

Abstract
Streptophyte green algae, ancestors of Embryophytes, occur frequently in terrestrial habitats being exposed to high light intensities, water scarcity and potentially toxic metal cations under acidic conditions. The filamentous Zygogonium ericetorum synthesizes a purple vacuolar ferrous pigment, which is lost after aplanospore formation. However, it is unknown whether this cellular reorganization also removes excessive iron from the protoplast and how Z. ericetorum copes with high concentrations of aluminium. Here we show that aplanospore formation shifts iron into the extracellular space of the algal filament. Upon germination of aplanospores, aluminium is bound in the parental cell wall. Both processes reduce iron and aluminium in unpigmented filaments. Comparison of the photosynthetic oxygen production in response to light and temperature gradients in two different Z. ericetorum strains from an Austrian alpine and a Scottish highland habitat revealed lower values in the latter strain. In contrast, the Scottish strain showed a higher optimum quantum yield of PSII during desiccation stress followed by rehydration. Furthermore, pigmented filaments of both strains exhibited a higher light and temperature dependent oxygen production when compared to the unpigmented phenotype. Our results demonstrate a high metal tolerance of Z. ericetorum, which is crucial for surviving in acidic terrestrial habitats.
Keywords: aluminium, cell wall, green algae, iron, photosynthesis, zinc
The green alga Zygogonium ericetorum shows high iron and aluminium tolerance, and these metals are excluded from the cells by special protection mechanisms, allowing high photosynthetic performance in terrestrial habitats.
INTRODUCTION
Based on their primary adaptation to freshwater, distinct streptophyte green algae belonging to the Zygnematophyceae moved to the limnetic transition zone and further to dry land, which gave rise to the evolution of land plants in the Mid-Ordovician period (Becker and Marin 2009; Wickett et al.2014). Today, Zygnematophyceae occur worldwide in limnetic, hydro- and aero-terrestrial habitats (e.g. Škaloud 2009; Stancheva, Sheath and Hall 2012; Pichrtová, Hajek and Elster 2014a), representing a gradient of decreasing water availability, increasing light intensity, temperature fluctuation as well as metal toxicity due to their increased abundance in more acidic environments. Investigating green algae's physiological performance and cell morphology in these different habitats allows tracing adaptations, which were necessary for changing the primary limnetic to terrestrial live-style (Herburger and Holzinger 2015). Several studies focused on the photosynthetic performance of Zygnematophyceae under light, temperature, osmotic, water or nutrient starvation stress, while either natural field populations (Holzinger, Roleda and Lütz 2009; Aigner et al.2013; Pichrtová, Hajek and Elster 2014a), cultured algae (Kaplan et al.2013; Pichrtová et al.2013; Pichrtová, Kulichová and Holzinger 2014b; Herburger, Lewis and Holzinger 2015) or both (Hawes 1990; Stamenković and Hanelt 2013) were investigated. Many of these green algae use light very efficiently, indicated by a high photosynthetic performance under dim conditions, low light compensation and saturation points and lacking strong photoinhibition even far beyond light saturation. This high photophysiological plasticity coincides with the hydro- and aero-terrestrial lifestyle, since it was also found in several other streptophyte algae (e.g. Klebsormidiophyceae) from similar habitats (Kaplan et al.2012; Karsten, Herburger and Holzinger 2014; Herburger, Karsten and Holzinger 2016). Changing light conditions often go along with desiccation stress and cellular water loss strongly suppresses the photosynthetic performance of green algae in the vegetative state (Holzinger and Karsten 2013). It was shown recently that specialized resistant-cells (pre-akinetes or akinetes) formed by Zygnema after mild long-term desiccation stress (Pichrtová, Hajek and Elster 2014a) or nutrient starvation (Herburger, Lewis and Holzinger 2015) provide resistance against short-term desiccation stress. However, pre-akinetes exhibit lower maximum photosynthetic rates when compared with vegetative cells (Herburger, Lewis and Holzinger 2015), making the transformation into these resistant cells a trade-off between desiccation tolerance and a high metabolic activity. Another common response of Zygnematophyceae to abiotic stress is the accumulation of phenolic compounds (Remias et al.2011; Aigner et al.2013; Pichrtová et al.2013). In natural habitats, green cells of the filamentous streptophyte green algae Z. ericetorum (‘green morph’) accumulate high amounts of phenolics and turn purple (‘purple morph’; Alston 1958). The purple pigment revealed as a glycosylated and highly branched phenolic compound which forms complexes with Fe3+ (Newsome and van Breemen 2012), while a ferric (gallate)2 complex causes the colour (Newsome, Murphy and van Breemen 2013). Iron is an essential heavy metal trace element for all plants, as it is a major constituent of the cellular redox system including cytochromes, catalase, peroxidase as well as ferredoxin and superoxide dismutase (Karlusich, Lodeyro and Carrillo 2014). On the other hand, due to low pH values, excessive soluble iron (Fe2+) can cause oxidative stress by generating reactive oxygen species through the Fenton reaction (e.g. hydroxyl radicals; Cadenas 1989), which are a source of damage to lipids, proteins and nucleic acids (Kranner and Birtić 2005). Therefore, iron metabolism has to be adapted to the cellular demands to maintain a sufficient redox system but to avoid cellular damage because of iron induced oxidative stress (Kampfenkel, Van Montagu and Inzé 1995; Finney and O'Halloran 2003). Important regulators of iron homeostasis in plants are ferritins (Ravet et al.2009), a superfamily of iron storage proteins (Briat et al.2010). In the green algae Chlamydomonas reinhardtii, ferritins are involved in restoring the PSI level after iron starvation followed by iron repletion and protecting the photosynthetic apparatus under photo-oxidative stress during highlight conditions or iron overflow (Busch et al.2008). Ferritin-iron complexes of plants are localized in the plastid stroma (Waldo et al.1995) and mitochondria (Zancani et al.2004). In contrast, the ferrous phenolic compound responsible for the purple pigmentation in Z. ericetorum cells is stored in vacuoles due to its hydrophilic nature (Alston 1958). It has been assumed recently that Zygogonium cells can exclude excessive iron from the protoplast into the extracellular space by forming unpigmented aplanospores, while these spores can germinate into new green vegetative cells (Stancheva et al.2014). Besides iron, low pH values (<5.5), which are typical for habitats of Z. ericetorum, can also solubilize other metals into their potentially toxic cationic form (Foy, Chaney and White 1978). Zinc is amongst the most widespread metals in soils, where it is strongly adsorbed until the pH value decreases (Mertens and Smolders 2013), which allows uptake into plant cells (Klimmek et al.2001). Like iron, zinc is an essential trace metal for plants, but high levels of zinc increase the production of harmful reactive oxygen species (Prasad, Saradhi and Sharmila 1999). Another limiting factor of plant growth is aluminium (Ma, Ryan and Delhaize 2001). In contrast to iron and zinc, aluminium is not essential for plants (Rengel 1992), but its harmful effects occur at even micromolar concentrations (Ma, Ryan and Delhaize 2001). Due to their toxicity, coping with changing and high concentrations of these metal ions is crucial in acidic habitats with strongly fluctuating water availability: Low pH values make metals bioavailable and the evaporation of water can increase the concentration of these ions at even short time scales. However, little information is available on how the ancestors of land plants coped with such stress under natural conditions.
This study investigates cellular and photophysiological responses of Z. ericetorum to abiotic stresses prevailing in limnetic transitional habitats, with a focus on resistance against metal toxicity. Therefore, purple Z. ericetorum filaments were collected from two different natural field populations in Austria (high alpine habitat: ∼2300 m a.s.l.) and Scotland (forest zone layer: 45 m a.s.l.). The role of cellular reorganization processes (i.e. accumulation of purple pigmentation or formation of aplanospores) to cope with iron, aluminium and zinc stress as well as the photosynthetic response to light, temperature and desiccation gradients was investigated in laboratory experiments. We hypothesize that (i) iron becomes excluded from the protoplasts during aplanospore formation to avoid toxic concentration within the protoplast and that (ii) Z. ericetorum is generally well adapted to metal rich environments by either complexing or avoiding the uptake of metals into cells. Furthermore, we tested if (iii) the two Z. ericetorum isolates obtained from two different habitats exhibit distinct responses to light, temperature and desiccation gradients, which would imply a high adaptive capacity and phenotypic plasticity of this species. Finally, we investigated whether (iv) the ‘purple morphs’ of both Z. ericetorum isolates show higher photosynthetic oxygen productions in response to increasing light and temperature when compared with the respective ‘green morphs’.
MATERIALS AND METHODS
Sampling localities, algal origin and experimental setup
Purple Z. ericetorum filaments (Z. ericetorum AUT ‘purple morph’ = Zygogonium AUT-p) were collected in August 2013 on Mt Schönwieskopf (near Obergurgl, Tyrol, Austria). This site was previously sampled by Holzinger, Tschaikner and Remias (2010), Aigner et al. (2013) and Stancheva et al. (2014, 2016). Purple algal filaments submersed in soil water and original soil water were collected separately in polyethylene vials and transferred immediately to the Institute of Botany at the University of Innsbruck. Physiological measurements and microscopic examinations (see following paragraphs) of Zygogonium AUT-p were performed in filtered soil water and started the day after collection. The same day, subsamples were washed, transferred to 100 mL Bold's Basal Medium (BBM, pH = 6.8; Bischoff and Bold 1963) in 250 mL Erlenmeyer flasks. They were cultured in a dark/light regime of 16:8 h in a Percival PGC 6L thermostat (Percival Scientific, Perry, GA, USA) at 20°C and ∼33 μmol photons m−2 s−1 (adjusted with Osram Daylight Lumilux Cool White lamps L36W/840; Osram. Munich, Germany). In the dark period, temperature was reduced to 14.5°C. Algal filaments turned green (i.e. Z. ericetorum AUT ‘green morph’ = Zygogonium AUT-g) within 10–12 days. All physiological measurements and microscopic examinations of Zygogonium AUT-g started two weeks after subsampling. A second purple sample of Z. ericetorum (Z. ericetorum SCOT ‘purple morph’ = Zygogonium SCOT-p) was collected in August 2014 near Glencoe (Lochaber, Scotland; coordinates in Table 1). Filaments were stored in original soil water obtained from the centre of the population. They were transferred to the laboratory in Innsbruck within one day and treated the same like the samples from Austria (see above) to establish Z. ericetorum SCOT ‘green morph’ (= Zygogonium SCOT-g) within ∼12 days and perform physiological and microscopic experiments. The methods applied to Zygogonium AUT-p, Zygogonium AUT-g, Zygogonium SCOT-p and Zygogonium SCOT-g are summarized in Fig. S1 (Supporting Information).
Table 1.
