

Abstract
An important regulator of fatty acid oxidation (FAO) is the allosteric inhibition of CPT-1 by malonyl-CoA produced by the enzyme acetyl-CoA carboxylase 2 (ACC2). Initial studies suggested that deletion of Acc2 (Acacb) increased fat oxidation and reduced adipose tissue mass but in an independently generated strain of Acc2 knockout mice we observed increased whole-body and skeletal muscle FAO and a compensatory increase in muscle glycogen stores without changes in glucose tolerance, energy expenditure or fat mass in young mice (12–16 weeks). The aim of the present study was to determine whether there was any effect of age or housing at thermoneutrality (29 °C; which reduces total energy expenditure) on the phenotype of Acc2 knockout mice. At 42–54 weeks of age, male WT and Acc2−/− mice had similar body weight, fat mass, muscle triglyceride content and glucose tolerance. Consistent with younger Acc2−/− mice, aged Acc2−/− mice showed increased whole-body FAO (24 h average respiratory exchange ratio=0.95±0.02 and 0.92±0.02 for WT and Acc2−/− mice respectively, P<0.05) and skeletal muscle glycogen content (+60%, P<0.05) without any detectable change in whole-body energy expenditure. Hyperinsulinaemic–euglycaemic clamp studies revealed no difference in insulin action between groups with similar glucose infusion rates and tissue glucose uptake. Housing Acc2−/− mice at 29 °C did not alter body composition, glucose tolerance or the effects of fat feeding compared with WT mice. These results confirm that manipulation of Acc2 may alter FAO in mice, but this has little impact on body composition or insulin action.
Keywords: insulin resistance, glucose metabolism, metabolism, acetyl CoA carboxylase, energy balance
Introduction
The oxidation of fatty acids in cells requires transport into mitochondria where the enzymes of β-oxidation are located. This transport utilises the enzyme carnitine palmitoyl transferase (CPT)-1 and an important regulator of CPT-1 activity is the metabolite malonyl-CoA, a product of the acetyl-CoA carboxylase (ACC) enzyme. There are two isoforms of ACC. Both Acc1 (Acaca) and Acc2 (Acacb) are expressed in the adipose tissue and liver, but the predominant form in the muscle and heart is Acc2 (Kreuz et al. 2009). Given its localisation on the mitochondrial outer membrane, deletion or inhibition of Acc2 would be expected to decrease the malonyl-CoA pool regulating CPT-1, thereby, enhancing the flux of FA into the mitochondria for oxidation. Accordingly, ACC2 was proposed to be an attractive target for the treatment of obesity on the assumption that an increase in the fatty acid utilisation would reduce fat stores. Consistent with this hypothesis, early studies using Acc2 knockout mice reported a lean phenotype with improved insulin action due to increased whole-body and muscle fatty acid oxidation (FAO) and higher energy expenditure (Abu-Elheiga et al. 2001, Abu-Elheiga et al. 2003, Choi et al. 2007). However, more recent studies using two separate strains of Acc2−/− mice produced using different targeting strategies and generated in different labs did not display lean phenotypes. One study showed that deletion of Acc2 produced the expected increase in muscle and whole-body FAO, but this change in substrate preference was not accompanied by altered energy expenditure, body composition or glucose tolerance in mice under both chow and high-fat diet (HFD) conditions (Hoehn et al. 2010). These Acc2−/− mice did, however, display a robust increase in muscle glycogen content and an elevation in conversion of carbohydrate to lipid, indicative of a re-channelling of excess carbohydrate into other anabolic pathways (Hoehn et al. 2010). Olson et al. (2010) also reported that whole-body and muscle specific deletion of Acc2−/− mice had limited influence on metabolic phenotype and body composition in mice.
