

Abstract
Degenerative brain changes in Alzheimer’s disease may occur in reverse order of normal brain development based on the retrogenesis model. This study tested whether evidence of reverse myelination was observed in mild cognitive impairment (MCI) using a data-driven analytic approach based on life span developmental data. Whole-brain high-resolution diffusion tensor imaging scans were obtained for 31 patients with MCI and 79 demographically matched healthy older adults. Comparisons across corpus callosum (CC) regions of interest (ROIs) showed decreased fractional anisotropy (FA) in the body but not in the genu or splenium; early-, middle-, and late-myelinating ROIs restricted to the CC revealed decreased FA in late- but not early- or middle-myelinating ROIs. Voxelwise group differences revealed areas of lower FA in MCI, but whole-brain differences were equally distributed across early-, middle-, and late-myelinating regions. Overall, results within the CC support the retrogenesis model, although caution is needed when generalizing these results beyond the CC.
Keywords: diffusion tensor imaging, magnetic resonance imaging, mild cognitive impairment, corpus callosum, Alzheimer’s disease
Introduction
White matter changes are well documented in Alzheimer’s dementia (AD) 1 -3 and its prodromal stages, including mild cognitive impairment (MCI). 4 -8 However, the pattern and underlying mechanism of these changes remain unclear. One reason for this uncertainty is that few diffusion tensor imaging (DTI) studies have tested a priori hypotheses based on a specified neuropathologic mechanism. The retrogenesis model of AD provides a testable model and posits that white matter degeneration reflects myelin breakdown that develops in a pattern that is the reverse of myelogenesis. 9,10,11 White matter pathways with large diameter fibers that myelinate first in development, such as primary motor fibers, are the last to be affected by AD. In contrast, white matter pathways with small diameter fibers that myelinate much later in normal development, such as neocortical association and allocortical fibers, are the first to be affected by the AD degenerative process. 12 Recent studies have demonstrated support for the retrogenesis model in patients with AD, 13,14,3 although support for this model is less clear in MCI. 13,14
Assumptions about the order of myelination throughout development vary across study methods used and contribute to difficulty in interpreting results within the context of the retrogenesis model (eg, volumetric imaging, diffusion imaging, and histological procedures may provide differing results). Consistent with the pattern of gray matter (GM) cortical development, 15 -17 pathological studies of myelination of human infants suggest a pattern of white matter development that is proximal to distal, projection to association, and occipital to frontal. 18,19 More recent studies have attempted to examine life span white matter changes in vivo utilizing magnetic resonance imaging (MRI) methodology, including white matter volumetric study, diffusion parameters, and tractography. Indeed, there is evidence that association tracts continue to develop in the 20s, followed by projection fibers in the 30s. 20,21 Further, in a cross-sectional study Westlye and colleagues 22 found that peaks in tract-based spatial statistics (TBSS) fractional anisotropy (FA), mean diffusivity (MD), and radial diffusivity occurred on average between 29 and 36 years, whereas white matter volume segmentation noted growth into the sixth decade. In normal aging, white matter pathways undergo a nonlinear, hierarchical maturation pattern such that there is a rapid acceleration of development (increases in FA and decreases in MD) during early life, a mid-life plateau, and a decline in later adulthood; however, there are regional differences in the course of these 3-phase models. Regions with frontotemporal connections typically develop at a slower rate than regions with less extensive connections. 22 -24 Those with protracted development, including portions of the corpus callosum (CC), have been shown to demonstrate rapid development in early childhood, a later plateau stage as well as accelerated decline in tract integrity with age. 22,24 -28 However, there are methodological considerations differentiating these studies, including MRI method (volumetric, DTI, and/or fiber tracking), regional versus whole-brain analysis, and participant inclusion. Overall, it appears that cortico–cortical association pathways followed by commissural pathways represent the latest myelinating regions in the brain.
The CC has frequently been targeted for investigating changes in early- versus late-myelinating regions in studies of aging and AD. 14,29 -35 It is often assumed that the genu is later myelinating relative to the splenium due to the general tendency of frontal regions to myelinate later than posterior regions 18 and the predominance of fibers connecting prefrontal regions in the genu versus temporoparietal fibers in the splenium. 36,37 Decreased white matter integrity within the CC has been previously reported in patients with AD relative to healthy controls 6,12,34,37 -42 and MCI. 8,12,31,32,43 -48 However, inconsistencies exist in the literature base regarding the location of abnormalities within the CC, when changes emerge across the disease course and the underlying mechanisms driving the changes. In particular, posterior callosal abnormalities (eg, splenium) have been hypothesized to reflect Wallerian degeneration attributable to cortical AD neuropathology, 12,32,44,47 -49 whereas degeneration of more anterior callosal regions (eg, genu) has been suggested to reflect retrogenesis of those areas. 12,32,44,47,48 The DTI studies of MCI and AD using ROI methods have frequently omitted the body of the CC 6,8,31,35,38,39,40,43,46,49,50,51 ; however, those studies that have included it or used a whole-brain voxelwise analysis approach have often demonstrated decreased white matter integrity in the body of the CC in MCI and AD. 12,32,37,44,48
Interestingly, some assumptions within the literature about the developmental order within the CC were recently challenged by data presented by Lebel and colleagues, 36 whose results suggest that the body of the CC should not be omitted from studies of MCI and AD. The authors divided the callosum into 7 separate regions, creating boundaries that are functionally as well as anatomically distinct. Although their lifetime white matter development findings were consistent with prior studies demonstrating an inverted-U curve of increased FA and decreased MD throughout adolescence and early adulthood, followed by a plateau and reverse growth in later adulthood, prior fit models were unable to assess relative timing of the 7 regions across the lifetime. 23 Their results suggested an “outer-to-inner” pattern of decreasing white matter integrity with age within the CC and indicate that examination of the entire callosum, including the body, is necessary to account for potential differences in neurodevelopmental timing.