Origin of Zygogonium AUT-p and Zygogonium SCOT-p.
| Isolate | Habitat | Metrological data |
|---|---|---|
| Zygogonium ericetorum AUT ‘purple morph’ (Zygogonium AUT-p) | Streamlet on Mt Schönwieskopf (∼2300 m a.s.l.), Tyrol, Austria (46.84804°N 11.01539°E); isolated on 18 August 2013; water depth 5–20 cm, open water surface; air temperature: 8.5°C | Minimum air temperature: −8°C to –6°C Maximum air temperature: −3°C to 12°C Monthly rainfall days: 11–18 Monthly precipitation: 36.3–120.3 mm Annual rainfall: 839 mm |
| Z. ericetorum SCOT ‘purple morph’ (Zygogonium SCOT-p) | Wayside near Glencoe (∼45 m a.s.l.), Lochaber, Scotland (56.684891°N −5.090494°E); isolated on 16 Aug 2014; algal filaments covered by thin water layer; air temperature: 16.9°C | Min. air temperature: −0.7°C to 9.7°C Max. air temperature: 5.5°C–17.4°C Monthly rainfall days: 14–20 Monthly precipitation: 77.0–248.3 mm Annual rainfall: 1809.4 mm |
Habitat characteristics and meteorological data (temperatures expressed as means, www.worldweatheronline.com, http://www.metoffice.gov.uk ) are given.
Iron and aluminium content in soil water
The iron and aluminium content of the soil water of Zygogonium AUT-p and Zygogonium SCOT-p were quantified colorimetrically by a Merck Spectroquant iron or aluminium test kit (1.14761.0001 (Fe), 1.14825.0001 (Al); concentration range 0.005–5 mg Fe L−1 or 0.02–1.2 mg Al L−1) according to the manufacturer's protocol (Merck, Darmstadt, Germany). For sample preservation prior analyses, 1 mL of nitric acid (65%; Sigma-Aldrich, Steinheim, Germany) per litre soil water was added to the soil water. A calibration curve was established by FeCl2·4 H2O or AlCl3 (both Sigma-Aldrich) and the metal content in the water expressed per mg L−1. The pH value of the soil water was measured in an untreated sample. Additionally, soil water of three other habitats in Obergurgl (Tyrol, Austria) located at least 500 m away from the habitat of Zygogonium AUT-p and predominated by Z. ericetorum (‘green morph’, field sample), Zygnema sp./ Z. ericetourm (‘green morph’, field sample) or Spirogyra sp. was analysed as described above for comparison. Iron and aluminium quantification were performed in four independent replicates (n = 4).
Metal quantification in Zygogonium AUT
Trace metal concentration (soluble Fe, Al and Zn) of Zygogonium AUT-p or Zygogonium AUT-g was determined by ICP OES (inductively coupled plasma/optical emission spectrometer; ULTIMA 2 Horiba Jobin Yvonne, Bensheim, Germany) equipped with an AS 80 auto sampler using the software ICP Analyst 5.2. The argon plasma torch (Air Liquide, >99 999%) was generated with a power of 1000 Watt and a flow rate of 12 L min−1. Dried algal samples were grinded with a mortar and extracted with methyl tert-butyl ether. Then, a phase separation against 20% ethanol was performed and both phases were evaporated and transferred to water and centrifuged at 13 000 g prior use. The clear supernatant was diluted ten times with nitric acid (67%–70%, c = 0.1 mmol L−1; Fisher Scientific, Vienna, Austria) in a 50 mL plastic screw tube. Samples were injected by a concentric Meinhard nebulizer in the concentric cyclone chamber. As calibration, the method of external standard was applied between 100 and 10.000 μg L−1. A multi standard was used (ICP-Standard-Solution 28 el. Roti Star, Carl Roth, Karlsruhe, Germany).
Haematoxylin staining and light microscopy
Haematoxylin was used frequently to stain iron and aluminium, which can be discriminated by colouration (Avwioro 2011; Fig. S2, Supporting Information). For haematoxylin staining, filaments of Zygogonium AUT-p and Zygogonium AUT-g were washed with A. dest. (3 × 10 min) followed by incubation in a 0.2% haematoxylin solution (Polle, Konzak and Kattrick 1978; Hollborn and Söhne, Leipzig, Germany) for 40 min at room temperature (RT) and washing (3 × 10 min in A. dest.). Stained and unstained (control) filaments were investigated with a Zeiss (Carl Zeiss AG, Jena, Germany) Axiovert 200M microscope (63 × 1.4 NA objective). Optical contrast was enhanced by using differential interference contrast (DIC). Images were captured with an Axiocam MRc5 camera using Zeiss Axiovision software. All images were further processed with the software Adobe Photoshop (CS5) version 12.1 (Adobe Systems, San José, CA, USA).
Morin Staining and confocal laser scanning microscopy
Morin (2',3,4',5,7-pentahydroxyflavone; VWR, Darmstadt, Germany) forms highly specific complexes with aluminium at pH = 5 (Browne, McColl and Driscoll 1990). Filaments of Zygogonium AUT–p and Zygogonium AUT-g were stained according to Navascués et al. (2012) with modifications: Filaments were rinsed in A. dest., washed in buffer (5 mM ammonium acetate (Sigma-Aldrich), pH = 5, 3 × 10 min), stained in morin (100 μM in buffer, 10 min) and washed in buffer (2 × 10 min). A Zeiss Pascal 5 CLSM system equipped with an argon laser (excitation 488 nm) on a Zeiss Axiovert 200 M was used to visualize morin (emission 505–550 nm, false colour green) and the chloroplast autofluorescence (emission 560 nm long pass filter, false colour red). A corresponding bright field image was merged with the autofluorescence image.
Transmission electron microscopy
Fixation for transmission electron microscopy (TEM) followed the protocol of Holzinger, Tschaikner and Remias (2010). Briefly, filaments of Zygogonium SCOT-p were fixed in sodium cacodylate puffer (10 mM, pH 6.8) containing 1.25% glutaraldehyde for 1.5 h. Filaments were transferred to 1% OsO4 and postfixed at 4°C for 16 h. After dehydration in increasing ethanol concentrations and propylene oxide, filaments were embedded in modified Spurr's resin, sectioned on a Leica ultracut (Leica Microsystems, Wetzlar, Germany), counterstained (uranyl acetate, Reynold's lead citrate) and examined with a Zeiss LIBRA 120 TEM at 80 kV connected to a Proscan 2 k SSCCD camera (Proscan Electronic Systems, Lagerlechfeld, Germany).
Photosynthetic oxygen measurements—light and temperature dependence
Photosynthesis irradiance (PI) curves were recorded according to Remias, Albert and Lütz (2010). Briefly, 3 mL of algal suspension (purple or green morph of Zygogonium AUT or Zygogonium SCOT) enriched with 2 mM NaHCO3 (final concentration) were filled in a thermostatic acrylic chamber on a magnetic stirrer and connected to a Presens Fibox 3 oxygen optode (Presens, Regensburg, Germany). Algae were exposed to eight light intensities (3–500 μmol photons m−2 s−1) at 20°C for always 6 min. In the dark period (6 min) directly before and after light measurements, respiratory oxygen consumption (R) was recorded and averaged to express R. After each measurement, chlorophyll (chl.) a was quantified according to Porra, Thompson and Kriedmann (1989) to estimate μmol O2 h−1 mg−1 chl. a in response to increasing light intensities as described by Herburger, Lewis and Holzinger (2015). PI curves were fitted by different models depending on whether photoinhibition occurred (Walsby 1997) or not (Webb, Newton and Starr 1974) to derive α, positive slope at limiting photon fluence rates (μmol O2 h−1 mg−1 chl. a (μmol photons−1 m−2 s−1)−1); Ic, light compensation point (μmol photons m−2 s−1); Ik, initial value of light-saturated photosynthesis (μmol photons m−2 s−1); and Pmax, maximum photosynthetic oxygen production under light saturation (μmol O2 h−1 mg−1 chl. a). To compare the PI curve kinetics of the purple or green morphs of Zygogonium AUT to Zygogonium SCOT and the purple to the green morph of Zygogonium AUT or Zygogonium SCOT to each other, linear regressions between the mean values of O2 production (μmol O2 h−1 mg−1 chl. a) at PAR 0, 3, 10, 14, 30, 70, 125, 290 and 500 μmol photons m−2 s−1 were calculated, and R2 was derived. Temperature dependence of photosynthetic oxygen production and respiratory consumption were examined according to Herburger, Lewis and Holzinger (2015). Three mL of algal suspension (purple or green morph of Zygogonium AUT or Zygogonium SCOT), enriched with 2 mM NaHCO3, were exposed to a temperature gradient (5°C–45°C in nine Δ5°C steps) at 100 μmol photons m−2 s−1. O2 production (gross photosynthesis) and consumption (respiration) were expressed in μmol O2 h−1 mg−1 chl. a, and net photosynthesis and gross photosynthesis:respiration (P/R) ratios were calculated for each temperature step.
Maximum quantum yield of PSII (Fv/Fm)—desiccation and rehydration response
Reduction and recovery of the maximum quantum yield of PSII (Fv/Fm) in response to desiccation followed by rehydration in Zygogonium SCOT-p and Zygogonium SCOT-g were measured according to Aigner et al. (2013). Algal filaments were transferred to Whatman (Whatman, Dassel, Germany) GF/F glass fibre filters (n = 6) in a desiccation chamber (Karsten, Herburger and Holzinger 2014) and dried at ambient air humidity and constant light (∼30 μmol photons m−2 s−1) and temperature (21 ± 0.5°C). A pulse-amplitude modulated fluorimeter (PAM 2500; Heinz Walz GmbH, Effeltrich, Germany) placed outside the chamber was used to measure the Fv/Fm value during desiccation and subsequent rehydration with filtered soil water (Zygogonium SCOT-p) or culture medium (Zygogonium SCOT-g).
Microscopic imaging PAM
The effective quantum yield of PSII (Y(II); false coloured image) and near-infrared remission (NIR, 780 nm) were visualized at the subcellular level with the microscopic version of an Imaging-PAM (M-series, Heinz Walz GmbH, Herburger and Holzinger 2015). Therefore, algal filaments of Zygogonium AUT-p and Zygogonium AUT-g exhibiting different cell morphologies (younger or older filaments, different stages of aplanospore formation and pigmentation) were collected by using a stereomicroscope (Lumar V12; Carl Zeiss AG) and transferred to welled slides. A modified Axio Scope A.1 epifluorescence microscope equipped with a Zeiss Fluar 40 × 1.3 NA objective and CCD Camera IMAG-K6 controlled with ImagingWinGigE (V2.45i) software was used for image generation. Measuring light for Y(II) determination was provided by a LED (620 nm). The signal/noise ratio was not modified by a SP-Routine.