FAO theoretically produces more ATP per carbon oxidised to CO2 than glucose but consumes 6–9% more oxygen for this process (Turner et al. 2014). This suggests that producing the same amount of ATP from FAO could be less efficient than carbohydrate oxidation in terms of ATP produced per oxygen consumed. The first aim in this study was to determine whether older Acc2−/− mice displayed alterations in body composition and insulin action, due to chronic changes in FAO over a more prolonged timeframe than our previous studies. Furthermore, because metabolic rate decreases with increasing ambient temperature (Abreu-Vieira et al. 2015) and housing temperature is known to have a marked effect on the whole-body substrate oxidation and metabolic profile (Liu et al. 2003, Castillo et al. 2011), our second aim was to determine whether the shift in fuel preference to fatty acids in Acc2−/− mice would alter the response to a chow and HFD when mice were housed under conditions of thermoneutrality. Materials and methods All surgical and experimental procedures performed were approved by the Garvan Institute/St Vincent’s Hospital Animal Ethics Committee and were in accordance with the National Health and Medical Research Council of Australia’s guidelines on animal experimentation.
Materials and methods
All surgical and experimental procedures performed were approved by the Garvan Institute/St Vincent’s Hospital Animal Ethics Committee and were in accordance with the National Health and Medical Research Council of Australia’s guidelines on animal experimentation.
Animals
Acc2−/− mice were generated as described previously (Hoehn et al. 2010). Acc2−/− and WT littermates were communally housed in temperature-controlled (22±0.5 °C standard housing or 29±0.5 °C for thermoneutral studies) and light-controlled (12 h light:12 h darkness cycle) rooms. They were fed ad libitum a standard chow diet (21% of calories from protein, 8% from fat, 71% from carbohydrate; Rat Maintenance Diet; Gordon Specialty Feeds, Sydney, NSW, Australia) or a HFD made in-house (45% of calories from fat, 20% from protein and 35% from carbohydrates) based on rodent diet no. D12451 (Research Diets, New Brunswick, NJ, USA). Mice were studied at ∼18 or 50 weeks of age. For thermoneutral studies, animals were acclimatised to the warmer room for 2–4 weeks prior to dietary intervention (4 weeks).
Respirometry
Oxygen consumption rate (VO2) and respiratory exchange ratio (RER) were measured under a consistent environmental temperature (22 °C) using an indirect calorimetry system (Oxymax series, Columbus Instruments, Columbus, OH, USA). The airflow was 0.6 l/min and measurements were recorded after a 24 h acclimatisation period in the metabolic chamber. VO2 and CO2 were measured in individual mice at 27 min intervals over a 24 h period.
Glucose tolerance test and fat mass determination
Mice were fasted for 5 h and were then injected intraperitoneally with glucose (2 g/kg) and blood glucose levels were monitored over time (ipGTT). Total fat mass was determined by dual X-ray absorptiometry using a PIXImus small-animal densitometer and associated software (PIXImus II; GE Medical Systems, Madison, WI, USA).
Surgical procedures
Older mice were anaesthetised with isoflurane anaesthesia (4% induction, 1.5–2% maintenance) for insertion of catheters into the left carotid artery and right jugular vein. Free catheter ends were tunnelled under the skin, externalised at the neck and sealed. Mice were then singly housed and monitored daily. Catheters were flushed every 1–2 days with heparinised saline to maintain patency.
Hyperinsulinaemic–euglycaemic clamp
Approximately 4–7 days post-surgery, and after a ∼5 h fast, a hyperinsulinaemic–euglycaemic clamp was conducted. Mice were conscious, unrestrained and were not handled during the procedure to minimise stress. At −90 min, a primed (5 μCi) continuous infusion (0.05 μCi/min) of [3,3H]glucose (PerkinElmer, Glen Waverley, Victoria, Australia) was commenced. At −30, −20, −10 and 0 samples were collected for basal glucose turnover (Rd) and glucose and insulin levels (−30 and 0). At time 0, the rate of [3,3H]glucose was increased to (0.1 μCi/min) and primed to (16 μU/kg), continuous (4 μU/kg per min) infusion of insulin commenced (Actrapid, Novo Nordisk, Copenhagen, Denmark). Glucose (25%) was infused at a variable rate to maintain glycaemia at ∼8 mM. Once blood glucose was stable, four sequential samples were taken for glucose turnover and insulin determination. A bolus of 2[14C]deoxyglucose (13 μCi; PerkinElmer) was then administered and blood sampled at 2, 5, 10, 15, 20 and 30 min for measurement of glucose uptake into specific tissues. Animals were then euthanised and organs removed, snap frozen in liquid nitrogen and stored at −80 °C for further analysis. This protocol is based on Ayala et al. (2006) and Charbonneau & Marette (2010).
