

Abstract
Somatosensory nerves transduce thermal, mechanical, chemical, and noxious stimuli caused by both endogenous and environmental agents. The cell bodies of these afferent neurons are located within the sensory ganglia. Sensory ganglia innervate a specific organ or portion of the body. For instance, the dorsal root ganglia (DRG) are located in the vertebral column and extend processes throughout the body and limbs. The trigeminal ganglia are located in the skull and innervate the face, and upper airways. Vagal afferents of the nodose ganglia extend throughout the gut, heart, and lungs. The nodose neurons control a diverse array of functions such as: respiratory rate, airway irritation, and cough reflexes. Thus, to understand and manipulate their function, it is critical to identify and isolate airway specific neuronal sub-populations. In the mouse, the airways are exposed to a fluorescent tracer dye, Fast Blue, for retrograde tracing of airway-specific nodose neurons. The nodose ganglia are dissociated and fluorescence activated cell (FAC) sorting is used to collect dye positive cells. Next, high quality ribonucleic acid (RNA) is extracted from dye positive cells for next generation sequencing. Using this method airway specific neuronal gene expression is determined.
Keywords: Neuroscience, Issue 110, Sensory neuroscience, nodose ganglia, mouse, lungs, retrograde fluorescence tracing, Fast Blue, fluorescence activated cell sorting (FACS), RNA sequencing
Introduction
Somatosensory nerves transduce thermal, mechanical, chemical, and noxious stimuli caused by both endogenous and environmental agents. The cell bodies of these afferent nerves are located in sensory ganglia, such as the dorsal root, trigeminal, or nodose ganglia. Each sensory ganglion innervates specific regions of the body and contains cells that innervate separate organs and tissues within that region. For instance, the dorsal root ganglia (DRG) are located in the vertebral column and extend processes throughout the body and limbs, while the trigeminal ganglia are located in the skull, containing neurons that innervate the face, eyes, meninges or upper airways1,2. The nodose ganglia of the vagus nerve is located in the neck below the skull and contains cell bodies that extend nerve fibers throughout the gastrointestinal tract, heart, and lower airways and lungs3. In humans the nodose ganglion stands alone, however, in the mouse it is fused with the jugular ganglion, which also innervates the lungs4. This fused ganglion is often called the jugular/nodose complex, vagal ganglion, or simply nodose ganglion5. Here, it is referred to as the nodose ganglion.
Afferent fibers of the nodose pass information from the viscera to the nucleus of the solitary tract (NTS) in the brainstem. Sensory input to this unique ganglion controls a diverse array of functions, such as gut motility6, heart rate7, respiration8,9, and irritant-activated respiratory responses10,11. With this diversity of functions and innervated organs, it is critical to target and isolate organ-specific subpopulations of the nodose ganglion in order to study individual neuronal pathways. However, given the small size of the nodose and the limited number of neurons it contains this is not a trivial task. Each mouse nodose ganglion contains roughly 5,000 neurons12 in addition to an extensive population of supporting satellite cells. Of the 5,000 nodose neurons, only 3 - 5% innervate the airways. Therefore, any functional, morphological or molecular changes within airway-innervating neurons, due to respiratory stimulation or pathologies, will be lost in the densely packed nodose ganglion.
To solve this problem, a method was developed to identify and isolate neurons that innervate the airways. The airways were exposed to a fluorescent tracer dye to identify the subsequent innervating nodose neurons. Fast Blue was picked up by neurons and travels quickly to their cell bodies where it is retained for up to eight weeks13-15. Once identified, a gentle, yet efficient, dissociation protocol was used to preserve dye labeling and cell viability for fluorescent activated cell (FAC) sorting. Sorted cells are used to extract high quality ribonucleic acid (RNA) to determine gene expression or for other downstream molecular analysis. This protocol provides a useful and robust technique for isolating sensory neurons that innervate a tissue of interest.
Protocol
Procedures involving animal subjects have been approved by the Institutional Animal Care and Use Committee (IACUC) of Duke University.
1. Intranasal Administration of Fast Blue
For Fast Blue, administer the dye at least 2 days before euthanizing the mouse. The dye will persist for up to eight weeks.