Statistical evaluation of the data
Comparisons of soil water's iron and aluminium contents of five different habitats (n = 4), photosynthetic parameters derived from PI curves (α, Ic, Ik, Pmax, R; n = 3), effects of temperature (O2 production and consumption, P:R ratios; n = 3) or desiccation (Fv/Fm; n = 6) on photosynthesis were performed by one-way analysis of variance (ANOVA) followed by Tukey's post hoc test (P < 0.05) to find homogeneous subgroups of significantly different means. Analyses were carried out in Origin 8.5 software (OriginLab Corporation, Northampton, MA, USA).
RESULTS
Habitat characteristics
Zygogonium AUT-p filaments formed dense purple layers submersed into a shallow puddle (depth: ∼5–20 cm), which was drained by a streamlet (Fig. S3A, Supporting Information). Occasionally, aggregates of green filaments were found in deeper layers of the puddle and covered by the ‘purple morph’. Compared to the surrounding, vascular plants were less abundant inside the drained population of Zygogonium AUT-p (Fig. S3A, Supporting Information). On the sides with lower water availability (i.e. outside the pond and streamlet), Zygogonium AUT-p was scarce. The Scottish site was dryer compared to the habitat in Obergurgl (Fig. S3B, Supporting Information), since it was not drained by a streamlet and rainwater was the only water source. In this habitat, a thin layer of dark purple Zygogonium SCOT-p filaments coated by a thin water film covered the gravelly soil (Fig. S3C, Supporting Information), while green filaments (<5%) or co-occurring vascular plants were scarce (Fig. S3B, Supporting Information). Occasionally, filamentous cyanobacteria were observed in both habitats. Detailed habitat characteristic including metrological data are listed in Table 1.
Iron and aluminium content in soil water
The iron and aluminium content between the soil water of habitats of Zygogonium AUT-p, Zygogonium SCOT-p and the soil water of three other habitats predominated by zygnematalean green algae in Obergurgl (Austria) differed significantly (Fig. 1). The highest iron contents were measured in the habitats of Zygogonium AUT-p (1.09 mg Fe L−1) and Zygogonium SCOT-p (1.14 mg Fe L−1; Fig. 1). Habitats predominated by a ‘green morph’ of Z. ericetorum (0.18 mg Fe L−1), a ‘green morph’ of Z. ericetorum and Zygnema sp. (0.09 mg Fe L−1) or Spirogyra sp. (0.08 mg Fe L−1) contained significantly less iron (Fig. 1). The highest aluminium content occurred in the habitat of Zygogonium AUT-p (0.29 mg Al L−1; Fig. 1). Significantly, lower values occurred in the site of Zygogonium SCOT-p (0.17 mg Al L−1; Fig. 1). The aluminium content was very low in habitats predominated by Zygnema sp. and a ‘green morph’ of Z. ericetorum and not measurable in the habitat of Spirogyra sp. due to too low concentrations (Fig. 1). The pH values in the soil water samples of Zygogonium AUT-p and Zygogonium SCOT-p (∼4.6) were lower when compared with the three other sites (∼6.2–8.6; Fig. 1).
Figure 1.

Comparison of the photometrically detected iron and aluminium content (n = 4) and pH value of soil water from Zygogonium AUT-p, Zygogonium SCOT-p and three different Austrian habitats containing a ‘green morph’ of Zygogonium, a mixture of Zygnema sp. and a ‘green morph’ of Zygogonium or Spirogyra sp. (n = 4 ± SD). Significant differences between the groups are indicated by lower case (iron content) or capital (aluminium content) letters. Data were analysed by one-way ANOVA followed by Tukey's post hoc test (P < 0.05).
Metal quantification in Zygogonium AUT
Filaments of Zygogonium AUT-p contained much more Al, Zn and Fe when compared to Zygogonium AUT-g, where aluminium showed higher values compared to zinc and iron (Table 2).
Table 2.
Mean (n = 2) and range (parentheses) of metal concentrations (μg mg dry mass−1) in Zygogonium AUT-p and Zygogonium AUT-g measured by ICP OES.
| Al | Zn | Fe | |
|---|---|---|---|
| Zygogonium AUT-p | 0.938 | 0.844 | 0.459 |
| (0.683–1.193) | (0.567–1.121) | (0.187–0.730) | |
| Zygogonium AUT-g | 0.248 | 0.003 | 0.054 |
| (0.194–0.303) | (0.002–0.003) | (0.042–0.066) |
Light microscopy
Cell morphology and ultrastructure of vegetative and conjugating filaments of Zygogonium AUT-p have been described in detail previously (Holzinger, Tschaikner and Remias 2010; Aigner et al.2013; Stancheva et al.2014, 2016). Thus, light microscopic examination in this study focused on Zygogonium SCOT-p and Zygogonium SCOT-g, which also exhibited vegetative cells with a high morphological variability (Fig. 2). Around >95% of the filaments obtained from the natural population were purple with one centrally located nucleus, two plate-like shaped chloroplasts surrounded by transparent particles, which were scarce in the cell periphery (Fig. 2A). Rarely (∼2%), filaments exhibited spherical dark purple inclusions near one cell pole and dentate outer cell walls (Fig. 2C). After two weeks of cultivation in BBM, the purple pigmentation was lost (Fig. 2B), while ∼25% of the filaments stored in soil water from the habitat (pH = ∼4.6) remained purple for several weeks. H-shaped cell wall structures were observed frequently in both the purple and green morph (Fig. 2A and B). Aplanospores occurred frequently and appeared similar as described in detail by Stancheva et al. (2014), including the spores being attached to the cross cell walls of the vegetative cell and dark cytoplasmic residue outside the spores (Fig. 2D and E). Occasionally (∼1% of filaments), a type of cell encystment was observed, where encysted cells occupied the centre of vegetative cells (Fig. 2F and G). These encysted cells contained ‘highly ordered’ transparent particles in the periphery of their lumen, and they were embedded in dark cytoplasmic residue (Fig. 2F and G). Encysted cells lacked highly condensed cytoplasm and cell organelles common in aplanospores. Germination of encysted cells was not observed. In contrast, aplanospores germinated into new vegetative cells, pushing the cytoplasmic residue outside the cells to H-shaped cell wall structures (Fig. 2H), where fragmentation took place frequently (not shown). This process occurred in natural field samples (Zygogonium AUT-p) and in subsamples after transferring them to BBM. In contrast, aplanospore formation and germination was not observed in Austrian habitats predominated by the ‘green morphs’ of Z. ericetorum.
Figure 2.

Light micrographs of vegetative cells (A–C, F–H) and aplanospores (D, E) of Zygogonium SCOT (A, C–H: Zygogonium SCOT-p, B: Zygogonium SCOT-g). (A), (B) H-shaped cell wall structures (arrowheads). (C) One spherical dark purple inclusion per cell (arrow), dentate longitudinal cell walls. (D) Small aplanospores; brownish or purple granular cytoplasmic residue outside the spores. (E) Aplanospores embedded in brownish cytoplasmic residue and; numerous transparent particles in the periphery of the spore (arrows). (F) Encysted cells with two prominent chloroplasts. (G) Encysted cells surrounded by ‘highly ordered’ spherical bodies (arrows). (H) Purple filament formed by germination of aplanospores as indicated by the brownish remnant of cytoplasmic residue; H-shaped cell wall structures (arrowheads). Bars = 20 μm.
Haematoxylin staining
Incubating Zygogonium AUT-p in haematoxylin stained the cell walls of ∼10% of the filaments entirely dark violet to blackish, while particularly strongly stained sheaths covered H-shaped cell wall structures between individual cells (Fig. 3A). Frequently, dark violet to blackish staining restricted to H-shaped cell wall structures occurred and lilac stained areas of the cell walls were found between these structures (Fig. 3B). In contrast, filaments lacking strong staining in or on the outer cell walls, showed dark violet to blackish staining of spherical bodies in the protoplast with a maximum in the cell periphery and cross cell walls (Fig. 3C). In filaments depositing cell wall material close to the cross cell walls (i.e. beginning aplanospore formation), blackish staining was restricted to chloroplast-free areas (Fig. 3D and E). Aplanospores lacked staining, while it was particularly strong in the cytoplasmic residue outside the spores (Fig. 3F). Germinated aplanospores, which were either purple (Fig. 3G) or green (Fig. 3H), lacked staining, which was restricted to delimited areas adjacent to the spores. Occasionally, green filaments showed lilac haematoxylin staining in specific areas of the outer and cross cell walls (Fig. 3I), while cross-wall protuberances (Fig. 3J) and H-shaped cell wall structures were stained frequently (Fig. 3H).
Figure 3.

Haematoxylin staining of Zygogonium AUT-p (A–G) and Zygogonium AUT-g (H–J) filaments. (A) Strong violet staining in the cell and in H-shaped cell wall structure (arrows). (B) Lilac (arrowheads) and dark violet to blackish staining in the cell wall. (C) Dark violet to blackish staining inside the protoplast (arrowheads) and cross cell walls (arrows). (D, E) Strong and sharply delineated staining in the protoplast near cell poles (arrow); deposition of cell wall material (arrowheads). (F) Strong blackish staining in the extracellular space outside unpigmented aplanospores. (G) Germinated aplasnpores with lilac staining in the cell wall (arrowheads). (H) Lilac staining in H-shaped cell wall structure, (I) longitudinal and cross cell walls and (J) cross cell wall protuberances. Bars = 20 μm.
Morin staining
Aluminium was detected by morin staining in both Zygogonium AUT-p and Zygogonium AUT-g, with a maximum in H-shaped cell wall structures (Fig. 4). Zygogonium AUT-p showed stronger labelling in the cell walls and inside one third of the cells in the pyrenoid regions (Fig. 4B). In contrast, Zygogonium AUT-g lacked staining inside cells (Fig. 4D). Visualization of the chloroplast autofluorescence by confocal laser scanning microscopy (CLSM) revealed variably shaped chloroplasts in vegetative cells, ranging from a rounded to irregularly rounded appearance with several protrusions in one plane (Fig. 4A and C). Controls, where morin was omitted, lacked fluorescence in the green channel (not shown).
Figure 4.