Analytical methods
Blood and plasma glucose levels were determined using an Accu-check II glucometer (Roche Diagnostics, Castle Hill, NSW, Australia). Insulin levels were determined using a sensitive rat RIA kit (Millipore, Missouri, USA). Triglyceride and glycogen content were measured in tissues as described previously (Bruce et al. 2007, Hoy et al. 2009). The rate of basal and clamp glucose disappearance (Rd) was determined using steady-state equations. Clamp hepatic glucose output was determined by subtracting the glucose infusion rate (GIR) from Rd. For determination of tissue glucose uptake, tissue samples were homogenised in water. After centrifugation, one aliquot was counted to determine total 2[14C]DG and 2[14C]DGP radioactivity. Another aliquot was run through a column of AG 1-X8 resin (Bio-Rad Laboratories) which retains 2[14C]DGP and elutant containing 2[14C]DG was counted. 2[14C]DGPtissue is the difference between total 14C and 2[14C]DG measures. Using this 2[14C]DGPtissue, glucose uptake into tissues determined using the following equation:
where 2[14C]DGPtissue is the 2[14C]DGP radioactivity in the muscle (in dpm/g), AUC 2[14C]DGplasma is the area under the plasma 2[14C]DG disappearance curve (in dpm/min per ml), and (arterial glucose) is the average blood glucose (in mmol/l)(Ayala et al. 2007).
Western blotting was conducted as previously described (Brandon et al. 2015). ACC antibody was from Cell Signaling Technology (Danvers, MA, USA).
Statistical analysis
Data are expressed as means±S.E.M. Differences between groups were determined by t-test or two-way ANOVA as required. If the two-way ANOVA reached significance, a Bonferroni’s post hoc test was conducted. All statistical analysis was done using GraphPad Prism (Version 6 for Windows, GraphPad Software, San Diego, CA, USA). Significance was set at P≤0.05.
Results
Effect of Acc2 deletion on body weight and fat mass of 1 year old mice kept at 22 °C
Successful Acc2 deletion was determined via western blot in the gastrocnemius muscle (Fig. 1A). Since Acc2 is the predominant isoform in the muscle, there is only one band observed in the WT and no bands in Acc2−/− mice.
Figure 1.
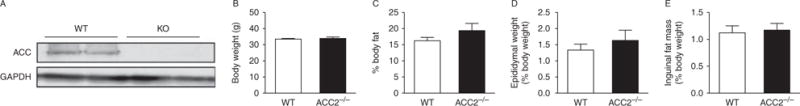
Body weight and fat mass are similar between WT and Acc2−/− mice. (A) Representative western blot demonstrating deletion of Acc2 in gastrocnemius muscle. (B) Body weight, (C) percent fat mass as determined by DEXA and individual weights for (D) epididymal and (E) inguinal fat pads in WT and Acc−/− mice. Data are expressed as mean±S.E.M. n=8–11 animals per group. Animals were 42–52 weeks of age.
Body weights were similar between Acc2−/− and WT animals (Fig. 1B). Total fat mass, as determined by DEXA, was not different between groups (Fig. 1C) and there was no difference in the weight of individual fat pads between genotypes (Fig. 1D and E).
Whole-body and muscle parameters
Whole-body calorimetry showed that, similar to younger animals (Hoehn et al. 2010), older Acc2−/− mice showed increased whole-body FAO (24 h average RER=0.95±0.02 and 0.92±0.02 for WT and Acc2−/− mice respectively, P<0.05; Fig. 2A and B). This occurred without observable changes in energy expenditure (Fig. 2C and D) and without differences in glucose tolerance (Fig. 2E). There were no differences between the groups in muscle triglyceride content (Fig. 3A). However, there was an increase in muscle glycogen content in the Acc2−/− group (Fig. 3B).
Figure 2.
