

Abstract
In many species, the loss of alpha matriarchs is associated with a number of negative outcomes such as troop fission, eviction, wounding, and reduced vitality. However, whether the dramatic consequences of their loss are due to their role as an old experienced figure or to their alpha status remains unclear. In a retrospective study, we tested that in a semi-free ranging colony of rhesus macaques (Macaca mulatta), the removal of a non-alpha matriarch, who had a large set of kin, led to changes in behavior and physiological stress within her matriline. Following her removal, her matriline increased in aggression, vigilance, and social grooming. Additionally, hierarchical stability, measured by levels of rank changes, decreased within her matriline, and levels of intense aggression by high-ranking animals were more frequent, as well as matrilineal wounding. Although ordinal rank was positively associated with higher chronic hair cortisol concentrations (HCCs) in the months before the matriarch’s removal, following her removal, only those who experienced large increases in rank within her matriline displayed higher HCCs. Changes in matrilineal stability, aggression, behavior, and HCCs within the other two matrilines in the troop were not evident, although caution is needed due to the small sample sizes. We conclude that the removal of the non-alpha matriarch led to matrilineal instability, characterized by higher levels of aggression and subsequent vigilance, rank changes, physiological stress, and grooming. We suggest that non-alpha matriarchs with a large number of kin and social support can be integral to the stability of matrilines.
Introduction
Among long-living animals, old females can play a crucial role for the survival of their family or social group. This support can come indirectly if these females use the experience they have accumulated over the years to lead the group and protect it from external threats [1–3] or directly by providing care to the future generations [4] or, at least in some cases, by controlling intra-group conflicts [5]. For example, in African elephants (Loxodonta africana), family units are commonly led by the oldest female, who exhibits the necessary social and ecological knowledge to coordinate her family and better recognize dangers [1, 6–8]. Accordingly, older female elephants were found to be better at discriminating between familiar and stranger groups [6], and at displaying more appropriate responses to predators [1] than young females, which can explain why old females tend to have larger families with more calves [6]. Similarly, among orcas (Orcinus orca), menopausal old females use their ecological knowledge to guide their families towards feeding sources, especially during periods of food scarcity [2], a trait that likely increases their offspring’s survival [9]. In chimpanzees (Pan troglodytes), females have been observed acting as mediators to reconcile individuals (generally males) during aggressive interactions [10–12], while in humans (Homo sapiens) the presence of grandmothers enhances grandchildren’s survival by increasing their foraging efforts [13–16].
Collectively, these studies have highlighted the importance of older females for their social group, and there is evidence that their loss can have dramatic effects on the rest of the group. In elephants, the death of matriarchs results in family unit splitting [17, 18] and increases the likelihood of calf death [19]. In ring-tailed lemurs (Lemur catta), the loss of the troop’s matriarch led to the eviction of distantly related kin [20], while in captive rhesus macaques (Macaca mulatta), matrilines with matriarchs received fewer wounds than matrilines without matriarchs [21]. However, since in many species matriarchs are often also alpha females, it is unclear whether the disruptive consequences of their loss on the family socio-dynamics are due to their influence as experienced females or rather to the important role of alpha females in the group. This is because high-ranking individuals commonly stabilize the social group through their interventions in ongoing conflicts in the attempt to appease opponents [5, 10, 22, 23]. Accordingly, the loss of dominant individuals has been found to increase social instability [11, 24, 25], to change social behaviors in the group [11, 26–28] and to affect individuals’ stress levels [29]. However, a detailed analysis of the behavioral and physiological consequences of the death or removal of non-alpha matriarchs is currently lacking.
Rhesus macaques are good models for studying the consequences of the removal of non-alpha matriarchs. They form large multi-male multi-female groups with an average group size between 10 and 80 individuals [30–32]. Male macaques generally emigrate to neighboring communities around sexual maturity at 4–5 years of age [33], while females remain in the natal group and form linear dominance hierarchies on the basis of their matrilineal kinship. Thus, matriarchs with large families are likely to both have and provide substantial social support (as mothers often aid their offspring in conflicts [34]) as well as to influence social dynamics within their matriline.
Here, we use the definition of a matriarch as the oldest living member of a family (where family is defined as descended from a common female ancestor). As females in our troop have their first offspring around 3–5 years of age, a family’s matriarch has at least a grandmother status. Because rank in rhesus macaques is determined by birth rank (i.e. mother’s rank) and not age [35], a matriarch is not necessarily the alpha female. We tested the hypothesis that the removal of a non-alpha matriarch in a captive colony of rhesus macaques significantly changed behavior and physiology in her matriline but not within the other matrilines present in the troop. At the time of this study, this matriarch, H1, had seven offspring in the population, and had birthed a total of 13 offspring overall, making her the matriarch with the largest number of extant offspring in the troop. Two of H1’s sons were high-ranking males (2nd and 3rd) in the troop, and her two younger sisters were the troop’s alpha and beta females (with the beta’s eldest daughter, P1, outranking H1, and P2 and PA1 beginning to ascend the hierarchy but not yet ranking above H1, see Fig 1). Thus, although H1 was not one of the top three ranking females in the troop, it is likely that she exerted a strong influence on matrilineal dynamics.
Fig 1. Pedigree of Matriline 3 members from a larger troop of rhesus macaques at the LCE field station.