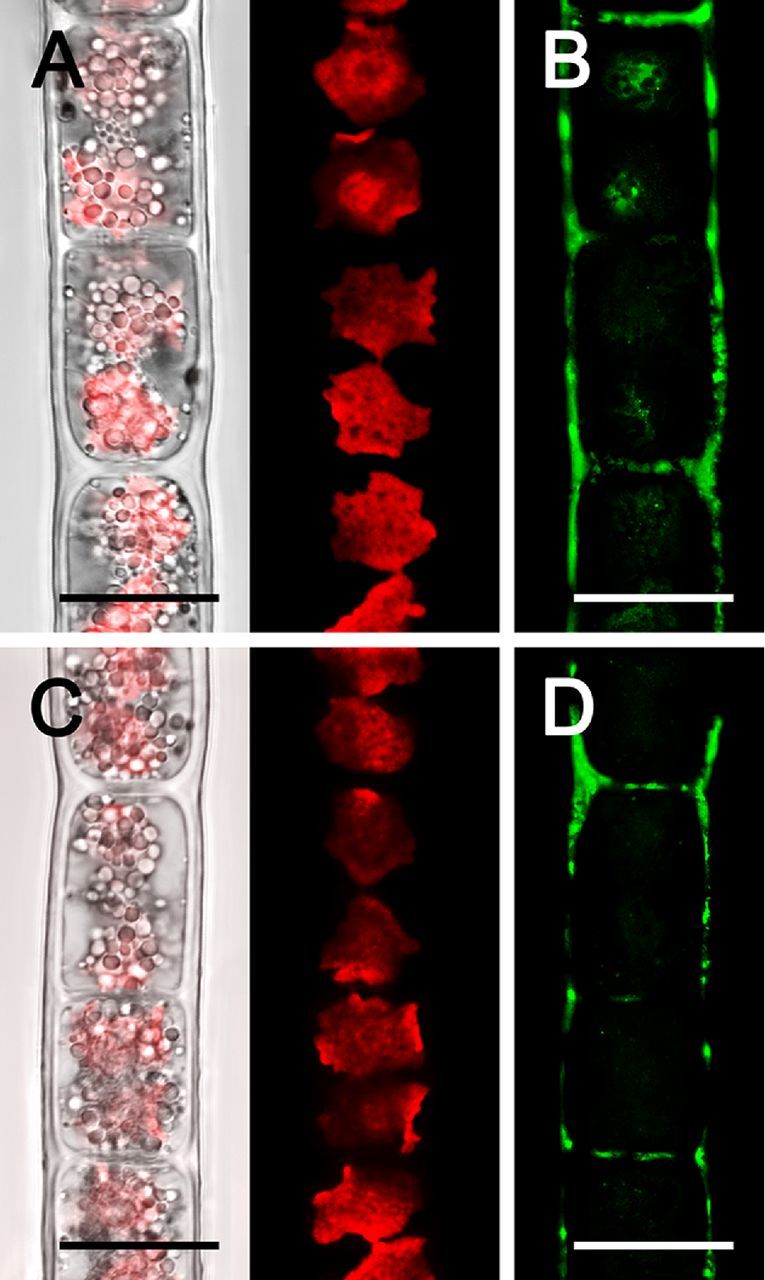
Confocal laser scanning micrographs (A, C) and morin (green) staining (B, D) of Zygogonium AUT-p (A, B) and Zygogonium AUT-g (C, D). (A, C) Bright field image merged with corresponding chloroplast autofluorescence (red) image. Bars = 20 μm.
Transmission electron microscopy
Each cell of Zygogonium SCOT-p contained two plate-like chloroplasts with an enlarged central part, holding one pyrenoid, which was invaginated by thylakoid membranes and surrounded by one layer of large starch grains (Fig. 5A–C). Occasionally, additional starch grains were deposited throughout the chloroplast (Fig. 5A). Plastoglobules were scarce (Fig. 5B). Sometimes, thylakoid membranes containing wings protruded from the central part of the chloroplast (Fig. 5C). Cells were filled with numerous vacuoles with different sizes (diameter 0.1–9 μm) and appearances (electron—opaque or granulated contents; Fig. 5A and B). Several Golgi bodies were located closely to the chloroplasts (Fig. 5B). Longitudinal and cross cell walls were layered and had a diameter of 0.6–3.5 μm (Fig. 5B and D). Prominent layered H-shaped cell wall structures between individual cells were observed frequently, which ruptured towards their edges (Fig. 5D). Occasionally, mitochondria were distinctly enlarged and cristae bloated (Fig. 5E).
Figure 5.
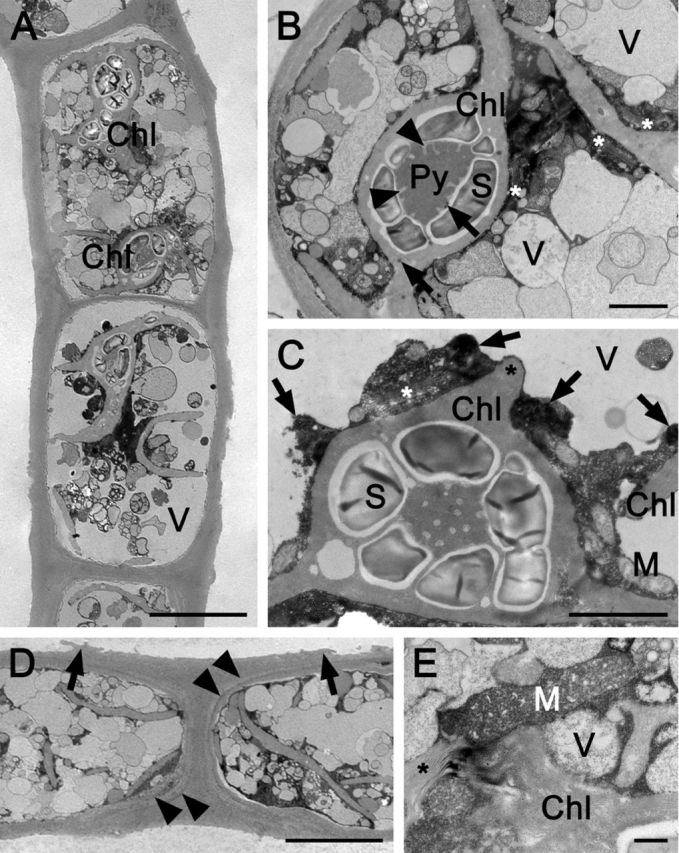
Transmission electron micrographs of longitudinal sections through Zygogonium SCOT-p. (A) Overview of two cells. (B) Detail of the protoplast: one pyrenoid with tubular invaginations, starch grains; plastoglobules (arrows); numerous vacuoles; Golgi bodies (asterisks). (C) Detail of a chloroplast with protruding wing (asterisk), pyrenoid, starch grains, mitochondria, Golgi bodies (asterisk), electron-dense structures (arrows) and vacuoles. (D) H-shaped cell wall structure; cell wall layers outside the innermost layer surrounding the cells are marked with arrowheads; H-shaped structures detach toward their edges (arrows). (E) Enlarged mitochondrion with distinctly bloated cristae; Golgi bodies (asterisk). Chl chloroplast, Py pyrenoid, M mitochondrion, S starch grain and V vacuole. Bars = 10 μm (a, d), 2 μm (b, c) and 0.5 μm (e).
Light dependence of photosynthesis
Photosynthetic oxygen production and respiratory consumption in response to light gradients depended on the habitats the algae were obtained from and whether pigmentation was present or not (Fig. 6A). Zygogonium AUT-p showed the highest maximum O2 production (Pmax) and respiratory consumption (R), followed by Zygogonium SCOT-p (Fig. 6A; Table 3). In general, both ‘green morphs’ exhibited lower Pmax and R values when compared to the ‘purple morphs’, while the lowest values were measured in Zygogonium SCOT-g (Fig. 6A; Table 3). In contrast to Zygogonium AUT, R did not differ significantly between the green and purple morph of Zygogonium SCOT (Table 3). Zygogonium AUT exhibited significantly (P < 0.05) higher α and Ic values compared to Zygogonium SCOT, while the two morphs of the same isolate did not differ significantly (P < 0.05) from each other (Table 3). In contrast, the Ik values of both ‘purple morphs’ were similar but significantly (P < 0.05) lower in the ‘green morphs’, while Zygogonium SCOT-g exhibited the lowest value (Table 3). Linear regression between the photosynthetic oxygen production and respiratory consumption from PAR 0 to 500 μmol photons m−2 s−1 in the purple and green morph of Zygogonium AUT and Zygogonium SCOT revealed a strong linear correlation (Fig. 6B). Similarly, the light curve data of the two different ‘purple morphs’ or ‘green morphs’, respectively, were strongly linearly correlated to each other (Fig. 6B).
Figure 6.
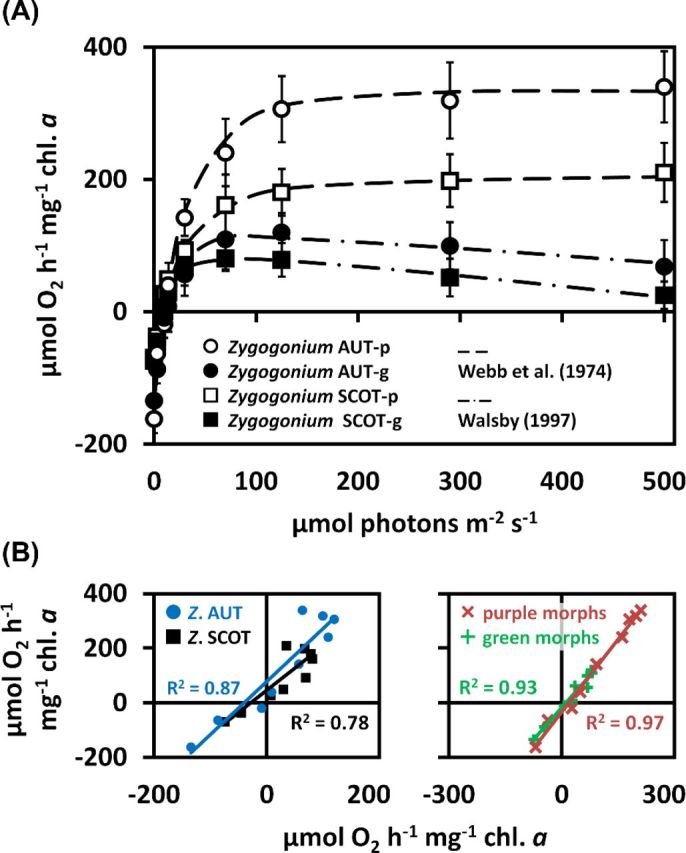
(A) Photosynthetic O2 production and respiratory consumption as a function of increasing PAR up to 500 μmol photons m−2 s−1 measured in the purple and green morphs of Zygogonium AUT and Zygogonium SCOT (n = 3 ± SD). Data points of the ‘purple morphs’ and ‘green morphs’ were fitted according to Webb, Newton and Starr (1974) and Walsby (1997), respectively. (B) Linearly correlated O2 production/consumption at increasing PAR. Right: correlations of the ‘purple morph’ (x-axis) with the ‘green morph’ (y-axis) of Zygogonium AUT or Zygogonium SCOT. Left: correlations of the two ‘purple morphs’ or ‘green morphs’ of Zygogonium AUT (y-axis) and Zygogonium SCOT (x-axis) to each other. Coefficients of determination (R2) are shown.
