

Abstract
Episodic memory relies on a distributed network of brain regions, with the hippocampus playing a critical and irreplaceable role. Few studies have examined how changes in this network contribute to episodic memory development early in life. The present addressed this gap by examining relations between hippocampal functional connectivity and episodic memory in 4- and 6-year-old children (n = 40). Results revealed similar hippocampal functional connectivity between age groups, which included lateral temporal regions, precuneus, and multiple parietal and prefrontal regions, and functional specialization along the longitudinal axis. Despite these similarities, developmental differences were also observed. Specifically, 3 (of 4) regions within the hippocampal memory network were positively associated with episodic memory in 6-year-old children, but negatively associated with episodic memory in 4-year-old children. In contrast, all 3 regions outside the hippocampal memory network were negatively associated with episodic memory in older children, but positively associated with episodic memory in younger children. These interactions are interpreted within an interactive specialization framework and suggest the hippocampus becomes functionally integrated with cortical regions that are part of the hippocampal memory network in adults and functionally segregated from regions unrelated to memory in adults, both of which are associated with age-related improvements in episodic memory ability.
Keywords: Hippocampus, Functional connectivity, Episodic memory, Early childhood, Interactive specialization
1. Introduction
The hippocampus is critical for episodic memory in adults (e.g., Aggleton and Brown, 2006, Cabeza and Nyberg, 2000, Davachi et al., 2003, Eichenbaum et al., 2007, Ranganath et al., 2004, Scoville and Milner, 1957, Yonelinas et al., 2005; see Spaniol et al., 2009 for review). Developmental changes in hippocampal structure and function have been shown to be related to improvement in episodic memory in school-aged children and adolescents (e.g., DeMaster et al., 2014, DeMaster and Ghetti, 2012, Ghetti et al., 2010, Ofen et al., 2007, Østby et al., 2012; see Ghetti and Bunge, 2012 for review). Overall, these studies suggest positive correlations exist between hippocampal volume and delayed recall and that changes in hippocampal function (as measured by task-based fMRI) contribute to age-related improvements in episodic memory. However, relatively little is known about how the hippocampus is related to memory development earlier in life. This gap is particularly unfortunate since behavioral studies have consistently identified early childhood (4–6 years) as a time of rapid improvement in episodic memory (e.g.,. Bauer et al., 2012; Drummey and Newcombe, 2002; Sluzenski et al., 2006, Riggins, 2014). Theoretical arguments have been made proposing how developmental changes in the hippocampus may be related to the development of episodic memory ability during early childhood (e.g., Jabès and Nelson, 2015, Lavenex and Lavenex, 2013, Riggins, 2012, Serres, 2001), however, this association has not been empirically examined in human children. This gap is likely due to the multiple challenges associated with acquiring functional neuroimaging measures in young children. To overcome these challenges and address this gap in the literature, the present research examined relations between resting-state functional connectivity and behavioral measures of episodic memory ability assessed outside the scanner in 4- and 6-year-old children.
1.1. Episodic memory development during early childhood
Behavioral studies examining development of episodic memory in children collectively suggest that early childhood is an important time of change. Specifically, several studies suggest that the ability to recall contextual details associated with events shows striking improvement between 4 and 6 years of age (e.g., Bauer et al., 2012, Drummey and Newcombe, 2002, Sluzenski et al., 2006, for review see Riggins, 2012). For instance, a recent longitudinal study examined rates of change in 4- to 10-year-old children's ability to recall both individual items (i.e., novel facts, such as bananas grow in bunches called hands) and contextual details associated with these items (i.e., from whom the fact was learned, a puppet or experimenter) across a 1-week delay (Riggins, 2014). Results revealed that whereas memory for items increased linearly between 4 and 10 years of age, memory for contextual details associated with these items showed accelerated change between 5 and 7 years of age.
Such empirical findings from controlled laboratory-based studies fit well with reports of children's ability to recall events from their own lives (i.e., autobiographical memories). For example, another longitudinal investigation examined 4- to 13-year-old children's ability to recall their earliest memories across a 2-year period (Peterson et al., 2011). Results indicated minimal consistency in the memories reported in 4- to 6-year-old children (either in the specific events recalled or the details of these events), but that consistency increased dramatically after this period. Based on these findings, the authors argued memories in early childhood are particularly fragile and especially prone to forgetting (see Bauer, 2015 for similar argument). However, the role of the hippocampus and associated networks in these changes remain relatively unstudied.
1.2. Hippocampal development during early childhood
Based on the known importance of the hippocampus for episodic memory in adults, it has been proposed that changes in hippocampal function during early childhood are related to observed improvements in episodic memory (Jabès and Nelson, 2015, Lavenex and Lavenex, 2013, Riggins, 2012, Serres, 2001). Neuroanatomical data from non-human primates suggest that developmental changes in synaptic connectivity within the hippocampus continue throughout early childhood (i.e., until 5–7 years of age, Jabès et al., 2010, Lavenex and Lavenex, 2013, Serres, 2001). Because circuitry in the dentate gyrus is critical for adult-like memory formation, its protracted developmental profile suggests that adult-like memory formation in humans may not be expected until the end of early childhood (Serres, 2001). Protracted development of the hippocampus may account for behavioral memory phenomena such as poor recall of contextual details in younger compared to older children and infantile or childhood amnesia (i.e., adults’ inability to recall autobiographical memories from very early in life).
To date, only one previous study has examined relations between hippocampal structure and episodic memory in early childhood (Riggins et al., 2015). In this study, associations between episodic memory and volume of subregions (head, body, tail) of the hippocampus were examined in 4- and 6-year-old children. Results revealed significant positive relations between episodic memory and volume of anterior regions of the hippocampus in both the left and right hemispheres for 6- but not 4-year-old children. These results suggested not only that relations between the hippocampus and episodic memory show significant developmental differences during early childhood, but they may be specific to certain subregions. These findings are consistent with results from fMRI studies in school-aged children (i.e., 8 years of age and older) that also suggest that changes in anterior hippocampal regions may be particularly relevant for episodic memory development (Ghetti et al., 2010; Maril et al., 2010; Paz-Alonso et al., 2008). However, relations between hippocampal function and memory behavior in early childhood (i.e., 3–6 years) were not addressed. Acquiring data addressing hippocampal function in children this young is challenging as typical task-based methods require children to remain motionless in the scanner while performing challenging memory tasks and likely accounts for why this question has not been explored previously.
1.3. Hippocampal functional connectivity at rest
One method that can be utilized to overcome the challenge of acquiring functional neuroimaging data from young children is task-free or resting-state functional connectivity MRI (rs-fcMRI). Previous research in adults has demonstrated that correlated patterns of intrinsic, spontaneous, low-frequency oscillations in brain activity can be detected in the absence of a specific task (Biswal et al., 1995, Fox and Raichle, 2007). These patterns, referred to as resting-state networks, are organized in functionally-relevant ways, as regions of these networks are typically co-activated during tasks designed to elicit distinct cognitive, social, or perceptual processes (Smith et al., 2009). The ability to identify brain networks independent of a task offers a significant advantage in the study of brain organization in development, particularly in young children, because the cognitive burden of performing a task while remaining motionless is eliminated (Casey et al., 2005, Uddin et al., 2010, for empirical examples see Emerson and Cantlon, 2012, Fareri et al., 2015, Gabard-Durnam et al., 2014). Moreover, use of rs-fcMRI has some particular advantages over traditional task-based approaches in that it (1) shifts focus to development of the entire network supporting memory and (2) results are independent of task-specific cognitive demands.