Deceased relatives are denoted by parentheses and italicized font. Underlines indicate males. (#) represents rank within sex (limited to #1–3 to show H1’s relationships to top-ranking members; Alpha male is from matriline 1). *denotes individuals with behavioral data
We tested this hypothesis by retrospectively examining dominance stability, vigilance, grooming, aggressive interactions, and hair cortisol concentrations (HCCs) within H1’s matriline, while also examining the other two matrilines within the troop and overall troop wounding, before and after her removal. We assessed HCC as a measure of chronic hypothalamic-pituitary- adrenocortical (HPA) activity. HCC measurement has several advantages over the collection of other biological samples (i.e. blood, urine, and feces), including the ability to measure chronic HPA activity without the need of multiple sampling and without the confounds of circadian and environmental variability [36–38]. We hypothesized that the matriarch provided stability within the matriline, and predicted that her removal would lead to 1) rank changes, and thus higher instability (prediction 1), 2) higher aggression rates (prediction 2), and 3) higher levels of wounding (prediction 3). An increase of rank instability and aggression rates following the matriarch’s removal should, in turn, increase individuals’ vigilant behaviors (prediction 4) and should increase HCCs as an index of chronic stress, especially among high-ranking animals (prediction 5) as their dominance positions are contested [29]. Finally, previous work has shown how social grooming can decrease social tension among primates [39, 40], and thus we predicted rates of social grooming would increase during the period of social instability (prediction 6). Finally, if social grooming can help alleviate chronic stress, we also predicted that monkeys with the highest frequency of grooming would have relatively lower hair cortisol concentrations (prediction 7). Because the matriarch is likely the most influential within her matriline, we did not predict any significant changes within the other two matrilines, which were presumably unaffected.
Materials and Methods
Subjects and housing
Subjects were 49 rhesus macaques (age range 3–17 years) born and reared at the Laboratory of Comparative Ethology (LCE) field station at the NIH Animal Center in Poolesville, Maryland, USA. Because there is an age-related decline in HCC in infants and juveniles that later stabilizes [41], we included all subjects above three years old. The field station is a 2.0 hectare outdoor open air enclosure consisting of natural vegetation, climbing structures, corncrib shelters, and a central pond measuring 0.09 hectares [42]. Continuous access to three indoor enclosures (2.74 x 5.79 x 4.27m) was also available, where animals had ad libitum access to water, as well as commercial lab diet (Purina Monkey Chow #5038, St. Louis, MO) and were given fresh fruits and vegetables twice daily. All procedures described below adhered to the NIH Guide for the Care and Use of Laboratory Animals and were approved by the NICHD Animal Care and Use Committee (ACUC). Furthermore, all applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Group structure
The troop consisted of three major matrilines, which are related individuals descending from a female line. Matriline 3 has occupied the top rank since 2009, when its major families overthrew the previously dominant matriline 1 [42] leading to severe injuries and therefore the permanent removal of most members of matriline 1. Matrilineal hierarchies were ranked in the order, from highest to lowest: matriline 3, matriline 4, and matriline 1 (matriline 2 was removed in 2004 for management reasons) [42]. The demographic composition of the troop is shown in Table 1. Subjects were 49 adults (matriline 3: 29; matriline 4: 15; matriline 1: 5).
Table 1. Troop composition at the LCE field station in 2014.
| Adults (3+) | Juveniles/infants (<2) | Total | |
|---|---|---|---|
| Matriline 3 | 29 (38%) | 14 (18%) | 43 (57%) |
| Matriline 4 | 15 (20%) | 11 (15%) | 26 (34%) |
| Matriline 1 | 5 (7%) | 2 (3%) | 7 (9%) |
| Total | 49 (64%) | 27 (36%) | 76 |
Numbers in parentheses represent proportion out of entire troop (%).
Demographic change: Removal of matriline 3 matriarch
In late September 2014, the troop’s eldest (but non-alpha) matriarch, H1 (18yrs), was removed from the field station due to chronic gastrointestinal issues that resulted in chronic diarrhea and weight loss and necessitated treatment that could not be provided in the field.
H1’s offspring remained in the troop after her removal, including six adults (five females, one male) and one juvenile female. However, the two youngest females were permanently removed from the population at the bi-annual health exam in February 2015 as part of routine population management.
Data collection and analysis
Behavioral data
Behavioral data were collected using modified frequency data sheets [43] from eight females and two males (see Fig 1), representing every major lineage within matriline 3, through continuous 5-minute focal animal interval sampling [44]. Data were also collected from the three highest-ranking females from matriline 4 and the two highest-ranking adults in matriline 1 (the troop’s alpha male and one female). Due to limited researcher availability, focal behavioral data were only collected on this subset of the population.
The 5-min samples were divided into 20, 15-second intervals. For each interval, every behavior that occurred within the 15 seconds was recorded. The frequency of intervals in which the behaviors occurred was then calculated. The maximum number of intervals an animal could perform a behavior therefore in one session was 20 intervals. Focal subjects received one weekly AM session (9:00–12:00) and one weekly PM session (12:00–17:00) for ten consecutive weeks from August to December 2014 by three observers (inter-rater reliability >85%). Data used were part of a long-term data collection procedure for unrelated projects, and therefore, coders were blind to the hypotheses of the study.
The focal animals within matriline 3 included: 1) the top three ranking females (H1’s sisters and niece: H3, H2, P1; see Fig 1); 2) a female who underwent a major rank change and rose above the matriarch’s offspring (CX); 3) a matriarch ranking immediately below H1’s family (BA1); 4) one son of the matriarch (2nd in the male hierarchy, K3), 5) one high-ranking male (CH2, distant cousin to H1), 6) and three intermediate ranking females within the matriline (B2, Y2, Y3). From the focal animals, we recorded data on both social (e.g. aggression, social grooming) and non-social (e.g. feeding, vigilance) behaviors (only behaviors in Table 2 were analyzed for this study). In order to calculate rank changes and dominance stability, we noted all instances of dominance interactions (i.e. supplant, threat, chase, attack, and submissive) within the entire troop through ad libitum observations [44]. We collected 600 focal observations (40 sessions/focal animal, 20 before and 20 after), for a total of 50 hours of observations (25 hours before and 25 hours after H1’s removal), as well as 1,258 ad libitum troop dominance interactions [44].
Table 2. Behaviors and definitions collected during focal observations.
| Behavior | Definition |
|---|---|
| Initiation of aggression | Displacements (taking the place of another), threats (open mouth, ears back, direct eye contact, includes lunges), attack (pin down and bite) |
| Grooming | Move hair apart and visual inspection, removal of debris; combination of given and received bouts |
| Vigilance | Scan the environment visually, at least 3 seconds |
Behavioral observations thus included the ten focal subjects from matriline 3 and the five focal subjects from matrilines 4 and 1, while ad libitum aggressive interactions included all 49 subjects from across the three matrilines.