Table 3.
Photosynthetic parameters of PI curves (O2) of the purple and green morph of Zygogonium AUT and Zygogonium SCOT, calculated by using the fitting models of Walsby 1997 (‘green morphs’) or Webb, Newton and Starr 1974 (‘purple morphs’).
| α | Ic | Ik | Pmax | R | |
|---|---|---|---|---|---|
| Zygogonium AUT-p | 11 ± 2.64a | 12.99 ± 2.02a | 28.42 ± 5.23a | 315.5 ± 28.38a | −161.93 ± 15.3a |
| Zygogonium AUT-g | 13.12 ± 2.17a | 11.77 ± 0.95a | 16.76 ± 2.99b | 113.86 ± 14.79b | −134.57 ± 12.72b |
| Zygogonium SCOT-p | 7.89 ± 1.27b | 7.5 ± 1.27b | 29.79 ± 8.01a | 148.95 ± 18.08c | −69.48 ± 6.64c |
| Zygogonium SCOT-g | 9.23 ± 1.83b | 8.08 ± 0.95b | 11.17 ± 1.12b | 103.81 ± 12.89b | −74.36 ± 11.64c |
Different lower case letters indicate significant differences between the values (α, Ic, Ik, Pmax or R) of the four different groups. They were determined by one-way ANOVA followed by Tukey's post hoc test (P < 0.05). α initial slope in the light-limiting range (μmol O2 h−1 mg−1 chl. a (μmol photons m−2 s−1)-1), Ic light compensation point (μmol photons m−2 s−1), Ik initial value of light-saturated photosynthesis (μmol photons m−2 s−1), Pmax maximum photosynthetic rate in the light-saturated range (μmol O2 h−1 mg−1 chl. a), R respiration rate in the dark (μmol O2 h−1 mg−1 chl. a).
Temperature dependence of photosynthesis
In all four samples investigated (Zygogonium AUT-p/g and SCOT-p/g), photosynthetic oxygen production and respiratory consumption were strongly temperature-dependent (Fig. 7A). Both Zygogonium AUT-p and Zygogonium SCOT-p exhibited a linearly increasing O2 production from 5°C to 20°C (AUT) or to 25°C (SCOT), followed by a decrease to negative values at 45°C. At 40°C, positive oxygen production was still measurable, while Zygogonium SCOT-p showed a higher O2 production at this temperature step when compared with Zygogonium AUT-p (Fig. 7A). Respiratory oxygen consumption increased linearly from 5°C and reached a broad temperature maximum between 25°C and 45°C (AUT) or 40°C and 45°C (SCOT; Fig. 7A). In general, both Zygogonium AUT-g and Zygogonium SCOT-g exhibited a lower photosynthetic oxygen production in response to increasing temperatures from 5°C to 45°C (Fig. 7A). Gross photosynthetic O2 production increased linearly from 5°C to 20°C (AUT) or 5°C to 20°C (SCOT), followed by a linearly decrease and reaching negative values at 45°C in the ‘green morphs’ (Fig. 7A).The lower temperature depended photosynthetic oxygen production in Zygogonium AUT-g and Zygogonium SCOT-g is also reflected by the gross photosynthesis:respiration (P:R) ratios, which were lower compared to the respective ‘purple morphs’ at temperature steps with positive net photosynthesis (AUT: 5°C–30°C; SCOT: 5°C–35°C; Fig. 7B). Both Austrian morphs showed the highest P:R ratios at 5°C, while in both Scottish morphs the highest P:R ratios values occurred at 10°C (Fig. 7B).
Figure 7.
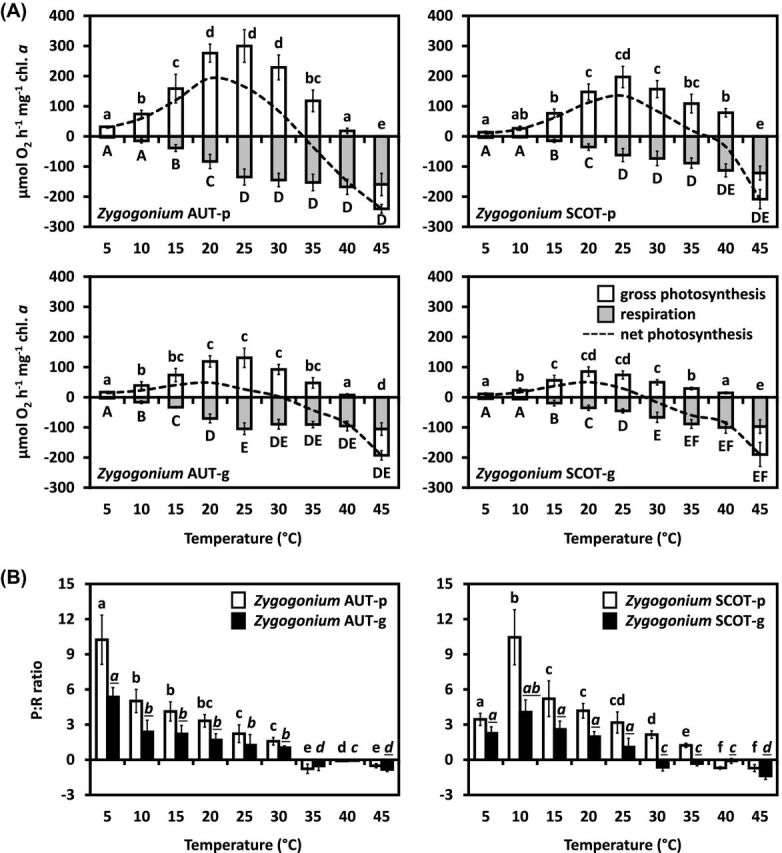
Temperature dependence of photosynthesis in the ‘purple morph’ and ‘green morph’ of Zygogonium AUT and SCOT. (A) Gross and net photosynthetic O2 production and respiratory consumption in response to increasing temperatures (0°C –45°C; n = 3 ± SD; SD values for net photosynthesis are excluded to increase legibility). (B) Gross photosynthesis: respiration (P:R) ratios. Significant temperature depended differences (lower case letters: gross photosynthesis, capital letters: respiration, lower case and cursive letters: P:R ratios) were determined by one-way ANOVA, followed by Tukey's post hoc test (P < 0.05). Significant different P:R ratios of the ‘purple morph’ and the ‘green morph’ of the same strain at the same temperature steps (underlined letters) were determined by standard two-sample t-tests (P < 0.05).
Optimum quantum yield of PSII (Fv/Fm)—desiccation response
In Zygogonium SCOT-p, desiccation at ambient air reduced the initial Fv/Fm value (0.57 ± 0.02) linearly after 90 min (P < 0.05), reaching 0 after 270 min (Fig. 8A). Subsequent rehydration with culture medium started recovery of Fv/Fm after 5 min., while after 300 min ∼40% of the initial value was restored (Fig. 8A). After 1 day of recovery, Fv/Fm increased to ∼60% of the initial value and remained unchanged (P < 0.05) for 4 days (Fig. 8A). In Zygogonium SCOT-g, the initial Fv/Fm value (0.55 ± 0.05) started decreasing significantly (P < 0.05) after 60 min of desiccation and reached ∼45% of the initial value after 240 min (Fig. 8B). After 270 min, Fv/Fm reached 0 (Fig. 8B). Rehydration started recovery of Fv/Fm immediately and after 300 min, ∼30% were restored (Fig. 8B). Fv/Fm increased to ∼60% after 1 day, followed by another significant increase (P < 0.05) to ∼90% of the initial Fv/Fm value after 4 days (Fig. 8B).
Figure 8.

Effect of desiccation followed by subsequent rehydration on the maximum quantum yield of PSII (Fv/Fm; n = 6 ± SD) of Zygogonium SCOT-p (A) and Zygogonium SCOT-g (B). The control Fv/Fm value is indicated as a dashed grey line on the rehydration charts (right). Significant different effects of desiccation and rehydration on the Fv/Fm values over time (small letters) were determined by one-way ANOVA followed by Tukey's post hoc test (P < 0.05).
Microscopic imaging PAM
Using the microscopic version of an Imaging, PAM allowed to visualize the effective quantum yield of PSII [Y(II)] on a cellular level. Most cells of Zygogonium AUT-p (Fig. 9A) and Zygogonium AUT-g (Fig. 9B) exhibited a similar Y(II) (∼0.61), while only in some chloroplasts the value was lower (Fig. 9A, B). Older filaments (indicated by cells with thick cell walls and filled with storage compounds) showed a lower Y(II) (Fig. 9C). Similarly, cells depositing cell wall material close to the cross cell walls (Fig. 9D), aplanospores (Fig. 9E), or encysted cells (Fig. 9F) showed a Y(II) which amounted to ∼50% of the value measured in usual vegetative cells. Germinated aplanospores and adjacent cells (Fig. 9G) as well as cells containing one conspicuous spherical inclusion (Fig. 9H) showed a similar Y(II) like usual green or purple cells.
Figure 9.

NIR and corresponding Y(II) images (false colour) of Zygogonium AUT filaments with different cell morphologies. The relative Y(II) as a percentages is indicated by the colour bar at the top. (A) Zygogonium AUT-p. (B) Zygogonium AUT-g. (C) Purple filament with older cells. (D) Cells depositing cell wall material close to cross cell walls (arrowheads). (E) Filament with numerous aplanospores close to cross cell walls of vegetative cells. (F) Encysted cells. (G) Germinated aplanospores as indicated by dark remnant of cytoplasmic residue. (H) Purple filament with one spherical inclusion per cell. Bars = 20 μm.
DISCUSSION
The colonization of land by zygnematalean green algae was one of the most pivotal events in the history of life on earth, as it founded the rise of Embryophytes, which shape the extant ecosystems on earth's surface (Waters 2003; Harholt, Moestrup and Ulvskov 2015). These multicellular land plants are characterized by a high level of functional specialization allowing them to partition complementary tasks among different cells, tissues or organs simultaneously (e.g. the mesophyll photosynthesizes, while cutinized epidermal tissues decrease water loss and roots accumulate absorbed heavy metal ions to prevent their harmful spread throughout the plant). In contrast, functional specialization in most algae is restricted to individual cells and the formation of different phenotypes of the same cell lineage (Grosberg and Strathmann 2007). In the vegetative stage, the zygnematalean alga Z. ericetorum forms different cell morphologies in response to the environment (Transeau 1951; Stancheva et al.2014, 2016). Therefore, this filamentous alga serves as an ideal model organism to investigate cellular reorganization processes, involving morphologically different cells in the ancestors of Embryophytes.
