

Abstract
Strains of the genus Gramella (family Flavobacteriacae, phylum Bacteroidetes) were isolated from marine habitats such as tidal flat sediments, coastal surface seawater and sea urchins. Flavobacteriaceae have been shown to be involved in the decomposition of plant and algal polysaccharides. However, the potential to decompose polysaccharides may differ tremendously even between species of the same genus. Gramella echinicola KMM 6050T (DSM 19838T) and Gramella portivictoriae UST040801-001T (DSM 23547T) have genomes of similar lengths, similar numbers of protein coding genes and RNA genes. Both genomes encode for a greater number of peptidases compared to ’G. forsetii’. In contrast to the genome of ’G. forsetii’, both genomes comprised a smaller set of CAZymes. Seven polysaccharide utilization loci were identified in the genomes of DSM 19838T and DSM 23547T. Both Gramella strains hydrolyzed starch, galactomannan, arabinoxylan and hydroxyethyl-cellulose, but not pectin, chitosan and cellulose (Avicel). Galactan and xylan were hydrolyzed by strain DSM 19838T, whereas strain DSM 23547T hydrolyzed pachyman and carboxy-methyl cellulose. Conclusively, both Gramella type strains exhibit characteristic physiological, morphological and genomic differences that might be linked to their habitat. Furthermore, the identified enzymes mediating polysaccharide decomposition, are of biotechnological interest.
Electronic supplementary material
The online version of this article (doi:10.1186/s40793-016-0163-9) contains supplementary material, which is available to authorized users.
Keywords: Carbohydrate active enzyme, Polysaccharide, Bioethanol, Gliding motility, Cellulose, Marine, Flavobacteriaceae, Bacteroidetes, GEBA, KMG I
Introduction
Strain UST040801-001T (=DSM 23547T = JCM 13192T = NBRC 101534T = NRRLB-41137T) is the type strain of G. portivictoriae [1] and strain KMM 6050T (=DSM 19838T =JCM 13510T =KCTC 12278T =LMG 22585T =NBRC 100593T) is the types train of G. echinicola [2], the type species of Gramella [2] of the family Flavobacteriaceae [3, 4]. G. echinicola KMM 6050T was isolated from the sea urchin Strongylocentrotus intermedius of the Sea of Japan [2], whereas G. portivictoriae UST040801-001T was isolated from sediment of the Victoria Harbor, Hong Kong [1]. All other Gramella known strains were isolated from marine habitats, such as tidal flat sediment [5–8] and coastal surface seawater [9, 10]. Many Flavobacteriaceae have been shown to harbour a great set of carbohydrate active enzymes, such as Zobelliagalactinovorans [11], Formosa agariphila [12], ’Gramella forsetii’ KT0803 [13]. However, the set of CAZymes within a genus may differ tremendously, as shown for Polaribacter [14] and Flavobacterium [15, 16]. Thus, we selected these Flavobacteriaceae type strains from different marine habitats to gain insights into their unknown polysaccharide decomposition potential (other than starch, cellulose and chitin).
Here we present the different sets of carbohydrate active enzymes, polysaccharide-utilization loci and peptidases of both Gramella genomes and a summary of their current classification, the set of known phenotypic features and a description of the permanent draft genome sequence and annotation derived from cultures of strains DSM 19838T and DSM 23547T. Furthermore, we investigated the polar lipid profiles, cell surface structures and gliding motility of these strains, as well as the hydrolysis of certain polysaccharides.
Organism information
Classification and features
The draft genome of G. echinicolaDSM 19838T has one full-length and one partial 16S rRNA gene sequence identical with the sequence from the original species description (AB681204, AY608409). The draft genome of G. portivictoriaeDSM 23547T has one full-length 16S rRNA gene sequence identical with the sequence from strain NBRC 101534T (AB681471) and 99 % similar with the sequence in the original species description (DQ002871) [1]. Based on 16S rRNA gene sequence similarity, closely related strains were TW-JL-80 (DQ073100, 98.1 %) from the South China Sea [17], MAR_2010_163 (JX854363, 97.3 %) from the North Sea [18] and the clone Vis_St18_35 (FN433421, 98.3 %) from the North Atlantic subtropical gyre [19]. A summary of the classification and general features of G. echinicolaDSM 19838T and G. portivictoriaeDSM 23547T is shown in Table 1.
Table 1.
Classification and general features of G. echinicola DSM 19838T and G. portivictoriae DSM 23547T in accordance with the MIGS recommendations [60], as developed by [25], List of Prokaryotic names with Standing in Nomenclature [61, 62] and the Names for Life database [63]
| MIGS ID | Property | DSM 19838T | DSM 23547T | ||
|---|---|---|---|---|---|
| Term | Evidence codea | Term | Evidence codea | ||
| Current | Domain Bacteria | TAS [64] | Domain Bacteria | TAS [64] | |
| classification | Phylum Bacteroidetes | TAS [65, 66] | Phylum Bacteroidetes | TAS [65, 66] | |
| Class Flavobacteriia | TAS [67, 68] | Class Flavobacteriia | TAS [67, 68] | ||
| Order Flavobacteriales | TAS [4, 69] | Order Flavobacteriales | TAS [4, 69] | ||
| Family Flavobacteriaceae | TAS [3, 4] | Family Flavobacteriaceae | TAS [3, 4] | ||
| Genus Gramella | TAS [2] | Genus Gramella | TAS [2] | ||
| Species Gramella echinicola |
TAS [2] | Species Gramella portivictoriae |
TAS [1] | ||
| Type strain KMM 6050T |
TAS [2] | Type strain UST040801-001T |
TAS [1] | ||
| Gram-stain | Negative | TAS [2] | Negative | TAS [1] | |
| Cell shape | Rod-shaped | TAS [2] | Rod-shaped | TAS [1] | |
| Motility | Motile, gliding | TAS [2] | Motile, gliding | TAS [1] | |
| Sporulation | Non-spore forming | TAS [2] | Non-spore forming | TAS [1] | |
| Temperature range | Mesophilic, 4–37 °C | TAS [2] | Mesophilic, 4–36 °C | TAS [1] | |
| Optimum temperature | 23–25 °C | TAS [2] | 28–30 °C | TAS [1] | |
| pH range; optimum | 4–11, 7–8 | TAS [2] | 6–10, 7–8 | TAS [1] | |
| MIGS-22 | Oxygen requirement | Strictly aerobic | TAS [2] | Strictly aerobic | TAS [1] |
| Carbon source | Carbohydrates, peptides | TAS [2] | Carbohydrates, peptides | TAS [1] | |
| Energy source | Chemoheterotroph | TAS [2] | Chemoheterotroph | TAS [1] | |
| MIGS-6 | Habitat | Marine, host, sea urchin | TAS [2] | Marine, sediment | TAS [1] |
| MIGS-6.3 | Salinity (% NaCl, w/v) | 1–15 % | TAS [2] | 1–6 % | TAS [1] |
| MIGS-15 | Biotic relationship | Commensal | TAS [2] | Free-living | TAS [1] |
| MIGS-14 | Pathogenicity | Not reported | NAS | Not reported | NAS |
| Biosafety level | 1 | TAS [70] | 1 | TAS [70] | |
| MIGS-4 | Geographic location | Troitsa Bay, Gulf of Peter the Great, Sea of Japan | TAS [2] | Victoria Harbour, Hong Kong | TAS [1] |
| MIGS-5 | Sample collection time | 1. Sep. 2002 | NAS | Before 2005 | NAS |
| MIGS-4.1 | Latitude | 42.64 | NAS | 22.31 | NAS |
| MIGS-4.2 | Longitude | 131.10 | NAS | 114.12 | NAS |
| Depth | 3 m | TAS [2] | not reported | ||
aEvidence codes - TAS traceable author statement (i.e., a direct report exists in the literature), NAS non-traceable author statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). Evidence codes are from the Gene Ontology project [71]
Figure 1 depicts a 16S rRNA gene sequence phylogenomic tree of the genera Gramella, Zunongwangia and other closely related Flavobacteriaceae. Gramella spp. Nedashkovskaya et al. 2005 are Gram-stain negative, rod-shaped, strictly aerobic Flavobacteriaceae that are cytochrom-oxidase and catalase positive, move by gliding, produce non-diffusible carotenoid pigments, but not flexirubin-like pigments [2]. G. echinicolaDSM 19838T produces extracellular polymeric substances, whereas G. portivictoriaeDSM 23547T produces appendages (Fig. 2). Colonies of both of these Gramella species are circular, convex with entire translucent margins and yellow–orange in color on marine agar (Fig. 2). Both strains grow at pH 6–10 and between 4 °C and 36 °C, with a temperature optimum at 23–25 °C for G. echinicola and 28–30 °C for G. portivictoriae [1, 2]. G. echinicola is able to grow in medium of higher salinity (1–15 % (w/v) NaCl) than G. portivictoriae (1–6 % (w/v) NaCl) [1, 2]. Both Gramella strains utilize d-arabinose, l-arabinose, d-glucose and d-sucrose [1, 2], d-fructose and trehalose [8]. G. portivictoriae UST040801-001T utilizes d-galactose, glycerol, d-mannitol, d-melibiose, d-sorbitol and starch [1], whereas G. echinicolaJCM 13510T utilizes d-xylose [7], but not d-lactose, d-mannose, d-mannitol, inositol, sorbitol, malonate and citrate [2]. A list of carbon sources utilized by both strains using the Biolog GN2 plate can be seen in Cho et al. [5].