Dominance rank and social instability
Dominance hierarchies were calculated via Elo-rating, a numerical system that tracks rank changes over time by constantly updating values based on wins and losses [45–49]. We generated Elo-ratings using the elo.sequence function provided by Neumann et al. [45] in R software (v 3.1.2), by setting animals’ initial rating at 1,000, and the k factor, which is weighted based on the probability of winning, at 200. Two main advantages of the use of Elo-rating over conventional matrix-based analyses include the ability to track rank changes over time, and accommodating variations in social dynamics due to a fluctuations in the study population [45, 47], making Elo-rating ideal for this study.
Social instability was calculated through the stability.index function derived from Neumann et al. [45]. This function measures the ratio of rank changes per individuals present over a given time period based on the derived Elo-ratings. As Elo-ratings can be arranged to reflect ordinal ranks, fluctuations in Elo-ratings reflect higher levels of instability. According to Neumann et al. [45], S values typically range from 0 to 0.5, where higher S values reflect higher levels of instability, and lower values reflect a relatively stable hierarchy. Stability measures were taken for the three months before and after H1’s removal for each matriline. Dominance ranks and stability measures thus included data from all 49 subjects across the three matrilines.
Severe aggression
Ad libitum records of severe aggression (defined as interactions resulting in tissue damage) were routinely recorded as part of the long-term data collection procedures for this colony of animals. For this study, instances of severe aggression from August to December 2014 were examined. Because these types of social interactions are rare, ad libitum sampling is the optimal recording method. Severe aggression was analyzed to determine whether H1’s removal resulted in an increase in this behavior for her matriline compared to matrilines 4 and 1.
Hair sample collection and cortisol assay
Hair samples were collected by shaving the back of the animals’ necks [36] during routine health exams in August 2014 and February 2015 for all 49 subjects across the three matrilines. Because these animals had been shaved as part of a longitudinal study every six months beginning in Fall 2012, the samples in August 2014 reflected hair cortisol concentrations (HCCs) that had accumulated since February 2014, and the February 2015 samples reflected HCCs that had accumulated since August 2014 (thus reflecting the timing of H1’s removal in September 2014). Samples were stored in a foil pouch at -80°C until shipment to the Hormone Assay Core Laboratory at the University of Massachusetts Amherst. Following Dettmer et al. [41], samples were weighed, washed twice with isopropanol and dried for 5–7 days under a fume hood. Samples were then ground to a fine powder with a ball mill grinder (MM200; Retsch, Newtown, PA) and incubated in methanol for 24 hours to extract cortisol from the samples. Aliquots of the methanol extract were dried down and reconstituted with assay buffer, then analyzed via enzyme immunoassay (EIA) using a salivary cortisol kit (#1–3002; Salimetrics, State College, PA). Resulting values (ug/dL) were converted to pg/mg for analysis. Inter- and intra-assay coefficients of variation were <8% based on aliquots of the same extracted pooled hair sample analyzed repeatedly across assays.
Data analysis
Due to the small sample size of matriline 1 (N = 5), data for matrilines 4 and 1 were analyzed together for all analyses.
To assess whether focal animals changed frequencies in their behaviors following H1’s removal, we compared rates of each behavior collected approximately three months before and after her removal using paired-sample t-tests for both matriline 3 and the combined matrilines 4 and 1.
Subjects were classified as high or low ranking (within their respective matriline) based on a median split of the Elo-ratings. Levels of intense aggression (i.e. chases and attacks) were compared using both focal and ad libitum data using paired-sample t tests to examine whether high ranking monkeys increased in intense aggression.
Severe aggression was analyzed as the proportion of events that occurred across each matriline, (i.e. out of all recorded events, how many were from each matriline) and Fisher’s exact tests were used to compare changes in proportion of events for each matriline before and after H1’s removal.
HCC values were log transformed to ensure normality prior to analysis. Seven subjects were missing a HCC measurement in August (three from matriline 3, three from matriline 4, one from matriline 1), as they could not be captured during routine exams. Two subjects were also missing a HCC measurement in February (both from matriline 4; one of which did not have any HCC data for either time point). One subject from matriline 3 was also excluded from the August HCC analysis (as well as the HCC change analysis), as the HCC measurement in August was well above six standard deviations from the mean. We tested the association between individuals’ HCCs and their Elo-rating using Pearson’s correlation test for both matriline 3 and matrilines 4/1. Additionally, we ran Spearman’s correlation test (owing to the smaller sample sizes) between HCC changes (from August to February, in pg/mg) and Elo-rating changes for high and low ranking monkeys. We predicted that an increase in rank would be associated with an increase in physiological stress among high-ranking monkeys in matriline 3 as their dominance positions were contested.
To examine whether frequencies of social grooming were associated with HCCs after the matriarch’s removal, we calculated a grooming frequency across the entire 6 months (from August to February; the total amount of time that HCC accumulated) for the 15 subjects of whom behavioral data were collected (ten from matriline 3, five from matrilines 4 and 1). We then tested the association between grooming frequency and February 2015 HCC (which reflected chronic activity since August) using Spearman’s correlation test.
All tests were two tailed with the significance level set at p<0.05. SPSS 22 was used for analyses.
Results
Social stability and behavioral changes
As expected (prediction 1), we found lower social stability after H1’s removal compared to the three months before: the stability index for matriline 3 increased from 0.039 to 0.128. Matrilines 4&1 had little change in stability (from .017 to .018). Furthermore, our focal observations revealed that in the months immediately following H1’s removal, there was a significant increase in the initiation of aggression (paired-sample t-test: t(9) = -3.20, P = 0.01, Fig 2A), in the time spent in vigilance (t(9) = -3.15, P = 0.01, Fig 2B), and in grooming time (t(9)) = -2.74, P = 0.02, Fig 2C) within matriline 3, supporting predictions 2, 4 and 6. No significant changes were found for the initiation of aggression (t(4) = 0.10, P = 0.93), vigilance (t(4) = -1.68, P = 0.17), or grooming (t(4) = -1.33, P = 0.25) for matrilines 4 and 1.
