

Abstract
Nontyphoidal Salmonella (NTS) invasive infections are an important cause of morbidity and mortality in sub-Saharan Africa. Several vaccines are in development to prevent these infections. We describe an NTS opsonophagocytic killing assay that uses HL-60 cells and baby rabbit complement to quantify functional antibodies elicited by candidate NTS vaccines.
TEXT
Invasive nontyphoidal Salmonella (NTS) infections are increasingly being recognized as an important cause of morbidity and mortality in infants and HIV-infected adults and children in sub-Saharan Africa (1). Multiple vaccines are currently in development, including live-attenuated vaccines, conjugate vaccines, and generalized modules for membrane antigens against invasive S. Typhimurium and S. Enteritidis (2–7). As these vaccines enter clinical development, it is important to establish their immunogenicity through relevant, well-characterized immunological assays. There is a growing realization within the vaccinology community that in addition to determining antibody levels through binding assays, it is important to determine the functional, antimicrobial capacity of these antibodies.
Although many studies have evaluated anti-Salmonella serum bactericidal and opsonophagocytic antibody activity (2, 8–15), there is a need for reproducible assays that can be used routinely to characterize functional antibody responses to Salmonella in a standardized manner. We previously developed a complement-dependent serum bactericidal antibody (SBA) activity assay that quantifies serum antibody responses to typhoidal (S. Typhi and S. Paratyphi A) and nontyphoidal (S. Typhimurium and S. Enteritidis) Salmonella (16). Here, we describe an assay that measures the opsonophagocytic capacity of NTS antibodies based on the well-characterized pneumococcus opsonophagocytic activity (OPA), which employs HL-60 phagocytic cells and a standardized source of complement, i.e., baby rabbit complement (BRC) (17, 18).
The Salmonella OPA assay was evaluated using serum samples from mice that had been vaccinated with live-attenuated NTS vaccines (7). BALB/c mice were orally immunized with 109 CFU of S. Typhimurium CVD 1931 or S. Enteritidis CVD 1944 suspended in phosphate-buffered saline (PBS) or given PBS alone on days 0, 28, and 56, as previously described (7). S. Typhimurium CVD 1931 and S. Enteritidis CVD 1944 are vaccine strains derived from invasive disease-associated clinical isolates (S. Typhimurium D65 and S. Enteritidis R11, respectively), which harbor deletions in guaBA and clpX. Serum samples from vaccinated mice taken 83 days postimmunization were designated immune, and serum samples from mice that received PBS were designated nonimmune.
HL-60 cells were maintained in RPMI 1640 complete medium (RPMI 1640 [Corning CellGro, Manassas, VA] supplemented with 10% [vol/vol] fetal bovine serum [FBS; HyClone, Waltham, MA] and 1% GlutaMAX, 1% sodium pyruvate, and 1% penicillin-streptomycin [Life Technologies]) and differentiated for 6 days in RPMI 1640 complete medium, 0.8% dimethylformamide (Fisher Scientific, Waltham, MA). Prior to use, cells were washed once in Hanks balanced salt solution (HBSS) (Life Technologies) and resuspended to 1 × 106 cells/ml in opsonization buffer (OPB) (HBSS, 0.1% gelatin [Sigma-Aldrich, St. Louis, MO], 10% FBS). Salmonella strains (S. Typhimurium D65 and S. Enteritidis SO1 [19]) from overnight cultures were diluted 1 in 1,000 in Hy-Soy medium (0.5% sodium chloride, 1% Hy-Soy [Kerry, Clackmannshire, United Kingdom], and 0.5% Hy-Yest [Kerry]), grown at 37°C to an optical density at 600 nm (OD600) of 0.3, and then diluted to 3 × 104 CFU/ml in OPB. Immune and nonimmune sera were heat-inactivated at 56°C for 20 min, and 2-fold serial dilutions in OPB (25 μl/well final volume) were performed in a U-bottomed 96-well microplate (Sigma-Aldrich, St. Louis, MO). The bottom row was left without sera to act as a negative control. To each well, 3 × 102 CFU of bacterial suspension in 10 μl OPB was added. Opsonization of bacteria was allowed to occur for 15 min at 37°C with 5% CO2. After incubation, 25 μl of BRC (lot 31235; Pel-Freez Biologicals, Rogers, AZ) was added to the wells at a final concentration of 12.5% (vol/vol), and 40 μl of differentiated HL-60 cells was added at a concentration of 4 × 104 cells/well. Plates were incubated at 37°C with shaking at 160 rpm for 45 min. Bacteria were enumerated by mixing the contents of each well by pipetting vigorously and performing viable counts in triplicate for each well. OPA titer was defined as the reciprocal of the highest serum dilution that produced >50% killing compared with the killing observed in the negative-control wells. Titers were determined from the mean bacterial counts in duplicate wells.
