

ABSTRACT
We have recently demonstrated that the Nlrp3 inflammasome can detect the presence of colorectal cancer (CRC) metastatic growth in the liver and limit its growth. Inflammasome signaling primed natural killer (NK) cells through Interleukin (IL)-18 and activated their ability to trigger FasL-induced apoptosis of the tumor.
KEYWORDS: Apoptosis, cancer, Fas, immunosurveillance, immunotherapy, innate immunity, NK cells, Nlrp3 inflammasome
Cancer immunosurveillance is a process in which both innate and adaptive immune cells cooperate to prevent tumor development. Upon sensing missing or altered self or the presence of tumor-derived danger signals, innate immune cells such as NK cells, macrophages and dendritic cells (DCs) provide first line defenses by directly killing tumor cells, secreting death-inducing cytokines or activating adaptive immunity by presenting tumor antigens to T cells including cytotoxic T lymphocytes (CTLs). In contrast to innate immunity, which is immediate and broad, adaptive immunity, mediated by T and B cells, provide long-lived antigen-specific responses. NK cells are important mediators of innate immunity and have recently been classified as a subset of cytotoxic innate lymphocytes (ILCs) that express the transcription factor NFIL3. As their name indicates, NK cells were initially discovered based on their capacity to kill cancer cells. In humans, low-circulating NK cell number is associated with increased cancer risk, whereas enhanced NK cell infiltration in tumor tissues is linked to better prognosis.1
A new frontier in cancer treatment is targeted immunotherapy that aims at boosting antitumor immune responses. Key clinical trials of immunotherapies have thus far been conducted with immune checkpoint inhibitors (antibodies against programmed cell death protein 1 (PD1), CTLA4 and others), co-stimulatory monoclonal antibodies, cancer vaccines and adoptive T cell therapy.2 These strategies, which modulate adaptive immunity, have shown efficacy in reducing tumor lesions as monotherapies, combination immunotherapies or in conjunction with conventional therapies (radiotherapy, chemotherapy, anti-angiogenics). The increasing evidence that NK cells can be educated and even generate long-term memory has rendered the prospect of targeting these cells of the innate immune system similarly attractive for immunotherapy. Current NK-based clinical trials include the administration of monoclonal antibodies against NK cell inhibitory receptors or the adoptive transfer of expanded allogenic NK cells, NK stable cell lines, such as NK-92 and KHYG-1, or NK cells genetically engineered to overexpress cytokines, chimeric tumor antigen receptors or Fc receptors.1
In addition to missing self- and altered self-recognition by inhibitory and activating receptors, respectively, a commensurate NK cell response depends on a continuum of paracrine signals for cell maturation and pre-activation, among which is the cytokine IL-18. The production of IL-18 depends on caspase-1 activation within the inflammasome, a macromolecular complex scaffolded by cytosolic pattern recognition receptors including the Nod-like receptors (NLRs), following the perception of microbial or sterile danger signals.3 A number of recent studies have implicated the inflammasome pathway in tumor immunology, ascribing seemingly contradictory roles in tumorigenesis versus tumor control.4 For instance, we and others have previously demonstrated that inflammasome IL-18 signaling regulates intestinal tissue repair following damage and CRC development by triggering the process of re-epithelialization.5 IL-18 signaling was also reported to favor metastasis and tumor growth by enhancing endothelial cell adhesion and vascular arrest of tumor cells,6 and by instating an immunoablative phenotype in NK cells through the expression of the immunosuppressive molecule PD-1.7 In contrast to these pro-tumorigenic effects, the inflammasome pathway also contributes to immunosurveillance, in combination with chemotherapies that induce immunogenic cell death8 or by synergizing with TLR signaling to induce effective antitumor adaptive immunity.9
Using an experimental model of CRC metastasis to the liver, we recently uncovered an innate immunity circuitry that connects cancer-induced Nlrp3 inflammasome activation in Kupffer cells to efficient NK cell tumoricidal activity.1 We demonstrated that the signal mediating the communication between these two innate immune cell populations is IL-18. Mice deficient in components of the Nlrp3 inflammasome (Nlrp3-/-, Ice-/-, Il18-/- and Il18r-/-) had a higher metastatic burden compared to wild-type (WT) mice. Antibiotics-mediated depletion of the microbiota did not impact tumor burden, suggesting that inflammasome activation was independent of the microbiota. Instead, cancer-derived factors primed and activated the Nlrp3 inflammasome leading to IL-18 maturation by caspase-1. IL-18 exerts anticancer effects though several mechanisms, including the production of IFNγ, which promotes a cancer-eradicating Th1 response, or by stimulating the cytotoxic activity of CTLs and NK cells. In our model, IL-18 restricted CRC metastatic growth in the liver independently of the adaptive immune system or IFNγ. Whereas Rag1-/- mice, which lack T, B and NKT cells, exhibited similar tumor control compared to WT mice, Rag1-/-/Il18-/- mice had an increased metastatic burden, suggesting that IL-18-induced tumor cytotoxicity independently of adaptive immunity effectors like CTLs. IL-18 was rather required for hepatic NK cell maturation and activation. Ablation of inflammasome signaling did not alter the capacity of hepatic NK cells to produce IFNγ, but resulted in decreased FasL surface expression. We propose a model in which the Nlrp3 inflammasome in Kupffer cells cooperates with NK cells in sensing carcinogenesis and boosts their tumoricidal activity through FasL-induced apoptosis. By identifying the nature of the innate immune system sensor and the mechanism by which this factor orchestrates tumor killing, our findings could have important implications for the further development of cancer immunotherapy, enabling potential therapeutic strategies that could harness this pathway to elicit tumor killing.