Fig. 1.
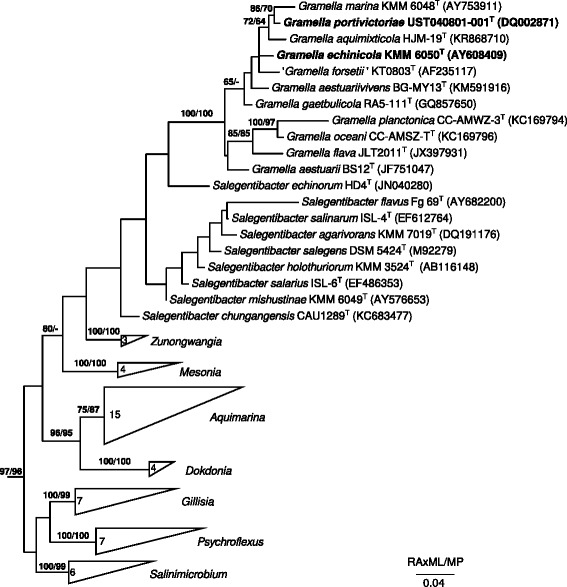
Phylogenetic tree the genus Gramella and closely related genera of the family Flavobacteriaceae. The tree was inferred from 1,409 aligned characters of the 16S rRNA gene sequence under the maximum likelihood (ML) and maximum parsimony [MP] criterion as previously described by Göker et al. [51]. The sequences of the LTP v. 121 database [52, 53] and from GenBank were aligned in ARB [54] using the SINA aligner [39] and manually corrected. The branches are scaled in terms of expected number of substitutions per site. Numbers adjacent to the branches are support values from 1,000 ML bootstrap replicates (left) and from 1,000 maximum-parsimony bootstrap replicates (right) if larger than 60 % [51]. Numbers in wedges represent the numbers of sequences. The tree was rooted using type strains of the genera Doktonia, Aquimarina, Salinimicrobium, Psychroflexus, Gillisia and Mesonia
Fig. 2.
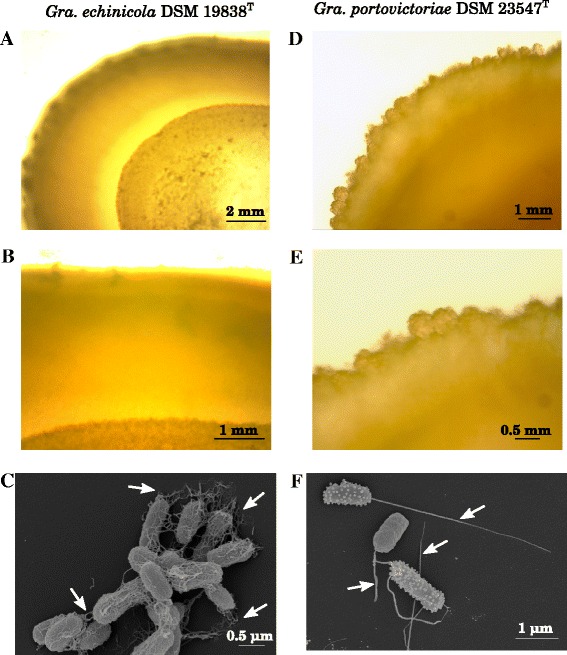
Gliding motility and scanning electron micrographs of G. echinicola DSM 19838T and G. portivictoriae DSM 23547T. (A-F) DSM 19838T and DSM 23547T were incubated on bacto marine soft agar (0.3 % agar) at 25 °C to visualize the gliding motility of these Gramella. (G-H) DSM 19838T and DSM 23547T were cultured in bacto marine broth at 25 °C and visualized by scanning electron microscopy. DSM 19838T expressed extracellular polymeric substances, EPS (arrows) whereas DSM 23547T produced appendages (arrows)
Chemotaxonomic data
Major fatty acids (>5 % of total) of G. echinicolaKMM 6050T are C15:0, anteiso-C15:0, iso-C15:0, iso-C16:0, iso-C16:1, and iso-C16:0 3-OH, iso-C17:0 3-OH and summed feature 3 (iso-C15:0 2-OH and/or C16:1 ω7c) [2]. Major fatty acids of G. portivictoriae UST040801-001T are almost identical with the exception that C15:0 was not identified but iso-C15:0 3-OH, iso-C17:1 ω9c [1]. The major polar lipids of strains DSM 19838T and DSM 23547T are phosphatidylethanolamine, five unidentified lipids (L1 – L2, L4 – L6) and two unidentified aminolipids (AL1 – AL2). One unidentified aminolipid (AL3) and three unidentified lipids (L2, L7 – L8) appeared as minor components (Fig. 3). As mentioned in the description of the genus Gramella, the major respiratory quinone in both strains is menachinone-6 whereas flexirubin-type pigments were not observed, only non-diffusible carotenoid pigments [2]. The DNA G + C content of the type strains was previously determined as 39.6 mol% of G. echinicolaKMM 6050T and 39.9 mol% of G. portivictoriae UST040801-001T [1, 2].
Fig. 3.
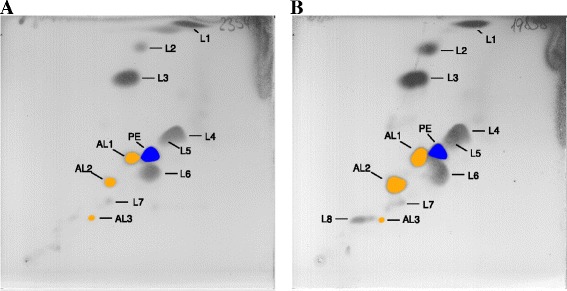
Polar lipids profiles of G. echinicola DSM 19838T and G. portivictoriae DSM 23547T. The polar lipids were extracted using a modified method of Bligh and Dyer [55] (see Tindall [56]) and separated by two-dimensional thin-layer chromatography using the solvents chloroform/methanol/water (65:2:4, by vol.) in the first dimension and chloroform/methanol/acetic acid/water (80:12:15:4, by vol.) in the second dimension at 25 °C, as described by Tindall et al. [21]. For identification of the total polar lipids plates were sprayed with molybdatophosphoric acid (5 % in ethanol) and specific spray reagents used to detect the functional head groups of the lipids, as described by Tindall et al. [21]. PE, phosphatidylethanolamine (blue, phospholipid); AL, amino lipid (yellow, amino lipid); L, polar lipid
Organic matter degradation
Both Gramella strains hydrolyze casein, gelatin, starch and Tweens 20, 40, 60 and 80 as well as esculin ferric citrate, but not agar, chitin or cellulose (CM-cellulose or filter paper) [1, 2, 6]. G. echinicola hydrolyzed DNA [2] whereas G. portivictoriae did not [1]. For strains KCTC 12278T and KCTC 22434T activity of acid phosphatase, alkaline phosphatase, naphthol-AS-BI-phosphohydrolase, esterase (C4), esterase lipase (C8), cystine arylamidase, leucine arylamidase, valine arylamidase and α-glucosidase, β-glucosidase were observed, but not the activity of β-glucuronidase, α-mannosidase, α-fucosidase, lipase (C14) and trypsin [5]. However, Shahina et al. [10] showed the activity of trypsin, α-chymotrypsin, α-glucosidase and N-acetyl-β-glucosaminidase for G. echinicolaKCTC 12278T. Nedashkovskaya et al. [2] showed β-galactosidase activity for G. echinicolaKMM 6050T and Cho et al. [5] showed the α-galactosidase activity for G. echinicolaKMM 12278T. Furthermore, G. portivictoriae UST040801-001T was described with positive α-chymotrypsin, lipase (C14), α-galactosidase, α-glucosidase, β-glucosidase, trypsin and naphthol-AS-BI-phosphohydrolase activity and without N-acetyl-β-glucosaminidase, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, tryptophan deaminase activity [1].
To get further insights into the polysaccharide decomposition potential of G. echinicolaDSM 19838T and G. portivictoriaeDSM 23547T, both strains were incubated in HaHa medium (12 mg/L carbon source mix, [18]) and marine broth (6 g/L carbon source mix, DSMZ medium 514, [20]) supplemented with different polysaccharides, casein and gelatine at 25 °C for up to 14 days (Fig. 4). Each 200 μL well of a microtiter plate was filled with a small portion of one of the AZO-CL-polysaccharides, −casein (Megazym, Bray, Ireland), charcoal-pectin, −gelatin (chapter 15.3.32.3, method 3 in [21]) and 100 μL medium. Each well was inoculated with 100 μL of a starved culture or 100 μL medium as control. Both Gramella type strains hydrolyzed casein and starch but did not hydrolyze chitosan or cellulose (Avicel), as described in previous studies [1, 2, 6], galactomannan, arabinoxylan and hydroxyethyl-cellulose, but not pectin (Fig. 4). Pachyman was hydrolyzed by strain DSM 23547T, whereas galactan and xylan were hydrolyzed by strain DSM 19838T.
Fig. 4.