Fig 2. Frequencies of behaviors within matriline 3 before and after H1’s removal.

Frequencies represent the average number of intervals (out of 400) the behavior occurred for each focal subject. A. Aggression frequency **p = 0.01. B. Grooming frequency **p = 0.01. C. Vigilance frequency *p = 0.02.
The ad libitum and focal data revealed that high-ranking monkeys from matriline 3 also exhibited an increase in the levels of intense aggression (i.e., chases and attacks; mean ± SE: 0.67 ± 0.33 before to 7.60 ± 1.81 after; t(14) = -3.86, P = 0.002) whereas low ranking animals did not show a significant change (t(13) = -1.629, P = 0.13). There was no significant change in intense aggression for high ranking monkeys (t(9) = -1.65, P = 0.13) or low ranking monkeys (t(9) = -2.25, P = 0.05) within matrilines 4 and 1.
Severe aggression
Contrary to our expectation (prediction 3), there was a reduction in overall troop severe aggression (from 9 to 6) compared to the period prior to H1's removal. However, when compared to the number of aggressive events experienced by the other matrilines, matriline 3 suffered proportionally more wounds after H1's removal. In the three months after her removal, no severe aggression was recorded for matrilines 4 and 1, while all recorded events occurred within matriline 3 (6 events total; P=0.04). In the three months prior to her removal, the matrilines had approximately equal numbers of severe aggression (four within matriline 3 and five within matriline 4 monkeys).
Rank changes and hair cortisol concentrations
There was no significant change in HCCs from August 2014 to February 2015 for either matriline 3 (t(24) = -1.05, P = 0.31) or matrilines 4 and 1 (t(13) = 0.15, P = 0.89). For matriline 3, there was a significant positive correlation between Elo-rating and HCCs prior to H1’s removal in August (Pearson correlation: r = 0.63, N = 25, P = 0.001; see Fig 3A), but there was no correlation after her removal (r = 0.27, N = 28, P = 0.16, see Fig 3B). No significant correlations between Elo-rating and HCC were evident for matrilines 4 and 1 for either August (Pearson correlation: r = -0.23, N = 16, P = 0.39) or February (Pearson correlation, r = -0.42, N = 18, P = 0.08).
Fig 3. Relationship between rank and HCCs within matriline 3 before (3a:left panel) and after (3b:right panel) H1’s removal.

When comparing individuals based on high or low ordinal rank in matriline 3, the change in Elo-rating was positively correlated with the change in HCC (from August to February; Spearman, rs = 0.71, N = 13, P<0.01, Fig 4A) for high-ranking monkeys, indicating that as top animals increased in dominance rank (and thus dominance assertion/increase in intense aggression), they experienced an increase in physiological stress, supporting prediction 5. There was no significant correlation for low-ranking animals (Spearman, rs = -0.434 N = 12, P = 0.16, Fig 4B) within matriline 3. No relationship between Elo-rating change and HCC change was found for matrilines 4 and 1 for either high-ranking monkeys (Spearman, rs = -0.32, N = 7, P = 0.48) or low-ranking monkeys (Spearman, rs = -0.04, N = 8, P = 0.93; see S1 Dataset for data used in this current study).
Fig 4. Relationship between rank change and HCC change in high-ranking (4a: left panel) and low-ranking (4b: right panel) rhesus macaques in matriline 3.

Rank and HCC changes reflect changes from August 2014 to February 2015.
Hair cortisol concentrations and grooming frequency
While the total grooming frequency was not significantly associated with February HCC for matriline 3 (Spearman, rs = -0.56, N = 10, P = 0.15, failing to support prediction 7), a significant negative association was evident when analyzing all subjects from which grooming data were collected (including the other two matrilines; Spearman, rs = -0.52, N = 15, P = .049; see Fig 5).
Fig 5. Total grooming frequency and HCC following H1’s removal.
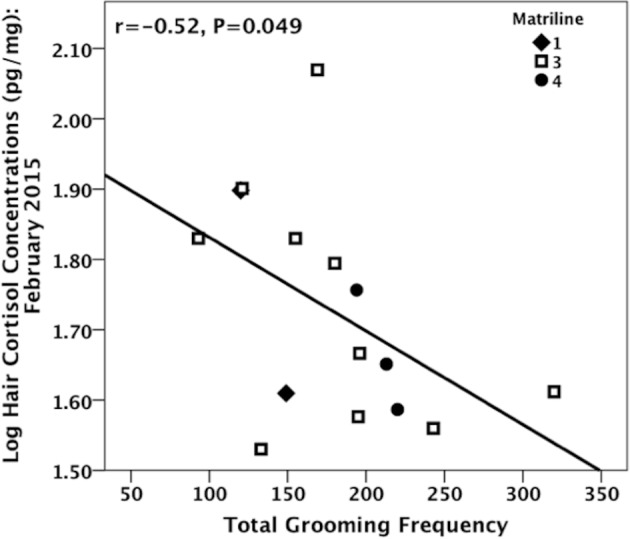
Frequency represents total number of intervals grooming occurred from August to February (when the HCC samples were taken).
Discussion
Our data indicate that a non-alpha matriarch exerted a strong influence on her matriline: with direct ties to the dominant females and a large set of kin, her social ties were significant enough to influence dominance stability within her matriline, although she was not the alpha. Accordingly, in the period following H1’s removal, her matriline endured a period of social instability, with increased aggression and higher rates of both vigilance and social grooming. Interestingly, we also found that large increases in Elo-rating after H1’s removal were associated with large increases in HCC. These findings suggest that individuals within her matriline may have experienced higher levels of chronic stress, probably because in a period of frequent rank changes, each individual struggled to exert dominance over others (hence the increase of chase and physical attacks). The lack of any significant relation between rank changes and hair cortisol for the other matrilines suggests that only H1’s matriline was affected by her removal. We found that her matriline experienced both behavioral and physiological changes that resemble the consequences of the loss or takeover of alpha individuals described in both this [25] and other mammalian species (e.g. chacma baboons, Papio ursinus [26, 50] naked mole-rat, Heterocephalus glaber [24]; chimpanzees, Pan troglodytes [11]).