Photophysiology of Zygogonium
The most striking cell differentiation of Z. ericetorum is the formation of purple pigmentation (Lagerheim 1895; Alston 1958; Holzinger, Tschaikner and Remias 2010; Stancheva et al.2014), which strongly influences the eco-physiological performance. In contrast to the Austrian and Scottish ‘green morphs’ established in the lab (i.e. Zygogonium AUT-g and Zygogonium SCOT-g), the field collected ‘purple morphs’ (Zygogonium AUT-p, Zygogonium SCOT-p) lacked photoinhibition at 500 μmol photons m−2 s−1. This corresponds to previous rETR measurements in a purple and green morph of Z. ericetorum isolated from Obergurgl (Austria; Aigner et al.2013). Interestingly, the light response kinetics of the green and purple morphs investigated in the present study were not strongly correlated throughout the applied light range (0–500 μmol photons m−2 s−1): while both the green and purple morph of Zygogonium AUT or Zygogonium SCOT, showed similar low light key figures (α and Ic value), the light saturation points and Pmax values were always significantly higher in the ‘purple morphs’. This indicates that the pigment acts as a sunscreen and protector against harmful irradiation. It absorbs highly in the UVR and PAR range (Newsome, Murphy and van Breemen 2013) and gets lost in the absence of UVR (Aigner et al.2013) or in standard growth medium exhibiting low concentration of iron, which is part of the purple pigment. Aigner et al. (2013) stated that Z. ericetorum lacks UV-absorbing mycosporine-like amino acids or secondary carotenoids, making phenolic compounds particularity important for photoprotection. Advantageously, phenolics do not contain nitrogen, which allows Z. ericetorum to occur in oligotrophic environments such as high alpine streamlets (Obergurgl, Austria) or gravely soils (Glen Coe, Scotland) or even xeric sandy soilcrusts (Hoppert et al.2004). Only upper layers of Z. ericetorum populations synthesize high amounts of the purple ferric (gallate)2 complex, which is part of a polysaccharide (MW 30 kDa) of variably linked and branched glucose monomers (Newsome, Murphy and van Breemen 2013). In these sun-exposed filaments, photosynthesis is rarely light limited and allows cells to produce and maintain high concentrations of this carbon-rich pigment by increasing the photosynthetic performance. Interestingly, both the purple and green morph of Zygogonium SCOT exhibited a significantly lower photosynthetic oxygen production and consumption compared to Zygogonium AUT, while the PI curve kinetics were very similar, pointing to a high phenotypic plasticity (Lürling 2003; Herburger, Karsten and Holzinger 2016). On the other hand, photosynthesis (Fv/Fm) in Zygogonium SCOT showed a higher resistance against desiccation stress and recovery upon rehydration when compared with Z. ericetorum isolated from Obergurgl (Austria; Aigner et al.2013). Since Zygogonium SCOT-p occurred in a gravely soil habitat, where rainwater was the only source of available water, a stimulation of defence mechanisms against cellular water loss via hardening can be assumed (Pichrtová, Hajek and Elster 2014a). Furthermore, measuring temperature dependence of photosynthesis revealed the highest P:R ratio to occur at low temperatures and in the ‘purple morphs’ (Zygogonium AUT: 5°C; Zygogonium SCOT: 10°C), which reflects the temperature regime in the respective natural habitats (Table S1, Supporting Information). Thus, the upper layers of purple algal filaments, which are exposed to the atmosphere, can gain sufficient amounts of fixed carbon, which is not used for respiration and available for glycosylated pigment and/or biomass synthesis (e.g. formation of aplanospores). This is also supported by the observation that Zygogonium SCOT-p exhibited several Golgi bodies close to the chloroplast, the primary location for carbohydrate synthesis in plants.
Formation of aplanospores removes iron from the protoplast
Using iron in acidic environments as a component of vacuolated sunscreen pigments is not only an effective strategy to cope with harmful solar and UV radiation. It also shifts excessive iron from the cytoplasm and chloroplasts into the vacuoles and reduces the risk of forming reactive oxygen species due to the Fenton reaction. The acidic habitats of Zygogonium AUT-p and Zygogonium SCOT-p (pH = ∼4.6) exhibited a much higher iron content (∼1.1 mg Fe L−1) when compared to more alkaline soil solutions in the region of Obergurgl (pH = ∼6.2–8.6) predominated by a naturally occurring ‘green morph’ of Z. ericetorum and other zygnematalean green algae (0.08–0.18 mg Fe L−1), Southwest Scotland (0.22 mg Fe L−1; Grieve 1990) or usual drinking water (0.30 Fe mg L−1; World Health Organization 1996). Excessive concentrations of heavy metals such as iron cause acute and chronic toxic effects on the metabolism of even metal tolerant plants (Nagajyoti, Lee and Sreekanth 2010). In algae, three principal mechanisms allow to maintain non-toxic concentrations of metal ions inside cells (Gaur and Rai 2001): (i) exclusion (e.g. binding to extracellular polysaccharides), (ii) prevention of bioavailability by complexing them inside the cells (e.g. binding to class III metallothioneins; Perales-Vela, Peña-Castro and Canizares-Villanueva 2006) and (iii) excretion from the protoplasts. Z. ericetorum cells use all three strategies to cope with iron-rich habitats by (i) binding iron to the extracellular matrix with a maximum in H-shaped cell wall structures between individual cells and by (ii) incorporating iron into a glucose-rich phenolic vacuolar compound. Finally, (iii) aplanospore formation shifts iron-rich compounds to cell poles of vegetative cells; as a consequence, iron is excluded from emerging aplanospores and rendered inert in the extracellular space surrounded by the parental cell wall. These are the areas, where cell detachment occurs frequently, which releases iron-rich residues from the filament into the environment and explains the lower iron content in the ‘green morph’. Microscopic imaging PAM revealed that the effective quantum yield of PSII [Y(II)] of Zygogonium AUT-g (low iron content) and Zygogonium AUT-p (high iron content) as well as germinated aplanospores was similar (∼0.61). This indicates a fine regulation of iron homeostasis, allowing Z. ericetorum to occur in acidic (pH = ∼4.6) iron rich as well as more alkaline habitats (pH = ∼6.2–6.4) with lower iron concentrations, where the formation of aplanospores as a mechanism of active iron exclusion was not observed.
Aluminium is abundant in the cell wall of the green and purple morph
Zygogonium AUT-p contained ∼3.5-fold more aluminium when compared with Zygogonium AUT-g as shown by ICP OES. Haematoxylin and morin staining revealed that both morphs enriched this metal in their cell walls, while only the ‘purple morph’ showed morin staining in some chloroplasts with a maximum in the pyrenoid regions. The accumulation of aluminium inside cells of the ‘purple morph’ might be explained by a higher aluminium concentration and a lower pH value (∼4.6) in the soil water when compared with the culture medium (pH = 6.8), where the ‘green morph’ was generated. Aluminium solubility and phytotoxicity are influenced by the pH value of the solution the organisms live in (Godbold, Jentschke and Marschner 1995; Crémazy, Campbell and Fortin 2013). In the streptophyte green algae Chara corallina, aluminium transport across the plasma membrane showed a maximum at pH 4.3 and dropped strongly at pH 5.2 (Taylor et al.2000). The ‘purple morphs’ of Z. ericetorum investigated in the present study occurred in habitats with a pH of ∼4.6, where transport of Al3+ across membranes is still possible (Taylor et al.2000), explaining the accumulation in chloroplasts, which was also found in several land plants (Cuenca, Herrera and Merida 1991; De Andrade et al.2011) but not in the ‘green morphs’ established in the lab. In contrast, aluminium accumulation in the cell wall is less pH depended (Taylor et al.2000). Like land plants, the cell walls of Zygnematophyceae contain homogalacturonan (Popper et al.2011), which consists of an α-D-1,4-galacturonan core (Willats et al.2001). In the cell wall, homogaladcturonan is de-methylesterified by pectin methylesterase enzymes (Domozych et al.2014), resulting into free carboxylic groups, which are the major binding sites of aluminium ions (Schmohl et al.2000). Under culture conditions, aluminium accumulation in the cell wall depends on the presence of lipid peroxidizing Fe2+, which mediates the production or leakage of new pectins as a primary ligand for aluminium binding (Chang, Yamamoto and Matsumoto 1999). Furthermore, the modulation of the pectin content modifies aluminium sensitivity in plant roots (Schmohl and Horst 2000; Eticha, Stass and Horst 2005), while the amount of de-methylesterified pectins correlates positively with aluminium accumulation and sensitivity due to increased binding of this metal to the more negatively charged cell wall (Horst et al.1999). Both the ‘purple morph’ of natural field populations of Z. ericetorum from Obergurgl (Austria; Holzinger, Tschaikner and Remias 2010) and Zygogonium SCOT-p can produce massive layered cell walls with a diameter up to 4 μm. Thus, the drastically higher aluminium content of Zygogonium AUT-p might be explained by different cell wall chemistries, namely a higher content of pectins and/or degree of de-methylation (Eticha, Stass and Horst 2005). In general, streptophyte green algae can modify their cell wall composition in response to environmental conditions (Herburger and Holzinger 2015) and increase the pectin content strongly under e.g. desiccation stress (Pichrtová, Kulichová and Holzinger 2014b, and references therein). Binding of Al3+ to the pectic matrix decreases the mechanical extensibility of the cell wall and reduces cell growth and development, likely by limiting the movement of wall-loosening enzymes (Eticha, Stass and Horst 2005). Interestingly, aplanospores of Z. ericetorum form their own cellulosic wall embedded in the pectin-rich parental cell wall (Stancheva et al.2014), while aluminium has a very low-binding affinity to cellulose (Chang, Yamamoto and Matsumoto 1999). This de novo formation of a cellulosic wall around the spores, followed by their germination inside the filament, allows cell division and filament growth independently of the stiff aluminium cross-linked pectic matrix of the parental wall. Haematoxylin staining in germinated aplanospores of green Zygogonium filaments showed that pectic cross-wall protuberances (as shown for Desmidium swartzii (Zygnematophyceae); Andosch et al.2015) and H-shaped cell wall structures between individual cells were particularly rich in aluminium. The latter structures, which are also stained by morin, are remnant of parental cell walls originated from several cell division and wall formation cycles which rupture and detach over time. After transferring the purple field samples of Zygogonium AUT-p to aluminium-free culture medium, green cells (Zygogonium AUT-g) still contained considerable amounts of aluminium, which was restricted to the cell wall. This might reflect the non-exchangeable fraction of aluminium (Tice, Parker and DeMason 1992), which is tightly bound to the pectic matrix (yet stainable by haematoxylin and morin) with a maximum in distinctly layered H-shaped cell wall pieces still attached to cells and germinated aplanospores. Over time, these pieces detach from the cells due to rupture or mechanical strain. They might be also related to cell detachment (Stancheva et al.2014; Herburger, Karsten and Holzinger 2016).