In adults, rs-fcMRI maps have been shown to reveal the full distribution of memory-related regions, as they coincide with regions showing activation across a variety of task-based memory studies (Vincent et al., 2006). Specifically, rs-fcMRI maps show robust correlations between BOLD signal in the hippocampus and several parietal regions (including precuneus, posterior cingulate, retrosplenial cortex, and bilateral inferior parietal lobule) as well as medial prefrontal regions. These regions are also often identified as the Default Mode Network (DMN), a network of regions that is preferentially active when individuals are not focused on tasks that demand external attention (e.g., when individuals are remembering past events, envisioning future events, or considering the perspectives of others, Buckner et al., 2008, Raichle et al., 2001). In addition, this network has been shown to be spatially distinct from other networks identified by different seed regions, such as the motion-sensitive MT complex (Vincent et al., 2006). Finally, specialization has also been shown along the longitudinal axis of the hippocampus, as functional connectivity maps differ between anterior and posterior hippocampal seed regions (Kahn et al., 2008, Poppenk and Moscovitch, 2011). Together, these studies report greater connectivity between the anterior hippocampus and lateral temporal regions (including the temporal poles), and greater connectivity between the posterior hippocampus and multiple regions including, frontal, parietal, posterior cingulate, and retrosplenial cortices. In addition, these rs-fcMRI studies suggest greater involvement of posterior regions of the hippocampus in episodic memory in adults, given its connectivity with multiple cortical areas known for their role in memory (e.g., frontal and parietal cortices).
Inter-subject variability in hippocampal network connectivity in adults has been shown to be related to variability in performance on episodic memory tasks. Specifically, connectivity between hippocampi during rest has been shown to predict adults’ performance during free recall (Wang et al., 2010a). Similarly, connectivity between the hippocampus and posterior cingulate and precuneus has been shown to predict adults’ performance on recollection tasks (whereas other cognitive tasks such as executive function or speed of processing did not; Wang et al., 2010b). Finally, these predictive associations have been shown to extend to atypical populations as well. For example, Roosendaal et al. (2010) demonstrated that differences in hippocampal functional connectivity arise even before memory is impaired at the behavioral level, suggesting they are causal, not consequential. Due to these associations with memory ability and the focus on memory in this paper, we will refer to this functional brain network as the “anterior/posterior hippocampal memory network;” however, we acknowledge that the hippocampus is involved in many behaviors in addition to memory (e.g., spatial navigation, emotion regulation, etc.).
Despite substantial reorganization of functional connectivity across development, several large-scale network properties appear to be preserved across the lifespan, suggesting that functional brain networks, even in children, are organized in manners similar to other complex systems (e.g., Power et al., 2010; Supekar et al., 2009). Previous research has established the presence and utility of resting-state functional connectivity early in development (e.g., Redcay et al., 2007, Uddin et al., 2010). In particular, findings from rs-fcMRI studies have added valuable knowledge regarding functional segregation and integration across short and long-range connections spanning the entire brain and have revealed important principles of functional brain development, including a shift from diffuse to focal activation patterns, and simultaneous pruning of local connectivity and strengthening of long-range connectivity with age (see Johnson, 2001, Supekar et al., 2009, Uddin et al., 2010; for theoretical argument; Fair et al., 2009 for empirical example in humans, and Webster et al., 1991 for empirical example in non-human primates). Although no developmental study has examined the relation between resting-state functional connectivity and episodic memory, this link has recently been established within other cognitive domains. For example, in school age-children, functional connectivity between distinct frontal and parietal regions has been shown to selectively predict math performance and connectivity between ventral-prefrontal cortex and fusiform gyrus has been shown to selectively predict performance on a face-matching task (Emerson and Cantlon, 2012).
1.4. Present study
The goal of the current study was to examine relations between development of hippocampal functional connectivity and episodic memory during early childhood. To achieve this goal, developmental differences in functional connectivity of the hippocampus during rest were examined in 4- and 6-year-old children and relations between connectivity and episodic memory ability were assessed. Based on previous literature in adults (e.g., Kahn et al., 2008, Poppenk and Moscovitch, 2011) and children (Riggins et al., 2015, see also DeMaster et al., 2014), suggesting functional specialization along the longitudinal axis of the hippocampus, both anterior and posterior hippocampal seeds were utilized. As reviewed above, although posterior regions appear to play a greater role in episodic memory in adults, there is evidence of anterior regions playing a more significant role in young children. Given that 4–6 years is a key transition period in episodic memory development, relations between both seeds as well as possible differential relations with age were investigated. Finally, given that development involves both progressive and regressive events (e.g., both segregation and integration of connections spanning the entire brain, e.g., Uddin et al., 2010) we probed whether any observed differences between the age groups fell “within” the anterior and posterior hippocampal memory networks or “outside” these networks.
2. Methods
This study was approved by the University's Institutional Review Board prior to data collection. Written informed consent was obtained from guardians of all participants. Children visited the laboratory for 3 different sessions as part of a larger study, which included behavioral assessments of memory, executive function, general cognitive abilities (Kaufman Brief Intelligence test, KBIT, (Kaufman and Kaufman, 1990)) and theory of mind, training in a mock scanner, and structural and functional MRI scans. Only data from the episodic memory assessment, KBIT, and MRI scans are included in this report (see Riggins et al., 2015 for relations between memory and hippocampal structure in children from this study).
2.1. Participants
A total of 75 4- (n = 36, 22 female, 14 male) and 6- (n = 39, 23 female, 16 male) year-old children participated in this study. All children were full-term, native English speakers, free of neurological damage, and had no history of developmental disorders, as determined by a parent-report questionnaire. A total of 52 children contributed memory data (29 4-year-old, 23 6-year-old children) and, of these, 40 also contributed useable fMRI data (21 4-year-old, 19 6-year-old children, see Table 1). Reasons for attrition are as follows: 4 children withdrew from the study, 3 were determined to be ineligible after enrollment (i.e., due to a history of seizures, behavioral problems, or limited knowledge of English), 4 did not successfully complete the MRI scan, 8 had excessive head motion during the MRI scan (i.e., >15% volumes were scrubbed), and 16 were not administered the memory assessment that is the focus of this paper.
Table 1.
Participant characteristics for children (n = 40) who contributed both fMRI and behavioral data.
| Characteristic | 4-year-olds (n = 21) | 6-year-olds (n = 19) | Group differences |
|---|---|---|---|
| Age (years) | 4.51 | 6.48 | t(38) = −20.88, p < .001 |
| Gender (male %) | 33.3 | 26.3 | χ2(1, 40) = 0.23, p = .63 |
| IQ (mean, SD) | 113 (11.40) | 116 (14.14) | t(38) = −.797, p = .43 |
| Motion | |||
| Scrubbed scans (mean, SD) | 5.05 (5.58) | 4.74 (6.44) | t(38) = .16, p = .87 |
| Mean FD (mean, SD) | 0.19 (0.09) | 0.20 (0.12) | t(38) = −.23, p = .82 |
2.2. Memory task
Episodic memory ability was examined using a source memory paradigm. In this paradigm, children were shown 36 items (commercially available toys) in one of two different locations (see Riggins and Rollins, 2015, Riggins et al., 2015 for similar paradigm). Locations were two rooms designed to be child-friendly and engaging (e.g., contained a stuffed “character” that resided in the room) and significant features of each room were made salient when the child entered the room (e.g., children unrolled the rug and placed it on the floor). In each room, children were first shown each item and allowed to interact with it in order to gain their attention and interest. Following this “baseline” assessment, the experimenter modeled one of three novel actions on each item (i.e., hug it, put it on your head, beat on it like a drum) and instructed the child to imitate the action. All items in location 1 were shown before the child moved to location 2. Each action was presented an equal number of times in each room. Location and item set order were counterbalanced between participants and items within sets were randomly presented, with the restriction that no more than two items with the same action were presented in a row.
After a delay of approximately 1 h, children were presented with the 36 target items seen during encoding and 18 novel items. Items were presented one at a time and the children were first asked to make old/new judgments. For items identified as “old”, children were also asked what action was paired with the item during encoding and which location they encountered it in. Experimenters recorded the action performed by the child and the location in which they placed the item during the session. Following the session, both action and location responses were assessed for accuracy. Consistent with the theoretical perspective that episodic memory consists of memory for both the item and the multiple types of details associated with it, items for which both the action and location details were correctly recalled served as the dependent measure of episodic memory. For items identified as “new” children were asked to place the item into a “new item” bin. In addition to age-appropriate instructions, five training trials were administered to ensure all children understood the task at encoding and retrieval. To ensure any observed effects were not the result of the items themselves, all 54 possible items were randomly divided into 6 sets of 9 items and then counterbalanced across conditions (i.e., location 1, location 2, and new).
