

Abstract
The Arabidopsis genome encodes numerous iron-containing proteins such as iron-sulfur (Fe-S) cluster proteins and hemoproteins. These proteins generally utilize iron as a cofactor, and they perform critical roles in photosynthesis, genome stability, electron transfer, and oxidation-reduction reactions. Plants have evolved sophisticated mechanisms to maintain iron homeostasis for the assembly of functional iron-containing proteins, thereby ensuring genome stability, cell development, and plant growth. Over the past few years, our understanding of iron-containing proteins and their functions involved in genome stability has expanded enormously. In this review, I provide the current perspectives on iron homeostasis in Arabidopsis, followed by a summary of iron-containing protein functions involved in genome stability maintenance and a discussion of their possible molecular mechanisms.
Keywords: Genome integrity, iron-containing protein, iron homeostasis
Introduction: Iron-Containing Proteins in Arabidopsis
Iron is an essential micronutrient for all organisms because it functions as a cofactor in a wide range of biological processes including DNA replication and repair.[1,2,3] Eukaryotic cells contain numerous iron-containing proteins, which can be mainly classified into three groups: Iron-sulfur (Fe-S) cluster proteins, hemoproteins, and non-heme/non-Fe-S proteins.[2,4] Fe-S proteins are characterized by their different structures with variable oxidation states, ranging from [2Fe-2S] diamonds, [3Fe-4S] intermediates, to [4Fe-4S] cluster cubes.[5,6] Examples of Fe-S proteins include DNA polymerases, DNA helicases, hydrogenases, nicotinamide adenine dinucleotide (NADH)-dehydrogenases, nitrogenases, ferredoxins, and aconitases.[1,7] Hemoproteins have a heme prosthetic group that allows them to carry out oxidative functions.[8] Examples of hemoproteins include cytochromes, hemoglobin, myoglobin, catalases, and peroxidases.[9] Non-heme/non-Fe-S proteins can be further subgrouped into three classes: Mononuclear non-heme iron enzymes, diiron proteins, and proteins involved in ferric iron transport.[10,11] This group of iron-containing proteins mainly includes the small subunit of ribonucleotide reductases (RNRs), superoxide dismutases (SODs), dioxygenases, pterin-dependent hydrolases, and lipoxygenases.[12,13]
The apo-forms of iron-containing proteins utilize iron as a cofactor to assemble their functional holoproteins.[1,14] The mammalian and yeast iron-containing proteins exhibit extensive functions in electron transfer, telomere maintenance, genome stability, and cell cycle control.[2,3] Similarly, the Arabidopsis genome also encodes numerous iron-containing proteins, including 107 putative and confirmed Fe-S proteins; 246 cytochrome P450 (CYP) members; 7 cytochrome b5 (Cb5) proteins; over 100 proteins involved in heme synthesis and heme binding; 12 putative and confirmed iron-containing dioxygenases; 4 iron-dependent SODs; and 3 RNR small subunits (from The Arabidopsis Information Resource, TAIR, http://www.arabidopsis.org/). Some of them have been directly characterized or implicated to play important roles in genome stability, such as DNA polymerases, DNA primase large subunits, DNA helicases, DNA glycosylases, peroxidase superfamily proteins, Cb5 proteins, cytochrome c (CYTc), SODs, and the small subunits of RNR [Table 1]. Additionally, some iron-containing proteins are also involved in the maintenance of genome stability through indirect ways. For instance, the biogenesis and maturation of cytosolic Fe-S proteins require the function of a dedicated Fe-S cluster assembly pathway, namely, cytosolic iron-sulfur cluster assembly (CIA) machinery.[1,14] The CIA pathway is necessary for transferring Fe-S clusters to target proteins such as DNA polymerases, DNA primases, DNA helicases, and RNRs in mammals and yeast.[1,15,16,17] Thus, the disruption of proteins involved in the CIA pathway possibly affects the stability of target proteins, resulting in genome instability [Table 1].
Table 1.
The putative and confirmed iron-containing proteins involved in genome stability in Arabidopsis
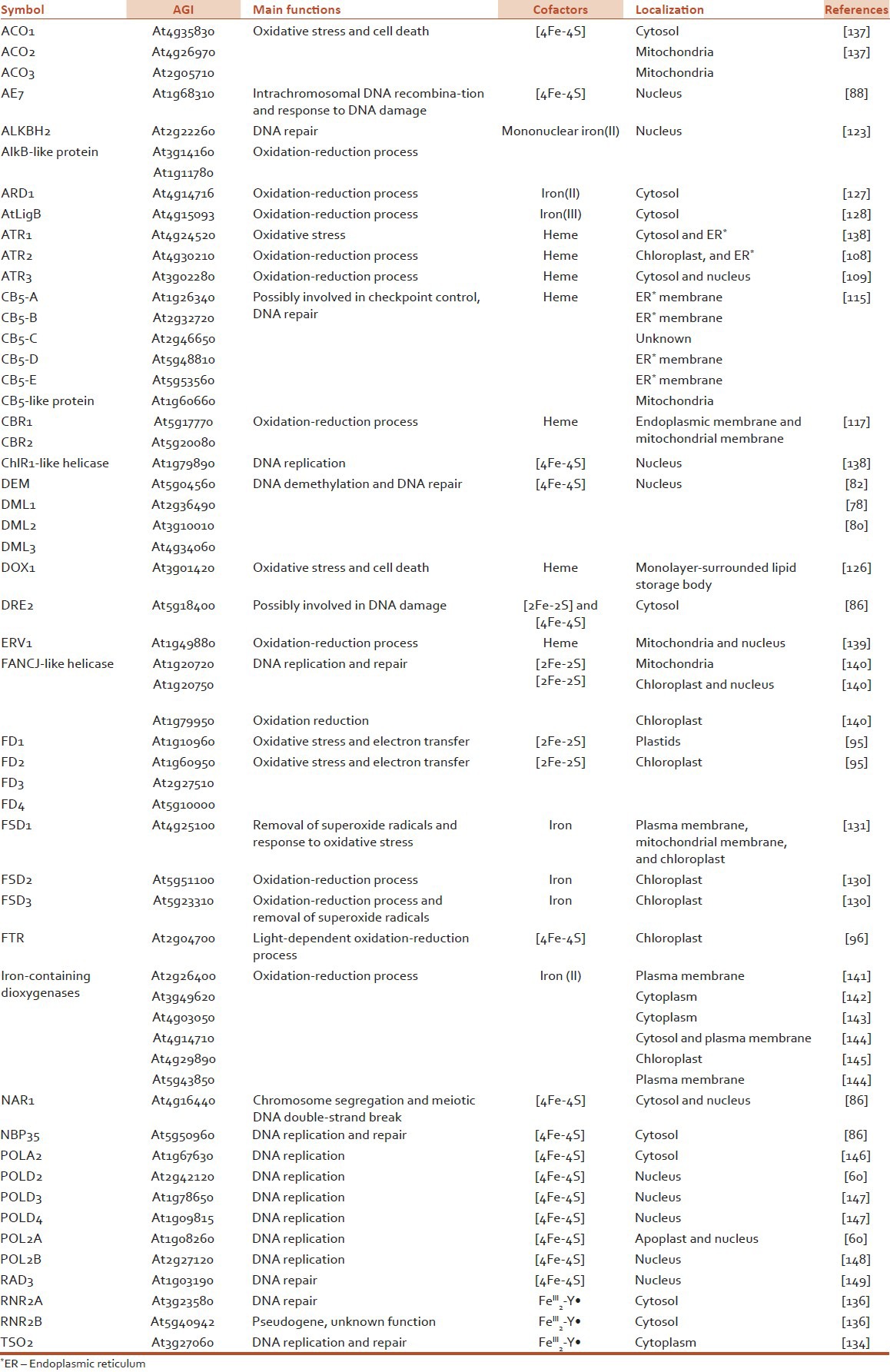
Iron Homeostasis in Arabidopsis
In Arabidopsis, iron homeostasis is achieved via strictly controlled systems for iron uptake at the cell plasma membrane in roots and for eliciting balanced iron distribution among cellular compartments, as well as systemic translocation.[18,19]
Iron uptake
Plants have evolved two distinct iron uptake strategies: Strategy I and strategy II,[20] which are responsible for iron acquisition in dicotyledenous plants/non-graminaceous monocots and in grasses, respectively.[21,22] Arabidopsis utilizes strategy I to absorb iron, which is tightly controlled by the root high-affinity iron uptake system.[23] This process is highly dependent on the expression of FRO2 (ferric reduction oxidase 2, At1g01580) and IRT1 (iron-regulated transporter 1, At4g19690).[23,24,25] FRO2 functions as a root ferric-chelate reductase, and IRT1 is required for the transport of ferrous iron across the plasma membrane [Figure 1].[24,26] Both of them specifically function in root iron uptake under iron-deficient conditions.[24,26] The expression of FRO2 and IRT1 is rapidly induced in iron-deficient conditions, whereas it is dramatically diminished in iron-sufficient conditions via posttranslational mechanisms.[23,27] The Arabidopsis genome also encodes an IRT1 paralog, namely, IRT2 (At4g19680). Studies have indicated that IRT2 cooperates with IRT1 and FRO2 to maintain iron homeostasis in root epidermal cells.[25]
Figure 1.