Polysaccharide hydrolysis by Gramella type strains G. echinicola DSM 19838T, G. portivictoriae DSM 23547T. Both strains were incubated in medium 514 (6 g/L carbon source mix) and HaHa (12 mg/L carbon source mix) for up to 14 days. G. echinicola DSM 19838T was incubated at 25 °C and G. portivictoriae DSM 23547T at 28 °C. Each 200 μL well of a microtiter plate was filled with a small portion of one of the AZO-CL-polysaccharides, −casein (Megazym, Bray, Ireland), charcoal-pectin, −gelatin in 100 μL medium. Each well was inoculated with 100 μL of a starved culture of the strains. The control wells were inoculated with 100 μL medium. The blue colour indicates the release of AZO- monomers and thus hydrolysis of the polysaccharide/peptide. A red-brown colour indicates growth of the strain (mixture of blue and yellow-orange). Black grains in the surrounding of the charcoal-pectin and -gelatine indicate hydrolysis
Genome sequencing information
Genome project history
G. portivictoriaeDSM 23547T and G. echinicolaDSM 19838T were selected for sequencing on the basis of their phylogenetic position [22] and are part of the Genomic Encyclopedia of Type Strains, Phase I: the one thousand microbial genomes project [23], a follow-up of the Genomic Encyclopedia of and Archaea: sequencing a myriad of type strains initiative [24] and the Genomic Standards Consortium project [25], which aim at increasing the number of key reference microbial genomes and to generate a large genomic basis for the discovery of genes encoding novel enzymes [26]. The genome project is deposited in the Genomes OnLine Database [27]. The permanent draft genome sequences are deposited in GenBank. Sequencing, finishing and annotation were performed by the DOE Joint Genome Institute [28]. A summary of the project information is shown in Table 2.
Table 2.
Genome sequencing project information
| MIGS ID | Property | Term | |
|---|---|---|---|
| DSM 19838T | DSM 23547T | ||
| MIGS-31 | Finishing quality | Level 2: Improved High-Quality Draft | Level 2: Improved High-Quality Draft |
| MIGS-28 | Libraries used | Illumina Std shotgun library, 2 × 150bp | Illumina Std shotgun library, 2 × 150bp |
| MIGS-29 | Sequencing platforms | Illumina HiSeq 2000 | Illumina HiSeq 2000 |
| MIGS-31.2 | Fold coverage | 123× | 122× |
| MIGS-30 | Assemblers | Velvet v. 1.1.04, ALLPATHS v. r41043 | Velvet v. 1.1.04, ALLPATHS v. r41043 |
| MIGS-32 | Gene calling method | Prodigal, GenePRIMP, IMG-ER | Prodigal, GenePRIMP, IMG-ER |
| Locus Tag | G530_RS01 | G529_RS01 | |
| NCBI project ID | 16158 | 16157 | |
| Genbank ID | AUHG00000000 | AUHF00000000 | |
| Genbank Date of Release | 2015-08-15 | 2013-12-12 | |
| GOLD ID | Gp0013656 | Gp0013657 | |
| BIOPROJECT | PRJNA185622 | PRJNA185621 | |
| MIGS-13 | Source Material Identifier | DSM 19838 | DSM 23547 |
| Project relevance | Tree of Life, GEBA-KMG | Tree of Life, GEBA-KMG | |
Growth conditions and genomic DNA preparation
Cultures of DSM 23547T and DSM 19838T were grown aerobically in DSMZ medium 514 [20] at 28 °C and 26 °C, respectively. Genomic DNA was isolated using Jetflex Genomic DNA Purification Kit (GENOMED 600100) following the standard protocol provided by the manufacturer but modified by an incubation time of 60 min, the incubation on ice overnight on a shaker, the use of an additional 50 μL proteinase K, and the addition of 200 μL protein precipitation buffer. DNA is available from the DSMZ through the DNA Bank Network [29].
Genome sequencing and assembly
The draft genomes of DSM 19838T and DSM 23547T were generated using the Illumina technology [30]. An Illumina standard shotgun library was constructed and sequenced using the Illumina HiSeq 2000 platform which generated 13,321,360 reads totaling 1,998.2 Mb for strain DSM 19838T and 9,930,650 reads totaling 1,489.6 Mb for strain DSM 23547T (Table 3).
Table 3.
Genome statistics
| DSM 19838T | DSM 23547T | |||
|---|---|---|---|---|
| Attribute | Number | % of Total | Number | % of Total |
| Genome size (bp) | 3,513,826 | 100.0 | 3,269,398 | 100.0 |
| DNA coding (bp) | 3,220,860 | 91.7 | 3,025,367 | 92.5 |
| DNA G + C (bp) | 1,296,572 | 36.9 | 1,292,347 | 39.5 |
| DNA, scaffolds | 18 | 100.0 | 8 | 100.0 |
| Total genes | 3,253 | 100.0 | 3,045 | 100.0 |
| Protein coding genes | 3,199 | 98.3 | 2,984 | 98.0 |
| RNA genes | 54 | 1.7 | 61 | 2.0 |
| Pseudo genes | 21 | 0.7 | 27 | 0.9 |
| Genes in internal clusters | 216 | 6.6 | 174 | 5.7 |
| Genes with function prediction | 2,464 | 75.8 | 2,302 | 75.6 |
| Genes assigned to COGs | 1,863 | 57.3 | 1,747 | 75.6 |
| Genes with Pfam domains | 2,564 | 78.8 | 2,409 | 79.1 |
| Genes with signal peptides | 334 | 10.3 | 347 | 11.4 |
| Genes with transmembrane helices | 766 | 23.6 | 662 | 21.7 |
| CRISPR repeats | 1 | 0.1 | 0 | 0.0 |
All general aspects of library construction and sequencing performed at the JGI can be found at the JGI website [31]. All raw sequence data were passed through DUK, a filtering program developed at JGI, which removes known Illumina sequencing and library preparation artifacts. The following steps were performed for assembly: filtered reads were assembled using Velvet [32], (2) 1–3 Kbp simulated paired end reads were created from Velvet contigs using wgsim [33], (3) sequence reads were assembled with simulated read pairs using Allpaths–LG [34]. Parameters for assembly steps were: (1) Velvet ("velveth 63 -shortPaired" and "velvetg -very clean yes -exportFiltered yes -min contig lgth 500 -scaffolding no -cov cutoff 10"), (2) wgsim ("wgsim -e 0–1 100–2 100 -r 0 -R 0 -X 0") (3) Allpaths–LG ("PrepareAllpathsInputs: PHRED 64 = 1 PLOIDY = 1 FRAG COVERAGE = 125 JUMP COVERAGE = 25 LONG JUMP COV = 50" and "RunAllpathsLG THREADS = 8 RUN = std shredpairs TARGETS = standard VAPI WARN ONLY = OVERWRITE = True").
The final draft assembly contained 18 contigs in a single scaffold for strain DSM 19838T and 11 contigs in two scaffolds for strain DSM 23547T. The total size of the genome of strain DSM 19838T is 3.5 Mbp and the final assembly is based on 430.3 Mbp of data, which provides a 122.6x average coverage of the genome. The total size of the genome of strain DSM 23547T is 3.3 Mbp and the final assembly is based on 396.8 Mbp of data, which provides a 121.5x average coverage of the genome.
Genome annotation
Genes were identified using Prodigal [35] as part of the DOE-JGI genome annotation pipeline [36], followed by manual curation using the JGI GenePRIMP pipeline [37]. The predicted CDSs were translated and used to search the National Center for Biotechnology Information non-redundant database, UniProt, TIGR-Fam, Pfam, PRIAM, KEGG, COG, and InterPro databases. The tRNAScanSE tool [38] was used to find tRNA genes, whereas ribosomal RNA genes were found by searches against models of the ribosomal RNA genes built from SILVA [39]. Other non-coding RNAs such as the RNA components of the protein secretion complex and the RNase P were identified by searching the genome for the corresponding Rfam profiles using INFERNAL [40]. Additional gene prediction analysis and manual functional annotation was performed within the Integrated Microbial Genomes-Expert Review platform [41] developed by the Joint Genome Institute, Walnut Creek, CA, USA [31]. CRISPRs were identified using the online CRIPSRFinder tool [42].
Genome properties
The assemblies of the draft genome sequence of DSM 19838T and DSM 23547T consist of one and two scaffolds amounting to 3,513,826 bp and 3,269,398 bp, respectively (Table 3). The G + C content of DSM 19838T is 36.9 %, which is 2.7 % less than the G + C content reported by Nedashkovskaya et al. [2], and thus shows a difference that surpasses the maximal range among strains belonging to the same species [43]. The G + C content of DSM 23547T is 39.5 % and similar to the G + C content reported by Lau et al. [1]. From the genome of DSM 19838T 3253 genes, 3199 protein-coding genes and 54 RNAs were predicted. From the genome of DSM 23547T 3,045 genes, 2,984 protein-coding genes and 61 RNAs were predicted. The majority of the protein-coding genes (DSM 19838T, 75.8 %; DSM 23547T, 75.6 %) were assigned a putative function while the remaining ones were annotated as hypothetical proteins. The distribution of genes into COGs functional categories is presented in Table 4.
Table 4.