Due to skewed distributions related to high performance on the recognition memory portion of the task, nonparametric Mann–Whitney U tests were used to examine differences in performance between 4- and 6-year-old children, on (1) recognition of old stimuli, (2) correct rejection of new stimuli, (3) d′ (sensitivity index). Between-sample t-tests were used to test group differences on the measure of episodic memory (i.e., recall of contextual details, which included both the location in which the old item was originally encountered and the action associated with it), which was more normally distributed (i.e., Kolmogorov–Smirnov p = 0.20).
The average delay between the episodic memory assessment and the MRI scan for the whole sample was 10 days (SD = 6 days, range = 1–25 days), which did not differ between the age groups, p = .22. (A similar pattern was observed for the subsample that contributed useable memory and fMRI data; n = 40, mean delay = 9 days, SD = 6 days, range = 1–25 days, p = .62.)
2.3. MRI data acquisition and preprocessing
All participants completed training in a mock scanner before MR data acquisition in order to become acclimated to the scanner environment and receive motion feedback. Participants were scanned in a Siemens 3.0-T scanner (MAGNETOM Trio Tim System, Siemens Medical Solutions, Erlangen, Germany) using a 12-channel coil. fMRI data were collected using an echo-planner imaging (EPI) sequence 180 EPI volumes consisting of 36 oblique interleaved slices with a 3.0 mm × 3.0 mm × 3.0 mm voxel size; 2 s TR; 24 ms TE; 3 mm slice thickness; 90° flip angle; 64 × 64 pixel matrix. The scan lasted for 6:06 min. Participants were asked to stay still and relax in the scanner while watching abstract objects (much like a screen saver) presented on a screen. During pilot testing, this manipulation proved to be effective in keeping young children still, yet remained devoid of any overt task instructions and yielded highly similar hippocampal maps compared to traditional fixation resting-state scans in a sample of young adults (see Supplemetary Material, Fig. S2; Emerson and Cantlon, 2012, Greicius et al., 2003, Vanderwal et al., 2015 for a similar approach). Structural data were collected using a high-resolution T1 magnetization-prepared rapid gradient-echo (MPRAGE) sequence consisting of 176 contiguous sagittal slices (1.0 mm × 1.0 mm × 1.0 mm voxel dimensions; 1900 ms TR; 2.52 ms TE; 900 ms inversion time; 9° flip angle; pixel matrix = 256 × 256). Functional and structural images were acquired in the same session.
Data were preprocessed using the Data Processing Assistant for Resting-state fMRI Advanced Edition (DPARSF-A, version 3.1) toolbox (Yan and Zang, 2010). The first 5 volumes of resting-state fMRI data were discarded from the analysis for each subject. The following steps were carried out in the preprocessing procedure: slice timing correction, head motion corrections, detrending, regression of nuisance variances, band-pass filtering (0.01–0.1 Hz), normalization, and smoothing (Gaussian kernel FWHM = 5 mm). The T1 image was segmented using the New Segment method implemented in SPM8. A child template was created by the Template-O-Matic toolbox (Wilke et al., 2008) and used in the New Segment procedure. DARTEL method was used to normalize the functional data from original space to MNI space. The nuisance variables included Friston24-motion parameters (Friston et al., 1996), the first five PCA components of white matter and CSF signal, and a binary dummy variable representing data points scrubbed based on head motion (Framewise Displacement (FD) > 0.7 mm, see Power et al., 2012). Specifically, we “scrubbed” our data using the cut option in the DPARSF-A toolbox, which excluded time points in which frame displacement was 0.7 mm or greater as well as the time point before and after these points. There was no relation between motion (i.e., mean FD) and performance on the memory task, p = 0.33, nor with age, p = .55, see Fig. 1. There were no differences in mean FD between the age groups (see Table 1, p = .82).
Fig. 1.

No significant relations were observed between motion (i.e., mean FD) and (A) performance on the episodic memory task (i.e., recall of contextual details), p = 0.33, nor (B) with age, p = .55.
2.4. Statistical analysis of fMRI data
In order to compute whole brain resting-state functional connectivity maps, two seed regions, bilateral anterior and bilateral posterior hippocampus, were generated by hand-tracing the hippocampus using anatomical landmarks on the left and right hemispheres of a pediatric template (Fonov et al., 2011) and resampling to functional resolution (3 mm3, see Fig. S1). The division between anterior and posterior hippocampus was determined based on the uncal apex, a standard and reliable anatomical landmark (Weiss et al., 2005). After preprocessing (see above), the average time courses from voxels within each seed region were extracted as reference signals (i.e., regressors). Correlation coefficients between the time courses from each seed region and time courses from a single voxel were calculated for every voxel in the brain. These correlation coefficients were then transformed with Fisher's r-to-z formula (z = 0.5*log((1 + r)/(1 − r))), yielding 2 z-maps (anterior and posterior) for second-level statistical analysis. One-sample t-tests were calculated on the z-map from each seed to detect anterior and posterior hippocampal memory networks in each age group using SPM. Additionally, in order to examine differences between the age groups and seed regions in functional connectivity, one group level linear mixed effects (LME) model was performed on the z-maps using 3dLME command within AFNI (Chen et al., 2013). The motion parameters, Age Group (4-year-old vs 6-year-old), Seed Location (anterior vs posterior) and Age Group-by-Seed Location interaction were included in the LME model on the z-maps. For the regions showing interactions, post hoc t-tests in SPSS were used to examine the directionality of these effects on the average coefficients extracted from these regions.
In order to examine relations between episodic memory performance and connectivity, a whole-brain multiple regression model in SPM was used to examine the relation between episodic memory scores (i.e., proportion of items for which both the action and location details were correctly recalled) and resting-state functional connectivity imputed using the two hippocampal seed regions (anterior and posterior, separately). This model included Age Group, Episodic Memory Performance, and the Age Group-by-Episodic Memory Performance interaction as independent variables while controlling for motion (mean FD) and IQ, in order to ensure any observed effects were not due to differences in motion or general intelligence. For regions showing interactions, the relation between memory performance and functional connectivity was examined in each group separately with and without continuous age as a covariate.
Finally, to determine if any observed interactions (without age as a covariate) were due to age-related differences in relations between episodic memory performance and connectivity “within” or “outside” the anterior and posterior hippocampal memory networks, results from the analyses described above (i.e., regions showing Age Group-by-Episodic Memory Performance interactions with mean FD and IQ as covariates) were then masked using the anterior or posterior hippocampal memory network maps as appropriate. Regions that remained after the masks were applied and the peak activation of the cluster fell clearly within the mask are referred to as falling “within” the anterior/posterior hippocampal memory network. Regions that were no longer apparent after the masks were applied are referred to as regions “outside” the anterior/posterior hippocampal memory networks.
Monte Carlo simulations were carried out using 3dClustSim in AFNI to determine the minimum cluster size and threshold in order to maintain an overall alpha at p < .05. Based on the simulation results, only clusters with a minimum of 79 voxels size and puncorrected < .01 were viewed as significant with multiple comparison correction (pcorrected < 0.05). All figures used the 4.5–8.5 year MNI Child Template (from Fonov et al., 2011) as the underlay.
3. Results
3.1. Memory performance
Results revealed that there was no difference between 4- and 6-year-old children's ability to recognize stimuli as old. However, consistent with previous literature (e.g., Bauer et al., 2012; Drummey and Newcombe, 2002; Riggins and Rollins, 2015, Riggins et al., 2015, Sluzenski et al., 2006), 4-year-old children were marginally less able to correctly reject new stimuli and recalled fewer contextual details1 compared to 6-year-old children (see Table 2).
Table 2.
Average behavioral performance for all 4- and 6-year-old children who contributed behavioral data for the episodic memory task (n = 52).
| 4 year olds (n = 29) |
6 year olds (n = 23) |
Group difference | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| Hits | 96.54% | 9.07 | 99.88% | 0.58 | p = .23 |
| Correct rejections | 92.16% | 21.44 | 98.79% | 5.79 | p = .09 |
| d′ | 3.98 | 1.53 | 4.56 | 0.37 | p = 0.08 |
| Contextual details | 42.45% | 12.51 | 49.55% | 15.11 | t(1, 50) = 1.86, p = .07 |
3.2. Whole brain connectivity
Whole brain connectivity maps for each age group (4 and 6 years) and seed region (anterior, posterior) are illustrated in Fig. 2 and listed in Table 3. These maps illustrate significant positive functional connectivity between the anterior hippocampus and multiple regions of the brain, including lateral temporal cortex and temporal poles, which have been shown in previous research in older individuals (Kahn et al., 2008, Poppenk and Moscovitch, 2011). For the posterior hippocampus, significant positive functional connectivity was also observed with a widespread set of brain regions, including posterior cingulate, precuneus, and multiple posterior parietal and prefrontal regions (Kahn et al., 2008, Poppenk and Moscovitch, 2011). Given the focus on memory in this investigation, in the following sections, these maps are referred to as the “anterior/posterior hippocampal memory network” maps.