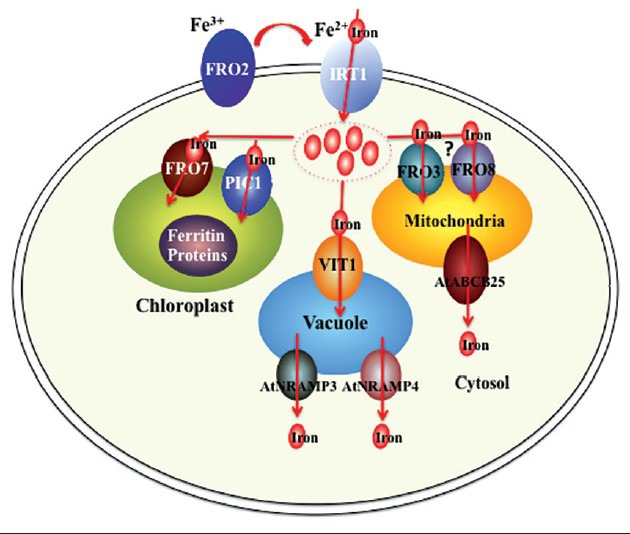
Iron uptake and intracellular trafficking in an Arabidopsis cell. The ferric iron (Fe3+) is primarily reduced to ferrous iron (Fe2+) by surface reductase FRO2. Iron uptake is carried out at the plasma membrane by iron transporter IRT1. When iron enters the cytosol, it can be delivered into the chloroplast, vacuole, and mitochondria by iron transporters. In the chloroplast, FRO7 is the main iron transporter. Moreover, PIC1 can mediate iron transport across the inner envelope of chloroplasts. The import iron is mainly stored in ferritin proteins (AtFER1-AtFER4). In the vacuole, iron accumulation and storage are controlled by the VIT1, AtNRAMP3, and AtNRAMP4 proteins. In the mitochondria, FRO3 and FRO8 are proposed to be required for iron transport. The ABC transporter protein AtABCB25 functions in iron efflux to the cytosol
Interestingly, the expression of many iron-regulated genes is induced by FIT (Fer-like Deficiency Induced Transcription Factor, At2g28160), which is a transcription factor that regulates iron uptake responses.[28] The fit mutant accumulates less iron in root and shoot tissues in comparison with wild-type plants.[28] Further studies indicate that FIT can regulate ferric chelate reductase activity and iron transport into plant roots.[28] This process is achieved by regulating FRO2 expression and by controlling protein accumulation of the IRT1.[28] Moreover, the Ib subgroup of the basic helix-loop-helix (bHLH) gene family (AtbHLH38, AtbHLH39, AtbHLH100, and AtbHLH101) in Arabidopsis also has been reported to participate in the regulation of iron uptake. AtbHLH38 and AtbHLH39 can interact with FIT, directly activating the expression of FRO2 and IRT1.[29] Recently, AtbHLH100 and AtbHLH101 have also been identified to interact with FIT.[30] Overexpression of FIT and AtbHLH101 in plants results in the constitutive expression of FRO2 and IRT1 in the roots, and accumulates more iron in the shoots.[30] However, the expression of FRO2 and IRT1 in roots and the iron content in shoots dramatically decrease in the triple knockout mutant of AtbHLH39, AtbHLH100, and AtbHLH101.[30] Moreover, the mediator subunit 16 (MED16, At4g04920) is reported to function in the regulation of iron uptake gene expression in Arabidopsis.[31] Lesion of MED16 significantly reduces the expression of FRO2 and IRT1 in Arabidopsis roots.[31] MED16 can interact with FIT and improves the binding of the FIT/Ib bHLH complex to FRO2 and IRT1 promoters under iron-deficient conditions.[31] Shk1 binding protein 1 (SKB1/AtPRMT5, At4g31120) is also reported to be involved in iron homeostasis in Arabidopsis.[32] The chromatin immunoprecipitation (ChIP) and genome-wide ChIP-seq results show that SKB1 associates with the chromatin of the Ib subgroup bHLH genes.[32] In addition, SKB1 can catalyze the symmetric dimethylation of histone H4R3 (H4R3sme2), and the level of H4R3sme2 positively corresponds to the iron status of plants.[32] These results indicate that SKB1-mediated H4R3sme2 regulates iron homeostasis in Arabidopsis.[32] Iron deficiency may increase the disassociation of SKB1 from bHLH genes in chromatin and decrease the level of H4R3sme2, thereby elevating the expression of bHLH genes and enhancing iron uptake.[32]
In addition, some studies suggest that there exist additional root-derived signals to control iron uptake.[23,33,34] Split-root experiments indicate that Fe(III) reductase activity is higher in the roots supplied with iron,[23,34] implying that the systemic signal generated by iron-deficient shoots is further modulated by a local, root-derived signal.[33]
Intracellular iron transport
Iron is required to be compartmentalized into different cellular organelles, such as chloroplasts, vacuoles, and mitochondria.[35] However, to date, much less is known about intracellular iron transport in Arabidopsis. Ferrous iron in roots is transported, it is suggested, into xylem via Ferroportin-1 (FPN1, At2g38460), where it forms a complex with citrate.[36] The complex is loaded into the phloem and further forms a complex with nicotianamine (NA), which is synthesized from S-adenosyl methionine by nicotianamine synthase (NAS).[36] Recent studies indicate that transcription factors MYB10 (At3g12820) and MYB72 (At1g56160) function in the iron-deficiency regulatory cascade to drive NAS4 gene expression.[36]
In addition, some proteins are probably involved in intracellular iron homeostasis according to the predicted organelle localization. For instance, FRO7 (At5g49740), a paralog of FRO2, might be involved in transporting iron into the chloroplast [Figure 1].[37] The expression of three members of the natural resistance-associated macrophage protein (NRAMP) metal transporter family, including AtNRAMP1 (At1g80830), AtNRAMP3 (At2g23150), and AtNRAMP4 (At5g67330), is regulated by the iron status.[38,39] Functional analyses have demonstrated that all three proteins are capable of transporting Fe and Mn.[40,41] AtNRAMP3 and AtNRAMP4 localize in the vacuole, where they can export metal ions into the cytosol.[38,42] Interestingly, AtNRAMP3 and AtNRAMP4 function, it is suggested, in the long-distance transport of metals due to their expressions in the stele of roots and in the vasculature of leaves and stems.[38,42] The vacuolar iron transporter 1 (VIT1, At2g01770) mediates iron accumulation in vacuoles and controls the localization of Fe in seeds.[43,44] A plastid protein PIC1 (permease in chloroplast 1, At2g15290) can regulate iron transport across the inner envelope of chloroplasts.[35]
Iron storage
In Arabidopsis, iron is mainly stored in the chloroplast, vacuole, and mitochondria, where it can be utilized by numerous iron-containing proteins.[18] The majority of iron is stored in chloroplast ferritin proteins.[44] Arabidopsis contains four ferritin members, namely, AtFer1-AtFer4 (At5g01600, At3g11050, At3g56090, and At2g40300, respectively) [Figure 1].[45] All these four ferritin proteins are predicted to target to the chloroplast, as they contain transit peptides required for delivering iron to the plastid.[46] Importantly, the expression of AtFer1, AtFer3, and AtFer4 is induced when plants are treated with excess iron.[45,46,47] In addition, iron can also be stored in the vacuole, where it is mainly controlled by AtNRAMP3, AtNRAMP4, and VIT1 iron transporters [Figure 1].[38,42,43] Moreover, AtABCB25 (also named AtSTA1, At5g58270), an ATP-binding cassette (ABC) transporter, has been shown to be important in iron efflux from the mitochondria to the cytosol [Figure 1].[48]
Iron translocation
Most iron acquired by the roots is ultimately delivered to above-ground portions of the plant via the xylem.[49] During this process, iron needs to cross several different membrane barriers.[23] Plants utilize a sophisticated system to deliver iron from root epidermal cells to leaf cells.[50] A variety of transporters have been identified to be involved in the iron translocation process in Arabidopsis, including (ferric reductase defective 3, At3g08040) FRD3 and the yellow stripe-like (YSL) family of proteins (YSL1-8). FRD3 facilitates citrate efflux into the xylem, and the mutation of FRD3 results in Fe localizing to the central vascular cylinder of the roots and failing to transport it to the aerial parts.[51] The YSL family of proteins transport, it is suggested, metals complexed with NA.[52] In Arabidopsis, YSL1 (At4g24120) and YSL3 (At5g53550) are important for iron transport and also responsible for loading Fe, Cu, and Zn from leaves into seeds.[53] YSL4 (At5g41000) and YSL6 (At3g27020) are suggested to control iron release from the chloroplast,[54] and they are also involved in iron transport and metal mobilization into seeds.[54] The transport functions of YSL1 and YSL2 (At5g24380) are shown to partially overlap with the function of YSL3 in vegetative structures, but they are distinct in the reproductive organs.[52] The functions of YSL7 and YSL8 in iron translocation have not been characterized.
The effects of iron homeostasis on protein functions
Maintaining iron homeostasis is critical for assembling prosthetic groups such as heme and Fe-S clusters.[55] Much less is known about the effects of iron homeostasis on protein stability in Arabidopsis, whereas studies in mammals have elucidated that iron homeostasis is governed in part through the regulated proteolysis of ferroportin (iron exporter), hypoxia-inducible factor (HIF), iron-regulatory proteins (IRPs), and an F-box/leucine-rich repeat protein (FBXL5).[56] Obviously, the critical roles played by iron in both enzymatic catalysis and protein structure contribute to the stability of iron regulation proteins and iron-containing proteins.[56]
Fe-S Cluster Proteins and Genome Stability
Numerous Fe-S cluster proteins are reported to function in genome stability maintenance in Arabidopsis. These proteins mainly include DNA polymerases, DNA helicases, DNA glycosylases, the large subunit of DNA primase, and components of the CIA system.
DNA polymerases and DNA primases
Eukaryotes mainly utilize three conserved polymerases (Polα, Pold, and Polε) to build DNA blocks.[57] Polα is associated with DNA primases to synthesize short RNA primers, which are subsequently utilized by Polε and Pold to synthesize the leading and lagging strands, respectively.[2,58,59] All the three DNA polymerases and the large subunit of primase contain a Fe-S cluster.[2] Arabidopsis has a DNA polymerase α (POLA2, At1g67630), three members of polymerase d (POLD2-At2g42120, POLD3-At1g78650, and POLD4-At1g09815), two catalytic subunits of Polε (POL2A-At1g08260 and POL2B-At2g27120), and a large subunit of DNA primase (At1g67320). Although the functions of Arabidopsis DNA polymerases are less characterized, their amino acid sequences share a high degree of similarity to their corresponding homologues in yeast and mammals, implicating their conserved functions in DNA replication.[60] In rice, the expression of DNA polymerase d catalytic subunit (POLD1, Os11g0186400) can be detected in mature leaves and is induced under ultraviolet (UV) irradiation treatment,[61] supporting the conclusion that POLD is required for DNA replication in plants.[62] In Arabidopsis, both POL2A and POL2B contain the conserved domains that are present in other eukaryotic homologues.[63] Mutations of POL2A result in DNA replication defects,[64] while pol2b mutants have no visible phenotypic effects.[63] These results suggest that POL2B is not essential for DNA replication or that POL2B functions redundantly with other DNA polymerases.