First, we determined the components required to demonstrate antibody-mediated OPA. Serum samples from mice immunized with CVD 1931 (S. Typhimurium D65 ΔguaBA ΔclpX) were tested for OPA with HL-60 cells and BRC in different combinations against an invasive clinical isolate of S. Typhimurium D65 from Mali, as shown in Fig. 1. In the absence of HL-60 cells, there was no detectable bacterial killing. Thus, at a concentration of 12.5%, BRC was unable to mediate killing, as shown previously (16). When HL-60 cells were incubated with bacteria and serum without BRC, limited OPA titers were observed for immune mice (1:400) but not for nonimmune mice. However, when BRC was included in the assay, the OPA titer for immune sera increased >100-fold. It was therefore determined that a sublethal concentration of 12.5% BRC is required for optimal OPA in this assay.
FIG 1.
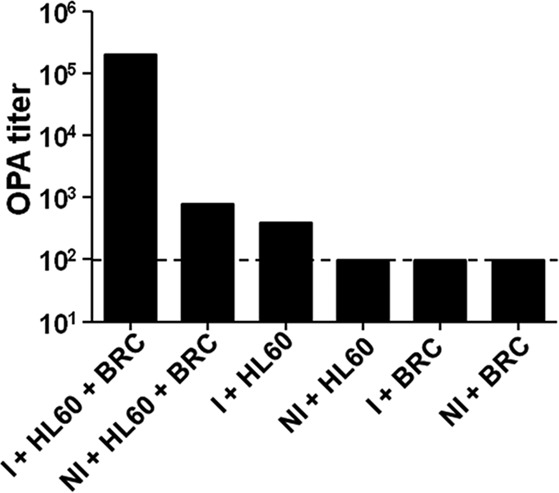
Optimization of OPA assay. To determine all of the components required for optimal opsonophagocytosis, we optimized the OPA assay using pooled immune serum produced by live-attenuated S. Typhimurium CVD 1931 (S. Typhimurium D65 ΔguaBA ΔclpX) and measured the killing of S. Typhimurium D65. We determined the effect of combining HL-60 cells and baby rabbit complement (BRC) and immune sera (I) or nonimmune sera (NI) in various combinations. OPA titer was determined as the reciprocal of the highest serum dilution that produced >50% killing in comparison to bacteria incubated with BRC and HL-60 cells. The dashed line represents the lowest dilution tested. Results shown are the titers obtained from a single experiment. Titers obtained for the conditions containing immune serum were verified by repeating the experiment using a different pool of immune serum from the same mice. Similar OPA titers were obtained (I + HL-60 + BRC, 1:204,800; I + HL60, 1:100; and I + BRC, 1:100).