High zinc content does not affect photosynthetic performance
In contrast to aluminium, Zygogonium AUT-g contained very low amounts of zinc, while Zygogonium AUT-p showed similar values like aluminium. Zinc can decrease growth, cell division rate and the photosynthetic activity of Zygnematophyceae (Volland et al.2011). Furthermore, uptake of zinc results into strongly enlarged mitochondria with bloated cristae (Andosch et al.2015), which was also observed in the ‘purple morph’ of Z. ericetorum isolated from Obergurgl (Austria; Holzinger, Tschaikner and Remias 2010) and Zygogonium SCOT-p. However, negative effects on photosynthesis were not observed, since Zygogonium AUT-p (high zinc content) showed similar Y(II) values when compared with Zygogonium AUT-g (low zinc content) and even a higher photosynthetic O2 production and respiratory consumption. Therefore, similar to Micrasterias denticulata (Zygnematophyceae) and several land plants (Volland et al.2011 and references therein), Zn might be detoxified by compartmentalization in the cell wall, vacuoles and mucilage vesicles, while the latter might be involved in removing excessive Zn from the protoplast.
CONCLUSION
Based on the observations in the present study, we consider the formation of aplanospores as a rejuvenation process, allowing Z. ericetorum to survive in acidic metal-rich habitats, where other zygnematalean algae (Zygnema sp., Spirogyra sp.) cannot occur: (i) Spore formation shifts excessive iron-rich compounds in the cell periphery and excludes them from emerging unpigmented aplanospores; (ii) spore germination allows filament growth despite high amounts of aluminium, iron and likely zinc in the pectin-rich parental cell wall matrix. Both the purple and green morph of Z. ericetorum, maintain a high photosynthetic performance in acidic (pH = ∼4.6) and neutral (pH = 6.8) environments, while the presence of the purple iron-rich pigment coincides with an considerable higher photosynthetic oxygen production in response to higher light intensities up to 500 μmol photons m−2 s−1 and temperature gradients (5°C–45°C). Different responses to light, temperature and desiccation gradients followed by rehydration in Zygogonium AUT and Zygogonum SCOT imply a high phenotypic plasticity and capability to respond to different limnetic transition habitats, which we consider to be crucial for the evolutionary landmark terrestrialization.
Supplementary Material
Acknowledgments
We sincerely thank Professor M. Schagerl, University of Vienna, Biocenter, Austria, for providing access to his Imaging-PAM. S. Obwegeser, University of Innsbruck, is acknowledged for help in TEM sectioning and image generation; FH Professor A. Jäger, Applied University Upper Austria in Wels, for access to the Horiba ICP and K. Krennhuber for help in element measurements.
SUPPLEMENTARY DATA
FUNDING
This study was funded by (FWF) projects and to A.H.
Conflict of interest. None declared.
REFERENCES
- Aigner S, Remias D, Karsten U, et al. Unusual phenolic compounds contribute to ecophysiological performance in the purple-colored green alga Zygogonium ericetorum (Zygnematophyceae, Streptophyta) from a high-alpine habitat. J Phycol. 2013;49:648–60. doi: 10.1111/jpy.12075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alston RE. An investigation of the purple vacuolar pigment of Zygogonium ericetorum and the status of ‘algal anthocyanins’ and ‘phycoporphyrins’. Am J Bot. 1958;45:688–92. [Google Scholar]
- Andosch A, Hoftberger M, Lütz C, et al. Subcellular sequestration and impact of heavy mentals on the ultrastructure and physiology of the multicellular freshwater alga Desmidium swartzii. Int J Mol Sci. 2015;16:10389–410. doi: 10.3390/ijms160510389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avwioro G. Histochemical uses of haematoxylin—a review. J Phys Conf Ser. 2011;1:24–34. [Google Scholar]
- Becker B, Marin B. Streptophyte algae and the origin of embryophytes. Ann Bot. 2009;103:999–1004. doi: 10.1093/aob/mcp044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff HW, Bold HC. Phycological studies IV. Some Soil Algae from Enchanted Rock and Related Algal Species. Austin: University of Texas Publications; 1963. pp. 1–95. [Google Scholar]
- Briat JF, Ravet K, Arnaud N, et al. New insights into ferritin synthesis and function highlight a link between iron homeostasis and oxidative stress in plants. Ann Bot. 2010;105:811–22. doi: 10.1093/aob/mcp128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Browne BA, McColl JG, Driscoll CT. Aluminum speciation using morin: I. Morin and its complexes with aluminum. J Environ Qual. 1990;19:65–72. [Google Scholar]
- Busch A, Rimbauld B, Naumann B, et al. Ferritin is required for rapid remodeling of the photosynthetic apparatus and minimizes photo-oxidative stress in response to iron-availability in Chlamydomonas reinhardtii. Plant J. 2008;55:201–11. doi: 10.1111/j.1365-313X.2008.03490.x. [DOI] [PubMed] [Google Scholar]
- Cadenas E. Biochemistry of oxygen toxicity. Annu Rev Biochem. 1989;58:79–110. doi: 10.1146/annurev.bi.58.070189.000455. [DOI] [PubMed] [Google Scholar]
- Chang YC, Yamamoto Y, Matsumoto H. Accumulation of aluminium in the cell wall pectin in cultured tobacco (Nicotiana tabacum L.) cells treated with a combination of aluminium and iron. Plant Cell Environ. 1999;22:1009–17. [Google Scholar]
- Crémazy A, Campbell PG, Fortin C. The biotic ligand model can successfully predict the uptake of a trivalent ion by a unicellular alga below pH 6.50 but not above: possible role of hydroxo-species. Environ Sci Technol. 2013;47:2408–15. doi: 10.1021/es3038388. [DOI] [PubMed] [Google Scholar]
- Cuenca G, Herrera R, Merida T. Distribution of aluminium in accumulator plants by X‐ray microanalysis in Richeria grandis Vahl leaves from a cloud forest in Venezuela. Plant Cell Environ. 1991;14:437–41. [Google Scholar]
- De Andrade LRM, Barros LMG, Echevarria GF, et al. Al-hyperaccumulator Vochysiaceae from the Brazilian Cerrado store aluminum in their chloroplasts without apparent damage. Environ Exp Bot. 2011;70:37–42. [Google Scholar]
- Domozych DS, Sørensen I, Popper ZA, et al. Pectin metabolism and assembly in the cell wall of the charophyte green alga Penium margaritaceum. Plant Physiol. 2014;165:105–18. doi: 10.1104/pp.114.236257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eticha D, Stass A, Horst WJ. Cell‐wall pectin and its degree of methylation in the maize root‐apex: significance for genotypic differences in aluminium resistance. Plant Cell Environ. 2005;28:1410–20. [Google Scholar]
- Finney LA, O'Halloran TV. Transition metal speciation in the cell: insights from the chemistry of metal ion receptors. Science. 2003;300:931–6. doi: 10.1126/science.1085049. [DOI] [PubMed] [Google Scholar]
- Foy CD, Chaney RT, White MC. The physiology of metal toxicity in plants. Annl Rev Plant Physiol. 1978;29:511–66. [Google Scholar]
- Gaur JP, Rai LC. Heavy metal tolerance in algae. In: Rai LC, Gaur JP, editors. Algal Adaptation to Environmental Stresses: Physiological, Biochemical and Molecular Mechanisms. New York: Springer; 2001. pp. 363–88. [Google Scholar]
- Godbold DL, Jentschke G, Marschner P. Solution pH modifies the response of Norway spruce seedlings to aluminum. Plant Soil. 1995;171:175–8. [Google Scholar]
- Grieve IC. Variations in chemical composition of the soil solution over a four-year period at an upland site in southwest Scotland. Geoderma. 1990;46:351–62. [Google Scholar]
- Grosberg RK, Strathmann RR. The evolution of multicellularity: a minor major transition? Ann Rev Ecol Evol S. 2007;38:621–54. [Google Scholar]
- Harholt J, Moestrup Ø, Ulvskov P. Why plants were terrestrial from the beginning. Trends Plant Sci. 2015 doi: 10.1016/j.tplants.2015.11.010. [DOI] [PubMed] [Google Scholar]
- Hawes I. Effects of freezing and thawing on a species of Zygnema (Chlorophyta) from the Antarctic. Phycologia. 1990;29:326–31. [Google Scholar]
- Herburger K, Holzinger A. Localization and quantification of callose in the streptophyte green algae Zygnema and Klebsormidium: correlation with desiccation tolerance. Plant Cell Physiol. 2015;56:2259–70. doi: 10.1093/pcp/pcv139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herburger K, Karsten U, Holzinger A. Entransia and Hormidiella, sister lineages of Klebsormidium (Streptophyta), respond differently to light, temperature, and desiccation stress. Protoplasma. 2016 doi: 10.1007/s00709-015-0889-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herburger K, Lewis LA, Holzinger A. Photosynthetic efficiency, desiccation tolerance and ultrastructure in two phylogenetically distinct strains of alpine Zygnema sp. (Zygnematophyceae, Streptophyta): role of pre-akinete formation. Protoplasma. 2015;252:571–89. doi: 10.1007/s00709-014-0703-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holzinger A, Karsten U. Desiccation stress and tolerance in green algae: consequences for ultrastructure, physiological, and molecular mechanisms. Front Plant Sci. 2013;4 doi: 10.3389/fpls.2013.00327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holzinger A, Roleda M, Lütz C. The vegetative arctic green alga Zygnema is insensitive to experimental UV exposure. Micron. 2009;40:831–8. doi: 10.1016/j.micron.2009.06.008. [DOI] [PubMed] [Google Scholar]
- Holzinger A, Tschaikner A, Remias D. Cytoarchitecture of the desiccation-tolerant green alga Zygogonium ericetorum. Protoplasma. 2010;243:15–24. doi: 10.1007/s00709-009-0048-5. [DOI] [PubMed] [Google Scholar]
- Hoppert M, Reimer R, Kemmling A, et al. Structure and reactivity of a biological soil crust from a xeric sandy soil in central Europe. Geomicrobiol J. 2004;21:183–91. [Google Scholar]
- Horst WJ, Schmohl N, Kollmeier M, et al. Does aluminium affect root growth of maize through interaction with the cell wall – plasma membrane – cytoskeleton continuum? Plant Soil. 1999;215:163–74. [Google Scholar]