DNA helicases
DNA helicases are highly conserved enzymes that function to unwind DNA in order to provide a single-stranded DNA for replication, RNA transcription, DNA repair, and recombination.[65,66] Defects of helicases are generally associated with genomic instability in yeast and mammals.[66] The Arabidopsis genome encodes numerous proteins exhibiting helicase activities, and these proteins include AtFANCM (At1g35530), AtINO80 (At5g57300), AtMER3 (or AtRCK, At3g27730), AtRAD54 (At3g19210), AtRAD5A (At5g22750), AtRECQ2 (At1g31360), AtRECQ4A (At1g10930), AtSRS2 (At4g25120), AtSWR1 (At5g37055), RAD3 (UVH6) (At1g03190), three homologues of human Fanconi anemia group J protein (FANCJ) helicase (At1g20720, At1g20750, and At1g79950), and a homologue of human ChlR1 helicase (At1g79890) (according to TAIR). Among the proteins listed above, only RAD3, the homologues of FANCJ helicase, and the homologue of ChlR1 helicase contain a Fe-S cluster. The Arabidopsis RAD3 (also known as UVH6) is a homologue of the human XPD and yeast RAD3. Both XPD and yeast RAD3 are essential helicases, with roles in the repair of damaged DNA through the nucleotide excision repair (NER) mechanism.[67,68] The Fe-S cluster in yeast RAD3 is essential for the coupling of adenosine triphosphate (ATP) hydrolysis to DNA translocation and for targeting the helicase to the DNA junction.[69] The uvh6-1 mutant is hypersensitive to both UV-C and UV-B irradiation, implicating its important role in DNA repair.[70] The human FANCJ helicase has been identified to catalyze the unwinding of duplex DNA and G-quadruplex structures in an ATP hydrolysis-dependent manner to ensure genomic stability.[71] The Arabidopsis genome encodes three FANCJ-like proteins, but their functions are still not characterized. ChlR1 belongs to the FANCJ-like DNA helicase family and it contains a DEAH/DEAD box,[71] which is required for unwinding nucleic acids and is involved in various aspects of RNA metabolism.[71] The deletion of ChlR1 in mammalian cells can lead to DNA damage accumulation, suggesting its important role in efficient DNA repair during DNA replication.[72] However, the function of Arabidopsis ChlR1-like helicase remains unclear.
DNA glycosylases
DNA glycosylases can recognize and excise mismatched or altered bases through the base excision repair (BER) mechanism.[73] Generally, all glycosylases function in a similar way; they cleave the N-glycosylic bond between the target base and the sugar-phosphate backbone of the DNA,[74] thereby releasing a free base and leaving an apurinic/apyrimidinic (AP) site.[75] Arabidopsis has 26 DNA glycosylases that extensively function in the DNA repair process;[76,77] of them, only the DEM (DNA glycosylase DEMETER, At5g04560), DML1 (DEMETER-LIKE 1, also known as AtROS1, At2g36490), DML2 (At3g10010), and DML3 (At4g34060) proteins contain a Fe-S cluster. These iron-containing DNA glycosylases extensively function in DNA methylation.[78,79,80] The DME gene is mainly expressed in the central cells before fertilization and is required for the DNA demethylation of the maternal allele in the endosperm that establishes gene imprinting.[81,82] DME also functions, it has been suggested, in a protein complex, providing promoter specificity for base excision and DNA nicking of the maternal genome.[83] Interestingly, AtROS1 possesses both DNA glycosylase and lyase activities against methylated DNA, but not unmethylated DNA.[78] The atros1 knockout mutant is hypersensitive to genotoxic stresses such as methyl methanesulfonate (MMS) and hydrogen peroxide (H2O2), suggesting that ROS1 is involved in the DNA repair process by repressing homology-dependent transcriptional gene silencing via the demethylation of target promoter DNA.[78] Moreover, the Arabidopsis genome encodes two additional paralogs of AtROS1, namely, DML2 and DML3.[80] Hypermethylation of cytosine residues has been observed in dml2 and dml3 mutants relative to wild-type plants.[80] These results indicate the important roles of DML2 and DML3 in removing DNA methylation from improperly-methylated cytosines.[80]
Components of CIA machinery
The assembly of Fe-S cluster is carried out via ISC (iron-sulfur cluster, in mitochondria), CIA, and SUF (sulfur mobilization, in plastid) machineries in plants.[84] Eukaryotes share conserved mechanisms for the synthesis of Fe-S clusters and their insertions into apoproteins.[85] The biogenesis of Fe-S proteins contains two major steps:
A Fe-S cluster is assembled on a scaffold complex, and
The Fe-S cluster is dislocated from the scaffold and transferred to specific apoproteins.[1,85]
Some of the CIA components have been found to function in DNA replication and repair processes in different organisms. However, little is known about the roles of ISC and SUF members in genome stability.
The CIA machinery members in Arabidopsis include NAR1 (At4g16440), CIA1 (At2g26060), NBP35 (At5g50960), AE7 (At1g68310), MET18 (also named MMS19, At5g48120), DRE2 (At5g18400), TAH18 (also named ATR3, At3g02280), ATM3 (At5g58270), and ERV1 (At1g49880).[86] The yeast CIA pathway proteins NAR1, CIA1, CIA2, and MET18, and their corresponding human homologues IOP1, CIAO1, MIP18, and MMS19, it is proposed, transfer Fe-S clusters to the target proteins.[1,16,17,87] More importantly, both the human and yeast MMS19 proteins interact with numerous Fe-S proteins, including Pold, DNA primase, Dna2, XPD, RTEL1, and FANCJ.[16] Similarly, the Arabidopsis AE7-CIA1-NAR1-MET18 complex has been indicated to facilitate the transfer of Fe-S clusters to the target apoproteins such as ACO (aconitase) and ROS1. In addition, the Arabidopsis CIA pathway has also been suggested in the maintenance of nuclear genome integrity through Fe-S proteins involved in DNA metabolism.[88] Mutations of CIA members including AE7 and ATM3 lead to the accumulation of DNA damage and the increase of homologous recombination (HR) rates.[88] Therefore, it is highly possible that genomic integrity defects in ae7 and atm3 mutants result from inefficient assembly of the Fe-S cluster proteins involved in DNA replication and repair.[88] Taken together, the CIA pathway plays a critical role in maintaining genome integrity, due to the importance of the Fe-S proteins in DNA replication and repair.
Other Fe-S proteins
In addition, some Fe-S proteins are also involved in genome stability. For instance, the aconitase family proteins, including ACO1 (At4g35830), ACO2 (At4g26970), and ACO3 (At2g05710), play roles in catalyzing the conversion of citrate to isocitrate.[89] The yeast ACO1 can bind to mitochondrial DNA (mtDNA) and mediate its maintenance; as a result, the disruption of ACO1 causes mtDNA instability.[89] In Arabidopsis, ACO1 can specifically bind to the 5′ untranslated region (5′-UTR) of CSD2 (At2g28190, a SOD) in vitro,[90] whereas ACO2 affects CSD2 gene expression and may function in response to oxidative stresses.[91] ACO3 is required for controlling seed abscisic acid (ABA) sensitivity and seedling establishment.[92] Furthermore, ferredoxin (Fd) family proteins, including FD1 (At1g10960), FD2 (At1g60950), FD3 (At2g27510), and FD4 (At5g10000) also contain a [2Fe-2S] cluster. The FD family proteins have been demonstrated to extensively involve oxidative stresses,[93,94,95] suggesting their critical roles in maintaining genome stability. The Arabidopsis ferredoxin/thioredoxin reductase (FTR, At2g04700) contains a [4Fe-4S] cluster and plays a key role in the light-dependent redox regulatory system.[96] The ftr knockout mutant exhibits significant sensitivity to oxidative stresses.[96]
Hemoproteins and DNA Stability
Heme is a diverse cofactor and it participates in a wide range of chemical reactions, such as electron transfer, oxygen activation, and gene regulation.[97] Heme-containing proteins, also termed hemoproteins, are essential for the physiology and viability of living organisms, and contribute to diverse functions including respiration, oxygen carriage, cellular signaling, and apoptosis.[98] Mutations of hemoproteins are always associated with the induction of reactive oxygen species (ROS),[83,99] which can damage lipids, proteins, and DNA. The Arabidopsis hemoproteins involved in genome stability mainly include CYP reductase (CPR), Cb5, and CYTc.
CPRs
The CYP superfamily proteins utilize heme as a cofactor and function in the oxidation/reduction of endogenous or exogenous compounds.[100] CYPs require a CPR to transfer electrons from reduced nicotinamide adenine dinucleotide phosphate (NADPH) to their substrates.[101] The NADPH-dependent CPR generally localizes in the endoplasmic reticulum (ER) membrane and serves as the electron donor of CYPs.[102,103] CPR contains multidomains such as three cofactor-binding domains (flavin mononucleotide [FMN], flavin adenine dinucleotide [FAD], and NADPH) and a linker domain situated between the FMN and FAD/NADPH domains.[104]
CPR is the most imperative redox partner of CYPs.[101] Human CPR and NADPH-dependent CPR can act as sources of endogenous oxidative DNA damage and are required for genome stability.[105] The Arabidopsis genome encodes 246 P450 genes, which can be grouped into 72 families.[106] Their biological functions range from the synthesis of macromolecules, hormones, and signaling molecules, and to the metabolism of xenobiotics.[106] However, the functions of numerous P450 genes remain unclear. The Arabidopsis genome encodes two authentic and one putative CPR genes, namely, ATR1 (At4g24520), ATR2 (At4g30210), and ATR3 (At3g02280), respectively. ATR1 is required for electron transfer from NADP to CYPs in microsomes.[107] In particular, it can provide electrons to heme oxygenase and Cb5.[107] ATR2 contributes to the first oxidative step of the phenylpropanoid general pathway.[108] ATR3 serves as a diflavin reductase and is essential for Arabidopsis embryo development.[109] Interestingly, ATR3 exhibits CYTc reductase activity, but not P450 reductase activity.[109] The yeast two-hybrid screening has identified that ATR3 can interact with two Fe-S proteins, the human CIAPIN1 and the yeast Dre2 protein.[109] These results suggest that ATR3 may function in genome stability either by ways similar to CYTc or by interacting with Fe-S proteins.