Using our optimized OPA assay, we determined OPA titers of serum samples from individual mice immunized with live-attenuated NTS vaccines. We tested samples from groups that received the live-attenuated S. Typhimurium vaccine strain CVD 1931 (S. Typhimurium D65 ΔguaBA ΔclpX) against the target strain S. Typhimurium D65 from which the vaccine was derived. As expected, significantly higher OPA titers were measured in postvaccination sera than in samples from PBS-treated controls (Fig. 2A). Similarly, when we tested serum samples from mice immunized with the live-attenuated S. Enteritidis vaccine strain CVD 1944 (S. Enteritidis R11 ΔguaBA ΔclpX) against the virulent target strain S. Enteritidis S01, also isolated from the blood of a Malian child, we observed significantly higher OPA titers than for samples from mice treated with PBS (Fig. 2B). To account for possible lot-to-lot variation in BRC from the supplier, we picked 10 different serum samples from mice immunized with the live-attenuated S. Enteritidis vaccine strain CVD 1944 (S. Enteritidis R11 ΔguaBA ΔclpX) that were previously tested (Fig. 2B, open circles) for OPA titers with BRC lot 31325 and retested them in an OPA assay with BRC lots 31325, 34636, and 19837. In our hands, we observed the OPA titers of the individual serum samples to be exactly the same for all three BRC lots. Furthermore, no differences in OPA titers were observed when the 10 immune serum samples were tested a second time on a different day with the same lot of BRC (lot 31325). A significant correlation was seen between OPA titers and anti-lipopolysaccharide (anti-LPS) serum IgG titers for S. Typhimurium and S. Enteritidis (Spearman's correlation coefficients of 0.85 and 0.88, respectively; P < 0.0001 for both) (Fig. 3). Using seroconversion as the parameter, all of the S. Typhimurium- and S. Enteritidis-vaccinated mice showed a >4-fold increase in OPA titer compared to preimmune serum, while 20 of 21 S. Typhimurium-vaccinated mice and 19 of 20 S. Enteritidis-vaccinated mice seroconverted serum IgG anti-LPS titer (Table 1). The fact that the S. Typhimurium- and S. Enteritidis-vaccinated mice were protected (86% and 76% vaccine efficacies, respectively) against a lethal challenge (7) suggests that the OPA assay not only provides a measure of biological function but appears to be highly sensitive in detecting seroconversion following vaccination. The specificity of the OPA antibodies remains to be determined.
FIG 2.
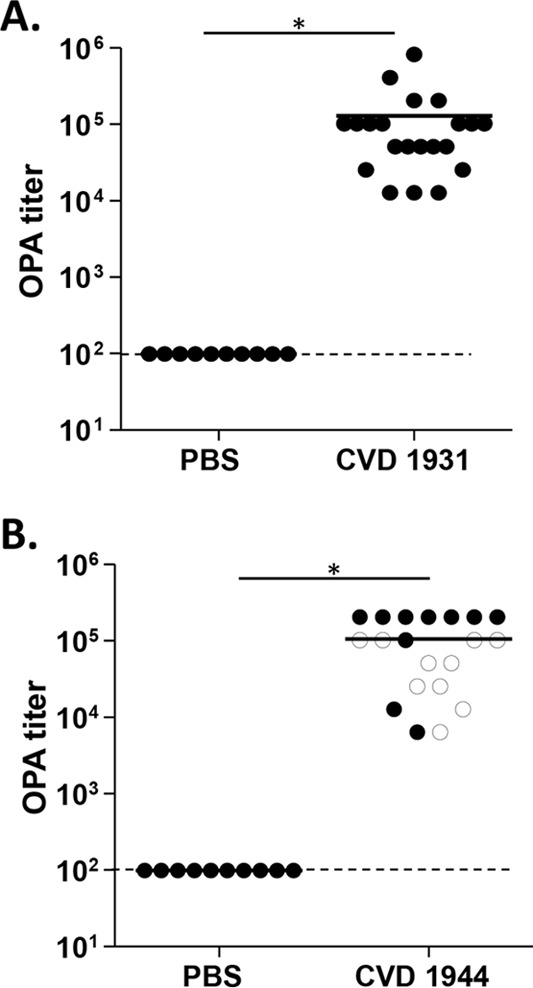
OPA titers for individual mice immunized with S. Typhimurium CVD 1931 and S. Enteritidis CVD 1944. (A) OPA titers against wild-type S. Typhimurium D65 in serum samples from individual mice immunized with CVD 1931 (S. Typhimurium D65 ΔguaBA ΔclpX) or PBS. (B) OPA titers against wild-type S. Enteritidis SO1 in serum samples from individual mice immunized with CVD 1944 (S. Enteritidis R11 ΔguaBA ΔclpX) or PBS. OPA titer was determined as the reciprocal of the highest serum dilution that produced >50% killing in comparison to bacteria incubated with BRC and HL-60 cells. Open circles, serum samples that were retested; *, P < 0.05 (Student's t test). Dashed line, the lowest dilution tested.