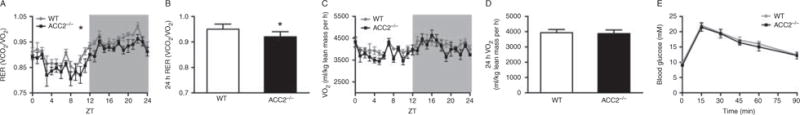
Deletion of Acc2 increased whole-body fatty acid oxidation without altering energy expenditure or glucose tolerance. (A) Respiratory exchange ratio (RER), (B) 24h average RER, (C) oxygen consumption (VO2), (D) 24h average VO2, (E) glucose tolerance in WT and ACC−/− mice. Data are expressed as mean±S.E.M. *P<0.05 vs WT (average 24h values). n=8–11 animals per group. Animals were 42–52 weeks of age.
Figure 3.
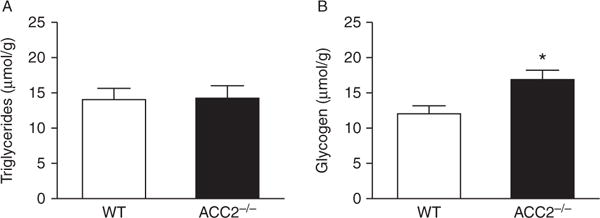
Muscle of Acc2−/− mice has similar triglyceride and increased glycogen levels. (A) Triglyceride and (B) glycogen content in the gastrocnemius muscle of WT and Acc2−/− mice. Data are expressed as mean±S.E.M. *P<0.05 vs WT. n=8–11 animals per group. Animals were 42–52 weeks of age.
Hyperinsulinaemic–euglycaemic clamp studies
Under basal conditions, WT and Acc2−/− mice showed similar blood glucose and plasma insulin levels (Fig. 4A and B). There was also no difference in basal whole-body glucose disappearance (Fig. 4D). Under insulin-stimulated conditions, there was no difference between the groups in GIR (Fig. 4C), whole-body glucose disappearance, hepatic glucose output (Fig. 4E) or glucose uptake into various tissues (Fig. 4F), indicating similar whole-body and tissue insulin sensitivity.
Figure 4.
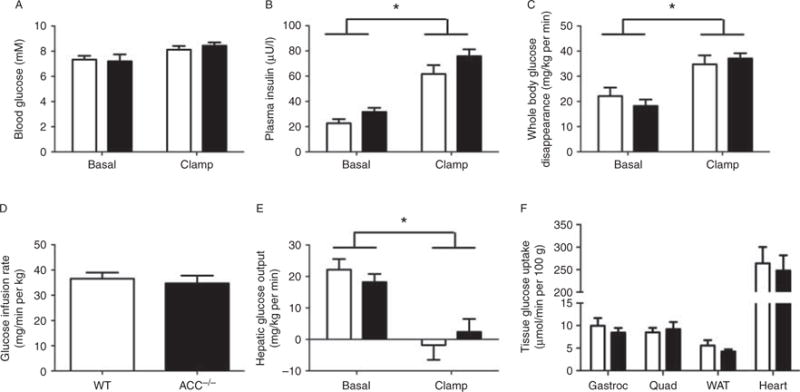
Hyperinsulinaemic–euglycaemic clamp studies showing Acc2−/− mice have similar insulin sensitivity to WT mice. (A) Blood glucose and (B) plasma insulin levels in the basal and insulin-stimulated conditions. (C) Glucose infusion rate. (D) Whole-body glucose disappearance and (E) hepatic glucose output in the basal and insulin-stimulated conditions. (F) Glucose uptake into muscle (quadriceps, gastrocnemius), white adipose tissue (WAT; epididymal) and heart of WT and Acc2−/− mice. Data are expressed as mean±S.E.M. *P<0.05 vs basal state. n=6–8 animals per group. Animals were 42–52 weeks of age.
Effect housing at 29 °C and HFD on body weight, fat mass and glucose tolerance of Acc2 knockout mice
There was no observable difference between WT and Acc2−/− mice housed at 29 °C for any of the parameters measured. Animals fed a HFD were heavier when compared with the chow-fed animals irrespective of genotype (Fig. 5A). This was due to a higher fat mass in the HFD group (Fig. 5B, C and D). While there was a worsening of glucose tolerance in animals fed a HFD, this was not affected by genotype (Fig. 5E and F). Acc2−/− animals housed at thermoneutrality did exhibit the increased glycogen content in the muscle demonstrated in mice housed at 22 8C, and this was unaffected by diet (Fig. 6A). There was no difference in triglyceride content between genotypes kept at 29 °C, but there was a clear increase in both genotypes when fed a HFD (Fig. 6B).