Fig. 2.
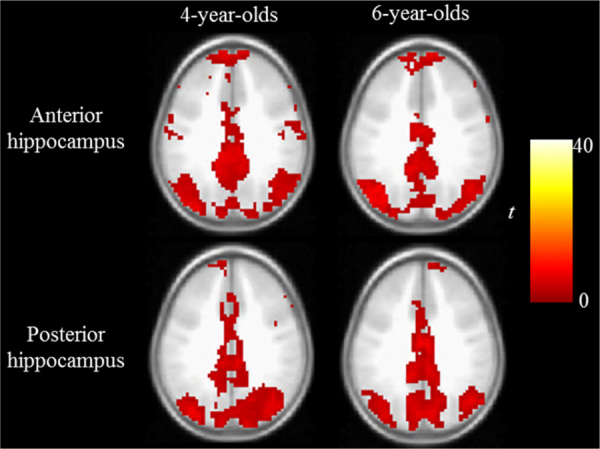
Whole brain functional connectivity maps with bilateral hippocampal seeds for each age group (4- or 6-year-old) and seed location (anterior or posterior, p < .05 corrected, z = 33). Analyses indicated no significant differences or interactions between the age groups, however, multiple regions show differences between seed region location (see text and Table 3). These maps are referred to in the text as the anterior hippocampal memory network and posterior hippocampal network map, respectively.
Table 3.
Peak coordinates (MNI) of regions showing significant functional connectivity for both anterior and posterior seed locations (p < .05 corrected, z = 33).
| Structure | k | x | y | z | t |
|---|---|---|---|---|---|
| Anterior hippocampus | |||||
| Left hippocampus | 36,181 | −21 | −15 | −18 | 39.79 |
| Right hippocampus | |||||
| Calcarine/precuneus | |||||
| Medial frontal gyrus | |||||
| Midbrain | |||||
| Ventral striatum | |||||
| Right middle temporal gyrus | |||||
| Bilateral fusiform | |||||
| Anterior and posterior cingulate cortex | |||||
| Inferior frontal gyrus | |||||
| Thalamus | |||||
| Amygdala | |||||
| Left middle temporal gyrus | |||||
| Cerebellum posterior lobe | |||||
| Posterior hippocampus | |||||
| Right parahippocampal gyrus | 33,176 | 27 | −24 | −15 | 35.73 |
| Left parahippocampal gyrus | |||||
| Hippocampus | |||||
| Thalamus | |||||
| Lingual/calcarine/precuneus | |||||
| Anterior and posterior cingulate | |||||
| Orbital medial frontal gyrus | |||||
| Ventral striatum | |||||
| Fusiform | |||||
| Amygdala | |||||
| Cuneus | |||||
| Midbrain | |||||
The LME model revealed main effects of seed location in 12 regions between the anterior and posterior hippocampal connectivity maps. Eight of these regions showed greater connectivity with anterior than posterior hippocampus, whereas the other 4 regions showed the opposite pattern, see Table 4. However, this model did not reveal any significant differences between the age groups or significant interactions between age group and seed region.
Table 4.
Regions from LME analyses showing significant differential connectivity between anterior vs posterior hippocampal seed regions.
| k | MNI coordinates |
Peak F value | |||
|---|---|---|---|---|---|
| x | y | z | |||
| Regions showing greater connectivity with anterior vs posterior hippocampus | |||||
| Right postcentral gyrus | 2856 | −30 | 27 | 66 | 50.12 |
| Left superior frontal gyrus | |||||
| Bilateral precentral gyrus | |||||
| Bilateral SMA | |||||
| Bilateral paracentral lobule | |||||
| Right anterior parahippocampal gyrus | 1672 | −24 | 18 | −27 | 100 |
| Right uncus | |||||
| Left middle temporal gyrus | 397 | 66 | 33 | −9 | 29.42 |
| Left precuneus | 258 | 0 | 54 | 9 | 25.86 |
| Left cingulate gyrus | |||||
| Left posterior cingulate | |||||
| Right anterior cingulate cortex | 152 | −6 | −27 | 21 | 32.23 |
| Left angular gyrus | 145 | 45 | 60 | 30 | 16.75 |
| Left postcentral gyrus | 90 | 54 | 12 | 42 | 18.43 |
| Right inferior frontal gyrus | 84 | −57 | −33 | −6 | 52.65 |
| Regions showing greater connectivity with posterior vs anterior hippocampus | |||||
| Right posterior parahippocampus gyrus | 1702 | −33 | 39 | −12 | 100 |
| Right caudate | |||||
| Right middle occipital gyrus | |||||
| Left posterior parahippocampal gyrus | 715 | 30 | 39 | −9 | 100 |
| Left fusiform gyrus | |||||
| Left caudate | |||||
| Right angular gyrus | 107 | −54 | 63 | 27 | 21.92 |
| Right putamen | 98 | −27 | −24 | 3 | 20.24 |
3.3. Relations between whole-brain connectivity and episodic memory performance
Given previous literature and significant differences in connectivity between anterior and posterior seeds, relations between these seed locations and episodic memory performance (i.e., proportion of items for which both the action and location details were correctly recalled) were examined separately. Including continuous age as a covariate in the LME analyses did not significantly change any results, thus only the results without continuous age as a covariate are reported.
For the anterior hippocampal seed, episodic memory was found to be negatively related to connectivity in both 4- and 6-year-old children in two regions, including precuneus and ventral medial prefrontal cortex (VMPFC) (see Fig. S3A). These effects were qualified by significant interactions between age group (4 vs 6) and episodic memory, which were observed in 5 regions. As described above, after identifying these regions, we then applied masks generated from the anterior hippocampal connectivity map in order to determine if they were “within” or “outside” the anterior hippocampal memory network. Results showed that 2 regions showing significant interactions fell ‘within’ the anterior hippocampal memory network map, namely the precuneus and right superior frontal gyrus (SFG), see Table 5/Fig. 3A. In both these regions, episodic memory was positively related to functional connectivity in 6-year-olds, but negatively related in 4-year-olds (Fig. 3A). Specifically, increased connectivity from anterior hippocampus to right SFG and precuneus was associated with better episodic memory performance in 6-year-old children, whereas in 4-year-old children increased connectivity was related to worse episodic memory performance. The remaining 3 regions fell ‘outside’ the anterior hippocampus memory network, which included left middle temporal gyrus (MTG), right MTG, and right inferior frontal gyrus (IFG), see Table 5/Fig. 4. These regions showed the opposite pattern compared to regions within the anterior hippocampal memory network, such that episodic memory was negatively related to functional connectivity in 6-year olds, but positively related in 4-year olds (Fig. 4). Specifically, increased connectivity from anterior hippocampus to left and right MTG and right IFG were associated with worse episodic memory performance in older children, but with better episodic memory performance in younger children.
Table 5.
Regions showing different relations between functional connectivity and episodic memory performance (i.e., recall of contextual details) between 4- and 6-year-old children for anterior and posterior hippocampal seed regions.
| Seed | Region(s) | Hemisphere | k | MNI coordinates |
Peak t value | Interactiona | ||
|---|---|---|---|---|---|---|---|---|
| x | y | z | ||||||
| Regions “within” the hippocampal memory networks | ||||||||
| Anterior hippocampal network | Precuneus | Left/Right | 170 | 3 | −63 | 18 | 3.84 | 6+/4− |
| Superior frontal gyrus | Right | 81 | 21 | 27 | 57 | 4.11 | 6+/4− | |
| Posterior hippocampal network | Superior temporal gyrus | Right | 95 | 45 | 6 | −21 | 4.48 | 6+/4− |
| Middle temporal gyrus | Right | 193 | 57 | −48 | −12 | −4.66 | 6−/4+ | |
| Regions “outside” the hippocampal memory networks | ||||||||
| Anterior hippocampal network | Middle temporal gyrus | Left | 100 | −57 | −63 | 0 | −4.43 | 6−/4+ |
| Middle temporal gyrus | Right | 188 | 60 | −51 | −12 | −3.75 | 6−/4+ | |
| Inferior frontal gyrus | Right | 102 | 57 | 21 | 27 | −4.21 | 6−/4+ | |
6+ represents positive correlation in 6-year-old group; 6− represents negative correlation in 6-year-old group; 4+ represents positive correlation in 4-year-old group; 4− represents negative correlation in 4-year-old group.