FIG 3.
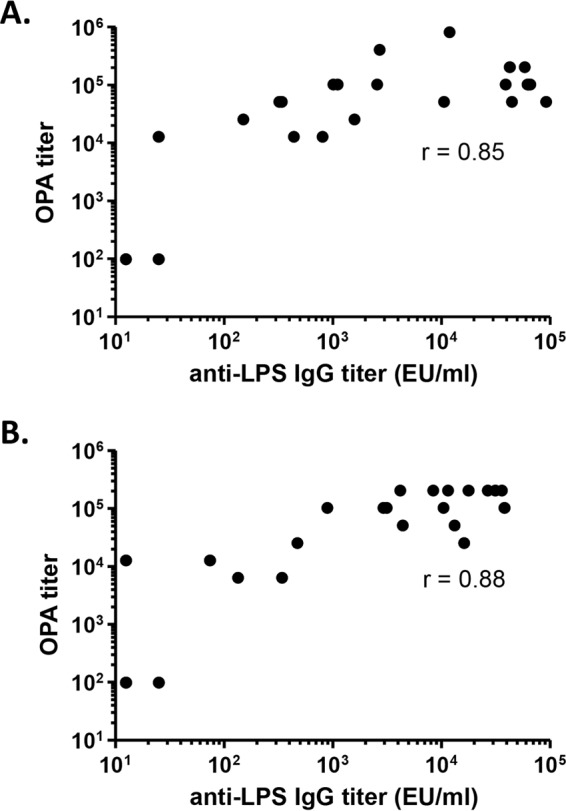
Correlation between anti-LPS serum IgG and OPA titers. Titers elicited by S. Typhimurium CVD 1931 (S. Typhimurium D65 ΔguaBA ΔclpX) (A) and S. Enteritidis CVD 1944 (S. Enteritidis R11 ΔguaBA ΔclpX) (B). Data were analyzed using Spearman's correlation coefficient.
TABLE 1.
Seroconversion and anti-LPS serum IgG and OPA geometric mean titers (GMT)
| Vaccine | Anti-LPS serum IgG titer |
OPA titer |
||||
|---|---|---|---|---|---|---|
| Preimmune GMT (95% CI) | Immune GMT (95% CI) | Seroconversiona | Preimmune GMT (95% CI) | Immune GMT (95% CI) | Seroconversiona | |
| S. Typhimurium CVD 1931 | 12.5 (12.5–12.5) | 4,337 (1,423–13,218) | 20/21 | 100 (100–100) | 71,223 (43,412–116,849) | 21/21 |
| S. Enteritidis CVD 1944 | 12.5 (12.5–12.5) | 3,210 (1,104–9,329) | 19/20 | 100 (100–100) | 65,258 (36,925–115,331) | 20/20 |
No. of mice with ≥4-fold rise in titer.