Number of genes associated with the general COG functional categories
| Code | DSM 19838T | DSM 23547T | Description | ||
|---|---|---|---|---|---|
| Value | % age | Value | % age | ||
| J | 188 | 9.2 | 178 | 9.3 | Translation, ribosomal structure and biogenesis |
| A | – | – | – | – | RNA processing and modification |
| K | 108 | 5.3 | 99 | 5.2 | Transcription |
| L | 97 | 4.7 | 88 | 4.6 | Replication, recombination and repair |
| B | 1 | 0.1 | 1 | 0.1 | Chromatin structure and dynamics |
| D | 23 | 1.1 | 22 | 1.2 | Cell cycle control, cell division, chromosome partitioning |
| V | 63 | 3.1 | 54 | 2.8 | Defense mechanisms |
| T | 80 | 3.9 | 70 | 3.7 | Signal transduction mechanisms |
| M | 183 | 8.9 | 168 | 8.8 | Cell wall/membrane biogenesis |
| N | 15 | 0.7 | 19 | 1.0 | Cell motility |
| U | 21 | 1.0 | 19 | 1.0 | Intracellular trafficking and secretion |
| O | 102 | 5.0 | 91 | 4.8 | Posttranslational modification, protein turnover, chaperones |
| C | 101 | 4.9 | 107 | 5.6 | Energy production and conversion |
| G | 115 | 5.6 | 102 | 5.3 | Carbohydrate transport and metabolism |
| E | 182 | 8.9 | 188 | 9.8 | Amino acid transport and metabolism |
| F | 58 | 2.8 | 58 | 3.0 | Nucleotide transport and metabolism |
| H | 127 | 6.2 | 129 | 6.7 | Coenzyme transport and metabolism |
| I | 93 | 4.5 | 91 | 4.8 | Lipid transport and metabolism |
| P | 107 | 5.2 | 104 | 5.4 | Inorganic ion transport and metabolism |
| Q | 51 | 2.5 | 44 | 2.3 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 218 | 10.6 | 189 | 9.9 | General function prediction only |
| S | 114 | 5.6 | 87 | 4.6 | Function unknown |
| X | 5 | 0.2 | 3 | 0.2 | Mobilome: prophages, transposons |
| – | 1,390 | 42.7 | 1,298 | 42.6 | Not in COGs |
Insights from the genome sequence
Comparative genomics
We present a brief comparative genomics analysis of Gramella echinicola and Gramella portivictoriae with a selection of its closest phylogenetic neighbors (according to Fig. 1), 'Gramella forsetii' and Zunongwangia profunda. The genomes of these strains differ significantly in their size with 3.5 Mbp (Gramella echinicola), 3.3 Mbp (Gramella portivictoriae), 3.8 Mbp ('Gramella forsetii') and 5.1 Mbp (Zunongwangia profunda).
An estimate of the overall similarity among these four strains was generated with the Genome-to-Genome Distance Calculator (GGDC 2.0) [44, 45]. It calculates intergenomic distances by comparing two respective genomes to obtain HSPs (high- scoring segment pairs) and, then infers distances via a set of formulae (1, HSP length/total length; 2, identities/HSP length; 3, identities/total length). Formula 2 is robust against the use of incomplete genome sequences and the recommended choice [45]. For convenience the GGDC also reports model-based DDH estimates (digital DDH or dDDH) along with their confidence intervals [45].
The result of this comparison is shown in Table 5 and yields a dDDH value below 22 % throughout, i.e., clearly underlines the expected status of distinct species. With 21.3 % dDDH Gramella echinicola has the highest similarity to 'Gramella forsetii', whereas Gramella portivictoriae has the lowest similarity to Zunongwangia profunda with 18.2 % dDDH. The comparison of Gramella echinicola and Gramella portivictoriae yielded 18.4 % dDDH.
Table 5.
Pairwise comparison of Gramella echinicola and Gramella portivictoriae with 'Gramella forsetii' and Zunongwangia profunda using the GGDC 2.0 (Genome-to-Genome Distance Calculator). Digital DDH (dDDH) and the respective confidence intervals (C.I.) are specified for GGDC’s recommended formula 2
| Strain 1 | Strain 2 | % dDDH | % C.I. |
|---|---|---|---|
| G. echinicola DSM 19838T | ’G. forsetii’ KT0803 | 21.3 | 2.3 |
| ’G. forsetii’ KT0803 | G. portivictoriae DSM 23547T | 18.6 | 2.3 |
| G. echinicola DSM 19838T | G. portivictoriae DSM 23547T | 18.4 | 2.3 |
| ’G. forsetii’ KT0803 | Zunongwangia profunda SM A87T | 20.4 | 2.3 |
| G. echinicola DSM 19838T | Zunongwangia profunda SM A87T | 18.6 | 2.3 |
| G. portivictoriae DSM 23547T | Zunongwangia profunda SM A87T | 18.2 | 2.3 |
Gliding motility
As given in the description of the genus, all Gramella are motile by gliding [2]. We identified all of the genes in the genomes of both type strains that are essential for gliding- motility (Table 6). Furthermore, we observed different modes of gliding-motility on marine soft agar (medium 514 with 0.3 % agar) for both strains. Interestingly, the observed modes of gliding-motility corroborate the observed cellular morphologies (Fig. 2). G. echinicolaDSM 19838T moved by gliding with smooth and entire translucent margins and produced extracellular polymeric substances. In contrast, G. portivictoriaeDSM 23547T formed micro-colonies surrounding the original colony and produced appendages at the cell surface (Fig. 2).
Table 6.
Gliding motility-related genes in strain DSM 19838T and DSM 23547T compared to genes in F. johnsoniae studied by McBride and Zhu [72]
| G. echinicola 19838T | G. portivictoriae 23547T | F. johnsoniae ATCC 17061T | ||
|---|---|---|---|---|
| Locus tag prefix | G530_RS01 | G529_RS01 | FJOH_ | |
| Gliding motility | + | + | + | |
| Adhesin-like | ||||
| remA | – | – | 0808 | |
| remB | 04710 | 03110 | 1657 | |
| sprB | 00190 | – | 0979 | |
| ATP-binding cassette transporter | ||||
| gldA | 13745 | 03925 | 1516 | |
| gldF | 00125 | 12395 | 2722 | |
| gldG | 00120 | 12390 | 2721 | |
| Additional proteins | ||||
| gldB a | 05595 | 08905 | 1793 | |
| gldC | 05600 | 08910 | 1794 | |
| gldD a | 03500 | 02145 | 1540 | |
| gldE | 03505 | 02150 | 1539 | |
| gldH a | 01530 | 00125 | 0890 | |
| gldJ a | 05045 | 08395 | 1557 | |
| peptidoprolyl isomerase (Flavobacteriia, protein folding) | ||||
| gldI | 12360 | 06845 | 2369 | |
| Type IX secretion system (secretion of RemA/RemB) | ||||
| gldK a | 14425 | 05780 | 1853 | |
| gldL a | 14430 | 05775 | 1854 | |
| gldM a | 14435 | 05770 | 1855 | |
| gldN a | 14440 | 05765 | 1856, 1857 | |
| sprA a | 04685 | 03085 | 1653 | |
| sprE a | 01675 | 00280 | 1051 | |
| sprT a | 15350 | 04170 | 1466 | |
aessential gliding motility genes after McBride and Zhu [72]
Peptidases
The MEROPS [46] annotation was carried out by searching the sequences against MEROPS 9.10 (access date: 2014.10.16, version: pepunit.lib) as described by Hahnke et al. [15]. G. echinicolaDSM 19838T processes 161 peptidases the majority of which were 68 metallo (M) and 62 serine (S) peptidases (Table 7 and Table S1 in Additional file 1). Furthermore, the genome contained 17 simple peptidase inhibitors (Table 7 and Table S2 in Additional file 1). G. portivictoriaeDSM 23547T processes 181 peptidases the majority of which were 81 metallo (M) and 72 serine (S) peptidases (Table 7 and Table S3 in Additional file 1). The genome contained 21 simple peptidase inhibitors (Table 7 and Table S4 in Additional file 1).
Table 7.
Peptidases and simple peptidase inhibitors in the genome of strains DSM 19838T and DSM 23547T
| Peptidase | Number of genes | |
|---|---|---|
| family | DSM 19838T | DSM 23547T |
| M01 | 5 | 4 |
| M03 | 2 | 2 |
| M12 | 2 | 2 |
| M13 | 1 | 1 |
| M14 | 6 | 7 |
| M15 | 1 | 1 |
| M16 | 6 | 5 |
| M19 | 1 | 1 |
| M20 | 6 | 5 |
| M23 | 8 | 10 |
| M24 | 4 | 4 |
| M28 | 6 | 5 |
| M38 | 12 | 6 |
| M41 | 1 | 1 |
| M42 | 1 | 1 |
| M43 | 2 | 1 |
| M48 | 2 | 3 |
| M49 | 2 | 0 |
| M50 | 1 | 1 |
| M56 | 3 | 1 |
| M57 | 1 | 1 |
| M61 | 2 | 2 |
| M75 | 1 | 1 |
| M79 | 3 | 1 |
| M97 | 2 | 2 |
| A08 | 1 | 1 |
| A28 | 1 | 1 |
| S01 | 1 | 2 |
| S06 | 0 | 1 |
| S08 | 2 | 3 |
| S09 | 22 | 19 |
| S10 | 1 | 1 |
| S12 | 9 | 4 |
| S13 | 1 | 1 |
| S14 | 2 | 2 |
| S15 | 1 | 0 |
| S16 | 3 | 3 |
| S24 | 1 | 2 |
| S26 | 1 | 1 |
| S33 | 15 | 13 |
| S41 | 6 | 4 |
| S41 | 6 | 4 |
| S51 | 1 | 1 |
| S54 | 4 | 4 |
| S66 | 1 | 1 |
| N11 | 0 | 1 |
| C01 | 1 | 0 |
| C26 | 6 | 6 |
| C40 | 4 | 4 |
| C44 | 5 | 5 |
| C45 | 1 | 1 |
| C56 | 4 | 4 |
| C82 | 1 | 1 |
| T02 | 2 | 2 |
| T03 | 0 | 1 |
| U32 | 2 | 2 |
| I4 | 1 | 1 |
| I39 | 18 | 15 |
| I43 | 1 | 0 |
| I87 | 1 | 1 |
Carbohydrate active enzymes
G. echinicolaDSM 19838T and G. portivictoriaeDSM 23547T harboured a large set of 127 and 119 CAZymes, respectively, comprising 37–39 glycoside hydrolases, 2–5 polysaccharide lyases, 9–14 carbohydrate esterases, 9–10 carbohydrate binding modules and 55–61 glycoside transferases (Table 8 and Table S5 and S6 in Additional file 1).