Cb5
Cb5s are ubiquitous hemoproteins and typically associate with the ER and outer mitochondrial membranes.[110] In higher eukaryotes, Cb5 functions as an electron donor for the desaturation of acyl-CoA fatty acids (FAs), sphingolipid long-chain base hydroxylation and desaturation, FA hydroxylation, sterol desaturation, and cytochrome P450-mediated reactions.[110] In plants, Cb5 plays a primary role in providing electrons for the synthesis of the polyunsaturated FAs linoleic acid (18:2) and a-linolenic acid (18:3),[111] which contributes to the integrity of cellular membranes.[112] Interestingly, previous studies have demonstrated that some Cb5-like proteins play critical roles in genome stability maintenance. For instance, a Cb5-like protein Dap1 is required for resistance to DNA damage agent methyl MMS in yeast.[113] Moreover, Irc21 protein also contains a Cb5-like domain and has been revealed to function in checkpoint control, DNA repair, and genome stability.[114]
The Arabidopsis genome encodes five Cb5 members (CB5A-E, referring to At1g26340, At2g32720, At2g46650, At5g48810, and At5g53560, respectively) and one Cb5-like protein (At1g60660).[115] Four of them including, CB5A, -B, -D, and -E are predicted to localize in the ER membrane.[115,116] However, little is known about the specific functions of individual Cb5 proteins. These proteins share a high degree of amino acid sequence similarities, suggesting the high possibility that they function redundantly. Two Cb5 reductases, namely, CBR1 (At5g17770) and CBR2 (At5g20080) are present in the endoplasmic membrane and the inner mitochondrial membrane, respectively.[117] CBR1 is essential for a functional male gametophyte.[117] Although both CBR1 and CBR2 are also predicated to function as FAD/NAD-binding oxidoreductases, further studies are still needed to uncover their biological roles.
CYTc
CYTc is a small hydrophilic hemoprotein that is extensively present in the mitochondrial inner membrane.[118] CYTc participates in many biological processes, such as respiration, apoptosis, cell death, oxidative stress, DNA damage, energetic metabolism, protein folding, and translational regulation.[119,120] In Arabidopsis, CYTc is encoded by CYTC-1 (At1g22840) and CYTC-2 (At4g10040). The knocking-out of both genes causes lethality in plants, whereas the individual mutants have no visible phenotype.[118] These results suggest that CYTC-1 and CYTC-2 function redundantly. Moreover, plants with decreased CYTc exhibit developmental delay, the alteration of stress-responsive gene expression, and the reduction of ROS levels,[118] implying that CYTc in Arabidopsis also functions as conservatively as other eukaryotes.
Other Iron-Containing Proteins and DNA Stability
Besides Fe-S proteins and hemoproteins, Arabidopsis also contains many other iron-containing proteins involved in genome stability, such as dioxygenases, SODs, and RNRs.
Dioxygenases
Some dioxygenases can utilize iron to incorporate into active sites for the assembly of holoproteins.[121,122] These iron-containing dioxygenases have been identified to be involved in the DNA repair process. For instance, the Fe(II)/2-oxoglutarate-dependent dioxygenase alpha-ketoglutarate-dependent dioxygenase (AlkB) is extensively present in Escherichia coli and in mammals.[123] The AlkB protein performs a conserving function in the oxidative removal of damaged DNA via alkylation.[124] Failure to remove damaged DNA generally leads to cytotoxicity or mutagenesis during DNA replication.[124] The Arabidopsis genome encodes several AlkB homologues, including ALKBH2 (At2g22260), At3g14160, and At1g11780. In them, the ALKBH2 protein can protect Arabidopsis against DNA methylation damage.[124] The alkbh2 knockout mutants are hypersensitive to the MMS.[123] However, functions of the other two genes have not been characterized. Additionally, another nine genes are also annotated as iron-containing dioxygenases, and these are At2g26400, At3g01420, At3g49620, At4g03050, At4g14710, At4g14716, At4g15093, At4g29890, and At5g43850. Of them, At3g01420, also termed DOX1, encodes a α-dioxygenase that protects plants against oxidative stress and cell death.[125] Recently, it has been shown to be an important component that positively regulates programmed cell death (PCD).[126] The At4g14716 gene encodes an acireductone dioxygenase 1 (ARD1) and functions as an effector of the β subunit of heterotrimeric G protein, which may participate in the synthesis of ethylene.[127] The At4g15093 gene, or AtLigB, is annotated as an extradiol ring-cleavage enzyme, and contributes to arabidopyrone (AP) biosynthesis.[128] The functions of the other six putative iron-containing dioxygenases have not been characterized.
SODs
Numerous environmental stresses can result in the abnormal induction of superoxide within plant tissues.[129] Plants commonly utilize SOD to detoxify the excess ROS.[129] The Arabidopsis genome encodes one manganese SOD (MSD1, At3g10920), three copper/zinc SODs (CSD1-At1g08830, CSD2-At2g28190, and CSD3-At5g18100), and three iron SODs (FSD1-At4g25100, FSD2-At5g51100, and FSD3-At5g23310). The iron-dependent FSD1 is abundantly present in the plasma membrane, mitochondrial membrane, and different fractions of chloroplast such as stroma, envelope, and the peripheral thylakoid.[130] The stromal localization implies its important role in photosynthesis due to the ability to scavenge ROS in the water-water cycle.[131] Notably, the expression of FSD1 is dramatically induced in low Cu levels.[132] FSD2 and FSD3 play essential roles in early chloroplast development.[130] The fsd2-1fsd3-1 double mutant plants exhibit a severe albino phenotype and are hypersensitive to oxidative stress.[130] In vivo and in vitro studies have confirmed that FSD2 and FSD3 proteins can form a heteromeric protein complex in the chloroplast, suggesting that the FSD2-FSD3 complex functions in the scavenging of ROS to facilitate the maintenance of early chloroplast development by protecting it against ROS.[130]
RNRs
RNRs are critical enzymes that catalyze a rate-limiting step in the synthesis of deoxyribonucleotides (dNTPs), thereby generating the precursors needed for DNA replication and repair.[133,134] Eukaryotic RNRs comprise the large subunits (α or R1) and small subunits (β or R2), of which only R2 subunits utilize iron to sustain a diferric tyrosyl radical (FeIII2-Y·) cofactor.[2] Previous studies in yeast and mammals have revealed that the defects of RNRs result in imbalanced dNTP pools in vivo, which generally leads to increased DNA mutations, DNA breaks, cell death, and p53-dependent apoptosis.[2,135] The Arabidopsis genome encodes three R2-like proteins, including TSO2 (At3g27060), RNR2A (At3g23580), and RNR2B (At5g40942). Interestingly, the individual genes contribute to unique aspects of the cellular response to DNA damage. For instance, the expression of RNR2A and RNR2B is specifically activated by the replication-blocking agent hydroxyurea (HU) but not by DNA double-strand break inducer bleomycin (BLM).[136] On the other hand, the transcription of TSO2 is only induced in response to BLM.[136] The tso2 single and tso2rnr2a double mutants show extreme sensitivity to UV-C light.[134] Importantly, these mutants exhibit increased DNA damage and PCD relative to the wild-type.[134] These results further indicate that the R2 subunits of RNRs function critically in genome stability maintenance.
Summary
The Arabidopsis iron-containing proteins involved in genome stability share a high degree of functional conservation with mammals and yeast. Interestingly, some proteins, such as DNA polymerase d and ε, FANCJ helicase homologues, iron SODs, and small subunits of RNR, evolve several copies in Arabidopsis relative to mammals and yeast. As a result, some of them may function redundantly. Therefore, their single mutants have no visible phenotype in DNA damage stresses, resulting in difficulties to characterize their functions. Moreover, plants also evolve numerous CYP proteins that function in the oxidation/reduction of endogenous or exogenous compounds. However, the functions of individual genes in this superfamily are poorly understood, especially in genome stability maintenance.
Although studies have extensively demonstrated that iron-containing proteins are required for genome stability, limited information is available regarding their functional mechanisms. Several iron-containing proteins such as DNA helicase RAD3 and small subunits of RNR directly participate in DNA replication and repair. Some iron-containing DNA glycosylases are involved in genome stability due to their participation in DNA methylation. Some CIA components of Fe-S proteins are involved in genome stability possibly because they can transfer Fe-S clusters to target proteins such as Pold, DNA primase, Dna2, XPD, RTEL1, and FANCJ. Importantly, numerous hemoproteins in Arabidopsis function in electron transport; as such, defects in them generally cause the induction of ROS, which can damage DNA, proteins, and lipids. Taken together, although considerable progress has been made in the past years, further studies are still needed to uncover the functions of these iron-containing proteins, especially in genome stability maintenance.
Acknowledgments
We apologize to our colleagues whose work is not cited due to limitations of space.
Footnotes
Source of Support: Nil.
Conflict of Interest: None declared.