Figure 5.
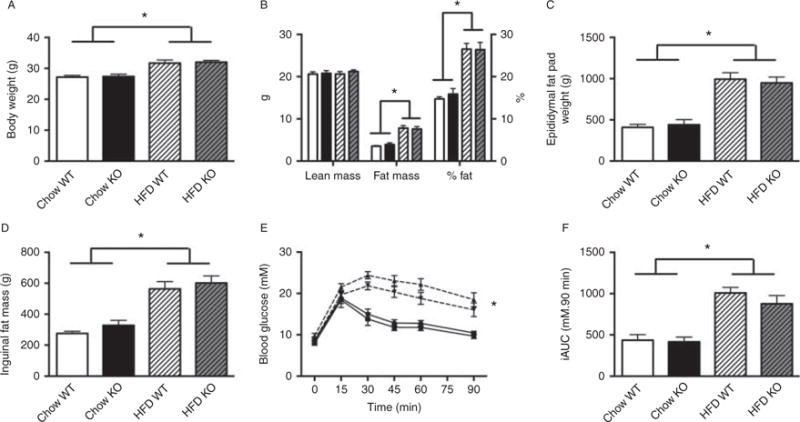
Housing Acc2−/− mice at thermoneutral temperatures (29 °C) does not alter body weight, body composition or glucose tolerance on chow or high-fat diet (HFD). (A) Body weight, (B) body composition as determined by DEXA, (C) epididymal and (D) inguinal fat mass weights, (E) ipGTT curve and (F) iAUC in chow and HFD mice. White bars, chow WT; black bars, chow Acc2−/−; white hatched bars, HFD WT; grey hatched bars, HFD Acc2−/−. *P<0.05 effect of diet. n=5–11 animals per group. Animals were 16–20 weeks of age.
Figure 6.
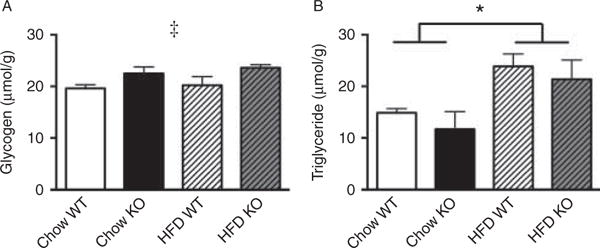
Acc2−/− mice have similar muscle triglyceride content that increases equally after a high-fat diet, while there is an increased muscle glycogen content in Acc2−/− mice that is unaffected by diet. (A) Glycogen and (B) triglyceride content in the gastrocnemius muscle of WT and Acc2−/− mice. Data are expressed as mean±S.E.M. ‡P<0.05 effect of genotype; *P<0.05 effect of diet. n=5–11 animals per group. Animals were 16–20 weeks of age.
Discussion
The glucose-fatty acid cycle, first reported more than 50 years ago by Randle et al. (1963), describes the competition between glucose and fatty acids for oxidation in muscle and adipose tissue to provide energy to support all cellular processes (Hue & Taegtmeyer 2009). This idea proposes that when there is excess of one type of fuel, or when one fuel substrate is preferentially oxidised, then the other is channelled into storage pathways. Our findings in Acc2−/− mice, both in the present study and in our previous work (Hoehn et al. 2010), are completely consistent with this model, as the enhanced oxidation of FA without any concomitant increase in energy expenditure leads to diversion of other potential energy substrates like carbohydrate to other pathways including storage as glycogen in muscle.