Table 8.
Carbohydrate active enzymes (CAZy) in the genome of strains DSM 19838T and DSM 23547T
| CAZy | Number of genes | |
|---|---|---|
| family | DSM 19838T | DSM 23547T |
| GH2 | 1 | 2 |
| GH3 | 4 | 3 |
| GH5 | 1 | 3 |
| GH9 | 0 | 1 |
| GH13 | 6 | 2 |
| GH15 | 1 | 1 |
| GH16 | 5 | 3 |
| GH17 | 1 | 1 |
| GH20 | 1 | 0 |
| GH23 | 2 | 2 |
| GH26 | 1 | 2 |
| GH27 | 0 | 1 |
| GH28 | 1 | 0 |
| GH29 | 1 | 0 |
| GH31 | 1 | 1 |
| GH32 | 1 | 4 |
| GH37 | 1 | 0 |
| GH43 | 2 | 1 |
| GH63 | 0 | 1 |
| GH65 | 0 | 1 |
| GH73 | 1 | 1 |
| GH88 | 1 | 0 |
| GH97 | 1 | 1 |
| GH105 | 1 | 0 |
| GH113 | 1 | 1 |
| GH130 | 0 | 1 |
| GHa | 1 | 3 |
| CE1 | 1 | 0 |
| CE4 | 3 | 1 |
| CE8 | 1 | 0 |
| CE11 | 1 | 1 |
| CE12 | 1 | 0 |
| CE14 | 3 | 2 |
| CEa | 4 | 2 |
| PL6 | 0 | 1 |
| PL7 | 0 | 2 |
| PL9 | 1 | 0 |
| PL12 | 0 | 1 |
| PL17 | 0 | 1 |
| CBM38 | 0 | 1 |
| CBM48 | 2 | 2 |
| CBM50 | 4 | 4 |
| CBM57 | 2 | 1 |
| CBMa | 2 | 1 |
| GT2 | 29 | 26 |
| GT4 | 18 | 16 |
| GT5 | 2 | 1 |
| GT8 | 0 | 1 |
| GT9 | 2 | 1 |
| GT10 | 2 | 0 |
| GT19 | 1 | 1 |
| GT20 | 1 | 1 |
| GT28 | 1 | 1 |
| GT30 | 1 | 1 |
| GT51 | 3 | 3 |
| GT83 | 1 | 1 |
| GTa | 0 | 2 |
| AA1 | 1 | 0 |
| AA6 | 0 | 1 |
| AA12 | 1 | 0 |
| AAa | 0 | 2 |
agenes attributed to an enzyme class, but not to a family
Polysaccharide utilization loci
Kabisch et al. [13] investigated ’G. forsetii’ KT0803 for its ability to decompose laminarin-like, α-1,4-linked-glucose and alginate-like polysaccharides. The two PULs involved in either the decomposition of laminarin-like polysaccharides or α-1,4-linked glucose-polymers (glycogen, starch and amylose) were as well found in G. portivictoriaeDSM 23547T and G. echinicolaDSM 19838T (Figure S1, Figure S2 in Additional file 2). Both PULs were greatly conserved among other closely related genera (see Fig. 1) and within the Flavobacteriaceae. The PUL involved in the decomposition of alginate-like polysaccharides was found in G. portivictoriaeDSM 23547T, but not in G. echinicolaDSM 19838T (Figure S3 and Figure S4 in Additional file 2). This PUL was not conserved among other closely related genera, but greatly distributed within the Flavobacteriaceae. Interestingly, the PULs of the Salegentibacter and Aquimarina were highly syntenic with those of Gramella, whereas the PULs of Gillisia, Mesonia, Zunongwangia, Psychroflexus, Salinimicrobium and Dokdonia had additional genes. One PUL that potentially encodes for the decomposition of sulfated β-d-glucosides (Figure S5 in Additional file 2) and one for the decomposition of β-d-fructans (levans) (Figure S6 in Additional file 2) was found in all three Gramella and in other closely related Flavobacteriaceae. A PUL that was found only in G. echinicolaDSM 19838T comprised pectin-like polysaccharide decomposing CAZymes and genes of the pectate degradation pathway (Fig. 5, Figure S7 in Additional file 2). A similar set of genes was found in a PUL of Flavobacterium johnsoniae UW101T, which was hypothesized to be involved in pectin decomposition [16].
Fig. 5.
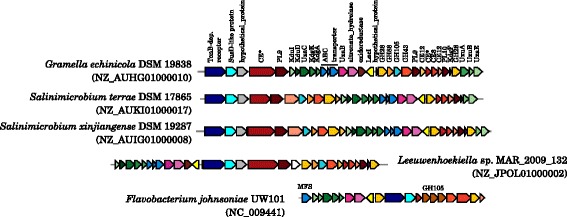
A pectin-like PUL of G. echinicola DSM 19838T and other Flavobacteriaceae. A similar PUL was identified in Flavobacterium johnsoniae UW101T by McBride et al. [16]. Locus tags are given below both the first and last gene of the loci. Accession numbers in brackets are GenBank accession numbers of the corresponding contig. Investigation of syntenic loci was done using MultiGeneBlast [57]. A description of glycoside hydrolase (GH), polysaccharide lyase (PL) and carbohydrate esterase (CE) families can be seen at the CAZy homepage [58, 59]. The pectin-like polysaccharide decomposition pathway, encoded by these genes, is shown in Figure S6 in the Additional file 2. SusD, SusD-like protein; LacI, LacI family transcriptional regulator; MFS, major facilitator superfamily transporter; KduD, 2-keto-3-deoxy-d-gluconate-dehydrogenase; UxaB, altronate oxidoreductase; UxaC, glucuronate isomerase; KdgA, 2-keto-3-deoxygluconate-6-phosphate aldolase; KdgF, pectin degradation protein; KduI, 5-dehydro-4-deoxy- d-glucuronate isomerase; KdgK, 2-dehydro-3-deoxygluconokinase; UxuA, mannonate dehydratase; UxuB, d-mannonate oxidoreductase; UxaE, d-tagaturonate epimerase
Surprisingly, we found a PUL in G. portivictoriaeDSM 23547T, Salinimicrobium terraeDSM 17865T and some other Flavobacteriaceae (Fig. 6) comprising typical cellulases/hemicellulases, such as GH5 (cellulase family A), GH9 (cellulase family E) and GH26 (cellulase family I). However, Salinimicrobium terraeDSM 17865T was described to be unable to hydrolyze carboxymethyl-cellulose and filter paper. Lau et al. [1] showed β-glucosidase activity by G. portivictoriaeDSM 23547T, but no decomposition of carboxymethyl-cellulose. The authors tested cellulose decomposition using a 0.5 % CMC overlay agar as described by McCammon et al. [47]. As mentioned above, we could show that G. portivictoriaeDSM 23547T is able to hydrolyze hydroxyethyl-cellulose, but not Avicel-cellulose. Thus we additionally tested this strain for the decomposition of AZO-CL carboxymethyl-cellulose, Whatman filter No. 1 cellulose and cellulose of cigarette paper. In HaHa medium and marine broth strain DSM 23547T hydrolyzed AZO-CL carboxymethyl-cellulose, but not the Whatman filter.
Fig. 6.
