the Glu-Gly domain. Further studies are underway to assess the effect of glutamylation of RPGRORF15 on its ability to maintain the integrity of its interactome and/or interact with additional proteins. Overall, these studies will provide new insights into the mode of regulation of ciliary protein trafficking and pathogenesis of associated ciliopathies.
MATERIALS AND METHODS
Animals
All studies involving animals were approved by the Institutional Animal Care and Use Committee of UMASS Medical School. Wild type C57BL6/J and Rpgrrd9 mice were procured from Jackson Labs; Rpgrnull mice were obtained from Dr Tiansen Li (National Eye Institute). Only hemizygous male mice were used in this study as Rpgr is an X-linked gene.
Antibodies
The GT335 (1:500 dilution) and polyE (1:200 dilution) antibodies were obtained from AdipoGen (San Diego, CA); anti-anti-acetylated α-tubulin (1:1000 dilution) and anti-polyglutamylated tubulin B3 (1:500 dilution) were obtained from Sigma-Aldrich; anti-detyrosinated antibody was obtained from Abcam; anti-Xpress antibody (1:200 dilution) was procured from Invitrogen. Anti-RPGR antibody (1:500 dilution) was raised against an N-terminal epitope, which is common to both RPGRconst and RPGRORF15 isoforms. Detailed characterization of this antibody was previously reported (Ghosh et al., 2010; Rao et al., 2015).
Plasmids, cell culture, transient transfection and IP
The human cDNA encoding full-length human RPGRconst and RPGRORF15 were cloned into pcDNA4 (Invitrogen), which expressed N-terminally X-press-tagged recombinant proteins, and sequence verified. hTERT-RPE1 cells (ATCC) were maintained in DMEM/F12 (Invitrogen) supplemented with 10% fetal bovine serum and penicillin/streptomycin and tested for contamination. Transient transfections were performed using Lipofectamine 2000 (Invitrogen). Cells were lysed 48 h post-transfection in lysis buffer containing 25 mM Tris, 150 mM NaCl, 1 mM EDTA, 1% NP-40, 5% glycerol (pH 7.4) with Complete protease inhibitors (Roche) and lysates were spun at 18,000×g for 15 min at 4°C. Equal amount of protein was analyzed by SDS-PAGE and immunoblotting, as described (Khanna et al., 2005). Immunoprecipitation of mouse retinal extracts was performed, as described (Khanna et al., 2005). All experiments were repeated three independent times.
Immunofluorescence
Staining of mouse retina was performed as described (Li et al., 2013). Cryosections of mouse retinas fixed in 4% paraformaldehyde were permeabilized and blocked using 5% normal goat serum followed by incubation with primary antibody overnight at 4°C. Slides were then washed three times with phosphate buffered saline and further incubated with secondary antibody for 1 h at room temperature. Hoechst 33342 (Life Technologies) was added (1 µg/ml) to label the nuclei and the sections were then mounted (Fluoromount; Electron Microscopy Services, Hatfield, PA) under glass coverslips and visualized using Leica TCS Sp5 II laser microscope (Leica Microsystems).
Acknowledgements
We thank Dr Carlos A. Murga-Zamalloa for constructive discussions during the early stages of this work.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
H.K. designed research and wrote the paper; K.N.R., M.A. and H.K. performed experiments.
Funding
This work is supported by grants from the National Institutes of Health [grant number RO1-EY022372] and Foundation Fighting Blindness [grant number C-CMM-0815-0675-UMASS04].