Because fatty acids are a more reduced form of substrate than carbohydrate or amino acids, the complete oxidation of fatty acids theoretically yields more ATP per carbon atom than oxidation of glucose carbons. However, more oxygen atoms are consumed for complete oxidation of fatty acid carbons (Turner et al. 2014) leading to the idea that increased FAO is accompanied by increased energy expenditure (as measured by oxygen consumption). Therefore, preferential whole body oxidation of FA has been hypothesised to impact on the metabolic and body composition phenotype of animals. Despite initial reports that this was the case in one line of Acc2−/− mice, we, and others have firmly established that preferentially promoting the oxidation of lipid substrates through deletion of Acc2 has little effect on body composition and insulin action (Hoehn et al. 2010, Olson et al. 2010). However, because our previous findings were in younger animals, it was possible that chronic changes in FAO over the longer term may have some impact on whole-body energy metabolism that was not evident in younger mice. In the present study, examining 9- to 12-month-old Acc2−/− mice, increases in whole-body FAO and glycogen deposition in muscle were still present, without any significant change in energy expenditure, body composition or insulin sensitivity as measured by the euglycaemic–hyperinsulinaemic clamp. Therefore, these results add further weight to our previous conclusions that increasing FAO alone, even over an extended period of time, is insufficient to alter body weight and fat mass, but instead leads to the channelling of carbohydrate into storage pathways (Hoehn et al. 2010).
The marked discrepancies between different lines of Acc2−/− mouse have been documented in a range of published articles. The first Acc2−/− mice generated were reported to have increased FAO, higher energy expenditure and better insulin sensitivity (determined by clamp), despite being hyperphagic (Abu-Elheiga et al. 2001, 2003, Choi et al. 2007). These findings have not been repeated in other studies (Hoehn et al. 2010, Olson et al. 2010) and these discrepancies may be related to differences in many factors, including variations in gene-targeting strategies (Hoehn et al. 2012). That is, Acc2−/− mice generated by Wakil and colleagues targeted a different exon to other groups and their mice are also engineered to overexpress hypoxanthine-guanine phosphoribosyltransferase (Hprt) (presumably in every cell of the animal), an important enzyme in the purine salvage pathway, which is not present in WT mice (Abu-Elheiga et al. 2001). Although there is little information about the effects of over-expression of Hprt, the deletion or mutation of this gene in humans causes Lesch–Nyhan syndrome, an X-linked recessive disorder characterised by uric acid overproduction and neurological manifestations (Torres & Puig 2007). However, in addition to the Acc2−/− mouse line used for the current studies, Olson et al. (2010) reported no alterations in body weight, fat mass or glucose homeostasis in different lines of whole-body and muscle-specific Acc2 knockouts or if the Acc2−/− were crossed with obese leptin-deficient mice (ob/ob).
There are some recent data showing that treatment of obese, insulin-resistant db/db mice for 70 days with a selective inhibitor of ACC2 can lower circulating glucose and triglycerides and improve glucose tolerance and glucose uptake in muscle (Glund et al. 2012). However, the inhibitor had no effect on body weight or fat mass and neither energy expenditure nor whole-body fat oxidation were determined in this study. Therefore, it is not clear whether the beneficial effects of this inhibitor were due solely to increased FAO relieving lipid-induced insulin resistance in muscle or whether a similar improvement would be seen if the ACC2 inhibitor was delivered to HFD-fed animals.
Housing at, or near, thermoneutral temperature for mice should be a consideration when assessing phenotypes, especially in relation to potential differences in energy metabolism and obesity. Standard housing conditions for mice vary between facilities but are normally between 20 and 25 °C, the thermoneutral conditions for lightly clothed humans. Mice, however, have a thermoneutral zone that is closer to 30 °C (Maloney et al. 2014). This means that under most housing conditions, mice are subject to chronic cold stress. This is an important aspect to consider since it has been shown that this temperature difference can change parameters of the cardiovascular system (e.g. heart rate), response to infections and metabolic rate (reviewed in Overton (2010)). Interestingly, there are examples in the literature where ambient temperature has a significant effect on phenotype. The first example is the UCP1 deficient mouse, which was reported to be resistant to a HFD weight gain when housed at standard temperatures (20 °C), but when the temperature was raised to 27 °C, these animals showed accelerated body weight gain (Liu et al. 2003). Another example is the type 2 deiodinase (Dio2) knockout mouse, which had better glucose tolerance than WT animals on a chow diet at standard housing temperature but this was lost when animals were housed at 30 °C. In fact, Dio2−/− mice were more glucose intolerant when placed on a HFD at this increased temperature (Castillo et al. 2011). In the present study, Acc2−/− mice housed at 29 °C displayed no differences compared with WT littermates with regard to body weight, fat mass or glucose tolerance on a chow diet and all these parameters changed similarly when both genotypes were placed on a HFD.