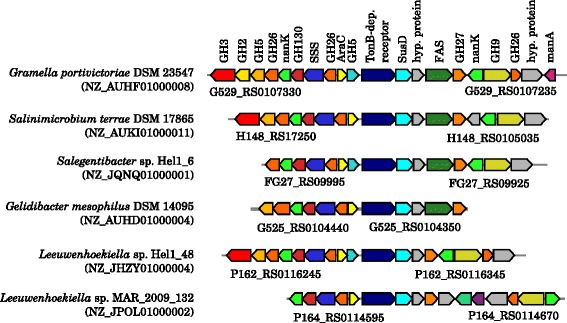
A cellulose/hemicellulose-like PUL of G. portivictoriae DSM 23547T and other Flavobacteriaceae. Locus tags are given below both the first and last gene of the loci. Accession numbers in brackets are GenBank accession numbers of the corresponding contig. Investigation of syntenic loci was done using MultiGeneBlast [57]. A description of glycoside hydrolase (GH), polysaccharide lyase (PL) and carbohydrate esterase (CE) families can be seen at the CAZy homepage [58, 59]. SusD, SusD-like protein; AraC, AraC family transcriptional regulator; manA, Man-6-P isomerase; nanK, GlcNAc-2-epimerase; FAS, FAS1 domain protein; SSS, sodium:solute symporter
Conclusion
All three of the genome-sequenced Gramella spp. sequenced to date were isolated from marine habitats, Gramella echinicolaDSM 19838T was isolated from a sea urchin, G. portivictoriaeDSM 23547T from the sediment and ’G. forsetii’ KT0803 from surface seawater. In contrast to ’G. forsetii’ (48.7 peptidases Mbp−1) [14, 48], both G. echinicolaDSM 19838T and G. portivictoriae DSM 23547T have a greater number peptidases, 68 Mbp−1 and 81 Mbp−1, respectively. The observed dominance of metallo (M), serine (S) and cysteine (C) peptidase families was already reported by Xing and Hahnke et al. [14] and seems to be a general feature among Flavobacteriaceae. Interestingly, while both G. echinicolaDSM 19838T and G. portivictoriae DSM 23547T have a similar amount of CAZymes (119 and 127), CAZymes Mbp−1 (36.1 and 36.4) and CAZy families (44 and 45), the genome of ’G. forsetii’ comprised a larger amount of CAZymes (164 overall and 43.2 Mbp−1) and a greater diversity of CAZy families (54) [13, 14]. We observed different polysaccharide decomposition capabilities among the Gramella which might be linked to the nutrient composition of the habitats they were isolated from. Whether the laminarin-like and the starch/amylose-like PUL is a common feature of Gramella needs to be assessed once further Gramella genomes are available. Furthermore, the link between the coincidence of the observed gliding-motility modes, the cellular morphologies and certain environmental conditions has to be investigated in detail. For example, Gramella oceani and Muricauda ruestringensis, both producing appendages, were isolated from marine intertidal sediment [6, 49]. Bruns et al. [49] and Hahnke et al. [50] assumed that such appendages are connections between the cells or serve as anchor to mediate surface attachment and particle formation.
Taxonomic and nomenclatural proposals
Based on the new morphological (gliding, EPS, appendages), physiological (polysaccharide hydrolysis) and genomic observations (DNA G + C content, CAZymes, PUL, peptidases) we propose the emendation of Gramella echinicolaDSM 19838T Nedashkovskaya et al. [2] emend. Shahina et al. [10] and the emendation of Gramella portivictoriae Lau et al. [5].
Emended description of Gramella echinicola Nedashkovskaya et al. [2] emend. Shahina et al. [10]
The description of Gramella echinicola is as given by Nedashkovskaya et al. [2] and Shahina et al. [10], with the following emendations. The major polar lipids are phosphatidylethanolamine, together with a number of unidentified lipids, that included seven polar lipids that did not stain with any of the specific spray reagents (L1 – L8) and two amino lipids (AL1 – AL3) that together with their specific Rf values, that can be deduced from Fig. 3 and their staining behavior, may serve as reference points for future work where chromatographic conditions are the same. The G + C content is 36.9 %. Production of extracellular polymeric substances. Hydrolyses aesculin, galactomannan, arabinoxylan, galactan, xylan and hydroxyethyl-cellulose, but not Avicel-cellulose, pectin and chitosan.
Emended description of Gramella portivictoriae Lau et al. [1]
The description of Gramella portivictoriae is as given by Lau et al. [1], with the following emendations. The major polar lipids are phosphatidylethanolamine, together with a number of unidentified lipids, that included seven polar lipids that did not stain with any of the specific spray reagents (L1 – L7) and two amino lipids (AL1 – AL3) that together with their specific Rf values, that can be deduced from Fig. 3 and their staining behavior, may serve as reference points for future work where chromatographic conditions are the same. Appendages at the cell surface. Hydrolyses aesculin, galactomannan, arabinoxylan, pachyman and hydroxyethyl-cellulose, but not Avicel-cellulose, pectin and chitosan.
Acknowledgements
The authors gratefully acknowledge the help of Andrea Schütze, DSMZ, for growing cells of DSM 19838T and of Susanne Schneider, DSMZ, for growing the cells of DSM 23547T, Evelyne Brambilla, DSMZ, for DNA extraction and quality control, and Anja Frühling for polar lipid extraction and thin-layer chromatography. This work was performed under the auspices of the US Department of Energy’s Office of Science, Biological and Environmental Research Program, and by the University of California, Lawrence Berkeley National Laboratory under contract No. DE-AC02-05CH11231, Lawrence Livermore National Laboratory under Contract No. DE-AC52-07NA27344.
Funding
AL was supported by the St. Petersburg State University grant (No 1.38.253.2015). RLH and IP were supported by the Bundesministerium für Ernährung und Landwirtschaft No. 22016812 (PI: Brian J. Tindall). The publication of this article was funded by the Open Access fund of the Leibniz Association.
Authors’ contributions
IP, RLH, MG, HPK and NCK designed research and project outline. SH performed CAZy and MEROPS analysis. JPMK and RLA performed comparative genomics. IP and RLH investigated gliding motility, CAZymes and PUL. MR performed electron microscopy. RLH, SV and BTI investigated the polar lipids. IP, RLH, JPMK and BJT drafted the manuscript that was critically reviewed and polished by RLH, JPMK, BTI, MG and HPK. AL, JH, ST, MH, TBKR, MH, AP, NNI, KM, VM and TW performed genome sequencing, assembly and annotation. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- AZO-CL
Azurine-crosslinked
- CAZy
Carbohydrate active enzymes
- EPS
Extracellular polymeric substances
- PUL
Polysaccharide utilization loci
Additional files
Peptidases or homologues in the genome of Gramella echinicola DSM 19838T. Table S2. Simple peptidases inhibitors in the genome of Gramella echinicola DSM 19838T. Table S3. Peptidases or homologues in the genome of Gramella portivictoriae DSM 23547T. Table S4. Simple peptidases inhibitors in the genome of Gramella portivictoriae DSM 23547T. Table S5. Carbohydrate active enzymes (CAZymes) in the genome of Gramella echinicola DSM 19838T. Table S6. Carbohydrate active enzymes (CAZymes) in the genome of Gramella portivictoriae DSM 23547T. (PDF 261 kb)
The laminarin-like PUL. Figure S2. The 1,4-linked glucose-polymer-like PUL. Figure S3. The alginate-like PUL. Figure S4. Part of the alginate-like polysaccharide decom- position pathway. Figure S5. The sulfated β-d-glucoside PUL. Figure S6. The two combined β-d-fructans PUL. Figure S7. Part of the pectin-like polysaccharide decomposition pathway. (PDF 642 kb)
References
- 1.Lau SCK, Tsoi MMY, Li X, Plakhotnikova I, Dobretsov S, Wong PK, Qian PY. Gramella portivictoriae sp. nov., a novel member of the family Flavobacteriaceae isolated from marine sediment. Int J Syst Evol Microbiol. 2005;55:2497–500. doi: 10.1099/ijs.0.63824-0. [DOI] [PubMed] [Google Scholar]
- 2.Nedashkovskaya OI, Kim SB, Lysenko AM, Frolova GM, Mikhailov VV, Bae KS, Lee DH, Kim IS. Gramella echinicola gen. nov., sp. nov., a novel halophilic bacterium of the family Flavobacteriaceae isolated from the sea urchin Strongylocentrotus intermedius. Int J Syst Evol Microbiol. 2005;55:391–4. doi: 10.1099/ijs.0.63314-0. [DOI] [PubMed] [Google Scholar]
- 3.Bernardet JF. Family I. Flavobacteriaceae Reichenbach 1992b, 327VP (Effective publication: Reichenbach 1989b, 2013.) emend. Bernardet, Segers, Vancanneyt, Berthe, Kersters and Vandamme 1996, 145 emend. Bernardet, Nakagawa and Holmes 2002, 1057. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al., editors. Bergey’s Manual of Systematic Bacteriology. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, vol. 4. 2nd ed. New York: Springer; 2011. p. 106–314.