Anesthetize the mouse with light inhalation anesthesia (2.5% sevoflurane) until breathing starts to slow.
Use a 200 µl pipette with filtered tips to slowly instill 40 µl of dye solution (0.4 mM Fast Blue, 1% dimethyl sulfoxide, DMSO, in phosphate buffered saline, PBS) into the nostril, pausing occasionally to ensure the mouse is aspirating the solution (Figure 1A).
Hold the mouse vertically, head up, and gently massage the chest to ensure the dye spreads throughout the lungs.
2. Preparations on Dissection and Analysis Day
Prepare 10 ml Ganglia Dissociation Solution (GaDS) by combining the ingredients as specified in Table 1.
Perform RNA extraction in a dedicated lab area. Before beginning the experiment, clean surfaces with 70% ethanol, 10% bleach, and RNase decontamination reagent. This includes the RNA centrifuge, any tube racks, and pipettes. Ensure there are dedicated filter tip boxes for RNA use only. Do not use this equipment and area for DNA preparation, as this will contaminate the samples.
Mix fresh 70%, and 80% ethanol with RNase-free water. Prepare 1 ml per sample to be used in RNA extraction.
| Ingredient | Amount |
| Advanced DMEM/f12 | 9.5 ml |
| glutamine | 100 µl |
| HEPES (10 mM) | 100 µl |
| N2 supplement | 100 µl |
| B27 (no vitamin A) | 200 µl |
| NGF (50 µg/ml) | 10 µl |
Table 1. Reagent Mixture for Ganglia Dissociation Solution (GaDS).
3. Dissection Procedure
Fill 1.5 ml tubes with 500 µl GaDS, one for each experimental sample, one for a positive sorting control, and one for a negative sorting control. Store tubes on ice throughout dissection.
Dissect out the nodose ganglion and make two partial cuts to facilitate dissociation, then put it in the experimental sample tube. Please refer to the following JoVE protocol for dissection of the nodose ganglia16 and DRG17.
Dissect and partially cut non-labeled ganglia, such as the dorsal root ganglia (DRG), for the sorting controls. Use two ganglia for the positive control and two ganglia for the negative control.
To obtain enough nodose neurons for successful sorting, combine ganglia from 5 mice into one experimental sample tube containing GaDS.
4. Sensory Ganglia Dissociation
Pipette out 500 µl GaDS, leaving ganglia in the tube. Wash ganglia once by adding 1 ml PBS, wait for ganglia to settle to the bottom of the tube, then pipette out PBS.
Add 1 ml GaDS and 22 µl of digestion enzyme (collagenase I/II and protease mix, 2.5 mg/ml in H2O) to each tube.
Add 20 µl Fast Blue (5.26 mM) to the positive control tube.
Put the tubes in a water bath or heating block already set at 37 °C. Note: The time of ganglia digestion depends on the age of the digestion enzyme. Within 1 month of dissolving the digestion enzyme, digest for 30 min, within 2 - 6 months digest for 45 min. If the enzyme is over 6 months old, digest ganglia for 60 min.
Every 15 min shake tubes briefly and flick them to ensure ganglia are covered by the solution.
- While ganglia are being digested, prepare density gradients using a particle solution (colloidal silica particles coated with polyvinylpyrrolidone).
- Mix 12% particle solution in GaDS, 500 µl per sample.
- Mix 28% particle solution in GaDS, 500 µl per sample.
- Slowly and carefully add 400 µl 12% density solution on top of 400 µl 28% density solution in a fresh 1.5 ml tube for each sample. Do not allow the two layers to mix. Two distinct layers should be visible in the tube; one darker than the other.
While ganglia are being digested, label new 1.5 ml collection tubes for sorting (one Fast Blue positive and one Fast Blue negative tube per sorting sample). Depending on the sorting facility requirements, these tubes must have detachable lids so as to not interfere with the cell sorter.
After the appropriate digestion time, remove GaDS with digestion enzyme and wash ganglia twice with 1 ml PBS. Add 200 µl of fresh GaDS to each tube.