References
- Anand M. and Khanna H. (2012). Ciliary transition zone (TZ) proteins RPGR and CEP290: role in photoreceptor cilia and degenerative diseases. Expert Opin. Ther. Targets 16, 541-551. 10.1517/14728222.2012.680956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Besharse J. C. and Bok D. (2011). The Retina and Its Disorders. Amsterdam; Boston: Academic Press. [Google Scholar]
- Bird A. C. (1987). Clinical investigation of retinitis pigmentosa. Prog. Clin. Biol. Res. 247, 3-20. [PubMed] [Google Scholar]
- Churchill J. D., Bowne S. J., Sullivan L. S., Lewis R. A., Wheaton D. K., Birch D. G., Branham K. E., Heckenlively J. R. and Daiger S. P. (2013). Mutations in the X-linked retinitis pigmentosa genes RPGR and RP2 found in 8.5% of families with a provisional diagnosis of autosomal dominant retinitis pigmentosa. Invest. Ophthalmol. Vis. Sci. 54, 1411-1416. 10.1167/iovs.12-11541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daiger S. P., Bowne S. J. and Sullivan L. S. (2007). Perspective on genes and mutations causing retinitis pigmentosa. Arch. Ophthalmol. 125, 151-158. 10.1001/archopht.125.2.151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edde B., Rossier J., Le Caer J. P., Desbruyeres E., Gros F. and Denoulet P. (1990). Posttranslational glutamylation of alpha-tubulin. Science 247, 83-85. 10.1126/science.1967194 [DOI] [PubMed] [Google Scholar]
- Fishman G. A. (1978). Retinitis pigmentosa. Genetic percentages. Arch. Ophthalmol. 96, 822-826. 10.1001/archopht.1978.03910050428005 [DOI] [PubMed] [Google Scholar]
- Fishman G. A., Farber M. D. and Derlacki D. J. (1988). X-linked retinitis pigmentosa Profile of clinical findings. Arch. Ophthalmol. 106, 369-375. 10.1001/archopht.1988.01060130395029 [DOI] [PubMed] [Google Scholar]
- Ghosh A. K., Murga-Zamalloa C. A., Chan L., Hitchcock P. F., Swaroop A. and Khanna H. (2010). Human retinopathy-associated ciliary protein retinitis pigmentosa GTPase regulator mediates cilia-dependent vertebrate development. Hum. Mol. Genet. 19, 90-98. 10.1093/hmg/ddp469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heckenlively J. R., Yoser S. L., Friedman L. H. and Oversier J. J. (1988). Clinical findings and common symptoms in retinitis pigmentosa. Am. J. Ophthalmol. 105, 504-511. 10.1016/0002-9394(88)90242-5 [DOI] [PubMed] [Google Scholar]
- Hong D.-H., Pawlyk B. S., Shang J., Sandberg M. A., Berson E. L. and Li T. (2000). A retinitis pigmentosa GTPase regulator (RPGR)-deficient mouse model for X-linked retinitis pigmentosa (RP3). Proc. Natl. Acad. Sci. USA 97, 3649-3654. 10.1073/pnas.97.7.3649 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janke C., Rogowski K. and Van Dijk J. (2008). Polyglutamylation: a fine-regulator of protein function? ‘Protein Modifications: beyond the usual suspects’ review series. EMBO Rep. 9, 636-641. 10.1038/embor.2008.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khanna H., Hurd T. W., Lillo C., Shu X., Parapuram S. K., He S., Akimoto M., Wright A. F., Margolis B., Williams D. S. et al. (2005). RPGR-ORF15, which is mutated in retinitis pigmentosa, associates with SMC1, SMC3, and microtubule transport proteins. J. Biol. Chem. 280, 33580-33587. 10.1074/jbc.M505827200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee G.-S., He Y., Dougherty E. J., Jimenez-Movilla M., Avella M., Grullon S., Sharlin D. S., Guo C., Blackford J. A. Jr, Awasthi S. Jr et al. (2013). Disruption of Ttll5/stamp gene (tubulin tyrosine ligase-like protein 5/SRC-1 and TIF2-associated modulatory protein gene) in male mice causes sperm malformation and infertility. J. Biol. Chem. 288, 15167-15180. 10.1074/jbc.M113.453936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L., Khan N., Hurd T., Ghosh A. K., Cheng C., Molday R., Heckenlively J. R., Swaroop A. and Khanna H. (2013). Ablation of the X-linked retinitis pigmentosa 2 (Rp2) gene in mice results in opsin mislocalization and photoreceptor degeneration. Invest. Ophthalmol. Vis. Sci. 54, 4503-4511. 10.1167/iovs.13-12140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magiera M. M. and Janke C. (2013). Investigating tubulin posttranslational modifications with specific antibodies. Methods Cell Biol. 115, 247-267. 10.1016/B978-0-12-407757-7.00016-5 [DOI] [PubMed] [Google Scholar]