References
- 1.Lill R. Function and biogenesis of iron-sulphur proteins. Nature. 2009;460:831–8. doi: 10.1038/nature08301. [DOI] [PubMed] [Google Scholar]
- 2.Zhang C. Essential functions of iron-requiring proteins in DNA replication, repair and cell cycle control. Protein Cell. 2014;5:750–60. doi: 10.1007/s13238-014-0083-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang C, Zhang F. Iron homeostasis and tumorigenesis: Molecular mechanisms and therapeutic opportunities. Protein Cell. 2015;6:88–100. doi: 10.1007/s13238-014-0119-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Crichton RR. 3rd ed. Chichester, UK: John Wiley & Sons; 2009. Iron Metabolism: From Molecular Mechanisms to Clinical Consequences; pp. 223–69. [Google Scholar]
- 5.Johnson DC, Dean DR, Smith AD, Johnson MK. Structure, function, and formation of biological iron-sulfur clusters. Annu Rev Biochem. 2005;74:247–81. doi: 10.1146/annurev.biochem.74.082803.133518. [DOI] [PubMed] [Google Scholar]
- 6.Prabhulkar S, Tian H, Wang X, Zhu JJ, Li CZ. Engineered proteins: Redox properties and their applications. Antioxid Redox Signal. 2012;17:1796–822. doi: 10.1089/ars.2011.4001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lill R, Dutkiewicz R, Elsässer HP, Hausmann A, Netz DJ, Pierik AJ, et al. Mechanisms of iron-sulfur protein maturation in mitochondria, cytosol and nucleus of eukaryotes. Biochim Biophys Acta. 2006;1763:652–67. doi: 10.1016/j.bbamcr.2006.05.011. [DOI] [PubMed] [Google Scholar]
- 8.Ouattara M, Pennati A, Devlin DJ, Huang YS, Gadda G, Eichenbaum Z. Kinetics of heme transfer by the Shr NEAT domains of Group A Streptococcus. Arch Biochem Biophys. 2013;538:71–9. doi: 10.1016/j.abb.2013.08.009. [DOI] [PubMed] [Google Scholar]
- 9.Murphy ER, Sacco RE, Dickenson A, Metzger DJ, Hu Y, Orndorff PE, et al. BhuR, a virulence-associated outer membrane protein of Bordetella avium, is required for the acquisition of iron from heme and hemoproteins. Infect Immun. 2002;70:5390–403. doi: 10.1128/IAI.70.10.5390-5403.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dlouhy AC, Outten CE. The iron metallome in eukaryotic organisms. Met Ions Life Sci. 2013;12:241–78. doi: 10.1007/978-94-007-5561-1_8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miner KD, Klose KE, Kurtz DM., Jr An HD-GYP cyclic di-guanosine monophosphate phosphodiesterase with a non-heme diiron-carboxylate active site. Biochemistry. 2013;52:5329–31. doi: 10.1021/bi4009215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Andreini C, Bertini I, Cavallaro G, Najmanovich RJ, Thornton JM. Structural analysis of metal sites in proteins: Non-heme iron sites as a case study. J Mol Biol. 2009;388:356–80. doi: 10.1016/j.jmb.2009.02.052. [DOI] [PubMed] [Google Scholar]
- 13.Costas M, Mehn MP, Jensen MP, Que L Jr. Dioxygen activation at mononuclear nonheme iron active sites: Enzymes, models, and intermediates. Chem Rev. 2004;104:939–86. doi: 10.1021/cr020628n. [DOI] [PubMed] [Google Scholar]
- 14.Couturier J, Touraine B, Briat JF, Gaymard F, Rouhier N. The iron-sulfur cluster assembly machineries in plants: Current knowledge and open questions. Front Plant Sci. 2013;4:259. doi: 10.3389/fpls.2013.00259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Weerapana E, Wang C, Simon GM, Richter F, Khare S, Dillon MB, et al. Quantitative reactivity profiling predicts functional cysteines in proteomes. Nature. 2010;468:790–5. doi: 10.1038/nature09472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gari K, León Ortiz AM, Borel V, Flynn H, Skehel JM, Boulton SJ. MMS19 links cytoplasmic iron-sulfur cluster assembly to DNA metabolism. Science. 2012;337:243–5. doi: 10.1126/science.1219664. [DOI] [PubMed] [Google Scholar]
- 17.Stehling O, Vashisht AA, Mascarenhas J, Jonsson ZO, Sharma T, Netz DJ, et al. MMS19 assembles iron-sulfur proteins required for DNA metabolism and genomic integrity. Science. 2012;337:195–9. doi: 10.1126/science.1219723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vigani G, Zocchi G, Bashir K, Philippar K, Briat JF. Cellular iron homeostasis and metabolism in plant. Front Plant Sci. 2013;4:490. doi: 10.3389/fpls.2013.00490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zheng SJ. Iron homeostasis and iron acquisition in plants: Maintenance, functions and consequences. Ann Bot. 2010;105:799–800. doi: 10.1093/aob/mcq082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Deng X, Eriksson M. Two iron-responsive promoter elements control expression of FOX1 in Chlamydomonas reinhardtii. Eukaryot Cell. 2007;6:2163–7. doi: 10.1128/EC.00324-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Von Wiren N, Mori S, Marschner H, Romheld V. Iron inefficiency in Maize Mutant ys1 (Zea mays L.cv Yellow-Stripe) is caused by a defect in uptake of iron phytosiderophores. Plant Physiol. 1994;106:71–7. doi: 10.1104/pp.106.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Vasconcelos MW, Clemente TE, Grusak MA. Evaluation of constitutive iron reductase (AtFRO2) expression on mineral accumulation and distribution in soybean (Glycine max.L) Front Plant Sci. 2014;5:112. doi: 10.3389/fpls.2014.00112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vert GA, Briat JF, Curie C. Dual regulation of the Arabidopsis high-affinity root iron uptake system by local and long-distance signals. Plant Physiol. 2003;132:796–804. doi: 10.1104/pp.102.016089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vert G, Grotz N, Dédaldéchamp F, Gaymard F, Guerinot ML, Briat JF, et al. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell. 2002;14:1223–33. doi: 10.1105/tpc.001388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vert G, Barberon M, Zelazny E, Séguéla M, Briat JF, Curie C. Arabidopsis IRT2 cooperates with the high-affinity iron uptake system to maintain iron homeostasis in root epidermal cells. Planta. 2009;229:1171–9. doi: 10.1007/s00425-009-0904-8. [DOI] [PubMed] [Google Scholar]
- 26.Robinson NJ, Procter CM, Connolly EL, Guerinot ML. A ferric-chelate reductase for iron uptake from soils. Nature. 1999;397:694–7. doi: 10.1038/17800. [DOI] [PubMed] [Google Scholar]
- 27.Connolly EL, Fett JP, Guerinot ML. Expression of the IRT1 metal transporter is controlled by metals at the levels of transcript and protein accumulation. Plant Cell. 2002;14:1347–57. doi: 10.1105/tpc.001263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Colangelo EP, Guerinot ML. The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell. 2004;16:3400–12. doi: 10.1105/tpc.104.024315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yuan Y, Wu H, Wang N, Li J, Zhao W, Du J, et al. FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res. 2008;18:385–97. doi: 10.1038/cr.2008.26. [DOI] [PubMed] [Google Scholar]
- 30.Wang N, Cui Y, Liu Y, Fan H, Du J, Huang Z, et al. Requirement and functional redundancy of Ib subgroup bHLH proteins for iron deficiency responses and uptake in Arabidopsis thaliana. Mol Plant. 2013;6:503–13. doi: 10.1093/mp/sss089. [DOI] [PubMed] [Google Scholar]
- 31.Zhang Y, Wu H, Wang N, Fan H, Chen C, Cui Y, et al. Mediator subunit 16 functions in the regulation of iron uptake gene expression in Arabidopsis. New Phytol. 2014;203:770–83. doi: 10.1111/nph.12860. [DOI] [PubMed] [Google Scholar]
- 32.Fan H, Zhang Z, Wang N, Cui Y, Sun H, Liu Y, et al. SKB1/PRMT5-mediated histone H4R3 dimethylation of Ib subgroup bHLH genes negatively regulates iron homeostasis in Arabidopsis thaliana. Plant J. 2014;77:209–21. doi: 10.1111/tpj.12380. [DOI] [PubMed] [Google Scholar]
- 33.Giehl RF, Lima JE, von Wirén N. Regulatory components involved in altering lateral root development in response to localized iron: Evidence for natural genetic variation. Plant Signal Behav. 2012;7:711–3. doi: 10.4161/psb.20337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Schikora A, Schmidt W. Iron stress-induced changes in root epidermal cell fate are regulated independently from physiological responses to low iron availability. Plant Physiol. 2001;125:1679–87. doi: 10.1104/pp.125.4.1679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Duy D, Wanner G, Meda AR, von Wirén N, Soll J, Philippar K. PIC1, an ancient permease in Arabidopsis chloroplasts, mediates iron transport. Plant Cell. 2007;19:986–1006. doi: 10.1105/tpc.106.047407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Palmer CM, Hindt MN, Schmidt H, Clemens S, Guerinot ML. MYB10 and MYB72 are required for growth under iron-limiting conditions. PLoS Genet. 2013;9:e1003953. doi: 10.1371/journal.pgen.1003953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mukherjee I, Campbell NH, Ash JS, Connolly EL. Expression profiling of the Arabidopsis ferric chelate reductase (FRO) gene family reveals differential regulation by iron and copper. Planta. 2006;223:1178–90. doi: 10.1007/s00425-005-0165-0. [DOI] [PubMed] [Google Scholar]
- 38.Lanquar V, Lelièvre F, Bolte S, Hamès C, Alcon C, Neumann D, et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005;24:4041–51. doi: 10.1038/sj.emboj.7600864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lanquar V, Ramos MS, Lelièvre F, Barbier-Brygoo H, Krieger-Liszkay A, Krämer U, et al. Export of vacuolar manganese by AtNRAMP3 and AtNRAMP4 is required for optimal photosynthesis and growth under manganese deficiency. Plant Physiol. 2010;152:1986–99. doi: 10.1104/pp.109.150946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Curie C, Briat JF. Iron transport and signaling in plants. Annu Rev Plant Biol. 2003;54:183–206. doi: 10.1146/annurev.arplant.54.031902.135018. [DOI] [PubMed] [Google Scholar]