Our results are consistent with findings reported from a variety of species (chacma baboons, Papio ursinus [51–54]; wild dogs, Lycaon pictus [55]; long-tailed macaques, Macaca fascicularis [56]; African cichlid fish, Haplochromis burtoni [57]) showing that social stress in dominant individuals can be linked to social instability and the use of intense aggression by dominants to affirm their position. Interestingly, we found a positive association between dominance rank and hair cortisol before H1’s removal, suggesting that high-ranking individuals may experience more chronic physiological stress than subordinates under stable conditions. This result is at first puzzling since in the period preceding the matriarch’s removal, we found stable rank relationships within the matriline, and social stability is commonly associated with higher stress levels in subordinates, who receive continuous harassment from dominants (chacma baboons [29]; but see [58] for a discussion of moderating factors that influence the relationship between social rank and physiological stress in nonhuman primates). However, this relationship might be explained by the relatively new, but stable, hierarchy present in the troop (circa 1 year) due to a social overthrow within the matriline in late December 2013/early January 2014 [47]. It is possible therefore that there might have been some degree of social tension within the matriline even in the period before H1’s removal, that her presence might have helped to hold in check, through, for instance, policing interventions [22]. Consequently, after the removal of the matriarch, and the threat to this relatively new hierarchy, this social tension was no longer under control, leading to higher levels of social instability and stress. Unfortunately, we do not have data on policing behavior to test this hypothesis directly in this study. However, we do know that H1 managed conflict among her offspring and following her removal, there was a surge in offspring conflicts, as well as challenges to their dominance positions (e.g. CX).
Notably, we did not find any association between dominance rank and HCC in the months following H1’s removal, probably because only a restricted number of individuals experienced heightened levels of stress during social instability, namely those high-ranking monkeys who increased in Elo-rating and intense aggression. Furthermore, since grooming has been shown to decrease animal stress levels [59–62] it is plausible that the increase in social grooming we found in H1’s matriline after her removal might have been a strategy monkeys used to counter-act the increased instability, and this, in turn, minimized the surge in social and physiological stress in this period. In wild chacma baboons, females who experienced increased stress levels following the removal of close relatives displayed higher rates of grooming which likely helped them to return cortisol concentrations to baseline levels [63]. While we did not find a significant relationship between total grooming frequency and HCCs specifically for H1’s matriline, we did find a significant negative relationship when analyzing all subjects for whom we had behavioral data recorded, indicating that social grooming may be an important mechanism to reduce chronic physiological stress. It is possible that with more subjects from her matriline, we would have found a significant relationship between hair cortisol and grooming specifically for her matriline. Importantly, our work is one of the few to relate social rank to HCCs in primates [64,65], and mammals in general, although one study found a positive correlation between rank and HCC in hyraxes (Procavis capensis [66]). Our study suggests that HCCs may be useful in assessing how social dynamics influence chronic stress in group-living mammals.
Following H1’s removal, instances of severe aggression were only recorded for her matriline, indicating that high intensity aggression was more frequent within her matriline. These findings are underscored by our observation of increased chases and attacks by the high-ranking moneys of matriline 3 after H1’s removal. The increase in dominance instability, and severe physical aggression, were the likely consequences of monkeys trying to reaffirm their rank. In support of this, one month after the matriarch’s removal, CX had undergone a major rank increase and ranked above all seven of H1’s offspring. During this time, she was also observed with a large tear wound. This wound was likely a byproduct of CX attempting to increase in rank (successfully), as well as the unwillingness of the others to sacrifice their positions (those attempting to reaffirm their position). Unfortunately, wounds are rarely ever seen occurring, so it is impossible to know exactly who inflicted this wound. The fact that no severe aggression was observed within the other matrilines suggests that they were likely not struggling to assert dominance over one another, a notion supported by their relatively stable hierarchies. Furthermore, the increased levels of intense aggression including those resulting in tissue damage may have resulted from changes in submissive signaling. Submissive signals (such as the silent bared teeth display) have been related to lower levels of severe aggression [67] and are associated with greater dominance relationship certainty [68]. While submissive displays were recorded, they were originally not a focus of ad libitum sampling and occurred too infrequently during focal observations, and therefore direct comparisons were not available during this study. Therefore, an interesting opportunity exists in the future to study the role and changes in submissive signaling and resultant levels of aggression during a time of social instability such as after the loss of a matriarch.
As a likely result of the increased levels of aggression and observed tissue damage, there was also an increase in the frequency of vigilance. In a time of rank changes and monkeys attempting to reaffirm their rank, vigilance may have been a buffering mechanism from both directed and redirected aggression. Additionally, monkeys may have increased in vigilance to monitor the social interactions of others. Indeed, female mountain gorillas (Gorilla gorilla beringei) spend more time monitoring individuals whom they had aggressive interactions [69] and may play a role in conflict-avoidance in tufted capuchins (Cebus apella; [70]).
Importantly, it is worth noting that the matriarch’s removal happened in the midst of the breeding season and thus the variation in aggression rates and possibly wounding rates we found in our study might be related to increased competition during the breeding season [71, 72]. However, we did not find any evidence for changes in observed severe aggression in the previous 2013 breeding season: with matrilines 3 and 4 each observed with approximately five instances of tissue damage in each time period identical to this study. In addition, we found a reduction in the overall number of severe aggression resulting in tissue damage in 2014 (from before the start of the breeding season to after), indicating that high levels of aggression may not be limited to the breeding season. Furthermore, because our findings (grooming, vigilance, aggression, HCC, rank, instability, etc.) were only significant for the matriarch’s matriline (matriline 3), and not the other matrilines (matrilines 4 and 1), it is unlikely that the findings are solely a result of the breeding season alone. However, data from matrilines 4 and 1 should be taken with caution due to the small sample sizes.