- 4.Reichenbach H. Reichenbach H. Order 1. Cytophagales Leadbetter 1974, 99AL. In: Staley JT, Bryant MP, Pfennig N, Holt JT, editors. Bergey’s Manual of Systematic Bacteriology. New York: Springer; 1989. pp. 2011–3. [Google Scholar]
- 5.Cho SH, Chae SH, Cho M, Kim TU, Choi S, Han JH, Kim YT, Joung Y, Joh K, Nedashkovskaya OI, Kim SB. Gramella gaetbulicola sp. nov., a member of the family Flavobacteriaceae isolated from foreshore soil. Int J Syst Evol Microbiol. 2011;61:2654–8. doi: 10.1099/ijs.0.027599-0. [DOI] [PubMed] [Google Scholar]
- 6.Hameed A, Shahina M, Lin S-Y, Liu Y-C, Lai W-A, Young C-C. Gramella oceani sp. nov., a zeaxanthin-producing bacterium of the family Flavobacteriaceae isolated from marine sediment. Int J Syst Evol Microbiol. 2014;64:2675–81. doi: 10.1099/ijs.0.059881-0. [DOI] [PubMed] [Google Scholar]
- 7.Jeong SH, Jin HM, Jeon CO. Gramella aestuarii sp. nov., isolated from a tidal flat, and emended description of Gramella echinicola. Int J Syst Evol Microbiol. 2013;63:2872–8. doi: 10.1099/ijs.0.048694-0. [DOI] [PubMed] [Google Scholar]
- 8.Park J-M, Park S, Won S-M, Jung Y-T, Shin K-S, Yoon J-H. Gramella aestuariivivens sp. nov., isolated from a tidal flat. Int J Syst Evol Microbiol. 2015;65:1262–7. doi: 10.1099/ijs.0.000093. [DOI] [PubMed] [Google Scholar]
- 9.Liu K, Li S, Jiao N, Tang K. Gramella flava sp. nov., a member of the family Flavobacteriaceae isolated from seawater. Int J Syst Evol Microbiol. 2013;64:165–8. doi: 10.1099/ijs.0.051987-0. [DOI] [PubMed] [Google Scholar]
- 10.Shahina M, Hameed A, Lin SY, Lee RJ, Lee MR, Young CC. Gramella planctonica sp. nov., a zeaxanthin-producing bacterium isolated from surface seawater, and emended descriptions of Gramella aestuarii and Gramella echinicola. Antonie van Leeuwenhoek. Int J Gen Mol Microbiol. 2014;105:771–9. doi: 10.1007/s10482-014-0133-4. [DOI] [PubMed] [Google Scholar]
- 11.Thomas F, Barbeyron T, Tonon T, Génicot S, Czjzek M, Michel G. Characterization of the first alginolytic operons in a marine bacterium: From their emergence in marine Flavobacteriia to their independent transfers to marine Proteobacteria and human gut Bacteroides. Environ Microbiol. 2012;14:2379–94. doi: 10.1111/j.1462-2920.2012.02751.x. [DOI] [PubMed] [Google Scholar]
- 12.Mann AJ, Hahnke RL, Huang S, Werner J, Xing P, Barbeyron T, Huettel B, Stüber K, Reinhardt R, Harder J, Glöckner FO, Amann RI, Teeling H. The genome of the alga-associated marine flavobacterium Formosa agariphila KMM 3901T reveals a broad potential for degradation of algal polysaccharides. Appl Environ Microbiol. 2013;79:6813–22. doi: 10.1128/AEM.01937-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kabisch A, Otto A, König S, Becher D, Albrecht D, Schüler M, Teeling H, Amann RI, Schweder T. Functional characterization of polysaccharide utilization loci in the marine Bacteroidetes ‘Gramella forsetii’ KT0803. ISME J. 2014;8:1492–502. doi: 10.1038/ismej.2014.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Xing P, Hahnke RL, Unfried F, Markert S, Huang S, Fuchs BM, Barbeyron T, Harder J, Schweder T, Glöckner FO, Amann RI, Teeling H. Niche separation of two Polaribacter strains isolated from the German Bight of the North Sea during a spring diatom bloom. ISME J. 2014;9:1410–22. doi: 10.1038/ismej.2014.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hahnke RL, Stackebrandt E, Meier-Kolthoff JP, Tindall BJ, Huang S, Rohde M, Lapidus A, Han J, Trong S, Haynes M, Reddy TBK, Huntemann M, Pati A, Ivanova NN, Mavromatis K, Markowitz V, Woyke T, Göker M, Kyrpides NC, Klenk H-P. High quality draft genome sequence of Flavobacterium rivuli type strain WB 3.3-2T (DSM 21788T), a valuable source of polysaccharide decomposing enzymes. Stand Genomic Sci. 2015;10:1–16. doi: 10.1186/s40793-015-0032-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McBride MJ, Xie G, Martens EC, Lapidus A, Henrissat B, Rhodes RG, Goltsman E, Wang W, Xu J, Hunnicutt DW, Staroscik AM, Hoover TR, Cheng YQ, Stein JL. Novel features of the polysaccharide-digesting gliding bacterium Flavobacterium johnsoniae as revealed by genome sequence analysis. Appl Env Microbiol. 2009;75:6864–75. doi: 10.1128/AEM.01495-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Du H, Jiao N, Hu Y, Zeng Y. Diversity and distribution of pigmented heterotrophic bacteria in marine environments. FEMS Microbiol Ecol. 2006;57:92–105. doi: 10.1111/j.1574-6941.2006.00090.x. [DOI] [PubMed] [Google Scholar]
- 18.Hahnke RL, Harder J. Phylogenetic diversity of Flavobacteria isolated from the North Sea on solid media. Syst Appl Microbiol. 2013;36:497–504. doi: 10.1016/j.syapm.2013.06.006. [DOI] [PubMed] [Google Scholar]
- 19.Gómez-Pereira PR, Fuchs BM, Alonso C, Oliver MJ, van Beusekom JEE, Amann R. Distinct flavobacterial communities in contrasting water masses of the North Atlantic Ocean. ISME J. 2010;4:472–87. doi: 10.1038/ismej.2009.142. [DOI] [PubMed] [Google Scholar]
- 20.List of growth media used at the DSMZ. http://www.dsmz.de/.
- 21.Tindall BJ, Sikorski J, Smibert RA, Krieg NR. Chapter 15 : Phenotypic Characterization and the Principles of Comparative Systematics. In: Reddy CA, Beveridge TJ, Breznak JA, Marzluf GA, Schmidt TM, Snyder LR, editors. Methods for General and Molecular Microbiology. 3rd ed. Washington, DC, USA: American Society of Microbiology; 2007. p. 330–93.
- 22.Klenk HP, Göker M. En route to a genome-based classification of Archaea and Bacteria? Syst Appl Microbiol. 2010;33:175–82. doi: 10.1016/j.syapm.2010.03.003. [DOI] [PubMed] [Google Scholar]
- 23.Kyrpides NC, Hugenholtz P, Eisen JA, Woyke T, Göker M, Parker CT, Amann R, Beck BJ, Klenk H-P, et al. Genomic Encyclopedia of Bacteria and Archaea: sequencing a myriad of type strains. PLoS Biol. 2014;12:e1001920. doi: 10.1371/journal.pbio.1001920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wu D, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Ivanova NN, Kunin V, Goodwin L, Wu M, Tindall BJ, Hooper SD, Pati A, Lykidis A, Spring S, Anderson IJ, D’haeseleer P, Zemla A, Singer M, Lapidus A, Nolan M, Copeland A, Han C, Chen F, Cheng J-F, Lucas S, Kerfeld C, Lang E, Gronow S, Chain P, Bruce D, Rubin EM, Kyrpides NC, Klenk H-P, Eisen JA. A phylogeny-driven genomic encyclopaedia of Bacteria and Archaea. Nature. 2009;462:1056–60. doi: 10.1038/nature08656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Field D, Amaral-Zettler L, Cochrane G, Cole JR, Dawyndt P, Garrity GM, Gilbert J, Glöckner FO, Hirschman L, Karsch-Mizrachi I, Klenk HP, Knight R, Kottmann R, Kyrpides N, Meyer F, Gil IS, Sansone SA, Schriml LM, Sterk P, Tatusova T, Ussery DW, White O, Wooley J. The Genomic Standards Consortium. PLoS Biol. 2011;9:8–10. doi: 10.1371/journal.pbio.1001088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Piao H, Froula J, Du C, Kim TW, Hawley ER, Bauer S, Wang Z, Ivanova N, Clark DS, Klenk HP, Hess M. Identification of novel biomass-degrading enzymes from genomic dark matter: Populating genomic sequence space with functional annotation. Biotechnol Bioeng. 2014;111:1550–65. doi: 10.1002/bit.25250. [DOI] [PubMed] [Google Scholar]
- 27.Pagani I, Liolios K, Jansson J, Chen IMA, Smirnova T, Nosrat B, Markowitz VM, Kyrpides NC. The Genomes OnLine Database (GOLD) v.4: Status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res. 2012;40:D571–9. doi: 10.1093/nar/gkr1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mavromatis K, Land ML, Brettin TS, Quest DJ, Copeland A, Clum A, Goodwin L, Woyke T, Lapidus A, Klenk HP, Cottingham RW, Kyrpides NC. The fast changing landscape of sequencing technologies and their impact on microbial genome assemblies and annotation. PLoS One. 2012;7:e48837. doi: 10.1371/journal.pone.0048837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gemeinholzer B, Dröge G, Zetzsche H, Haszprunar G, Klenk H-P, Güntsch A, Berendsohn WG, Wägele J-W. The DNA Bank Network: The start from a German initiative. Biopreserv Biobank. 2011;9:51–5. doi: 10.1089/bio.2010.0029. [DOI] [PubMed] [Google Scholar]
- 30.Bennett S. Solexa Ltd. Pharmacogenomics. 2004;5:433–8. doi: 10.1517/14622416.5.4.433. [DOI] [PubMed] [Google Scholar]
- 31.DOE Joint Genome Institute. A DOE Office of Science user Facility of Lawrence Berkeley National Laboratory. DOE Jt Genome Inst.:http://jgi.doe.gov.