Robust, reproducible assays to measure immune responses elicited by candidate vaccines are required by regulatory agencies for eventual licensure. Here, we have adapted a well-characterized pneumococcus OPA assay accepted by regulatory agencies to evaluate pneumococcal conjugate vaccine formulations for measurement of OPA antibodies against Salmonella. Although the contribution of opsonophagocytic antibody activity in controlling Salmonella infections is unknown, the assay described herein may be used to characterize NTS vaccine immune responses, allowing quantification of functional antibody titers. The robustness of this assay, a measure of its reproducibility, was determined by using different lots of BRC and testing the assay on different days where the sera were subjected to repeated freeze-thaw cycles. We found the assay to be reproducible, with no differences observed in the OPA titers of the immune sera. We previously observed a similar robustness for SBA titers using immune mouse serum samples. Although SBA assays for NTS have been described, the need for an OPA assay is still compelling, as immune sera may lack SBA activity but still possess OPA that represents a correlate of protection for vaccines. Together with the SBA assay that we described previously, these assays may be useful in determining NTS correlates of protection, thereby allowing vaccine developers to predict vaccine performance.
ACKNOWLEDGMENT
This work was supported by a grant from NIH/NIAID (2 U54 AI057168) (principal investigator, M.M.L.).
REFERENCES
- 1.Feasey NA, Dougan G, Kingsley RA, Heyderman RS, Gordon MA. 2012. Invasive non-typhoidal Salmonella disease: an emerging and neglected tropical disease in Africa. Lancet 379:2489–2499. doi: 10.1016/S0140-6736(11)61752-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ault A, Tennant SM, Gorres JP, Eckhaus M, Sandler NG, Roque A, Livio S, Bao S, Foulds KE, Kao SF, Roederer M, Schmidlein P, Boyd MA, Pasetti MF, Douek DC, Estes JD, Nabel GJ, Levine MM, Rao SS. 2013. Safety and tolerability of a live oral Salmonella typhimurium vaccine candidate in SIV-infected nonhuman primates. Vaccine 31:5879–5888. doi: 10.1016/j.vaccine.2013.09.041. [DOI] [PubMed] [Google Scholar]
- 3.MacLennan CA, Martin LB, Micoli F. 2014. Vaccines against invasive Salmonella disease: current status and future directions. Hum Vaccin Immunother 10:1478–1493. doi: 10.4161/hv.29054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Simon R, Tennant SM, Wang JY, Schmidlein PJ, Lees A, Ernst RK, Pasetti MF, Galen JE, Levine MM. 2011. Salmonella enterica serovar Enteritidis core O polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. Enteritidis. Infect Immun 79:4240–4249. doi: 10.1128/IAI.05484-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Simon R, Wang JY, Boyd MA, Tulapurkar ME, Ramachandran G, Tennant SM, Pasetti M, Galen JE, Levine MM. 2013. Sustained protection in mice immunized with fractional doses of Salmonella Enteritidis core and O polysaccharide-flagellin glycoconjugates. PLoS One 8:e64680. doi: 10.1371/journal.pone.0064680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stefanetti G, Rondini S, Lanzilao L, Saul A, MacLennan CA, Micoli F. 2014. Impact of conjugation chemistry on the immunogenicity of S. Typhimurium conjugate vaccines. Vaccine 32:6122–6129. doi: 10.1016/j.vaccine.2014.08.056. [DOI] [PubMed] [Google Scholar]