Fig. 3.

Regions “within” the hippocampal memory networks showing significant interactions between age group and episodic memory performance in predicting whole brain connectivity when the (A) anterior and (B) posterior hippocampus seeds were used. SFG: superior frontal gyrus. MTG: middle temporal gyrus; IFG: inferior frontal gyrus. (A) Anterior hippocampal memory network. (B) Posterior hippocampal memory network.
Fig. 4.

Regions “outside” the hippocampal memory networks showing significant interactions between age group and episodic memory performance in predicting whole brain connectivity when the anterior hippocampus seed was used. Note: No significant findings were observed for the posterior hippocampal seed. STG: superior temporal gyrus. MTG: middle temporal gyrus. Anterior hippocampal memory network.
For the posterior hippocampus seed, episodic memory in both 4- and 6-year-old children was found to be positively related to connectivity in right middle temporal gyrus (MTG) and negatively related to connectivity in VMPFC (see Fig. S3B). This effect was qualified by significant interactions between age group and episodic memory in 2 regions. As with the anterior hippocampal seed, we then applied a mask generated from the posterior hippocampal connectivity map to these results in order to determine if these regions were “within” or “outside” the posterior hippocampal memory network. Significant interactions between age group and memory for contextual details were observed in 2 regions ‘within’ the posterior hippocampal memory network map, including right superior temporal gyrus (STG) and right MTG, see Table 5/Fig. 3B. Similar to the two regions within the anterior hippocampal memory network, increased connectivity from posterior hippocampus to right STG was associated with better episodic memory performance in 6-year olds but worse episodic memory performance in 4-year olds. In contrast, connectivity from posterior hippocampus to right MTG was associated with worse episodic memory performance in older children, but better episodic memory performance in younger children. No significant interactions were observed ‘outside’ the posterior hippocampal memory network map.
4. Discussion
The current study sought to elucidate relations between functional development of the hippocampus and episodic memory during early childhood by examining developmental differences in correlations between functional connectivity of the hippocampus during rest and episodic memory ability outside of the scanner. First, results revealed that hippocampal resting-state functional connectivity (rs-fcMRI) was similar in 4- and 6-year-old children. Specifically, consistent with findings in adults, functional connectivity was observed between the hippocampus and many regions of the brain, including precuneus, posterior cingulate, retrosplenial cortex, bilateral inferior parietal regions, and medial prefrontal regions (see Vincent et al., 2006), and this connectivity showed specialization along the longitudinal axis of the hippocampus as anterior and posterior regions showed significant differences in connectivity (Kahn et al., 2008, Poppenk and Moscovitch, 2011). However, despite these similarities in overall hippocampal functional connectivity, developmental differences between 4- and 6-year-old children were observed in relations between connectivity and episodic memory ability in 7 different regions. Four of these regions fell “within” the hippocampal memory network and 3 fell “outside” the hippocampal memory network, suggesting both functional integration and segregation may be occurring during this period. Overall, these findings are consistent with an interactive specialization framework, which suggests that functionally relevant brain development during early childhood is not the result of the maturation of brain regions, but rather the organization (or re-organization) of patterns of connectivity between regions (Johnson, 2001).
Specifically, within the anterior/posterior hippocampal memory networks, 3 of the 4 regions identified showed positive associations between functional connectivity and episodic memory ability in 6-year-old children. That is, increased connectivity between the anterior hippocampus and both the precuneus and right SFG and increased connectivity between the posterior hippocampus and STG was related to better memory performance. This pattern is similar to previous findings in adults (Wang et al., 2010a, Wang et al., 2010b), which show that greater functional connectivity between the hippocampus and multiple regions within the hippocampal memory network (including the precuneus) is related to better performance on memory tasks. It is also consistent with research in adults suggesting that, during rest, individuals engage many of these regions (i.e., posterior cingulate, medial PFC, bilateral parietal cortex) during rest to, among other things, recall or reminisce about past experiences (Buckner et al., 2008). Finally, it is consistent with the previous structural MRI study in early childhood, which showed relations between volume of anterior hippocampus and episodic memory in 6-year-old children (Riggins et al., 2015).
In contrast, in 4-year-old children, connectivity between these regions within the anterior/posterior hippocampal memory networks were negatively related to memory performance. The reason for this developmental difference is not clear. However it may be that earlier reliance on a more “mature” network may not yet be advantageous for behavioral performance. We return to this point below. But first, it is worth discussing 4-year-old children's performance on the task. Although these younger children recalled fewer details on the episodic memory task compared to 6-year-old children, their performance was still above chance levels. This finding is consistent with previous behavioral research suggesting that 4-year-old children can remember certain aspects of events, but that these details are recalled much less reliably than in older children (Bauer et al., 2012; Drummey and Newcombe, 2002; Riggins and Rollins, 2015, Riggins et al., 2015, Sluzenski et al., 2006). As such, a commonly asked question is, how are 4-year olds accomplishing this task, and is it in a manner that differs from that in older children? One possibility is that 4-year-olds are engaging a different network(s) or set of regions to accomplish the same behavioral task. Such a dissociation has been shown previously in memory research in nonhuman primates. For example, focal lesions to the hippocampus impair recognition memory in adults but do not impact performance in infant monkeys (cf. Zola et al., 2000, Zeamer et al., 2010) suggesting use of different regions at different developmental periods. To explore this possibility we examined whether younger children were utilizing regions “outside” the anterior/posterior hippocampal memory networks (Johnson, 2001, Uddin et al., 2010, Webster et al., 1991). Results revealed 3 regions that showed positive associations between anterior hippocampal functional connectivity and episodic memory performance in 4-year-old children, namely bilateral MTG and IFG. Thus, data from the present study suggest it may be that younger children were relying more on regions “outside” the memory network to complete the episodic memory task. This finding is consistent with research in children in other cognitive domains, which suggest that younger individuals recruit a more distributed set of regions early in development and that more focal activation patterns or localized functions emerge over time (e.g., Durston et al., 2006). It is possible that, as children develop, reliance on regions “outside” the mature hippocampal memory network becomes detrimental to memory performance.
If it is indeed the case that young children rely on a different set or subset of brain regions to perform episodic memory tasks then it is possible that negative correlations between connectivity and memory performance in the 4-year-old age group arise because some of these children are transitioning from reliance on these regions to reliance on the more mature/canonical memory networks. This transitional phase may result in an initial decrement in performance that is subsequently followed by an increase (see Siegler, 1996 for similar description of change in the behavioral domain). An analogy that might make this more clear comes from behavioral learning of strategies, such as learning how to type. One can learn to be a fairly fast typist by using the “hunt and peck” index fingers only strategy. However, when one then learns the proper technique (i.e., using all fingers, home row, etc.) they experience an initial decrement in performance (i.e., slower typing speed) before experiencing an increase in typing speed that, eventually, exceeds the speed of the old, less mature method. A similar situation could be occurring at the neural level if 4-year-olds are transitioning from reliance on a distributed set of unspecialized regions to the more “mature” hippocampal memory network. There is some evidence to support this notion, namely that, with age, children come to rely more heavily on strategic cognitive control processes and prefrontal regions during both the encoding and retrieval of memories (see Shing and Lindenberger, 2011 for elaboration).