Figure 1.
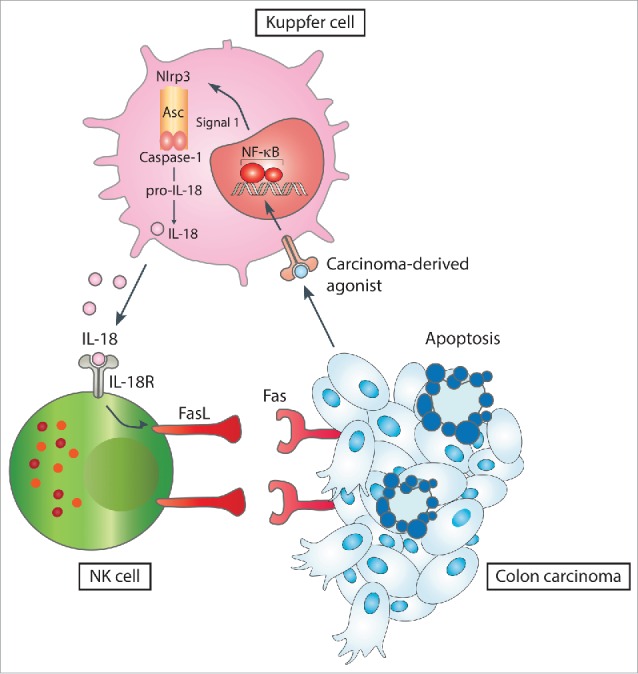
A regulatory circuitry within the innate immune system that links inflammasome stimulation by cancer-associated agonists to activation of NK cell tumoricidal function leading to tumor elimination at metastatic sites.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- 1.Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat. Rev. (2012); 12:239–52; PMID:22437937; http://dx.doi.org/ 10.1038/nri3174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Melero I. Berman DM, Aznar MA, Korman AJ, Pérez Gracia JL, Haanen J. Evolving synergistic combinations of targeted immunotherapies to combat cancer. Nat. Rev. Cancer (2015); 15:457–72; PMID:26205340; http://dx.doi.org/ 10.1038/nrc3973 [DOI] [PubMed] [Google Scholar]
- 3.Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat. Rev. (2013); 13:397–411; PMID:23702978; http://dx.doi.org/ 10.1038/nri3452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kent A, Blander JM. Nod-like receptors: key molecular switches in the conundrum of cancer. Front. Immunol. (2014); 5:185; PMID:24795727; http://dx.doi.org/ 10.3389/fimmu.2014.00185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Saleh M, Trinchieri G. Innate immune mechanisms of colitis and colitis-associated colorectal cancer. Nat. Rev. (2011); 11:9–20; PMID:21151034; http://dx.doi.org/ 10.1038/nri2891 [DOI] [PubMed] [Google Scholar]
- 6.Carrascal MT. Mendoza L, Valcárcel M, Salado C, Egilegor E, Tellería N, Vidal-Vanaclocha F, Dinarello CA. Interleukin-18 binding protein reduces b16 melanoma hepatic metastasis by neutralizing adhesiveness and growth factors of sinusoidal endothelium. Cancer Res. (2003); 63:491–7; PMID:12543807 [PubMed] [Google Scholar]
- 7.Terme M, Ullrich E, Aymeric L, Meinhardt K, Coudert JD, Desbois M, Ghiringhelli F, Viaud S, Ryffel B, Yagita H, et al. Cancer-induced immunosuppression: IL-18-elicited immunoablative NK cells. Cancer Res. (2012); 72:2757–67; PMID:22427351; http://dx.doi.org/ 10.1158/0008-5472.CAN-11-3379 [DOI] [PubMed] [Google Scholar]
- 8.Ghiringhelli F, Apetoh L, Tesniere A, Aymeric L, Ma Y, Ortiz C, Vermaelen K, Panaretakis T, Mignot G, Ullrich E, et al. Activation of the NLRP3 inflammasome in dendritic cells induces IL-1beta-dependent adaptive immunity against tumors. Nat. Med. (2009); 15:1170–8; PMID:19767732; http://dx.doi.org/ 10.1038/nm.2028 [DOI] [PubMed] [Google Scholar]
- 9.Garaude J, Kent A, van Rooijen N, Blander JM. Simultaneous targeting of toll- and nod-like receptors induces effective tumor-specific immune responses. Sci Transl Med. (2012); 8;4(120): 120ra16; PMID:22323829; http://dx.doi.org/26384545 10.1126/scitranslmed.3002868 [DOI] [PubMed] [Google Scholar]
- 10.Dupaul-Chicoine J, Arabzadeh A, Dagenais M, Douglas T, Champagne C, Morizot A, Rodrigue-Gervais IG, Breton V, Colpitts SL, Beauchemin N, et al. The Nlrp3 inflammasome suppresses colorectal cancer metastatic growth in the liver by promoting natural killer cell tumoricidal activity. Immunity (2015); 43:751–63; PMID:26384545; http://dx.doi.org/ 10.1016/j.immuni.2015.08.013 [DOI] [PubMed] [Google Scholar]