- Kampfenkel K, Van Montagu M, Inzé D. Effects of iron excess on Nicotiana plumbaginifolia plants (implications to oxidative stress) Plant Physiol. 1995;107:725–35. doi: 10.1104/pp.107.3.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan F, Lewis LA, Herburger K, et al. Osmotic stress in the Arctic and Antarctic green alga Zygnema sp. (Zygnemtales, Streptophyta): effects on photosynthesis and ultrastructure. Micron. 2013;44:317–30. doi: 10.1016/j.micron.2012.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan F, Lewis LA, Wastian J, et al. Plasmolysis effects and osmotic potential of two phylogenetically distinct alpine strains of Klebsormidium (Streptophyta) Protoplasma. 2012;249:789–804. doi: 10.1007/s00709-011-0324-z. [DOI] [PubMed] [Google Scholar]
- Karlusich JJP, Lodeyro AF, Carrillo N. The long goodbye: the rise and fall of flavodoxin during plant evolution. J Exp Bot. 2014;65:5161–78. doi: 10.1093/jxb/eru273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karsten U, Herburger K, Holzinger A. Dehydration, temperature and light tolerance in members of the aeroterrestrial green algal genus Interfilum (Streptophyta) from biogeographically different temperate soils. J Phycol. 2014;50:804–16. doi: 10.1111/jpy.12210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimmek S, Stan HJ, Wilke A, et al. Comparative analysis of the biosorption of cadmium, lead, nickel, and zinc by algae. Environ Sci Technol. 2001;35:4283–8. doi: 10.1021/es010063x. [DOI] [PubMed] [Google Scholar]
- Kranner I, Birtić S. A modulating role for antioxidants in desiccation tolerance. Integr Comp Biol. 2005;45:734–40. doi: 10.1093/icb/45.5.734. [DOI] [PubMed] [Google Scholar]
- Lagerheim G. Über das Phycoporphyrin, einen Conjugatenfarbstoff. Videnskab.-Selsk. Skrifter, I. Vol. 5. Mat.-Nat. Kl; 1895. pp. 3–25. [Google Scholar]
- Lürling M. Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction of defensive morphology. Ann Limnol Int J Lim. 2003;39:85–101. [Google Scholar]
- Ma JF, Ryan PR, Delhaize E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001;6:273–8. doi: 10.1016/s1360-1385(01)01961-6. [DOI] [PubMed] [Google Scholar]
- Mertens J, Smolders E. Zinc. In: Alloway BJ, editor. Heavy Metals in soils. Trace Metals and Metalloids in Soils and their Bioavailability. New York: Springer; 2013. pp. 465–93. [Google Scholar]
- Nagajyoti PC, Lee KD, Sreekanth TVM. Heavy metals, occurrence and toxicity for plants: a review. Environ Chem Lett. 2010;8:199–216. [Google Scholar]
- Navascués J, Pérez‐Rontomé C, Sánchez DH, et al. Oxidative stress is a consequence, not a cause, of aluminum toxicity in the forage legume Lotus corniculatus. New Phytol. 2012;193:625–36. doi: 10.1111/j.1469-8137.2011.03978.x. [DOI] [PubMed] [Google Scholar]
- Newsome AG, Murphy BT, van Breemen RB. Isolation and characterization of natural blue pigments from underexplored sources. Phys Meth Food Analysis. 2013:105–25. [Google Scholar]
- Newsome AG, van Breemen RB. Characterization of the purple vacuolar pigment of Zygogonium ericetorum alga. Planta Med. 2012;78:PJ20. [Google Scholar]
- Perales-Vela HV, Peña-Castro JM, Canizares-Villanueva RO. Heavy metal detoxification in eukaryotic microalgae. Chemosphere. 2006;64:1–10. doi: 10.1016/j.chemosphere.2005.11.024. [DOI] [PubMed] [Google Scholar]
- Pichrtová M, Hajek T, Elster J. Osmotic stress and recovery in field populations of Zygnema sp. (Zygnematophyceae, Streptophyta) on Svalbard (High Arctic) subjected to natural desiccation. FEMS Microbiol Ecol. 2014a;89:270–80. doi: 10.1111/1574-6941.12288. [DOI] [PubMed] [Google Scholar]
- Pichrtová M, Kulichová J, Holzinger A. Nitrogen limitation and slow drying induce desiccation tolerance in conjugating green algae (Zygnematophyceae, Streptophyta) from polar habitats. PLoS One. 2014b;9:e113137. doi: 10.1371/journal.pone.0113137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pichrtová M, Remias D, Lewis LA, et al. Changes in phenolic compounds and cellular ultrastructure of arctic and Antarctic strains of Zygnema (Zygnematophyceae, Streptophyta) after exposure to experimentally enhanced UV to PAR ratio. Microb Ecol. 2013;65:6–83. doi: 10.1007/s00248-012-0096-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polle E, Konzak CF, Kattrick JA. Visual detection of aluminum tolerance levels in wheat by hematoxylin staining of seedling roots. Crop Sci. 1978;18:823–7. [Google Scholar]
- Popper ZA, Michel G, Hervé C , et al. Evolution and diversity of plant cell walls: from algae to flowering plants. Annu Rev Plant Biol. 2011;62:567–90. doi: 10.1146/annurev-arplant-042110-103809. [DOI] [PubMed] [Google Scholar]
- Porra RJ, Thompson WA, Kriedmann PE. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA Bioenerg. 1989;975:384–94. [Google Scholar]
- Prasad KVSK, Saradhi PP, Sharmila P. Concerted action of antioxidant enzymes and curtailed growth under zinc toxicity in Brassica juncea. Environ and Exp Bot. 1999;42:1–10. [Google Scholar]
- Ravet K, Touraine B, Boucherez J, et al. Ferritins control interaction between iron homeostasis and oxidative stress in Arabidopsis. Plant J. 2009;57:400–12. doi: 10.1111/j.1365-313X.2008.03698.x. [DOI] [PubMed] [Google Scholar]
- Remias D, Albert A, Lütz C. Effects of realistically simulated, elevated UV irradiation on photosynthesis and pigment composition of the alpine snow alga Chlamydomonas nivalis and the arctic soil alga Tetracystis sp. (Chlorophyceae) Photosynthetica. 2010;48:269–77. [Google Scholar]
- Remias D, Schwaiger S, Aigner S, et al. Characterization of an UV- and VIS-absorbing, purpurogallinderived secondary pigment new to algae and highly abundant in Mesotaenium berggrenii (Zygnematophyceae, Chlorophyta), an extremophyte living on glaciers. FEMS Microbiol Ecol. 2011;79:638–48. doi: 10.1111/j.1574-6941.2011.01245.x. [DOI] [PubMed] [Google Scholar]
- Rengel Z. Role of calcium in aluminium toxicity. New Phytol. 1992;121:499–513. doi: 10.1046/j.1469-8137.2003.00821.x. [DOI] [PubMed] [Google Scholar]
- Schmohl N, Horst WJ. Cell wall pectin content modulates aluminium sensitivity of Zea mays (L.) cells grown in suspension culture. Plant Cell Environ. 2000;23:735–42. [Google Scholar]
- Schmohl N, Pilling J, Fisahn J, et al. Pectin methylesterase modulates aluminium sensitivity in Zea mays and Solanum tuberosum. Physiol Plant. 2000;109:419–27. [Google Scholar]
- Škaloud P. Species composition and diversity of aero-terrestrial algae and cyanobacteria of the Boreč Hill ventaroles. Fottea. 2009;9:65–80. [Google Scholar]
- Stamenković M, Hanelt D. Protection strategies of Cosmarium strains (Zygnematophyceae, Streptophyta) isolated from various geographic regions against excessive photosynthetically active radiation. Photochem Photobiol. 2013;89:900–10. doi: 10.1111/php.12083. [DOI] [PubMed] [Google Scholar]
- Stancheva R, Hall JD, Herburger K, et al. Phylogenetic position of Zygogonium ericetorum (Zygnematophyceae, Charophyta) from a high alpine habitat and ultrastructural characterization of unusual aplanospores. J Phycol. 2014;50:790–803. doi: 10.1111/jpy.12229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stancheva R, Herburger K, Sheath RG, et al. Conjugation morphology of Zygogonium ericetorum (Zygnematophyceae, Charophyta) from a high alpine habitat. J Phycol. 2016;52:131–4. doi: 10.1111/jpy.12363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stancheva R, Sheath RG, Hall JD. Systematics of the genus Zygnema (Zygnematophyceae, Charophyta) from Californian watersheds. J Phycol. 2012;48:409–22. doi: 10.1111/j.1529-8817.2012.01127.x. [DOI] [PubMed] [Google Scholar]
- Taylor GJ, McDonald-Stephens JL, Hunter DB, et al. Direct measurement of aluminum uptake and distribution in single cells of Chara corallina. Plant Physiol. 2000;123:987–96. doi: 10.1104/pp.123.3.987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tice KR, Parker DR, DeMason DA. Operationally defined apoplastic and symplastic aluminum fractions in root tips of aluminum-intoxicated wheat. Plant Physiol. 1992;100:309–18. doi: 10.1104/pp.100.1.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Transeau EN. The Zygnemataceae. Columbus, Ohio: The Ohio State University Press; 1951. [Google Scholar]
- Volland S, Andosch A, Milla M, et al. Intracellular metal compartmentalization in the green algal model system Micrasterias denticulata (Streptophyta) measured by transmission electron microscopy-coupled electron energy loss spectroscopy. J Phycol. 2011;47:565–79. doi: 10.1111/j.1529-8817.2011.00988.x. [DOI] [PubMed] [Google Scholar]
- Waldo GS, Wright E,Whang ZH, et al. Formation of the ferritin iron mineral occurs in plastids. Plant Physiol. 1995;109:797–802. doi: 10.1104/pp.109.3.797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waters ER. Molecular adaptation and the origin of land plants. Mol Phylogen Evol. 2003;29:456–63. doi: 10.1016/j.ympev.2003.07.018. [DOI] [PubMed] [Google Scholar]
- Wickett NJ, Mirarab S, Nguyen N, et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. P Natl Acad Sci USA. 2014;111:E4859–68. doi: 10.1073/pnas.1323926111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willats WG, Orfila C, Limberg G, et al. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls. Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J Biol Chem. 2001;276:19404–13. doi: 10.1074/jbc.M011242200. [DOI] [PubMed] [Google Scholar]
- Walsby AE. Numerical integration of phytoplankton photosynthesis through time and depth in a water column. New Phytol. 1997;136:189–209. [Google Scholar]
- Webb WL, Newton M, Starr D. Carbon dioxide exchange of Alnus rubra: a mathematical model. Oecologia. 1974;17:281–91. doi: 10.1007/BF00345747. [DOI] [PubMed] [Google Scholar]
- World Health Organization. Guidelines for Drinking-Water Quality. Health Criteria and Other Supporting Information. Geneva: World Health Organization; 1996. [Google Scholar]
- Zancani M, Peresson C, Biroccio A, et al. Evidence for the presence of ferritin in plant mitochondria. Eur J Biochem. 2004;271:3657–64. doi: 10.1111/j.1432-1033.2004.04300.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.