Use a 200 µl pipette, set to 100 µl volume, and pipette ganglia up and down several times to break cells apart. Repeat until no intact piece of tissue is seen. Avoid creating bubbles in the solution.
Pass dissociated cells through a 70 µm cell strainer.
Add another 100 µl GaDS to the original dissociation tube to pick up any remaining stray cells left and pass the additional solution through the cell strainer. Using a new pipette tip, collect any remaining cell-containing liquid that has passed through the strainer and clung to the mesh. The final volume of cells suspended in GaDS is 300 µl.
Carefully layer the 300 µl of cells on top of the previously made density gradient. Avoid bubbles and mixing of cells with the density layers.
Centrifuge for 10 min at 2,900 x g at RT.
- While cells are in the centrifuge, mix lysis buffer (1 ml per experimental sample) as follows: 10 µl 2-mercaptoethanol in 1 ml Buffer RLT (from RNA extraction kit).
- Add 300 µl lysis buffer to Fast Blue positive collection tube and 600 µl to Fast Blue negative collection tube (tubes labeled from step 4.7). This volume depends on the number of cells expected to be collected. For <2,000 cells, add 300 µl, for >7,000 cells add 600 µl. This will ensure the sorting sheath fluid does not significantly dilute the lysis buffer.
Once centrifugation is complete, carefully remove and discard the top 700 µl layer. This layer contains the majority of cell debris that will otherwise interfere with cell sorting.
Add 700 µl fresh GaDS to the remaining cell containing solution and pipette up and down several times to mix.
Centrifuge for 15 min at 2,900 x g to pellet cells.
While cells are in the centrifuge, mix sorting GaDS (500 µl per sample) by adding 5 µl DNase (10 mg/ml) to 1 ml GaDS.
Once centrifugation is done, carefully remove and discard supernatant, leaving the cell pellet.
Re-suspend cell pellet in 200 - 300 µl sorting GaDS by pipetting up and down with a 1,000 µl pipette set to 200 µl several times. Put samples on ice. Cells are now ready to be sorted.
5. Fluorescence Activated Cell (FAC) Sorting
Note: This section requires operational knowledge of a FACS sorter or the assistance of skilled personnel.
Add 1 µl propidium iodide (PI) stock solution (50 µg/ml) to each sample to test for cell viability. Split negative control into 2 tubes, one with PI and one without. PI binds DNA only in dead cells. If after the first few sorts there are no dead cells to label, the use of PI is not needed.
Transport cells to flow cytometry instrument, e.g., BD FACS AriaII running Diva 8 software, on ice.
Since the cell size is variable in this population, use a large nozzle (100 µm) for sorting. To decrease the amount of stress the cells experience and increase cell viability use low pressure (20 psi).
Use the FACS sorter software to set up the analysis plots, forward scatter, side scatter (SSC), Fast Blue, and PI, if needed.
To re-suspend cells, gently vortex the sample briefly before loading it into the sorter.
Begin with the negative control, no Fast Blue or PI, to set the gate threshold (Figure 2A). This will determine the amount of autofluorescence in the cell population. Optional: Run the negative sample with PI to identify whether there is a population of dead cells.
Sort the positive control sample (incubated with Fast Blue), to set compensation parameters.
Once the gates and parameters are set, begin sorting the experimental samples. Samples are sorted into the prepared collection tubes containing RNA lysis buffer.
Keep sorted samples on ice until the cell sorting is complete.
Begin RNA extraction step immediately after cells are sorted.
6. RNA Extraction
Vortex sorted cells in lysis buffer for 1 min to homogenize.
Add 1 volume of 70% ethanol and mix by pipetting up and down several times.
- Transfer up to 700 µl of the sample to a spin column. Centrifuge for 15 sec at 8,000 x g. Discard the flow-through.
- Repeat until the total volume of the sample is passed through the spin column.
Add 350 µl Buffer RW1 and continue the RNA purification process, including the DNase treatment step, according to the manufacturer's protocol for cells.
Once RNA has been extracted, test the RNA quality using a microfluidic electrophoresis system according to the manufacturer's specifications.