More generally, our results emphasize the importance of careful consideration in the management of a captive colony when deciding whether some individuals should be permanently removed. Although each individual primate colony is unique and management decisions will differ across facilities, it is important for researchers, colony managers, and veterinarians to consider that group members who play a key role for the stability of the social group should be identified, even if they are not among the alpha individuals. Our study shows how the removal of even non-alpha matriarchs significantly impacts matrilineal dynamics: their absence can trigger a period of social instability and heightened intense aggression. In the case in which socially significant individuals must be removed, extra monitoring is recommended to detecting potential changes in stability. Managers of captive colonies may therefore be able to use data generated via Elo-rating and rank changes (i.e. social instability) to examine how certain characteristics (age, rank, number of kin, rate of policing behavior, etc.) of an individual may be important in maintaining stability. This will then in turn allow managers to predict the effects of the removal of certain individuals in the future and guide future management decisions.
In conclusion, this study provides evidence that old-aged matriarchs may play a substantial role in family stability in group-living primates. When considering the stability of large family groups, it may be important to recognize the critical role that non-alpha matriarchs provide. Collectively, our findings demonstrate the power of specific individuals, including non-alpha group members, to exert strong influences within their group and the consequences following their loss.
Supporting Information
(XLSX)
Acknowledgments
We thank Kristen Byers for assistance with pedigree construction and Ryan McNeill for assistance with data collection. We are also grateful for the veterinary and animal care staff at the NIH Animal Center for their dedicated care.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Division of Intramural Research, National Institute of Child Health and Human Development, 1ZIAHD001107-32, awarded to SJS, https://www.nichd.nih.gov/about/org/dir/Pages/index.aspx. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.McComb K, Shannon G, Durant SM, Sayialel K, Slotow R, Poole J, et al. Leadership in elephants: the adaptive value of age. Proc R Soc B. 2011:rspb20110168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brent LJ, Franks DW, Foster EA, Balcomb KC, Cant MA, Croft DP. Ecological knowledge, leadership, and the evolution of menopause in killer whales Curr Biol. 2015;25(6):746–50. 10.1016/j.cub.2015.01.037 [DOI] [PubMed] [Google Scholar]
- 3.Davenport LC. Aid to a declining matriarch in the giant otter (Pteronura brasiliensis). PLOS One. 2010;5(6):e11385 10.1371/journal.pone.0011385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kachel AF, Premo LS, Hublin J-J. Grandmothering and natural selection. Proc R Soc B. 2011;278(1704):384–91. 10.1098/rspb.2010.1247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Beisner BA, McCowan B. Policing in nonhuman primates: Partial interventions serve a prosocial conflict management function in rhesus macaques. PLOS One. 2013;8(10):e77369 10.1371/journal.pone.0077369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McComb K, Moss C, Durant SM, Baker L, Sayialel S. Matriarchs as repositories of social knowledge in African elephants. Science. 2001;292(5516):491–4. [DOI] [PubMed] [Google Scholar]
- 7.Moss CJ. The demography of an African elephant (Loxodonta africana) population in Amboseli, Kenya. J Zool. 2001;255(02):145–56. [Google Scholar]
- 8.Wittemyer G, Douglas-Hamilton I, Getz WM. The socioecology of elephants: analysis of the processes creating multitiered social structures. Anim Behav. 2005;69(6):1357–71. [Google Scholar]
- 9.Foster EA, Franks DW, Mazzi S, Darden SK, Balcomb KC, Ford JK, et al. Adaptive prolonged postreproductive life span in killer whales. Science. 2012;337(6100):1313-. 10.1126/science.1224198 [DOI] [PubMed] [Google Scholar]
- 10.de Waal F. Chimpanzee politics: Sex and power among apes London, UK: Jonathan Cape; 1982. [Google Scholar]
- 11.Kaburu SS, Inoue S, Newton-Fisher NE. Death of the alpha: within-community lethal violence among chimpanzees of the Mahale Mountains National Park. Am J Primatol. 2013;75(8):789–97. 10.1002/ajp.22135 [DOI] [PubMed] [Google Scholar]
- 12.Nieuwenhuijsen K, de Waal F. Effects of spatial crowding on social behavior in a chimpanzee colony. Zoo Biol. 1982;1(1):5–28. [Google Scholar]
- 13.Hawkes K, O'Connell JF, Blurton Jones NG. Hadza women's time allocation, offspring provisioning, and the evolution of long postmenopausal life spans. Curr Anthropol. 1997;38(4):551–77. [Google Scholar]
- 14.Sear R, Mace R, McGregor IA. Maternal grandmothers improve nutritional status and survival of children in rural Gambia. Proc R Soc B. 2000;267(1453):1641–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sear R, Mace R. Who keeps children alive? A review of the effects of kin on child survival. Evol Hum Behav. 2008;29(1):1–18. [Google Scholar]
- 16.Croft DP, Brent LJ, Franks DW, Cant MA. The evolution of prolonged life after reproduction. Trends Ecol Evol. 2015;30(7):407–16. 10.1016/j.tree.2015.04.011 [DOI] [PubMed] [Google Scholar]
- 17.Douglas-Hamilton I, Douglas-Hamilton O. Among the elephants London: Collins & Harvill Press; 1975. [Google Scholar]
- 18.Douglas-Hamilton I, Bhalla S, Wittemyer G, Vollrath F. Behavioural reactions of elephants towards a dying and deceased matriarch. Appl Anim Behav Sci. 2006;100(1):87–102. [Google Scholar]