Although the “within” vs “outside” network approach is useful in showing how the field can move beyond strictly maturational arguments, this dichotomy is likely too simplistic to capture the complex interactions taking place in the developing brain (e.g., Pfeifer and Allen, 2012). This may be why one region within the posterior hippocampal memory network in the present study (i.e., right MTG), showed the opposite pattern than the other 3 regions, namely positive relations between connectivity and episodic memory performance in 4-year-old children and negative relations in 6-year-old children. Thus, future research should strive to build on the present work yet move beyond this dichotomized characterization and examine functional organization in increasingly complex ways (e.g., Supekar et al., 2009). However, the results presented here are an essential “first step” in this process and will be useful for identifying regions of interest that can be used in future studies.
The present findings are consistent with previous research in both developmental psychology and cognitive neuroscience. First, in terms of episodic memory performance, 4-year-old children were less likely than 6 year olds to recall contextual details associated with items, which is similar to previous research (Bauer et al., 2012; Drummey and Newcombe, 2002; Riggins and Rollins, 2015, Riggins et al., 2015, Sluzenski et al., 2006). However, it should be noted that the magnitude of this effect was smaller in the present study than in previous reports. This is likely due to the fact that the delay between encoding and retrieval was 1-h, compared to longer delays used in previous work (e.g., 1–2 days or 1-week). It is unclear from the present study if or how developmental differences in relations with functional connectivity would be affected if longer delays had been imposed. Second, in terms of whole brain measures of hippocampal functional connectivity, our findings are consistent with previous research in adults. Specifically, as with Vincent et al., 2006, the hippocampus showed significant connectivity with multiple, widespread regions, including the precuneus, parietal, and prefrontal cortical regions (cf. Fig. 2 in the present paper and Fig. 2 in Vincent et al., 2006). We also observed significant differences between connectivity in anterior vs posterior hippocampal seeds, suggesting specialization along the longitudinal axis of the hippocampus is present during childhood. Similar to research in adults, the anterior hippocampus showed greater connectivity with temporal regions and the posterior hippocampus showed greater connectivity with occipital regions (Kahn et al., 2008, Poppenk and Moscovitch, 2011). However, contrary to findings in adults, anterior hippocampus also showed greater connectivity with precuneus, anterior and posterior cingulate, and frontal cortex (Kahn et al., 2008, Poppenk and Moscovitch, 2011). Thus, although functional specialization was present, the specifics differed from adults. These differences are likely due to prolonged developmental changes that occur beyond 6 years of age (see Blankenship et al., 2015, Ghetti and Bunge, 2012).
The present study has several notable strengths, including focus on a young age range during which a known developmental transition occurs in memory ability (i.e., 4–6 years). To achieve this, we related resting-state functional connectivity measures to performance on an episodic memory task outside the scanner. This approach allowed for the examination of relations between hippocampal function and behavior in young children. In addition, great attention was paid to quality control in terms of acquiring data with very minimal artifact from both age groups. This was also coupled with adhering to current recommendations for analysis, including those designed to address the influence of motion (e.g., scrubbing, Power et al., 2012), 24 motion parameters (Friston et al., 1996) and multiple nuisance regressors (components from CSF and WM masks) to account for physiological noise (see Behzadi et al., 2007). Our analyses revealed there were no significant differences in motion between age groups and that motion was not correlated with either performance on the memory task nor age. Therefore, observed effects are likely not due to these nuisance variables.
Despite these strengths, there are also some limitations that should be noted. First, the present analyses only included a subset of children tested. Thus, results are conservative and can only be generalized to children similar to our sample who are able to undergo and complete fMRI testing. Second, given the sample size and distribution of gender within the age groups, differences between genders could not be addressed. This is an important direction for future research as previous research suggests a gender effect on hippocampal volume in children and adolescents (e.g., Gogtay et al., 2006). Third, although use of rs-fcMRI allowed for the examination of hippocampal function in young children, these findings would be strengthened if they were replicated using a traditional task-based fMRI paradigm in conjunction with resting-state analyses. Finally, longitudinal studies would be necessary to confirm that these differential relations between episodic memory performance and hippocampal connectivity between 4 and 6 years of age represent a developmental shift. Although challenging, efforts to collect task-based and longitudinal fMRI data from children as young as 4 years of age are underway in our lab.
This study is the first, to our knowledge, to address relations between functional development of the hippocampus and episodic memory during early childhood. Taken together, the present findings are consistent with an interactive specialization framework, which proposes that brain development is not a unidirectional maturational process, but rather results from complex and dynamic interactions between brain regions (Johnson, 2001). Specifically, our results suggest that during early childhood the hippocampus becomes progressively (1) integrated with cortical regions that are part of the hippocampal memory network in adults and (2) segregated from regions unrelated to memory in adults, both of which result in age-related improvements in episodic memory ability. This pattern has also been observed in empirical studies examining anatomical connections between MTL and ITG early in life in macaques that then disappear by adulthood, which suggests that “extra” connections may be useful early in life but not later (e.g., Webster et al., 1991). This pattern is also supported by findings from other rs-fcMRI studies in developmental samples (see Uddin et al., 2010 for review). These results provide (1) initial insight into neuroanatomical correlates of developmental change in behavior during a relatively unexplored, yet critical, age window, (2) possible mechanisms underlying variability in episodic memory ability during early childhood, and (3) typical development of the structural and functional architecture of the brain network supporting memory. Data that speak to these issues are important as they not only increase our understanding of the development of brain networks supporting behavior, but also have strong relevance to public health as they can potentially be used to develop interventions targeting memory early in life when impact may be largest (Nelson, 2000, Ramey and Ramey, 1998). Finally, results from the present investigation fill a significant gap in the literature between what is known regarding memory processes early vs later in life, which is essential for a complete understanding of memory ability to be achieved.
Footnotes
This research was supported by funding from the National Institutes of Health (HD079518), a Maryland Neuroimaging Center seed grant, the Behavioral and Social Sciences Dean's Research Initiative, and the Department of Psychology at the University of Maryland, College Park. The authors would like to thank the members of the Neurocognitive Development Lab and Developmental Social Cognitive Neuroscience Lab at the University of Maryland, especially, Daniel O’Young, Brieana Viscomi, Katherine Rice, and Lauren Weiss, for their contribution to this work, Jason Smith for assistance with data analysis, and the families who participated in this study. The authors also thank the Maryland Neuroimaging Center for their support in data acquisition. Portions of these data were presented at the Cognitive Neuroscience Society meeting in San Francisco, CA, May 2013 and the Society for Research in Child Development meeting in Seattle, WA, May 2013.
The pattern of findings was similar in the subset of children who contributed useable fMRI data (n = 40). However, due to decreases in power, differences were no longer marginal. However, the means were similar (e.g., memory for contextual details in 4-year-olds: M = 42.74%, SD = 11.87 and 6-year-olds: M = 47.52%, SD = 13.67, p = .24).
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.dcn.2016.02.002.