Representative Results
Using this method, airway-innervating neurons are labeled by intranasally instilling Fast Blue (Figure 1A). After two days, Fast Blue labeled cells appear in the nodose ganglia (Figure 1C). These cells make up 3 - 5% of the total neuronal population of the nodose ganglia. Other retrograde dyes that have been used for this purpose include DiI (1,1'-dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate) and Fluorogold. Lung exposure to DiI labels nodose neurons. However, DiI takes longer to reach the ganglion, 7 - 14 days after instillation18,19, therefore an actively transported dye was used. Fluorogold is an actively transported dye that has also been used to label airway neurons14,20. In our experience, however, once this dye reached the cell bodies in the nodose ganglion it was quickly picked up by satellite cells and not neurons (data not shown). Fast Blue is an alternate actively transported fluorescent dye that successfully, and quickly, labels DRG in rats21 and airway-innervating nodose neurons13,14. The Fast Blue approach, therefore, appears more robust and reproducible.
The dissection of the nodose ganglion has been previously described16 and its location is briefly illustrated in Figure 1B. Digestion begins immediately after the nodose ganglia have been excised. To prevent neurons from clumping together and allow for a cleaner sort, use a gentle digestion enzyme, clear any debris and add RNase to the ganglia sorting solution. Multiple digestion enzyme mixes were tested for effectiveness. A mild digestion enzyme mix22 was tried, however, for adult ganglion digestion it was not strong enough to break apart cells in a timely fashion. Alternatively, a previously used23 combination of papain, protease, and collagenase enzyme mixture, proved to be too aggressive and caused undesirable cell death (data not shown). The outlined digestion protocol, with the specified enzyme mix, was found to be the ideal for a timely digestion with high cell viability.
Previous methods have used manual cell picking with a glass pipette to separate labeled cells from unlabeled cells24. This technique is better suited for single cell analysis of a small number of cells. However, with a maximal rate of 48 cells/hr this technique is impractical when whole populations of hundreds or thousands of cells need to be collected. When the goal is to isolate high quality RNA, time becomes a key variable. It is important to begin RNA extraction as quickly as possible to prevent degradation. FAC sorting allows collection of the entire labeled population in ~45 min and RNA extraction is started immediately after.
When sorting neurons it is important to clear as much debris from broken neuronal processes. A density gradient was used to clear the debris from the dissociated samples. It is important to centrifuge at a high enough force to ensure that C-fiber neurons, which have a small radius, ~5 µm24, are forced through the gradient.
Figure 2A and B show the negative and positive controls used to set the sorting gates. As can be seen in Figure 2C, the dissociated Fast Blue labeled nodose cells sort into two populations. The cell numbers collected from this sample are listed in Figure 2D. Sorting efficiency for this method is 73 - 85%. When sorting neurons the forward scatter and side scatter parameters did not add specificity. Since neurons are not spherical these parameters were unable to reliably report cell size.
For high quality RNA yield, cells need to be sorted directly into the lysis buffer and RNA should be extracted immediately after sorting. A microfluidic electrophoresis system was used to test the quality of RNA (Figure 3). The 18S and 28S peaks are determined via gel electrophoresis used to calculate an RNA integrity number (RIN). For RNA sequencing the sample RIN should be greater than 7. This RNA sample is now ready to be sequenced.