In summary, this study provides further evidence that deletion of Acc2, although increasing whole-body FAO, does not alter energy expenditure, fat mass or insulin sensitivity even when genotype-dependent differences in substrate oxidation have been present for an extended period of time or animals are housed under conditions of thermoneutrality. These studies strongly support the view that switching between different substrates for energy metabolism, without any alteration in total fuel oxidation, is insufficient to alter body weight and that changes in total energy expenditure are required to drive weight loss.
Acknowledgments
The authors would like to thank the staff of the Garvan Biological Testing Facility.
Funding
This work was supported by a National Health and Medical Research Council (NHMRC) Program grant (535921) to D E J, E W K and G J C. D E J and G J C are currently supported by NHMRC Research Fellowships and NT is supported by an Australian Research Council (ARC) Future Fellowship.
Footnotes
Declaration of interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Author contribution statement
A E B, N T, G J C and E W K conceived and designed the experiments; A E B, N T, E S and S J L performed the experiments; A E B, N T and G J C analysed the data; A E B, N T, E W K, D E J, K L H and G J C wrote the paper.
References
- Abreu-Vieira G, Xiao C, Gavrilova O, Reitman ML. Integration of body temperature into the analysis of energy expenditure in the mouse. Molecular Metabolism. 2015;4:461–470. doi: 10.1016/j.molmet.2015.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abu-Elheiga L, Matzuk MM, Abo-Hashema KA, Wakil SJ. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-CoA carboxylase 2. Science. 2001;291:2613–2616. doi: 10.1126/science.1056843. [DOI] [PubMed] [Google Scholar]
- Abu-Elheiga L, Oh W, Kordari P, Wakil SJ. Acetyl-CoA carboxylase 2 mutant mice are protected against obesity and diabetes induced by high-fat/high-carbohydrate diets. PNAS. 2003;100:10207–10212. doi: 10.1073/pnas.1733877100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayala JE, Bracy DP, McGuinness OP, Wasserman DH. Considerations in the design of hyperinsulinemic–euglycemic clamps in the conscious mouse. Diabetes. 2006;55:390–397. doi: 10.2337/diabetes.55.02.06.db05-0686. [DOI] [PubMed] [Google Scholar]
- Ayala JE, Bracy DP, Julien BM, Rottman JN, Fueger PT, Wasserman DH. Chronic treatment with sildenafil improves energy balance and insulin action in high fat-fed conscious mice. Diabetes. 2007;56:1025–1033. doi: 10.2337/db06-0883. [DOI] [PubMed] [Google Scholar]
- Brandon AE, Tid-Ang J, Wright LE, Stuart E, Suryana E, Bentley N, Turner N, Cooney GJ, Ruderman NB, Kraegen EW. Overexpression of SIRT1 in rat skeletal muscle does not alter glucose induced insulin resistance. PLoS ONE. 2015;10:e0121959. doi: 10.1371/journal.pone.0121959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruce CR, Brolin C, Turner N, Cleasby ME, van der Leij FR, Cooney GJ, Kraegen EW. Overexpression of carnitine palmitoyltransferase I in skeletal muscle in vivo increases fatty acid oxidation and reduces triacylglycerol esterification. American Journal of Physiology Endocrinology and Metabolism. 2007;292:E1231–E1237. doi: 10.1152/ajpendo.00561.2006. [DOI] [PubMed] [Google Scholar]
- Castillo M, Hall JA, Correa-Medina M, Ueta C, Kang HW, Cohen DE, Bianco AC. Disruption of thyroid hormone activation in type 2 deiodinase knockout mice causes obesity with glucose intolerance and liver steatosis only at thermoneutrality. Diabetes. 2011;60:1082–1089. doi: 10.2337/db10-0758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charbonneau A, Marette A. Inducible nitric oxide synthase induction underlies lipid-induced hepatic insulin resistance in mice: potential role of tyrosine nitration of insulin signaling proteins. Diabetes. 