- 19.Foley C, Pettorelli N, Foley L. Severe drought and calf survival in elephants. Biol Lett. 2008;4(5):541–4. 10.1098/rsbl.2008.0370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Soma T, Koyama N. Eviction and Troop Reconstruction in a Single Matriline of Ring-Tailed Lemurs (Lemur catta): What happened when “grandmother” died? In: Masters J, Gamba M, Génin F, editors. Leaping ahead. Springer; 2012. p. 137–46. [Google Scholar]
- 21.Beisner BA, Jackson ME, Cameron AN, McCowan B. Detecting instability in animal social networks: genetic fragmentation is associated with social instability in rhesus macaques. PLOS One. 2011;6(1):e16365 10.1371/journal.pone.0016365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Flack JC, Girvan M, De Waal FB, Krakauer DC. Policing stabilizes construction of social niches in primates. Nature. 2006;439(7075):426–9. [DOI] [PubMed] [Google Scholar]
- 23.Jennings DJ, Carlin CM, Gammell MP. A winner effect supports third-party intervention behaviour during fallow deer, Dama dama, fights. Anim Behav. 2009;77(2):343–8. [Google Scholar]
- 24.Clarke F, Faulkes C. Dominance and queen succession in captive colonies of the eusocial naked mole–rat, Heterocephalus glaber. Proc R Soc B. 1997;264(1384):993–1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Oates-O'Brien RS, Farver TB, Anderson-Vicino KC, McCowan B, Lerche NW. Predictors of matrilineal overthrows in large captive breeding groups of rhesus macaques (Macaca mulatta). J Am Assoc Lab Anim Sci. 2010;49(2):196–201. [PMC free article] [PubMed] [Google Scholar]
- 26.Barrett L, Henzi SP, Lusseau D. Taking sociality seriously: the structure of multi-dimensional social networks as a source of information for individuals. Phil Trans R Soc B. 2012;367(1599):2108–18. 10.1098/rstb.2012.0113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kaburu SS, Newton-Fisher NE. Egalitarian despots: hierarchy steepness, reciprocity and the grooming-trade model in wild chimpanzees, Pan troglodytes. Anim Behav. 2015;99:61–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Franz M, McLean E, Tung J, Altmann J, Alberts SC. Self-organizing dominance hierarchies in a wild primate population. Proc R Soc B; 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sapolsky RM. The influence of social hierarchy on primate health. Science. 2005;308(5722):648–52. [DOI] [PubMed] [Google Scholar]
- 30.Lindburg DG, Rosenblum L. The rhesus monkey in North India: an ecological and behavioral study In: Rosenblum LA, editor. Primate behavior: developments in field and laboratory research. New York: Academic Press; 1971, pp 2–106. [Google Scholar]
- 31.Wenyuan Q, Yongzu Z, Manry D, Southwick CH. Rhesus monkeys (Macaca mulatta) in the Taihang mountains, Jiyuan county, Henan, China. Int J Primatol. 1993;14(4):607–21. [Google Scholar]
- 32.Southwick C, Yongzu Z, Haisheng J, Zhenhe L, Wenyuan Q. Population ecology of rhesus macaques in tropical and temperate habitats in China In: Fa JE, Lindburg DG, editors. Evolution and ecology of macaque societies. Cambridge: Cambridge University Press; 1996. pp 95–105. [Google Scholar]
- 33.Sade DS. A longitudinal study of social behavior of rhesus monkeys. The functional and evolutionary biology of primates In: Tuttle R, editor. Functional and evolutionary biology of primates. Chicago: Aldine-Atherton, Chicago; 1972. pp 378–398. [Google Scholar]
- 34.Bernstein I. Ehardt C. Agonistic aiding: kinship, rank, age, and sex influences. Am J Primatol; 1985; 8: 37–52. [DOI] [PubMed] [Google Scholar]
- 35.Vessey SH. Dominance among rhesus monkeys. Polit Psychol. 1984:623–8. [Google Scholar]
- 36.Davenport MD, Tiefenbacher S, Lutz CK, Novak MA, Meyer JS. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen Comp Endocrinol. 2006;147(3):255–61. [DOI] [PubMed] [Google Scholar]
- 37.Meyer JS, Novak MA. Minireview: hair cortisol: a novel biomarker of hypothalamic-pituitary-adrenocortical activity. Endocrinology. 2012;153(9):4120–7. 10.1210/en.2012-1226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Novak MA, Hamel AF, Kelly BJ, Dettmer AM, Meyer JS. Stress, the HPA axis, and nonhuman primate well-being: a review. App Anim Behav Sci. 2013;143(2):135–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Schino G, Scucchi S, Maestripieri D, Turillazzi PG. Allogrooming as a tension‐reduction mechanism: a behavioral approach. Am J Primatol. 1988;16(1):43–50. [DOI] [PubMed] [Google Scholar]
- 40.Judge P, Griffaton N, Fincke A. Conflict management by hamadryas baboons (Papio hamadryas hamadryas) during crowding: a tension‐reduction strategy. Am J Primatol. 2006;68(10):993–1006. [DOI] [PubMed] [Google Scholar]
- 41.Dettmer A, Novak M, Meyer J, Suomi S. Population density-dependent hair cortisol concentrations in rhesus monkeys (Macaca mulatta). Psychoneuroendocrinology. 2014;42:59–67. 10.1016/j.psyneuen.2014.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dettmer AM, Woodward RA, Suomi SJ. Reproductive consequences of a matrilineal overthrow in rhesus monkeys. Am J Primatol. 2015;77(3):346–52. 10.1002/ajp.22350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Novak M, Kinsey J, Jorgensen M, Hazen T. Effects of puzzle feeders on pathological behavior in individually housed rhesus monkeys. Am J Primatol. 1998;46(3):213–27. [DOI] [PubMed] [Google Scholar]
- 44.Altmann J. Observational study of behavior: sampling methods. Behaviour. 1974;49(3):227–66. [DOI] [PubMed] [Google Scholar]
- 45.Neumann C, Duboscq J, Dubuc C, Ginting A, Irwan AM, Agil M, et al. Assessing dominance hierarchies: validation and advantages of progressive evaluation with Elo-rating. Anim Behav. 2011;82(4):911–21. [Google Scholar]
- 46.Albers PC, de Vries H. Elo-rating as a tool in the sequential estimation of dominance strengths. Anim Behav. 2001;61(2):489–95. [Google Scholar]
- 47.Wooddell L, Kaburu SSK, Dettmer A, Suomi S. Elorating as a tool to measure rank changes and dominance stability in semi-free ranging rhesus macaques. Am J Primatol. 2015; 77(S1), 80. [Google Scholar]
- 48.Elo AE. The rating of chessplayers, past and present: Arco Pub.; 1978.