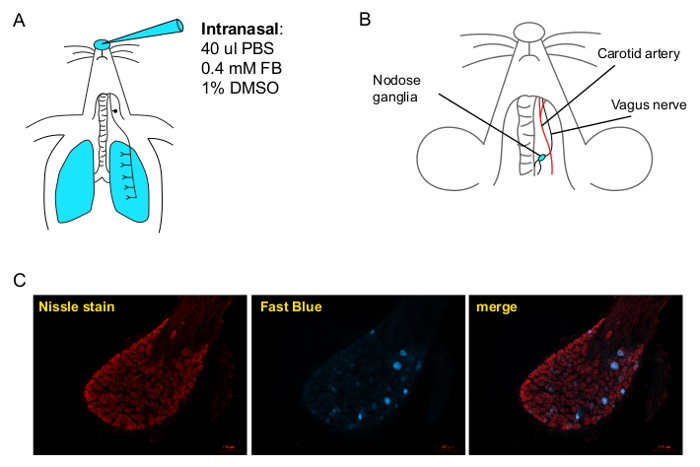 Figure 1. Fast Blue Labeling of Airway Innervating Neurons in the Nodose Ganglia of the Vagus Nerve. A) A representation of Fast Blue intranasal instillation into the lungs. Under light sevoflurane anesthesia, pipette 40 µl of Fast Blue in PBS with 1% DMSO into the nostrils of a mouse, making sure the mouse is aspirating in the fluid. B) A diagram showing where the nodose ganglion is located. It appears as a thickening of the vagus nerve, which runs along the carotid artery. C) After at least 2 days, Fast Blue appears in the nodose ganglia, marking airway neurons. The nodose is counterstained with red fluorescent Nissle stain, which stains all neurons. Scale bar is 100 µm. Please click here to view a larger version of this figure.
Figure 1. Fast Blue Labeling of Airway Innervating Neurons in the Nodose Ganglia of the Vagus Nerve. A) A representation of Fast Blue intranasal instillation into the lungs. Under light sevoflurane anesthesia, pipette 40 µl of Fast Blue in PBS with 1% DMSO into the nostrils of a mouse, making sure the mouse is aspirating in the fluid. B) A diagram showing where the nodose ganglion is located. It appears as a thickening of the vagus nerve, which runs along the carotid artery. C) After at least 2 days, Fast Blue appears in the nodose ganglia, marking airway neurons. The nodose is counterstained with red fluorescent Nissle stain, which stains all neurons. Scale bar is 100 µm. Please click here to view a larger version of this figure.
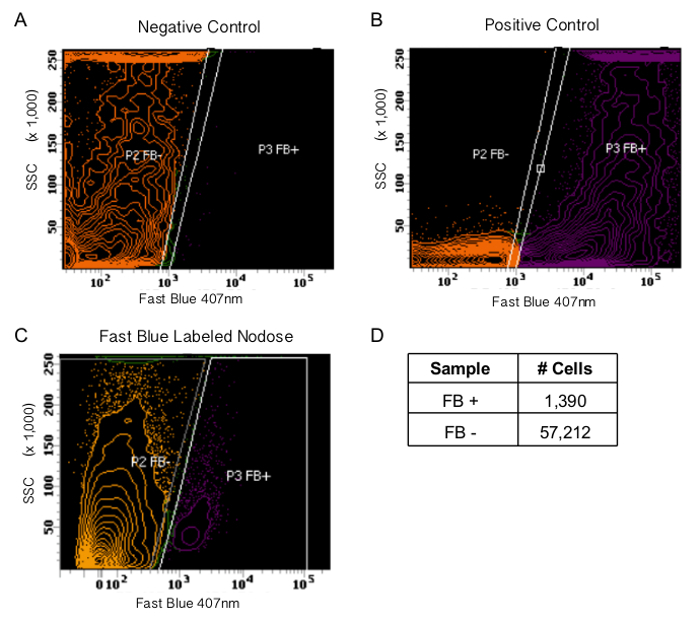 Figure 2. Fluorescence Activated Cell (FAC) Sorting Gates for Fast Blue Positive Cells. To set up sorting gates use the negative (A) and positive (B) controls to establish the Fast Blue positive population. The negative control (A) is dissociated DRG cells that do not innervate the lungs. The positive control (B) is dissociated DRG cells that are incubated with Fast Blue in vitro, after they have been dissected. Due to the non-spherical shape of dissociated neurons, the forward scatter and side scatter parameters will not help define the cell population. C) A representative sort of dissociated nodose ganglia cells of mice whose airways have been exposed to Fast Blue. D) The cell count for (C). Over 1,000 Fast Blue (FB) cells were collected. Please click here to view a larger version of this figure.