2010;59:861–871. doi: 10.2337/db09-1238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi CS, Savage DB, Abu-Elheiga L, Liu ZX, Kim S, Kulkarni A, Distefano A, Hwang YJ, Reznick RM, Codella R, et al. Continuous fat oxidation in acetyl-CoA carboxylase 2 knockout mice increases total energy expenditure, reduces fat mass, and improves insulin sensitivity. PNAS. 2007;104:16480–16485. doi: 10.1073/pnas.0706794104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glund S, Schoelch C, Thomas L, Niessen HG, Stiller D, Roth GJ, Neubauer H. Inhibition of acetyl-CoA carboxylase 2 enhances skeletal muscle fatty acid oxidation and improves whole-body glucose homeostasis in db/db mice. Diabetologia. 2012;55:2044–2053. doi: 10.1007/s00125-012-2554-9. [DOI] [PubMed] [Google Scholar]
- Hoehn KL, Turner N, Swarbrick MM, Wilks D, Preston E, Phua Y, Joshi H, Furler SM, Larance M, Hegarty BD, et al. Acute or chronic upregulation of mitochondrial fatty acid oxidation has no net effect on whole-body energy expenditure or adiposity. Cell Metabolism. 2010;11:70–76. doi: 10.1016/j.cmet.2009.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoehn KL, Turner N, Cooney GJ, James DE. Phenotypic discrepancies in acetyl-CoA carboxylase 2-deficient mice. Journal of Biological Chemistry. 2012;287:15801. doi: 10.1074/jbc.O112.356915. author reply 15802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoy AJ, Brandon AE, Turner N, Watt MJ, Bruce CR, Cooney GJ, Kraegen EW. Lipid and insulin infusion-induced skeletal muscle insulin resistance is likely due to metabolic feedback and not changes in IRS-1, Akt or AS160 phosphorylation. American Journal of Physiology. Endocrinology and Metabolism. 2009;297:E67–E75. doi: 10.1152/ajpendo.90945.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hue L, Taegtmeyer H. The Randle cycle revisited: a new head for an old hat. American Journal of Physiology. Endocrinology and Metabolism. 2009;297:E578–E591. doi: 10.1152/ajpendo.00093.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreuz S, Schoelch C, Thomas L, Rist W, Rippmann JF, Neubauer H. Acetyl-CoA carboxylases 1 and 2 show distinct expression patterns in rats and humans and alterations in obesity and diabetes. Diabetes/Metabolism Research and Reviews. 2009;25:577–586. doi: 10.1002/dmrr.997. [DOI] [PubMed] [Google Scholar]
- Liu X, Rossmeisl M, McClaine J, Riachi M, Harper ME, Kozak LP. Paradoxical resistance to diet-induced obesity in UCP1-deficient mice. Journal of Clinical Investigation. 2003;111:399–407. doi: 10.1172/JCI200315737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maloney SK, Fuller A, Mitchell D, Gordon C, Overton JM. Translating animal model research: does it matter that our rodents are cold? Physiology. 2014;29:413–420. doi: 10.1152/physiol.00029.2014. [DOI] [PubMed] [Google Scholar]
- Olson DP, Pulinilkunnil T, Cline GW, Shulman GI, Lowell BB. Gene knockout of Acc2 has little effect on body weight, fat mass, or food intake. PNAS. 2010;107:7598–7603. doi: 10.1073/pnas.0913492107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overton JM. Phenotyping small animals as models for the human metabolic syndrome: thermoneutrality matters. International Journal of Obesity. 2010;34(Suppl 2):S53–S58. doi: 10.1038/ijo.2010.240. [DOI] [PubMed] [Google Scholar]
- Randle PJ, Garland PB, Hales CN, Newsholme EA. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet. 1963;1:785–789. doi: 10.1016/S0140-6736(63)91500-9. [DOI] [PubMed] [Google Scholar]
- Torres RJ, Puig JG. Hypoxanthine-guanine phosophoribosyl-transferase (HPRT) deficiency: Lesch-Nyhan syndrome. Orphanet Journal of Rare Diseases. 2007;2:48. doi: 10.1186/1750-1172-2-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner N, Cooney GJ, Kraegen EW, Bruce CR. Fatty acid metabolism, energy expenditure and insulin resistance in muscle. Journal of Endocrinology. 2014;220:T61–T79. doi: 10.1530/JOE-13-0397. [DOI] [PubMed] [Google Scholar]