- Mendes Maia T., Gogendeau D., Pennetier C., Janke C. and Basto R. (2014). Bug22 influences cilium morphology and the post-translational modification of ciliary microtubules. Biol. Open 3, 138-151. 10.1242/bio.20146577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchison T. and Kirschner M. (1984). Microtubule assembly nucleated by isolated centrosomes. Nature 312, 232-237. 10.1038/312232a0 [DOI] [PubMed] [Google Scholar]
- Murga-Zamalloa C. A., Atkins S. J., Peranen J., Swaroop A. and Khanna H. (2010). Interaction of retinitis pigmentosa GTPase regulator (RPGR) with RAB8A GTPase: implications for cilia dysfunction and photoreceptor degeneration. Hum. Mol. Genet. 19, 3591-3598. 10.1093/hmg/ddq275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murga-Zamalloa C. A., Ghosh A. K., Patil S. B., Reed N. A., Chan L. S., Davuluri S., Peränen J., Hurd T. W., Rachel R. A. and Khanna H. (2011). Accumulation of the Raf-1 kinase inhibitory protein (Rkip) is associated with Cep290-mediated photoreceptor degeneration in ciliopathies. J. Biol. Chem. 286, 28276-28286. 10.1074/jbc.M111.237560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'hagan R. and Barr M. M. (2012). Regulation of tubulin glutamylation plays cell-specific roles in the function and stability of sensory cilia. Worm 1, 155-159. 10.4161/worm.19539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao K. N., Li L., Anand M. and Khanna H. (2015). Ablation of retinal ciliopathy protein RPGR results in altered photoreceptor ciliary composition. Sci. Rep. 5, 11137 10.1038/srep11137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raunser S. and Gatsogiannis C. (2015). Deciphering the tubulin code. Cell 161, 960-961. 10.1016/j.cell.2015.05.004 [DOI] [PubMed] [Google Scholar]
- Regnard C., Desbruyeres E., Huet J. C., Beauvallet C., Pernollet J. C. and Edde B. (2000). Polyglutamylation of nucleosome assembly proteins. J. Biol. Chem. 275, 15969-15976. 10.1074/jbc.M000045200 [DOI] [PubMed] [Google Scholar]
- Sergouniotis P. I., Chakarova C., Murphy C., Becker M., Lenassi E., Arno G., Lek M., MacArthur D. G., UCL-Exomes Consortium, Bhattacharya S. S. et al. (2014). Biallelic variants in TTLL5, encoding a tubulin glutamylase, cause retinal dystrophy. Am. J. Hum. Genet. 94, 760-769. 10.1016/j.ajhg.2014.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson D. A., Khan N. W., Othman M. I., Chang B., Jia L., Grahek G., Wu Z., Hiriyanna S., Nellissery J., Li T. et al. (2012). Rd9 is a naturally occurring mouse model of a common form of retinitis pigmentosa caused by mutations in RPGR-ORF15. PLoS ONE 7, e35865 10.1371/journal.pone.0035865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Dijk J., Rogowski K., Miro J., Lacroix B., Eddé B. and Janke C. (2007). A targeted multienzyme mechanism for selective microtubule polyglutamylation. Mol. Cell 26, 437-448. 10.1016/j.molcel.2007.04.012 [DOI] [PubMed] [Google Scholar]
- Van Dijk J., Miro J., Strub J.-M., Lacroix B., Van Dorsselaer A., Edde B. and Janke C. (2008). Polyglutamylation is a post-translational modification with a broad range of substrates. J. Biol. Chem. 283, 3915-3922. 10.1074/jbc.M705813200 [DOI] [PubMed] [Google Scholar]
- Verhey K. J. and Gaertig J. (2007). The tubulin code. Cell Cycle 6, 2152-2160. 10.4161/cc.6.17.4633 [DOI] [PubMed] [Google Scholar]
- Vervoort R., Lennon A., Bird A. C., Tulloch B., Axton R., Miano M. G., Meindl A., Meitinger T., Ciccodicola A. and Wright A. F. (2000). Mutational hot spot within a new RPGR exon in X-linked retinitis pigmentosa. Nat. Genet. 25, 462-466. 10.1038/78182 [DOI] [PubMed] [Google Scholar]
- Wolff A., De Nechaud B., Chillet D., Mazarguil H., Desbruyeres E., Audebert S., Edde B., Gros F. and Denoulet P. (1992). Distribution of glutamylated alpha and beta-tubulin in mouse tissues using a specific monoclonal antibody, GT335. Eur. J. Cell Biol. 59, 425-432. [PubMed] [Google Scholar]
- Yu I., Garnham C. P. and Roll-Mecak A. (2015). Writing and reading the Tubulin Code. J. Biol. Chem. 290, 17163-17172. 10.1074/jbc.R115.637447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Q., Acland G. M., Wu W. X., Johnson J. L., Pearce-Kelling S., Tulloch B., Vervoort R., Wright A. F. and Aguirre G. D. (2002). Different RPGR exon ORF15 mutations in Canids provide insights into photoreceptor cell degeneration. Hum. Mol. Genet. 11, 993-1003. 10.1093/hmg/11.9.993 [DOI] [PubMed] [Google Scholar]