- 32.Zerbino DR, Birney E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18:821–9. doi: 10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li H. wgsim. 2011. https://github.com/lh3/wgsim.
- 34.Gnerre S, Maccallum I, Przybylski D, Ribeiro FJ, Burton JN, Walker BJ, Sharpe T, Hall G, Shea TP, Sykes S, Berlin AM, Aird D, Costello M, Daza R, Williams L, Nicol R, Gnirke A, Nusbaum C, Lander ES, Jaffe DB. High-quality draft assemblies of mammalian genomes from massively parallel sequence data. Proc Natl Acad Sci U S A. 2011;108:1513–8. doi: 10.1073/pnas.1017351108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hyatt D, Chen G-L, Locascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010;11:119. doi: 10.1186/1471-2105-11-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Huntemann M, Ivanova NN, Mavromatis K, Tripp HJ, Paez-Espino D, Palaniappan K, Szeto E, Pillay M, Chen I-MA, Pati A, Markowitz VM, Kyrpides NC. The Standard Operating Procedure of the DOE-JGI Microbial Genome Annotation Pipeline (MGAP v.4) Keywords. Stand Genomic Sci. 2015:1–6. [DOI] [PMC free article] [PubMed]
- 37.Pati A, Ivanova NN, Mikhailova N, Ovchinnikova G, Hooper SD, Lykidis A, Kyrpides NC. GenePRIMP: a gene prediction improvement pipeline for prokaryotic genomes. Nat Methods. 2010;7:455–7. doi: 10.1038/nmeth.1457. [DOI] [PubMed] [Google Scholar]
- 38.Lowe TM, Eddy SR. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997;25:955–64. doi: 10.1093/nar/25.5.0955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig WG, Peplies J, Glöckner FO. SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007;35:7188–96. doi: 10.1093/nar/gkm864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.INFERNAL. Inference of RNA alignments. 2014. http://infernal.janelia.org.
- 41.Markowitz VM, Mavromatis K, Ivanova NN, Chen IMA, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics. 2009;25:2271–8. doi: 10.1093/bioinformatics/btp393. [DOI] [PubMed] [Google Scholar]
- 42.Grissa I, Vergnaud G, Pourcel C. CRISPRFinder: a web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007;35:W52–7. doi: 10.1093/nar/gkm360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Meier-Kolthoff JP, Klenk H-P, Göker M. Taxonomic use of DNA G + C content and DNA-DNA hybridization in the genomic age. Int J Syst Evol Microbiol. 2014;64:352–6. doi: 10.1099/ijs.0.056994-0. [DOI] [PubMed] [Google Scholar]
- 44.Auch AF, Klenk H-P, Göker M. Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Stand Genomic Sci. 2010;2:142–8. doi: 10.4056/sigs.541628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Meier-Kolthoff JP, Auch AF, Klenk H-P, Göker M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics. 2013;14:60. doi: 10.1186/1471-2105-14-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rawlings ND, Waller M, Barrett AJ, Bateman A. MEROPS: the database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014;42:D503–9. doi: 10.1093/nar/gkt953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McCammon A, Innest BH, Bowman JP, Franzmann PD, Dobson SJ, Holloway PE, Skerratt JH, Nichols PD, Rankin LM. Flavobacterium hibernum sp. nov., a lactose-utilizing bacterium from a freshwater Antarctic lake. 2015. [DOI] [PubMed] [Google Scholar]
- 48.Fernández-Gómez B, Richter M, Schüler M, Pinhassi J, Acinas SG, González JM, Pedrós-Alió C. Ecology of marine bacteroidetes: a comparative genomics approach. ISME J. 2013;7:1026–37. doi: 10.1038/ismej.2012.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bruns A, Rohde M, Berthe-Corti L. Muricauda ruestringensis gen. nov., sp. nov., a facultatively anaerobic, appendaged bacterium from German North Sea intertidal sediment. Int J Syst Evol Microbiol. 2001;51:1997–2006. doi: 10.1099/00207713-51-6-1997. [DOI] [PubMed] [Google Scholar]
- 50.Hahnke RL, Bennke CM, Fuchs BM, Mann AJ, Rhiel E, Teeling H, Amann R, Harder J. Dilution cultivation of marine heterotrophic bacteria abundant after a spring phytoplankton bloom in the North Sea. Environ Microbiol. 2015;17:3515–26. doi: 10.1111/1462-2920.12479. [DOI] [PubMed] [Google Scholar]
- 51.Göker M, Cleland D, Saunders E, Lapidus A, Nolan M, Lucas S, Hammon N, Deshpande S, Cheng J-F, Tapia R, Han C, Goodwin L, Pitluck S, Liolios K, Pagani I, Ivanova N, Mavromatis K, Pati A, Chen A, Palaniappan K, Land M, Hauser L, Chang Y-J, Jeffries CD, Detter JC, Beck B, Woyke T, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Kyrpides NC, Klenk H-P. Complete genome sequence of Isosphaera pallida type strain (IS1BT) Stand Genomic Sci. 2011;4:63–71. doi: 10.4056/sigs.1533840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yarza P, Ludwig W, Euzéby J, Amann R, Schleifer K-H, Glöckner FO, Rosselló-Móra R. Update of the All-Species Living Tree Project based on 16S and 23S rRNA sequence analyses. Syst Appl Microbiol. 2010;33:291–9. doi: 10.1016/j.syapm.2010.08.001. [DOI] [PubMed] [Google Scholar]
- 53.Yarza P, Richter M, Peplies J, Euzeby J, Amann R, Schleifer KH, Ludwig W, Glöckner FO, Rosselló-Móra R. The All-Species Living Tree project: A 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst Appl Microbiol. 2008;31:241–50. doi: 10.1016/j.syapm.2008.07.001. [DOI] [PubMed] [Google Scholar]
- 54.Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, Buchner A, Lai T, Steppi S, Jobb G, Förster W, Brettske I, Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, König A, Liss T, Lüßmann R, May M, Nonhoff B, Reichel B, Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M, Ludwig T, Bode A, Schleifer KH. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–71. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–7. doi: 10.1139/o59-099. [DOI] [PubMed] [Google Scholar]
- 56.Tindall BJ. A comparative study of the lipid composition of Halobacterium saccharovorum from various sources. Syst Appl Microbiol. 1990;13:128–30. doi: 10.1016/S0723-2020(11)80158-X. [DOI] [Google Scholar]
- 57.Medema MH, Takano E, Breitling R. Detecting sequence homology at the gene cluster level with MultiGeneBlast. Mol Biol Evol. 2013;30:1218–23. doi: 10.1093/molbev/mst025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lombard V, Golaconda Ramulu H, Drula E, Coutinho PM, Henrissat B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014;42:D490–5. doi: 10.1093/nar/gkt1178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cantarel BL, Coutinho PM, Rancurel C, Bernard T, Lombard V, Henrissat B. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for glycogenomics. Nucleic Acids Res. 2009;37:D233–8. doi: 10.1093/nar/gkn663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Thomson N, Wipat A, et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26:541–7. doi: 10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Euzéby JP. List of bacterial names with standing in nomenclature: A folder available on the Internet. Int J Syst Bacteriol. 1997;47:590–2. doi: 10.1099/00207713-47-2-590. [DOI] [PubMed] [Google Scholar]
- 62.Parte AC. LPSN–list of prokaryotic names with standing in nomenclature. Nucleic Acids Res. 2014;42:D613–6. doi: 10.1093/nar/gkt1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Garrity G. NamesforLife. BrowserTool takes expertise out of the database and puts it right in the browser. Microbiol Today. 2010;37:9. [Google Scholar]
- 64.Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci U S A. 1990;87:4576–9. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Krieg NR, Ludwig W, Euzéby J, Whitman WB. Phylum XIV. Bacteroidetes phyl. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al., editors. Bergey’s Manual of Systematic Bacteriology. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes., vol. 4. 2nd ed. New York: Springer; 2010. p. 25–469.
- 66.Euzeby J. Validation List N° 143. List of new names and new combinations previously effectively, but not validly, published. Int J Syst Evol Microbiol. 1980;62:225–420. doi: 10.1099/ijs.0.2008/000679-0. [DOI] [PubMed] [Google Scholar]
- 67.Bernardet JF. Class II. Flavobacteriia class. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al., editors. Bergey’s Manual of Systematic Bacteriology. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, vol. 4. 2nd ed. New York: Springer; 2010. p. 106–314.
- 68.Skerman V, Mc Gowan V, Sneath P. Validation List N° 145. List of new names and new combinations previously effectively, but not validly, published. Int J Syst Evol Microbiol. 1980;30:225–420. doi: 10.1099/00207713-30-1-225. [DOI] [Google Scholar]
- 69.Bernardet JF. Order I. Flavobacteriales ord. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al., editors. Bergey’s Manual of Systematic Bacteriology. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes, vol. 4. 2nd ed. New York: Springer; 2011.
- 70.BAuA 2010 – 2012 update, Classification of bacteria and archaea in risk groups. http://www.baua.de TRBA 466. :19.
- 71.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000;25:25–9. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McBride MJ, Zhu Y. Gliding motility and Por secretion system genes are widespread among members of the phylum bacteroidetes. J Bacteriol. 2013;195:270–8. doi: 10.1128/JB.01962-12. [DOI] [PMC free article] [PubMed] [Google Scholar]