Figure 2. Fluorescence Activated Cell (FAC) Sorting Gates for Fast Blue Positive Cells. To set up sorting gates use the negative (A) and positive (B) controls to establish the Fast Blue positive population. The negative control (A) is dissociated DRG cells that do not innervate the lungs. The positive control (B) is dissociated DRG cells that are incubated with Fast Blue in vitro, after they have been dissected. Due to the non-spherical shape of dissociated neurons, the forward scatter and side scatter parameters will not help define the cell population. C) A representative sort of dissociated nodose ganglia cells of mice whose airways have been exposed to Fast Blue. D) The cell count for (C). Over 1,000 Fast Blue (FB) cells were collected. Please click here to view a larger version of this figure.
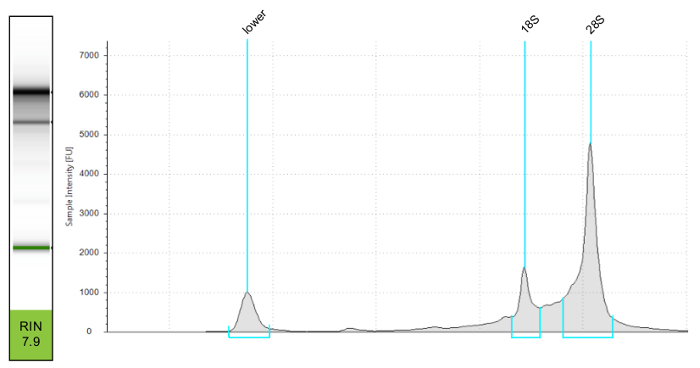 Figure 3. Assessment of RNA Quality and Assignment of RNA Integrity Number (RIN) Using a Microfluidic Electrophoresis System. Using an electrophoresis system the quality of RNA is calculated from the 18S and 28S peaks on the pictured gel image. The lower peak represents degraded RNA. Please click here to view a larger version of this figure.
Figure 3. Assessment of RNA Quality and Assignment of RNA Integrity Number (RIN) Using a Microfluidic Electrophoresis System. Using an electrophoresis system the quality of RNA is calculated from the 18S and 28S peaks on the pictured gel image. The lower peak represents degraded RNA. Please click here to view a larger version of this figure.
Discussion
This protocol describes a method to target airway-innervating neurons in the nodose ganglia of the vagus nerve. Once labeled, the ganglia are gently dissociated to optimally preserve cell numbers and viability. These neurons are then FAC sorted directly into lysis buffer and RNA is extracted. The significance of this protocol is the ability to target, isolate, and preserve the quality of a specific sensory cell population. Gene expression is described in this small population of neurons, and organ-specific functions and neural networks are identified.
The critical steps in this protocol include rapid progression through the dissociation process and subsequent cell sorting. It is important to schedule the sorting time such that it begins immediately after dissociation. It is also critical that all the RNA extraction reagents are fresh, the 70% and 80% ethanol solutions are made fresh, and all tubes and equipment for RNA extraction are clean and RNase free.
This protocol can be modified such that cells are sorted individually into well plates for single cell RNA extraction and sequencing. It can also be used to isolate cells from other sensory ganglia, such as dorsal root ganglia (DRGs). Dye injection of different body parts for tracing back to DRGs has been previously described17. This protocol can also be modified to collect cells for functional analysis. Instead of sorting cells into lysis buffer, cells are sorted into neuronal culture medium, GaDS. These cells are then cultured and used for functional studies such as calcium imaging, or electrophysiology.
If the Fast Blue positive population is smaller than the expected outcome, purchase new digestion enzyme. The age of the digestion enzyme is correlated to the efficiency of digestion. The digestion enzyme needs to be used within 1 year of purchase. In the case where the cell number is high, RNA is extracted and the quality tested. If the RNA is degraded this is an indication that the reagents are not fresh, or have been contaminated. In this case, be sure to thoroughly clean all RNA extraction areas, pipettes, racks, and anything that will come in contact with the sample tubes. Make fresh RNase and DNase free 70% and 80% ethanol. The water and ethanol used to make these solutions should also be kept separate from other lab supplies and used solely for RNA extraction.