Appendix A. Supplementary data
The following are the supplementary data to this article:
References
- Aggleton J.P., Brown M.W. Interleaving brain systems for episodic and recognition memory. Trends Cogn. Sci. 2006;10(10):455–463. doi: 10.1016/j.tics.2006.08.003. [DOI] [PubMed] [Google Scholar]
- Bauer P.J. A complementary processes account of the development of childhood amnesia and a personal past. Psychol. Rev. 2015;2:204–231. doi: 10.1037/a0038939. [DOI] [PubMed] [Google Scholar]
- Bauer P.J., Doydum A.O., Pathman T., Larkina M., Güler O.E., Burch M. It's all about location, location, location: children's memory for the “where” of personally experienced events. J. Exp. Child Psychol. 2012;113(4):510–522. doi: 10.1016/j.jecp.2012.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behzadi Y., Restom K., Liau J., Liu T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage. 2007;37(1):90–101. doi: 10.1016/j.neuroimage.2007.04.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blankenship S.L., Dougherty L.R., Redcay E., Riggins T. Development of hippocampal resting-state networks during childhood. Talk presented at the biennial meeting of the Society for Research on Child Development; Philadelphia, PA; 2015. [Google Scholar]
- Biswal B., Zerrin Yetkin F., Haughton V.M., Hyde J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn. Reson. Med. 1995;34(4):537–541. doi: 10.1002/mrm.1910340409. [DOI] [PubMed] [Google Scholar]
- Buckner R.L., Andrews-Hanna J.R., Schacter D.L. The brain's default network. Ann. N.Y. Acad. Sci. 2008;1124(1):1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- Cabeza R., Nyberg L. Imaging cognition II: an empirical review of 275 PET and fMRI studies. J. Cogn. Neurosci. 2000;12(1):1–47. doi: 10.1162/08989290051137585. [DOI] [PubMed] [Google Scholar]
- Casey B., Galvan A., Hare T.A. Changes in cerebral functional organization during cognitive development. Curr. Opin. Neurobiol. 2005;15(2):239–244. doi: 10.1016/j.conb.2005.03.012. [DOI] [PubMed] [Google Scholar]
- Chen G., Saad Z.S., Britton J.C., Pine D.S., Cox R.W. Linear mixed-effects modeling approach to FMRI group analysis. Neuroimage. 2013;73:176–190. doi: 10.1016/j.neuroimage.2013.01.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davachi L., Mitchell J.P., Wagner A.D. Multiple routes to memory: distinct medial temporal lobe processes build item and source memories. Proc. Natl. Acad. Sci. U. S. A. 2003;100(4):2157–2162. doi: 10.1073/pnas.0337195100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeMaster D., Pathman T., Lee J.K., Ghetti S. Structural development of the hippocampus and episodic memory: developmental differences along the anterior/posterior axis. Cereb. Cortex. 2014;24:3036–3045. doi: 10.1093/cercor/bht160. [DOI] [PubMed] [Google Scholar]
- DeMaster D., Ghetti S. Developmental differences in hippocampal and cortical contributions to episodic retrieval. Cortex. 2012;49(6):1482–1493. doi: 10.1016/j.cortex.2012.08.004. [DOI] [PubMed] [Google Scholar]
- Drummey A.B., Newcombe N.S. Developmental changes in source memory. Dev. Sci. 2002;5(4):502–513. [Google Scholar]
- Durston S., Davidson M.C., Tottenham N., Galvan A., Spicer J., Fossella J.A., Casey B.J. A shift from diffuse to focal cortical activity with development. Dev. Sci. 2006;9(1):1–8. doi: 10.1111/j.1467-7687.2005.00454.x. [DOI] [PubMed] [Google Scholar]
- Eichenbaum H., Yonelinas A.P., Ranganath C. The medial temporal lobe and recognition memory. Annu. Rev. Neurosci. 2007;30(1):123–152. doi: 10.1146/annurev.neuro.30.051606.094328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emerson R.W., Cantlon J.F. Early math achievement and functional connectivity in the fronto-parietal network. Dev. Cogn. Neurosci. 2012;2(Suppl. 1):S139–S151. doi: 10.1016/j.dcn.2011.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fair D.A., Cohen A.L., Power J.D., Dosenbach N.U.F., Church J.A., Miezin F.M.…Petersen S.E. Functional brain networks develop from a “local to distributed” organization. PLoS Comput. Biol. 2009;5(5):e1000381. doi: 10.1371/journal.pcbi.1000381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fareri D.S., Gabard-Durnam L., Goff B., Flannery J., Gee D.G., Lumian D.S., Caldera C., Tottenham N. Normative development of ventral striatal resting-state connectivity in humans. Neuroimage. 2015;118:422–437. doi: 10.1016/j.neuroimage.2015.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonov V., Evans A.C., Botteron K., Almli C.R., McKinstry R.C., Collins D.L. Unbiased average age-appropriate atlases for pediatric studies. Neuroimage. 2011;54(1):313–327. doi: 10.1016/j.neuroimage.2010.07.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox M.D., Raichle M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007;8(9):700–711. doi: 10.1038/nrn2201. [DOI] [PubMed] [Google Scholar]
- Friston K.J., Williams S., Howard R., Frackowiak R.S.J., Turner R. Movement-related effects in fMRI time-series. Magn. Reson. Med. 1996;35(3):346–355. doi: 10.1002/mrm.1910350312. [DOI] [PubMed] [Google Scholar]
- Gabard-Durnam L.J., Flannery J., Goff B., Gee D.G., Humphreys K.L., Telzer E.…Tottenham N. The development of human amygdala functional connectivity at rest from 4 to 23 years: a cross-sectional study. Neuroimage. 2014;95:193–207. doi: 10.1016/j.neuroimage.2014.03.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius M.D., Krasnow B., Reiss A.L., Menon V. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. U. S. A. 2003;100(1):253–258. doi: 10.1073/pnas.0135058100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghetti S., Bunge S.A. Neural changes underlying the development of episodic memory during middle childhood. Dev. Cogn. Neurosci. 2012;2(4):381–395. doi: 10.1016/j.dcn.2012.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghetti S., DeMaster D.M., Yonelinas A.P., Bunge S.A. Developmental differences in medial temporal lobe function during memory encoding. J. Neurosci. 2010;30(28):9548–9556. doi: 10.1523/JNEUROSCI.3500-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gogtay N., Nugent T.F., Herman D.H., Ordonez A., Greenstein D., Hayashi K.M.…Thompson P.M. Dynamic mapping of normal human hippocampal development. Hippocampus. 2006;16(8):664–672. doi: 10.1002/hipo.20193. [DOI] [PubMed] [Google Scholar]
- Jabès A., Nelson C.A. 20 years after “The ontogeny of human memory A cognitive neuroscience perspective”. Where are we? Reply to commentaries. Int. J. Behav. Dev. 2015 [Google Scholar]
- Jabès A., Lavenex P.B., Amaral D.G., Lavenex P. Quantitative analysis of postnatal neurogenesis and neuron number in the macaque monkey dentate gyrus. Eur. J. Neurosci. 2010;31:273–285. doi: 10.1111/j.1460-9568.2009.07061.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson M.H. Functional brain development in humans. Nat. Rev. Neurosci. 2001;2:475–483. doi: 10.1038/35081509. [DOI] [PubMed] [Google Scholar]
- Kaufman A.S., Kaufman N.L. Wiley Online Library; 1990. Kaufman Brief Intelligence Test. [Google Scholar]
- Kahn I., Andrews-Hanna J.R., Vincent J.L., Snyder A.Z., Buckner R.L. Distinct cortical anatomy linked to subregions of the medial temporal lobe revealed by intrinsic functional connectivity. J. Neurophysiol. 2008;100(1):129–139. doi: 10.1152/jn.00077.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavenex P., Lavenex P.B. Building hippocampal circuits to learn and remember: insights into the development of human memory. Behav. Brain Res. 2013 doi: 10.1016/j.bbr.2013.02.007. [DOI] [PubMed] [Google Scholar]
- Maril A., Davis P.E., Koo J.J., Reggev N., Zuckerman M., Ehrenfeld L., Rivkin M.J. Developmental fMRI study of episodic verbal memory encoding in children. Neurology. 2010;75:2110–2116. doi: 10.1212/WNL.0b013e318201526e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson C.A. Neural plasticity and human development: the role of early experience in sculpting memory systems. Dev. Sci. 2000;3(2):115–136. [Google Scholar]
- Ofen N., Kao Y.-C., Sokol-Hessner P., Kim H., Whitfield-Gabrieli S., Gabrieli J.D.E. Development of the declarative memory system in the human brain. Nat. Neurosci. 2007;10(9):1198–1205. doi: 10.1038/nn1950. [DOI] [PubMed] [Google Scholar]
- Østby Y., Tamnes C.K., Fjell A.M., Walhovd K.B. Dissociating memory processes in the developing brain: the role of hippocampal volume and cortical thickness in recall after minutes versus days. Cereb. Cortex. 2012;22(2):381–390. doi: 10.1093/cercor/bhr116. [DOI] [PubMed] [Google Scholar]