- 41.Thomine S, Wang R, Ward JM, Crawford NM, Schroeder JI. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc Natl Acad Sci U S A. 2000;97:4991–6. doi: 10.1073/pnas.97.9.4991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Thomine S, Lelièvre F, Debarbieux E, Schroeder JI, Barbier-Brygoo H. AtNRAMP3, a multispecific vacuolar metal transporter involved in plant responses to iron deficiency. Plant J. 2003;34:685–95. doi: 10.1046/j.1365-313x.2003.01760.x. [DOI] [PubMed] [Google Scholar]
- 43.Kim SA, Punshon T, Lanzirotti A, Li L, Alonso JM, Ecker JR, et al. Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1. Science. 2006;314:1295–8. doi: 10.1126/science.1132563. [DOI] [PubMed] [Google Scholar]
- 44.Ravet K, Touraine B, Boucherez J, Briat JF, Gaymard F, Cellier F. Ferritins control interaction between iron homeostasis and oxidative stress in Arabidopsis. Plant J. 2009;57:400–12. doi: 10.1111/j.1365-313X.2008.03698.x. [DOI] [PubMed] [Google Scholar]
- 45.Petit JM, Briat JF, Lobréaux S. Structure and differential expression of the four members of the Arabidopsis thaliana ferritin gene family. Biochem J. 2001;359:575–82. doi: 10.1042/0264-6021:3590575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Briat JF, Duc C, Ravet K, Gaymard F. Ferritins and iron storage in plants. Biochim Biophys Acta. 2010;1800:806–14. doi: 10.1016/j.bbagen.2009.12.003. [DOI] [PubMed] [Google Scholar]
- 47.Duc C, Cellier F, Lobréaux S, Briat JF, Gaymard F. Regulation of iron homeostasis in Arabidopsis thaliana by the clock regulator time for coffee. J Biol Chem. 2009;284:36271–81. doi: 10.1074/jbc.M109.059873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kushnir S, Babiychuk E, Storozhenko S, Davey MW, Papenbrock J, De Rycke R, et al. A mutation of the mitochondrial ABC transporter Sta1 leads to dwarfism and chlorosis in the Arabidopsis mutant starik. Plant Cell. 2001;13:89–100. doi: 10.1105/tpc.13.1.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Grennan AK. Identification of genes involved in metal transport in plants. Plant Physiol. 2009;149:1623–4. doi: 10.1104/pp.109.900287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jain A, Wilson GT, Connolly EL. The diverse roles of FRO family metalloreductases in iron and copper homeostasis. Front Plant Sci. 2014;5:100. doi: 10.3389/fpls.2014.00100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kobayashi T, Nishizawa NK. Iron uptake, translocation, and regulation in higher plants. Annu Rev Plant Biol. 2012;63:131–52. doi: 10.1146/annurev-arplant-042811-105522. [DOI] [PubMed] [Google Scholar]
- 52.DiDonato RJ Jr, Roberts LA, Sanderson T, Eisley RB, Walker EL. Arabidopsis Yellow Stripe-Like2 (YSL2): A metal-regulated gene encoding a plasma membrane transporter of nicotianamine-metal complexes. Plant J. 2004;39:403–14. doi: 10.1111/j.1365-313X.2004.02128.x. [DOI] [PubMed] [Google Scholar]
- 53.Waters BM, Chu HH, Didonato RJ, Roberts LA, Eisley RB, Lahner B, et al. Mutations in Arabidopsis yellow stripe-like1 and yellow stripe-like3 reveal their roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 2006;141:1446–58. doi: 10.1104/pp.106.082586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Divol F, Couch D, Conéjéro G, Roschzttardtz H, Mari S, Curie C. The Arabidopsis YELLOW STRIPE LIKE4 and 6 transporters control iron release from the chloroplast. Plant Cell. 2013;25:1040–55. doi: 10.1105/tpc.112.107672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Briat JF, Curie C, Gaymard F. Iron utilization and metabolism in plants. Curr Opin Plant Biol. 2007;10:276–82. doi: 10.1016/j.pbi.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 56.Thompson JW, Bruick RK. Protein degradation and iron homeostasis. Biochim Biophys Acta. 2012;1823:1484–90. doi: 10.1016/j.bbamcr.2012.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Miyabe I, Kunkel TA, Carr AM. The major roles of DNA polymerases epsilon and delta at the eukaryotic replication fork are evolutionarily conserved. PLoS Genet. 2011;7:e1002407. doi: 10.1371/journal.pgen.1002407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Schumacher SB, Stucki M, Hübscher U. The N-terminal region of DNA polymerase delta catalytic subunit is necessary for holoenzyme function. Nucleic Acids Res. 2000;28:620–5. doi: 10.1093/nar/28.2.620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang X, Ira G, Tercero JA, Holmes AM, Diffley JF, Haber JE. Role of DNA replication proteins in double-strand break-induced recombination in Saccharomyces cerevisiae. Mol Cell Biol. 2004;24:6891–9. doi: 10.1128/MCB.24.16.6891-6899.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Shultz RW, Tatineni VM, Hanley-Bowdoin L, Thompson WF. Genome-wide analysis of the core DNA replication machinery in the higher plants Arabidopsis and rice. Plant Physiol. 2007;144:1697–714. doi: 10.1104/pp.107.101105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Uchiyama Y, Hatanaka M, Kimura S, Ishibashi T, Ueda T, Sakakibara Y, et al. Characterization of DNA polymerase delta from a higher plant, rice (Oryza sativa L.) Gene. 2002;295:19–26. doi: 10.1016/s0378-1119(02)00822-3. [DOI] [PubMed] [Google Scholar]
- 62.Garcia E, Quiroz F, Uchiyama Y, Sakaguchi K, Vázquez-Ramos JM. Expression of a maize delta-type DNA polymerase during seed germination. Physiol Plant. 2006;127:268–76. [Google Scholar]
- 63.Ronceret A, Guilleminot J, Lincker F, Gadea-Vacas J, Delorme V, Bechtold N, et al. Genetic analysis of two Arabidopsis DNA polymerase epsilon subunits during early embryogenesis. Plant J. 2005;44:223–36. doi: 10.1111/j.1365-313X.2005.02521.x. [DOI] [PubMed] [Google Scholar]
- 64.Jenik PD, Jurkuta RE, Barton MK. Interactions between the cell cycle and embryonic patterning in Arabidopsis uncovered by a mutation in DNA polymerase epsilon. Plant Cell. 2005;17:3362–77. doi: 10.1105/tpc.105.036889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kellermayer R. The versatile RECQL4. Genet Med. 2006;8:213–6. doi: 10.1097/01.gim.0000214457.58378.1a. [DOI] [PubMed] [Google Scholar]
- 66.van Brabant AJ, Stan R, Ellis NA. DNA helicases, genomic instability, and human genetic disease. Annu Rev Genomics Hum Genet. 2000;1:409–59. doi: 10.1146/annurev.genom.1.1.409. [DOI] [PubMed] [Google Scholar]
- 67.Fan L, Fuss JO, Cheng QJ, Arvai AS, Hammel M, Roberts VA, et al. XPD helicase structures and activities: Insights into the cancer and aging phenotypes from XPD mutations. Cell. 2008;133:789–800. doi: 10.1016/j.cell.2008.04.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sung P, Prakash L, Matson SW, Prakash S. RAD3 protein of Saccharomyces cerevisiae is a DNA helicase. Proc Natl Acad Sci USA. 1987;84:8951–5. doi: 10.1073/pnas.84.24.8951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Pugh RA, Honda M, Leesley H, Thomas A, Lin Y, Nilges MJ, et al. The iron-containing domain is essential in Rad3 helicases for coupling of ATP hydrolysis to DNA translocation and for targeting the helicase to the single-stranded DNA-double-stranded DNA junction. J Biol Chem. 2008;283:1732–43. doi: 10.1074/jbc.M707064200. [DOI] [PubMed] [Google Scholar]
- 70.Liu Z, Hong SW, Escobar M, Vierling E, Mitchell DL, Mount DW, et al. Arabidopsis UVH6, a homolog of human XPD and yeast RAD3 DNA repair genes, functions in DNA repair and is essential for plant growth. Plant Physiol. 2003;132:1405–14. doi: 10.1104/pp.103.021808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wu Y, Brosh RM JR. DNA helicase and helicase-nuclease enzymes with a conserved iron-sulfur cluster. Nucleic Acids Res. 2012;40:4247–60. doi: 10.1093/nar/gks039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shah N, Inoue A, Woo Lee S, Beishline K, Lahti JM, Noguchi E. Roles of ChlR1 DNA helicase in replication recovery from DNA damage. Exp Cell Res. 2013;319:2244–53. doi: 10.1016/j.yexcr.2013.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jacobs AL, Schär P. DNA glycosylases: In DNA repair and beyond. Chromosoma. 2012;121:1–20. doi: 10.1007/s00412-011-0347-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Sun B, Latham KA, Dodson ML, Lloyd RS. Studies on the catalytic mechanism of five DNA glycosylases. Probing for enzyme-DNA imino intermediates. J Biol Chem. 1995;270:19501–8. doi: 10.1074/jbc.270.33.19501. [DOI] [PubMed] [Google Scholar]
- 75.Krokan HE, Standal R, Slupphaug G. DNA glycosylases in the base excision repair of DNA. Biochem J. 1997;325:1–16. doi: 10.1042/bj3250001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Agius F, Kapoor A, Zhu JK. Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation. Proc Natl Acad Sci U S A. 2006;103:11796–801. doi: 10.1073/pnas.0603563103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wyatt MD, Allan JM, Lau AY, Ellenberger TE, Samson LD. 3-methyladenine DNA glycosylases: Structure, function, and biological importance. Bioessays. 1999;21:668–76. doi: 10.1002/(SICI)1521-1878(199908)21:8<668::AID-BIES6>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 78.Gong Z, Morales-Ruiz T, Ariza RR, Roldán-Arjona T, David L, Zhu JK. ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell. 2002;111:803–14. doi: 10.1016/s0092-8674(02)01133-9. [DOI] [PubMed] [Google Scholar]