FAC sorting is a fast and efficient method for isolating Fast Blue positive cells, however, one limitation is that the cell density should be 1 to 2 million cells per ml buffer. This protocol pushes the limit of current cell sorter models. The number of cells in the nodose ganglion is small. To get a cell yield sufficient for the cell sorter, cells from five animals are pooled. The sorting volume described is 200 - 300 µl and has fewer than 100,000 suspended cells. This translates to a concentration of around 450,000 cells per ml, below the recommended density. One alternative to using a cell sorter is to handpick with a fluorescent microscope and glass pipette24. This technique is slower and has been used to collect 30 - 100 cells at a time. Therefore, this protocol offers a faster high-throughput method compared to existing methods.
The future applications of this protocol include the ability to determine the transcriptional profile of a specific population of airway-innervating nerves. The high quality RNA collected using this protocol can be used as a template for cDNA synthesis and next generation sequencing, and other techniques to characterize the transcriptome of the collected neurons. In this way, it can be determined which genes or pathways are specific to these neurons. This information will help to elucidate the functional role these neurons play in normal physiological conditions. Once basic expression patterns have been established, the system can be challenged by physical and chemical stimuli or by pathogens to determine their effects on gene regulation in lung-innervating neurons. Finally, by identifying which genes are regulated we can identify new pharmacological targets and start to investigate the effects of therapeutics to interfere with acute and chronic changes of nerve function in airway diseases.
Disclosures
The authors have nothing to disclose.
Acknowledgments
Supported by NIH grant R01HL105635 to SEJ. The authors would like to thank Diego V. Bohórquez for technical advice. We also thank R. Ian Cumming for technical assistance and performing the flow cytometry at the Duke Human Vaccine Institute Research Flow Cytometry Shared Resource Facility (Durham, NC). Flow cytometry was performed in the Regional Biocontainment Laboratory at Duke which received partial support for construction from the National Institutes of Health, National Institute of Allergy and Infectious Diseases (UC6-AI058607).
References
- Manteniotis S, et al. Comprehensive RNA-Seq Expression Analysis of Sensory Ganglia with a Focus on Ion Channels and GPCRs in Trigeminal Ganglia. PLoS One. 2013;8(11):1–30. doi: 10.1371/journal.pone.0079523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandewauw I, Owsianik G, Voets T. Systematic and quantitative mRNA expression analysis of TRP channel genes at the single trigeminal and dorsal root ganglion level in mouse. BMC Neurosci. 2013;14(1):21. doi: 10.1186/1471-2202-14-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paintal AS. Vagal sensory receptors and their reflex effects. Physiol Rev. 1973;53(1):159–227. doi: 10.1152/physrev.1973.53.1.159. [DOI] [PubMed] [Google Scholar]
- Springall DR, Cadieux A, Oliveira H, Su H, Royston D, Polak JM. Retrograde tracing shows that CGRP-immunoreactive nerves of rat trachea and lung originate from vagal and dorsal root ganglia. J Auton Nerv Syst. 1987;20(2):155–166. doi: 10.1016/0165-1838(87)90113-5. [DOI] [PubMed] [Google Scholar]
- Ricco MM, Kummer W, Biglari B, Myers AC, Undem BJ. Interganglionic segregation of distinct vagal afferent fibre phenotypes in guinea-pig airways. J Physiol. 1996;496(Pt 2):521–530. doi: 10.1113/jphysiol.1996.sp021703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao H, Sprunger LK, Simasko SM. Expression of transient receptor potential channels and two-pore potassium channels in subtypes of vagal afferent neurons in rat. Am J Physiol Gastrointest Liver Physiol. 2010;298(2):212–221. doi: 10.1152/ajpgi.00396.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhuo H, Ichikawa H, Helke CJ. Neurochemistry of the nodose ganglion. Prog Neurobiol. 1997;52(2):79–107. doi: 10.1016/s0301-0082(97)00003-8. [DOI] [PubMed] [Google Scholar]