- 7.Tennant SM, Wang JY, Galen JE, Simon R, Pasetti MF, Gat O, Levine MM. 2011. Engineering and preclinical evaluation of attenuated nontyphoidal Salmonella strains serving as live oral vaccines and as reagent strains. Infect Immun 79:4175–4185. doi: 10.1128/IAI.05278-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Trebicka E, Shanmugam NK, Mikhailova A, Alter G, Cherayil BJ. 2014. Effect of human immunodeficiency virus infection on plasma bactericidal activity against Salmonella enterica serovar Typhimurium. Clin Vaccine Immunol 21:1437–1442. doi: 10.1128/CVI.00501-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Trebicka E, Jacob S, Pirzai W, Hurley BP, Cherayil BJ. 2013. Role of antilipopolysaccharide antibodies in serum bactericidal activity against Salmonella enterica serovar Typhimurium in healthy adults and children in the United States. Clin Vaccine Immunol 20:1491–1498. doi: 10.1128/CVI.00289-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wahid R, Zafar SJ, McArthur MA, Pasetti MF, Levine MM, Sztein MB. 2014. Live oral Salmonella enterica serovar Typhi vaccines Ty21a and CVD 909 induce opsonophagocytic functional antibodies in humans that cross-react with S. Paratyphi A and S. Paratyphi B. Clin Vaccine Immunol 21:427–434. doi: 10.1128/CVI.00786-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gat O, Galen JE, Tennant S, Simon R, Blackwelder WC, Silverman DJ, Pasetti MF, Levine MM. 2011. Cell-associated flagella enhance the protection conferred by mucosally-administered attenuated Salmonella Paratyphi A vaccines. PLoS Negl Trop Dis 5:e1373. doi: 10.1371/journal.pntd.0001373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lindow JC, Fimlaid KA, Bunn JY, Kirkpatrick BD. 2011. Antibodies in action: role of human opsonins in killing Salmonella enterica serovar Typhi. Infect Immun 79:3188–3194. doi: 10.1128/IAI.05081-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pulickal AS, Gautam S, Clutterbuck EA, Thorson S, Basynat B, Adhikari N, Makepeace K, Rijpkema S, Borrow R, Farrar JJ, Pollard AJ. 2009. Kinetics of the natural, humoral immune response to Salmonella enterica serovar Typhi in Kathmandu, Nepal. Clin Vaccine Immunol 16:1413–1419. doi: 10.1128/CVI.00245-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.MacLennan CA, Gondwe EN, Msefula CL, Kingsley RA, Thomson NR, White SA, Goodall M, Pickard DJ, Graham SM, Dougan G, Hart CA, Molyneux ME, Drayson MT. 2008. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J Clin Invest 118:1553–1562. doi: 10.1172/JCI33998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gondwe EN, Molyneux ME, Goodall M, Graham SM, Mastroeni P, Drayson MT, MacLennan CA. 2010. Importance of antibody and complement for oxidative burst and killing of invasive nontyphoidal Salmonella by blood cells in Africans. Proc Natl Acad Sci U S A 107:3070–3075. doi: 10.1073/pnas.0910497107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Boyd MA, Tennant SM, Saague VA, Simon R, Muhsen K, Ramachandran G, Cross AS, Galen JE, Pasetti MF, Levine MM. 2014. Serum bactericidal assays to evaluate typhoidal and nontyphoidal Salmonella vaccines. Clin Vaccine Immunol 21:712–721. doi: 10.1128/CVI.00115-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Romero-Steiner S, Frasch C, Concepcion N, Goldblatt D, Kayhty H, Vakevainen M, Laferriere C, Wauters D, Nahm MH, Schinsky MF, Plikaytis BD, Carlone GM. 2003. Multilaboratory evaluation of a viability assay for measurement of opsonophagocytic antibodies specific to the capsular polysaccharides of Streptococcus pneumoniae. Clin Diagn Lab Immunol 10:1019–1024. doi: 10.1128/CDLI.10.6.1019-1024.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Romero-Steiner S, Frasch CE, Carlone G, Fleck RA, Goldblatt D, Nahm MH. 2006. Use of opsonophagocytosis for serological evaluation of pneumococcal vaccines. Clin Vaccine Immunol 13:165–169. doi: 10.1128/CVI.13.2.165-169.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tennant SM, Diallo S, Levy H, Livio S, Sow SO, Tapia M, Fields PI, Mikoleit M, Tamboura B, Kotloff KL, Nataro JP, Galen JE, Levine MM. 2010. Identification by PCR of non-typhoidal Salmonella enterica serovars associated with invasive infections among febrile patients in Mali. PLoS Negl Trop Dis 4:e621. doi: 10.1371/journal.pntd.0000621. [DOI] [PMC free article] [PubMed] [Google Scholar]