- 79.Lister R, O’Malley RC, Tonti-Filippini J, Gregory BD, Berry CC, Millar AH, et al. Highly integrated single-base resolution maps of the epigenome in Arabidopsis. Cell. 2008;133:523–36. doi: 10.1016/j.cell.2008.03.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ortega-Galisteo AP, Morales-Ruiz T, Ariza RR, Roldán-Arjona T. Arabidopsis DEMETER-LIKE proteins DML2 and DML3 are required for appropriate distribution of DNA methylation marks. Plant Mol Biol. 2008;67:671–81. doi: 10.1007/s11103-008-9346-0. [DOI] [PubMed] [Google Scholar]
- 81.Schoft VK, Chumak N, Choi Y, Hannon M, Garcia-Aguilar M, Machlicova A, et al. Function of the DEMETER DNA glycosylase in the Arabidopsis thaliana male gametophyte. Proc Natl Acad Sci U S A. 2011;108:8042–7. doi: 10.1073/pnas.1105117108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wöhrmann HJ, Gagliardini V, Raissig MT, Wehrle W, Arand J, Schmidt A, et al. Identification of a DNA methylation-independent imprinting control region at the Arabidopsis MEDEA locus. Genes Dev. 2012;26:1837–50. doi: 10.1101/gad.195123.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Choi Y, Gehring M, Johnson L, Hannon M, Harada JJ, Goldberg RB, et al. DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in arabidopsis. Cell. 2002;110:33–42. doi: 10.1016/s0092-8674(02)00807-3. [DOI] [PubMed] [Google Scholar]
- 84.Lill R, Mühlenhoff U. Maturation of iron-sulfur proteins in eukaryotes: Mechanisms, connected processes, and diseases. Annu Rev Biochem. 2008;77:669–700. doi: 10.1146/annurev.biochem.76.052705.162653. [DOI] [PubMed] [Google Scholar]
- 85.Netz DJ, Mascarenhas J, Stehling O, Pierik AJ, Lill R. Maturation of cytosolic and nuclear iron-sulfur proteins. Trends Cell Biol. 2014;24:303–12. doi: 10.1016/j.tcb.2013.11.005. [DOI] [PubMed] [Google Scholar]
- 86.Couturier J, Touraine B, Briat JF, Gaymard F, Rouhier N. The iron-sulfur cluster assembly machineries in plants: Current knowledge and open questions. Front Plant Sci. 2013;4:259. doi: 10.3389/fpls.2013.00259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Weerapana E, Wang C, Simon GM, Richter F, Khare S, Dillon MB, et al. Quantitative reactivity profiling predicts functional cysteines in proteomes. Nature. 2010;468:790–5. doi: 10.1038/nature09472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Luo D, Bernard DG, Balk J, Hai H, Cui X. The DUF59 family gene AE7 acts in the cytosolic iron-sulfur cluster assembly pathway to maintain nuclear genome integrity in Arabidopsis. Plant Cell. 2012;24:4135–48. doi: 10.1105/tpc.112.102608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Farooq MA, Pracheil TM, Dong Z, Xiao F, Liu Z. Mitochondrial DNA instability in cells lacking aconitase correlates with iron citrate toxicity. Oxid Med Cell Longev 2013. 2013:493536. doi: 10.1155/2013/493536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Arnaud N, Ravet K, Borlotti A, Touraine B, Boucherez J, Fizames C, et al. The iron-responsive element (IRE)/iron-regulatory protein 1 (IRP1)-cytosolic aconitase iron-regulatory switch does not operate in plants. Biochem J. 2007;405:523–31. doi: 10.1042/BJ20061874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Sunkar R, Kapoor A, Zhu JK. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell. 2006;18:2051–65. doi: 10.1105/tpc.106.041673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhao Z, Assmann SM. The glycolytic enzyme, phosphoglycerate mutase, has critical roles in stomatal movement, vegetative growth, and pollen production in Arabidopsis thaliana. J Exp Bot. 2011;62:5179–89. doi: 10.1093/jxb/err223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hanke GT, Kurisu G, Kusunoki M, Hase T. Fd: FNR electron transfer complexes: Evolutionary refinement of structural interactions. Photosynth Res. 2004;81:317–27. doi: 10.1023/B:PRES.0000036885.01534.b8. [DOI] [PubMed] [Google Scholar]
- 94.Hanke GT, Satomi Y, Shinmura K, Takao T, Hase T. A screen for potential ferredoxin electron transfer partners uncovers new, redox dependent interactions. Biochim Biophys Acta. 2011;1814:366–74. doi: 10.1016/j.bbapap.2010.09.011. [DOI] [PubMed] [Google Scholar]
- 95.Hanke G, Mulo P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ. 2013;36:1071–84. doi: 10.1111/pce.12046. [DOI] [PubMed] [Google Scholar]
- 96.Keryer E, Collin V, Lavergne D, Lemaire S, Issakidis-Bourguet E. Characterization of Arabidopsis mutants for the variable subunit of ferredoxin: Thioredoxin reductase. Photosynth Res. 2004;79:265–74. doi: 10.1023/B:PRES.0000017173.46185.3e. [DOI] [PubMed] [Google Scholar]
- 97.Stevens JM, Uchida T, Daltrop O, Ferguson SJ. Covalent cofactor attachment to proteins: Cytochrome c biogenesis. Biochem Soc Trans. 2005;33:792–5. doi: 10.1042/BST0330792. [DOI] [PubMed] [Google Scholar]
- 98.Mense SM, Zhang L. Heme: A versatile signaling molecule controlling the activities of diverse regulators ranging from transcription factors to MAP kinases. Cell Res. 2006;16:681–92. doi: 10.1038/sj.cr.7310086. [DOI] [PubMed] [Google Scholar]
- 99.Ohashi T, Mizutani A, Murakami A, Kojo S, Ishii T, Taketani S. Rapid oxidation of dichlorodihydrofluorescin with heme and hemoproteins: Formation of the fluorescein is independent of the generation of reactive oxygen species. FEBS Lett. 2002;511:21–7. doi: 10.1016/s0014-5793(01)03262-8. [DOI] [PubMed] [Google Scholar]
- 100.Danielson PB. The cytochrome P450 superfamily: Biochemistry, evolution and drug metabolism in humans. Curr Drug Metab. 2002;3:561–97. doi: 10.2174/1389200023337054. [DOI] [PubMed] [Google Scholar]
- 101.Gan L, von Moltke LL, Trepanier LA, Harmatz JS, Greenblatt DJ, Court MH. Role of NADPH-cytochrome P450 reductase and cytochrome-b5/NADH-b5 reductase in variability of CYP3A activity in human liver microsomes. Drug Metab Dispos. 2009;37:90–6. doi: 10.1124/dmd.108.023424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Jensen K, Møller BL. Plant NADPH-cytochrome P450 oxidoreductases. Phytochemistry. 2010;71:132–41. doi: 10.1016/j.phytochem.2009.10.017. [DOI] [PubMed] [Google Scholar]
- 103.Gutierrez A, Paine M, Wolf CR, Scrutton NS, Roberts GC. Relaxation kinetics of cytochrome P450 reductase: Internal electron transfer is limited by conformational change and regulated by coenzyme binding. Biochemistry. 2002;41:4626–37. doi: 10.1021/bi0159433. [DOI] [PubMed] [Google Scholar]
- 104.Aigrain L, Fatemi F, Frances O, Lescop E, Truan G. Dynamic control of electron transfers in diflavin reductases. Int J Mol Sci. 2012;13:15012–41. doi: 10.3390/ijms131115012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Heine T, Glatt H, Epe B. Human cytochrome P450 reductase can act as a source of endogenous oxidative DNA damage and genetic instability. Free Radic Biol Med. 2006;40:801–7. doi: 10.1016/j.freeradbiomed.2005.10.033. [DOI] [PubMed] [Google Scholar]
- 106.Werck-Reichhart D, Bak S, Paquette S. Cytochromes p450. Arabidopsis Book. 2002;1:e0028. doi: 10.1199/tab.0028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Fukuchi-Mizutani M, Mizutani M, Tanaka Y, Kusumi T, Ohta D. Microsomal electron transfer in higher plants: Cloning and heterologous expression of NADH-cytochrome b5 reductase from Arabidopsis. Plant Physiol. 1999;119:353–62. doi: 10.1104/pp.119.1.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Urban P, Mignotte C, Kazmaier M, Delorme F, Pompon D. Cloning, yeast expression, and characterization of the coupling of two distantly related Arabidopsis thaliana NADPH-cytochrome P450 reductases with P450 CYP73A5. J Biol Chem. 1997;272:19176–86. doi: 10.1074/jbc.272.31.19176. [DOI] [PubMed] [Google Scholar]
- 109.Varadarajan J, Guilleminot J, Saint-Jore-Dupas C, Piégu B, Chabouté ME, Gomord V, et al. ATR3 encodes a diflavin reductase essential for Arabidopsis embryo development. New Phytol. 2010;187:67–82. doi: 10.1111/j.1469-8137.2010.03254.x. [DOI] [PubMed] [Google Scholar]
- 110.Kumar R, Tran LS, Neelakandan AK, Nguyen HT. Higher plant cytochrome b5 polypeptides modulate fatty acid desaturation. PloS One. 2012;7:e31370. doi: 10.1371/journal.pone.0031370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Shanklin J, Cahoon EB. Desaturation and related modifications of fatty acids1. Annu Rev Plant Physiol Plant Mol Biol. 1998;49:611–41. doi: 10.1146/annurev.arplant.49.1.611. [DOI] [PubMed] [Google Scholar]
- 112.McConn M, Browse J. Polyunsaturated membranes are required for photosynthetic competence in a mutant of Arabidopsis. Plant J. 1998;15:521–30. doi: 10.1046/j.1365-313x.1998.00229.x. [DOI] [PubMed] [Google Scholar]
- 113.Mallory JC, Crudden G, Johnson BL, Mo C, Pierson CA, Bard M, et al. Dap1p, a heme-binding protein that regulates the cytochrome P450 protein Erg11p/Cyp51p in Saccharomyces cerevisiae. Mol Cell Biol. 2005;25:1669–79. doi: 10.1128/MCB.25.5.1669-1679.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Guénolé A, Srivas R, Vreeken K, Wang ZZ, Wang S, Krogan NJ, et al. Dissection of DNA damage responses using multiconditional genetic interaction maps. Mol Cell. 2013;49:346–58. doi: 10.1016/j.molcel.2012.11.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Nagano M, Ihara-Ohori Y, Imai H, Inada N, Fujimoto M, Tsutsumi N, et al. Functional association of cell death suppressor, Arabidopsis Bax inhibitor-1, with fatty acid 2-hydroxylation through cytochrome b 5. Plant J. 2009;58:122–34. doi: 10.1111/j.1365-313X.2008.03765.x. [DOI] [PubMed] [Google Scholar]