- Chang RB, Strochlic DE, Williams EK, Umans BD, Liberles SD. Vagal Sensory Neuron Subtypes that Differentially Control Breathing. Cell. 2015;161:1–12. doi: 10.1016/j.cell.2015.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczyńska K, Szereda-Przestaszewska M. Nodose ganglia-modulatory effects on respiration. Physiol Res. 2013;62:227–235. doi: 10.33549/physiolres.932412. [DOI] [PubMed] [Google Scholar]
- Taylor-Clark TE, Undem BJ. Sensing pulmonary oxidative stress by lung vagal afferents. Respir Physiol Neurobiol. 2011;178(3):406–413. doi: 10.1016/j.resp.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bautista DM, et al. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell. 2006;124(6):1269–1282. doi: 10.1016/j.cell.2006.02.023. [DOI] [PubMed] [Google Scholar]
- Ichikawa H, De Repentigny Y, Kothary R, Sugimoto T. The survival of vagal and glossopharyngeal sensory neurons is dependent upon dystonin. Neuroscience. 2006;137(2):531–536. doi: 10.1016/j.neuroscience.2005.08.081. [DOI] [PubMed] [Google Scholar]
- Hondoh A, et al. Distinct expression of cold receptors (TRPM8 and TRPA1) in the rat nodose-petrosal ganglion complex. Brain Res. 2010;1319:60–69. doi: 10.1016/j.brainres.2010.01.016. [DOI] [PubMed] [Google Scholar]
- Kummer W, Fischer A, Kurkowski R, Heym C. The sensory and sympathetic innervation of guinea-pig lung and trachea as studied by retrograde neuronal tracing and double-labelling immunohistochemistry. Neuroscience. 1992;49(3):715–737. doi: 10.1016/0306-4522(92)90239-x. [DOI] [PubMed] [Google Scholar]
- Choi D, Li D, Raisman G. Fluorescent retrograde neuronal tracers that label the rat facial nucleus: A comparison of Fast Blue, Fluoro-ruby, Fluoro-emerald, Fluoro-Gold and DiI. J Neurosci Methods. 2002;117(2):167–172. doi: 10.1016/s0165-0270(02)00098-5. [DOI] [PubMed] [Google Scholar]
- Calik MW, Radulovacki M, Carley DW. A Method of Nodose Ganglia Injection in Sprague-Dawley Rat. J Vis Exp. 2014. pp. e1–e5. [DOI] [PMC free article] [PubMed]
- Ramachandra R, McGrew S, Elmslie K. Identification of specific sensory neuron populations for study of expressed ion channels. J Vis Exp. 2013. p. e50782. [DOI] [PMC free article] [PubMed]
- Yu X, Hu Y, Ru F, Kollarik M, Undem BJ, Yu S. TRPM8 function and expression in vagal sensory neurons and afferent nerves innervating guinea pig esophagus. Am J Physiol - Gastrointest Liver Physiol. 2015;308(6):489–496. doi: 10.1152/ajpgi.00336.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwong K, Lee L-Y. PGE(2) sensitizes cultured pulmonary vagal sensory neurons to chemical and electrical stimuli. J Appl Physiol. 2002;93(4):1419–1428. doi: 10.1152/japplphysiol.00382.2002. [DOI] [PubMed] [Google Scholar]
- Joachim RA, et al. Stress induces substance P in vagal sensory neurons innervating the mouse airways. Clin Exp Allergy. 2006;36(8):1001–1010. doi: 10.1111/j.1365-2222.2006.02533.x. [DOI] [PubMed] [Google Scholar]
- Kaan TKY, et al. Systemic blockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour in rats. Brain. 2010;133(9):2549–2564. doi: 10.1093/brain/awq194. [DOI] [PubMed] [Google Scholar]
- Nakatani T, Minaki Y, Kumai M, Ono Y. Helt determines GABAergic over glutamatergic neuronal fate by repressing Ngn genes in the developing mesencephalon. Development. 2007;134(15):2783–2793. doi: 10.1242/dev.02870. [DOI] [PubMed] [Google Scholar]
- Lobo MK, Karsten SL, Gray M, Geschwind DH, Yang XW. FACS-array profiling of striatal projection neuron subtypes in juvenile and adult mouse brains. Nat Neurosci. 2006;9(3):443–452. doi: 10.1038/nn1654. [DOI] [PubMed] [Google Scholar]
- Usoskin D, et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat Neurosci. 2015;18:145–153. doi: 10.1038/nn.3881. [DOI] [PubMed] [Google Scholar]