- Paz-Alonso P.M., Ghetti S., Donohue S.E., Goodman G.S., Bunge S.A. Neurodevelopmental correlates of true and false recognition. Cereb. Cortex. 2008;18:2208–2216. doi: 10.1093/cercor/bhm246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson C., Warren K.L., Short M.M. Infantile amnesia across the years: a 2-year follow-up of children's earliest memories. Child Dev. 2011;82(4):1092–1105. doi: 10.1111/j.1467-8624.2011.01597.x. [DOI] [PubMed] [Google Scholar]
- Pfeifer J.H., Allen N.B. Arrested development? Reconsidering dual-systems models of brain function in adolescence and disorders. Trends Cogn. Sci. 2012;16(6):322–329. doi: 10.1016/j.tics.2012.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poppenk J., Moscovitch M. A hippocampal marker of recollection memory ability among healthy young adults: contributions of posterior and anterior segments. Neuron. 2011;72(6):931–937. doi: 10.1016/j.neuron.2011.10.014. [DOI] [PubMed] [Google Scholar]
- Power J.D., Fair D.A., Schlaggar B.L., Petersen S.E. The development of human functional brain networks. Neuron. 2010;67(5):735–748. doi: 10.1016/j.neuron.2010.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Power J.D., Barnes K.A., Snyder A.Z., Schlaggar B.L., Petersen S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage. 2012;59(3):2142–2154. doi: 10.1016/j.neuroimage.2011.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichle M.E., MacLeod A.M., Snyder A.Z., Powers W.J., Gusnard D.A., Shulman G.L. A default mode of brain function. Proc. Natl. Acad. Sci. U. S. A. 2001;98(2):676–682. doi: 10.1073/pnas.98.2.676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramey C.T., Ramey S.L. Early intervention and early experience. Am. Psychol. 1998;53(2):109–120. doi: 10.1037//0003-066x.53.2.109. [DOI] [PubMed] [Google Scholar]
- Ranganath C., Yonelinas A.P., Cohen M.X., Dy C.J., Tom S.M., D’Esposito M. Dissociable correlates of recollection and familiarity within the medial temporal lobes. Neuropsychologia. 2004;42(1):2–13. doi: 10.1016/j.neuropsychologia.2003.07.006. [DOI] [PubMed] [Google Scholar]
- Redcay E., Kennedy D.P., Courchesne E. fMRI during natural sleep as a method to study brain function during early childhood. Neuroimage. 2007;38(4):696–707. doi: 10.1016/j.neuroimage.2007.08.005. [DOI] [PubMed] [Google Scholar]
- Riggins T. Building blocks of recollection. In: Ghetti S., Bauer P.J., editors. Origins and Development of Recollection: Perspectives from Psychology and Neuroscience. Oxford University Press; New York, NY: 2012. [Google Scholar]
- Riggins T. Longitudinal investigation of source memory reveals qualitative differences between item memory and binding. Dev. Psychol. 2014;50:449–459. doi: 10.1037/a0033622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riggins T., Rollins L. Developmental changes in recollection and familiarity during early childhood: insights from event-related potentials. Child Dev. 2015;86:889–902. doi: 10.1111/cdev.12351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riggins T., Blankenship S., Mulligan E., Rice K., Redcay E. Episodic memory and structural development of the hippocampus during early childhood. Child Dev. 2015;86:1710–1718. doi: 10.1111/cdev.12445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roosendaal S.D., Schoonheim M.M., Hulst H.E., Sanz-Arigita E.J., Smith S.M., Geurts J.J.G., Barkhof F. Resting state networks change in clinically isolated syndrome. Brain. 2010;133(Pt 6):1612–1621. doi: 10.1093/brain/awq058. [DOI] [PubMed] [Google Scholar]
- Scoville W.B., Milner B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry. 1957;20(1):11–21. doi: 10.1136/jnnp.20.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serres L. Morphological changes of the human hippocampal formation from midgestation to early childhood. In: Nelson C.A., Luciana M., editors. Handbook of Developmental Cognitive Neuroscience. MIT Press; Cambridge, MA: 2001. pp. 45–58. [Google Scholar]
- Shing Y.L., Lindenberger U. The development of episodic memory: lifespan lessons. Child Dev. Perspect. 2011;5(2):148–155. [Google Scholar]
- Siegler R.S. Oxford University Press; New York: 1996. Emerging Minds: The Process of Change in Children's Thinking. [Google Scholar]
- Sluzenski J., Newcombe N.S., Kovacs S.L. Binding, relational memory, and recall of naturalistic events: a developmental perspective. J. Exp. Psychol.: Learn. Mem. Cognit. 2006;32(1):89–100. doi: 10.1037/0278-7393.32.1.89. [DOI] [PubMed] [Google Scholar]
- Smith S.M., Fox P.T., Miller K.L., Glahn D.C., Fox P.M., Mackay C.E.…Beckmann C.F. Correspondence of the brain's functional architecture during activation and rest. Proc. Natl. Acad. Sci. U. S. A. 2009;106(31):13040–13045. doi: 10.1073/pnas.0905267106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spaniol J., Davidson P.S.R., Kim A.S.N., Han H., Moscovitch M., Grady C.L. Event-related fMRI studies of episodic encoding and retrieval: meta-analyses using activation likelihood estimation. Neuropsychologia. 2009;47(8–9):1765–1779. doi: 10.1016/j.neuropsychologia.2009.02.028. [DOI] [PubMed] [Google Scholar]
- Supekar K., Musen M., Menon V. Development of large-scale functional brain networks in children. PLoS Biol. 2009;7(7):e1000157. doi: 10.1371/journal.pbio.1000157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uddin L.Q., Supekar K., Menon V. Typical and atypical development of functional human brain networks: insights from resting-state fMRI. Front. Syst. Neurosci. 2010;4 doi: 10.3389/fnsys.2010.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderwal T., Kelly C., Eilbott J., Mayes L.C., Castellanos F.X. Inscapes: a movie paradigm to improve compliance in functional magnetic resonance imaging. Neuroimage. 2015;122:222–232. doi: 10.1016/j.neuroimage.2015.07.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent J.L., Snyder A.Z., Fox M.D., Shannon B.J., Andrews J.R., Raichle M.E., Buckner R.L. Coherent spontaneous activity identifies a hippocampal-parietal memory network. J. Neurophysiol. 2006;96(6):3517–3531. doi: 10.1152/jn.00048.2006. [DOI] [PubMed] [Google Scholar]
- Wang L., Negreira A., LaViolette P., Bakkour A., Sperling R.A., Dickerson B.C. Intrinsic interhemispheric hippocampal functional connectivity predicts individual differences in memory performance ability. Hippocampus. 2010;20(3):345–351. doi: 10.1002/hipo.20771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L., Laviolette P., O’Keefe K., Putcha D., Bakkour A., Van Dijk K.R.A.…Sperling R.A. Intrinsic connectivity between the hippocampus and posteromedial cortex predicts memory performance in cognitively intact older individuals. Neuroimage. 2010;51(2):910–917. doi: 10.1016/j.neuroimage.2010.02.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster M.J., Ungerleider L.G., Bachevalier J. Connections of inferior temporal areas TE and TEO with medial temporal-lobe structures in infant and adult monkeys. J. Neurosci. 1991;11(4):1095–1116. doi: 10.1523/JNEUROSCI.11-04-01095.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss A.P., DeWitt I., Goff D., Ditman T., Heckers S. Anterior and posterior hippocampal volumes in schizophrenia. Schizophr. Res. 2005;73(1):103–112. doi: 10.1016/j.schres.2004.05.018. [DOI] [PubMed] [Google Scholar]
- Wilke M., Holland S.K., Altaye M., Gaser C. Template-O-Matic: a toolbox for creating customized pediatric templates. Neuroimage. 2008;41(3):903–913. doi: 10.1016/j.neuroimage.2008.02.056. [DOI] [PubMed] [Google Scholar]
- Yan C.-G., Zang Y.-F. DPARSF: a MATLAB toolbox for “pipeline” data analysis of resting-state fMRI. Front. Syst. Neurosci. 2010;4:13. doi: 10.3389/fnsys.2010.00013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yonelinas A.P., Otten L.J., Shaw K.N., Rugg M.D. Separating the brain regions involved in recollection and familiarity in recognition memory. J. Neurosci. 2005;25(11):3002–3008. doi: 10.1523/JNEUROSCI.5295-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeamer A., Heuer E., Bachevalier J. Developmental trajectory of object recognition memory in infant rhesus macaques with and without neonatal hippocampal lesions. J. Neurosci. 2010;30:9157–9165. doi: 10.1523/JNEUROSCI.0022-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zola S.M., Squire L.R., Teng E., Stefanacci L., Buffalo E.A., Clark R.E. Impaired recognition memory in monkeys after damage limited to the hippocampal region. J. Neurosci. 2000;20:451–463. doi: 10.1523/JNEUROSCI.20-01-00451.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.