- 116.Hwang YT, Pelitire SM, Henderson MP, Andrews DW, Dyer JM, Mullen RT. Novel targeting signals mediate the sorting of different isoforms of the tail-anchored membrane protein cytochrome b5 to either endoplasmic reticulum or mitochondria. Plant Cell. 2004;16:3002–19. doi: 10.1105/tpc.104.026039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Wayne LL, Wallis JG, Kumar R, Markham JE, Browse J. Cytochrome b5 reductase encoded by CBR1 is essential for a functional male gametophyte in Arabidopsis. Plant Cell. 2013;25:3052–66. doi: 10.1105/tpc.113.113324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Welchen E, Hildebrandt TM, Lewejohann D, Gonzalez DH, Braun HP. Lack of cytochrome c in Arabidopsis decreases stability of Complex IV and modifies redox metabolism without affecting Complexes I and III. Biochim Biophys Acta. 2012;1817:990–1001. doi: 10.1016/j.bbabio.2012.04.008. [DOI] [PubMed] [Google Scholar]
- 119.Jiang X, Wang X. Cytochrome C-mediated apoptosis. Annu Rev Biochem. 2004;73:87–106. doi: 10.1146/annurev.biochem.73.011303.073706. [DOI] [PubMed] [Google Scholar]
- 120.Huttemann M, Pecina P, Rainbolt M, Sanderson TH, Kagan VE, Samavati L, et al. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion. 2011;11:369–81. doi: 10.1016/j.mito.2011.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Grzyska PK, Appelman EH, Hausinger RP, Proshlyakov DA. Insight into the mechanism of an iron dioxygenase by resolution of steps following the FeIV=HO species. Proc Natl Acad Sci U S A. 2010;107:3982–7. doi: 10.1073/pnas.0911565107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Yi C, Jia G, Hou G, Dai Q, Zhang W, Zheng G, et al. Iron-catalysed oxidation intermediates captured in a DNA repair dioxygenase. Nature. 2010;468:330–3. doi: 10.1038/nature09497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Meza TJ, Moen MN, Vågbø CB, Krokan HE, Klungland A, Grini PE, et al. The DNA dioxygenase ALKBH2 protects Arabidopsis thaliana against methylation damage. Nucleic Acids Res. 2012;40:6620–31. doi: 10.1093/nar/gks327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Falnes PØ, Johansen RF, Seeberg E. AlkB-mediated oxidative demethylation reverses DNA damage in Escherichia coli. Nature. 2002;419:178–82. doi: 10.1038/nature01048. [DOI] [PubMed] [Google Scholar]
- 125.De León IP, Sanz A, Hamberg M, Castresana C. Involvement of the Arabidopsis alpha-DOX1 fatty acid dioxygenase in protection against oxidative stress and cell death. Plant J. 2002;29:61–2. doi: 10.1046/j.1365-313x.2002.01195.x. [DOI] [PubMed] [Google Scholar]
- 126.García-Marcos A, Pacheco R, Manzano A, Aguilar E, Tenllado F. Oxylipin biosynthesis genes positively regulate programmed cell death during compatible infections with the synergistic pair potato virus X-potato virus Y and Tomato spotted wilt virus. J Virol. 2013;87:5769–83. doi: 10.1128/JVI.03573-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Friedman EJ, Wang HX, Jiang K, Perovic I, Deshpande A, Pochapsky TC, et al. Acireductone dioxygenase 1 (ARD1) is an effector of the heterotrimeric G protein beta subunit in Arabidopsis. J Biol Chem. 2011;286:30107–18. doi: 10.1074/jbc.M111.227256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Weng JK, Li Y, Mo H, Chapple C. Assembly of an evolutionarily new pathway for α-pyrone biosynthesis in Arabidopsis. Science. 2012;337:960–4. doi: 10.1126/science.1221614. [DOI] [PubMed] [Google Scholar]
- 129.Kliebenstein DJ, Monde RA, Last RL. Superoxide dismutase in Arabidopsis: An eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 1998;118:637–50. doi: 10.1104/pp.118.2.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Myouga F, Hosoda C, Umezawa T, Iizumi H, Kuromori T, Motohashi R, et al. A heterocomplex of iron superoxide dismutases defends chloroplast nucleoids against oxidative stress and is essential for chloroplast development in Arabidopsis. Plant Cell. 2008;20:3148–62. doi: 10.1105/tpc.108.061341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Asada K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:601–39. doi: 10.1146/annurev.arplant.50.1.601. [DOI] [PubMed] [Google Scholar]
- 132.Abdel-Ghany SE, Pilon M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J Biol Chem. 2008;283:15932–45. doi: 10.1074/jbc.M801406200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Zhang Y, Wu H, Wang N, Fan H, Chen C, Cui Y, et al. Mediator subunit 16 functions in the regulation of iron uptake gene expression in Arabidopsis. New Phytol. 2014;203:770–83. doi: 10.1111/nph.12860. [DOI] [PubMed] [Google Scholar]
- 134.Wang C, Liu Z. Arabidopsis ribonucleotide reductases are critical for cell cycle progression, DNA damage repair, and plant development. Plant Cell. 2006;18:350–65. doi: 10.1105/tpc.105.037044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kumar D, Viberg J, Nilsson AK, Chabes A. Highly mutagenic and severely imbalanced dNTP pools can escape detection by the S-phase checkpoint. Nucleic Acids Res. 2010;38:3975–83. doi: 10.1093/nar/gkq128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Roa H, Lang J, Culligan KM, Keller M, Holec S, Cognat V, et al. Ribonucleotide reductase regulation in response to genotoxic stress in Arabidopsis. Plant Physiol. 2009;151:461–71. doi: 10.1104/pp.109.140053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Moeder W, Del Pozo O, Navarre DA, Martin GB, Klessig DF. Aconitase plays a role in regulating resistance to oxidative stress and cell death in Arabidopsis and Nicotiana benthamiana. Plant Mol Biol. 2007;63:273–87. doi: 10.1007/s11103-006-9087-x. [DOI] [PubMed] [Google Scholar]
- 138.Fukuchi-Mizutani M, Mizutani M, Tanaka Y, Kusumi T, Ohta D. Microsomal electron transfer in higher plants: Cloning and heterologous expression of NADH-cytochrome b5 reductase from Arabidopsis. Plant Physiol. 1999;119:353–62. doi: 10.1104/pp.119.1.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Farver O, Vitu E, Wherland S, Fass D, Pecht I. Electron transfer reactivity of the Arabidopsis thaliana sulfhydryl oxidase AtErv1. J Biol Chem. 2009;284:2098–105. doi: 10.1074/jbc.M806316200. [DOI] [PubMed] [Google Scholar]
- 140.Vandepoele K, Vlieghe K, Florquin K, Hennig L, Beemster GT, Gruissem W, et al. Genome-wide identification of potential plant E2F target genes. Plant Physiol. 2005;139:316–28. doi: 10.1104/pp.105.066290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Sudre D, Gutierrez-Carbonell E, Lattanzio G, Rellán-Álvarez R, Gaymard F, Wohlgemuth G, et al. Iron-dependent modifications of the flower transcriptome, proteome, metabolome, and hormonal content in an Arabidopsis ferritin mutant. J Exp Bot. 2013;64:2665–88. doi: 10.1093/jxb/ert112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Charron JB, Ouellet F, Houde M, Sarhan F. The plant Apolipoprotein D ortholog protects Arabidopsis against oxidative stress. BMC Plant Biol. 2008;8:86. doi: 10.1186/1471-2229-8-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Kliebenstein DJ, West MA, van Leeuwen H, Kim K, Doerge RW, Michelmore RW, et al. Genomic survey of gene expression diversity in Arabidopsis thaliana. Genetics. 2006;172:1179–89. doi: 10.1534/genetics.105.049353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Pommerrenig B, Feussner K, Zierer W, Rabinovych V, Klebl F, Feussner I, et al. Phloem-specific expression of Yang cycle genes and identification of novel Yang cycle enzymes in Plantago and Arabidopsis. Plant Cell. 2011;23:1904–19. doi: 10.1105/tpc.110.079657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Fey V, Wagner R, Braütigam K, Wirtz M, Hell R, Dietzmann A, et al. Retrograde plastid redox signals in the expression of nuclear genes for chloroplast proteins of Arabidopsis thaliana. J Biol Chem. 2005;280:5318–28. doi: 10.1074/jbc.M406358200. [DOI] [PubMed] [Google Scholar]
- 146.Takahashi K, Niwa H, Yokota N, Kubota K, Inoue H. Widespread tissue expression of nepenthesin-like aspartic protease genes in Arabidopsis thaliana. Plant Physiol Biochem. 2008;46:724–9. doi: 10.1016/j.plaphy.2008.04.007. [DOI] [PubMed] [Google Scholar]
- 147.Heyndrickx KS, Vandepoele K. Systematic identification of functional plant modules through the integration of complementary data sources. Plant Physiol. 2012;159:884–901. doi: 10.1104/pp.112.196725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ascencio-Ibáñez JT, Sozzani R, Lee TJ, Chu TM, Wolfinger RD, Cella R, et al. Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2008;148:436–54. doi: 10.1104/pp.108.121038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ly V, Hatherell A, Kim E, Chan A, Belmonte MF, Schroeder DF. Interactions between Arabidopsis DNA repair genes UVH6, DDB1A, and DDB2 during abiotic stress tolerance and floral development. Plant Sci. 2013;213:88–97. doi: 10.1016/j.plantsci.2013.09.004. [DOI] [PubMed] [Google Scholar]