- 49.Dettmer AM, Kaburu SS, Byers KL, Murphy AM, Soneson E, Wooddell LJ, et al. First‐time rhesus monkey mothers, and mothers of sons, preferentially engage in face‐to‐face interactions with their infants. Am J Primatol. 2016; 78 (2): 238–246. 10.1002/ajp.22503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Beehner J, Bergman T, Cheney D, Seyfarth R, Whitten P. The effect of new alpha males on female stress in free-ranging baboons. Anima Behav. 2005;69(5):1211–21. [Google Scholar]
- 51.Sapolsky RM. Cortisol concentrations and the social significance of rank instability among wild baboons. Psychoneuroendocrinology. 1992;17(6):701–9. [DOI] [PubMed] [Google Scholar]
- 52.Sapolsky RM. Endocrine aspects of social instability in the olive baboon (Papio anubis). Am J Primatol. 1983;5(4):365–79. [DOI] [PubMed] [Google Scholar]
- 53.Sapolsky RM. Endocrinology alfresco: psychoendocrine studies of wild baboons. Recent Prog Horm Res. 1993;48:437 [DOI] [PubMed] [Google Scholar]
- 54.Bergman T, Beehner J, Cheney D, Seyfarth R, Whitten P. Correlates of stress in free-ranging male chacma baboons, Papio hamadryas ursinus. Anim Behav. 2005;70(3):703–13. [Google Scholar]
- 55.Creel S, Creel NM, Mills MG, Monfort SL. Rank and reproduction in cooperatively breeding African wild dogs: behavioral and endocrine correlates. Behav Ecol. 1997;8(3):298–306. [Google Scholar]
- 56.Czoty PW, Gould RW, Nader MA. Relationship between social rank and cortisol and testosterone concentrations in male cynomolgus monkeys (Macaca fascicularis). J neuroendocrinol. 2009;21(1):68–76. 10.1111/j.1365-2826.2008.01800.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Fox HE, White SA, Kao MH, Fernald RD. Stress and dominance in a social fish. J Neurosci. 1997;17(16):6463–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Meyer J, Hamel A. Models of stress in nonhuman primates and their relevance for human pschopathlogy and endocrine dysfunction. ILAR J. 2014; 55(2): 347–360. 10.1093/ilar/ilu023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Boccia ML, Reite M, Laudenslager M. On the physiology of grooming in a pigtail macaque. Physiol Behav. 1989;45(3):667–70. [DOI] [PubMed] [Google Scholar]
- 60.Gust DA, Gordon TP, Hambright MK, Wilson ME. Relationship between social factors and pituitary-adrenocortical activity in female rhesus monkeys (Macaca mulatta). Horm Behav. 1993;27(3):318–31. [DOI] [PubMed] [Google Scholar]
- 61.Aureli F, Preston SD, de Waal F. Heart rate responses to social interactions in free-moving rhesus macaques (Macaca mulatta): a pilot study. J Comp Psychol. 1999;113(1):59 [DOI] [PubMed] [Google Scholar]
- 62.Shutt K, MacLarnon A, Heistermann M, Semple S. Grooming in Barbary macaques: better to give than to receive? Biol Lett. 2007;3(3):231–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Engh AL, Beehner JC, Bergman TJ, Whitten PL, Hoffmeier RR, Seyfarth RM, et al. Behavioural and hormonal responses to predation in female chacma baboons (Papio hamadryas ursinus). Proc R Soc B. 2006;273(1587):707–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Qin D, Rizak J, Feng X, Chu X, Yang S, Li C, et al. Social rank and cortisol among female rhesus macaques. (Macaca mulatta). Zool. Res. 2013; 34, E42–49. 10.3724/SP.J.1141.2013.E02E42 [DOI] [PubMed] [Google Scholar]
- 65.Qin D, Rizak J, Chu X, Li Z, Yang S, Lu L, et al. A spontaneous depressive pattern in adult female rhesus macaques. Sci Rep. 2015; 5:11267 10.1038/srep11267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Koren L, Mokady O, Geffen E. Social status and cortisol levels in singing rock hyraxes. Horm Behav. 2008;54(1):212–6. 10.1016/j.yhbeh.2008.02.020 [DOI] [PubMed] [Google Scholar]
- 67.Beisner B, McCowan B. Signaling context modulates social function of silent bared-teeth displays in rhesus macaques (Macaca mulatta). Am J Primatol. 2014; 76(2): 111–121. 10.1002/ajp.22214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Beisner B, Hannibal D, Finn K, Fushing H, McCowan B. Social power, conflict policing, and the role of subordination signals in rhesus macaque society. Am J Phys Anthropol. 2016; 160 (1): 102–112. 10.1002/ajpa.22945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Watts D. A preliminary study of selective visual attention in female mountain gorillas (Gorilla gorilla beringei). Primates; 1998; 39, 71–78. [Google Scholar]
- 70.Pannozzo P, Phillips K, Haas M, Mintz E. Social monitoring reflects dominance relationships in a small captive group of brown capuchin monkeys (Cebus apella). Ethology. 2007; 113(9): 881–888. [Google Scholar]
- 71.Mallow GK. Aggressive behavior during the breeding season of adult female rhesus monkeys (Macaca mulatta). Aggressive Behav. 1980;6(3):217–32. [DOI] [PubMed] [Google Scholar]
- 72.Ruehlmann TE, Bernstein IS, Gordon TP, Balcaen P. Wounding patterns in three species of captive macaques. Am J Primatol. 1988;14(2):125–34. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.