Given the variation in assumptions about the order of myelination, we aimed to take a data-driven approach to testing the retrogenesis model, with a focus on the CC. We defined myelination order according to previously acquired data across a large normative sample spanning ages 8 to 85 22 and used a priori, empirically defined ROIs to test the retrogenesis model within the CC and across the whole brain. We predicted that FA would be lower in patients with MCI relative to healthy older adults in late-myelinating regions. We further expected lower FA in the body of the CC in patients with MCI relative to healthy older adults based on recent data suggesting this region to be late myelinating relative to the genu and splenium. 36
Methods
Participants
One hundred and ten participants underwent neuropsychological testing and brain MRI. Participants were recruited from 2 overlapping studies conducted at VA Boston Healthcare System. Twenty-eight participants were selected (based on their agreement to undergo structural MRI) from a larger sample recruited from the community through the Harvard Cooperative Program on Aging (HCPA) Claude Pepper Older American Independence Center (OAIC). These participants responded to an HCPA newsletter asking for healthy community-dwelling older African Americans. Eighty-two participants were part of the Understanding Cerebrovascular and Alzheimer’s Risk in the Elderly (UCARE) program, recruited through the Boston University Alzheimer’s Disease Center (BUADC) based on the criteria of being neurologically healthy and having a first-degree family relative with AD. Data were analyzed on a subset of individuals presented previously. 7 Participants were excluded for the following: history of head trauma of “mild” severity or greater 52 (within our sample loss of consciousness did not exceed 15 minutes), more than 1 head injury, any neurological disorder including dementia (ie, Parkinson’s disease, AD, vascular dementia), severe psychiatric illness, or brain surgery. All participants were literate with at least a ninth-grade education. The VA Boston Healthcare System’s institutional review board approved the study according to the Helsinki Declaration, and informed consent was obtained from each participant.
Criteria for MCI
Retrospective diagnosis of MCI was made by applying the “comprehensive” criteria 53 based on neuropsychological test scores across 4 cognitive domains including memory, attention/processing speed, language, and executive functions, as described previously. 7 To contribute to MCI classification, at least 2 performances within a cognitive domain fell 1 standard deviation (SD) or more below published normative expectations for that domain. All participants scored ≥24 (caucasians) or ≥23 (African Americans) 54 on the Mini-Mental State Examination (MMSE). Participants were classified as amnestic MCI (aMCI) if memory was impaired (either alone or with additional domains impaired) and as nonamnestic MCI (naMCI) if domains other than memory were impaired. Participants were classified as healthy controls (HC) if performance on no more than 1 measure within a cognitive domain fell more than 1 SD below normative data. Using these criteria, 31 participants met criteria for MCI (14 aMCI and 17 naMCI), and 79 participants were classified as HC. Self-report measures included the Geriatric Depression Scale (GDS) 55 and the Lawton and Brody Instrumental Activities Of Daily Living (IADLs) questionnaire. 56
Neuroimaging Protocol
Five participants were scanned using a Siemens 1.5 Tesla Sonata system (Siemens Erlangen, Germany), with the following parameters: DTI: repetition time (TR) = 9000 milliseconds and echo time (TE) = 68 milliseconds, 60 slices total, acquisition matrix = 128 × 128 (field of view [FOV] = 256 × 256 mm), slice thickness = 2 mm (for 2-mm isotropic voxels) with 0 mm gap, with a b value = 700 s/mm2, 10 T2- and 60 diffusion-weighted images, and 1 image, the T2-weighted “low b” image with a b value = 0 s/mm2 as an anatomical reference volume. The remaining 105 participants were scanned on the upgraded Siemens 1.5 Avanto System, with slightly different parameters: DTI: TR = 7200 milliseconds and TE = 77 milliseconds, 60 slices total, acquisition matrix = 128 × 128 (FOV = 256 × 256 mm), slice thickness = 2 mm (for 2-mm isotropic voxels) with 0-mm gap, with a b value = 700 s/mm2, 10 T2- and 60 diffusion-weighted images, and 1 image, the T2-weighted “low b” image with a b value = 0 s/mm2 as an anatomical reference volume.
Image Processing
Diffusion data went through a multistep processing pipeline involving tools from the FreeSurfer image analysis suite and Functional Magnetic Resonance Imaging of the Brain (FMRIB) Software Library (FSL) (http://www.fmrib.ox.ac.uk.fsl/), specifically Dtifit from FMRIB’s Diffusion Toolbox 57 and TBSS, 58 part of FSL. 59 See our previous work 2,60 for additional details. Fractional anisotropy (FA), a measure of the diffusion displacement probability of water within white matter, was used as the primary metric of white matter integrity for this study. Lower FA is generally taken to indicate lower white matter integrity.
Regions of interest (ROIs) restricted to the TBSS skeleton for early-, middle-, and late-myelinating regions were created based on available data. Order of myelination of different white matter regions was empirically determined based on data presented in Westlye et al 22 wherein age trajectories of FA was determined from 430 healthy participants (age range 8-85). Summary data were provided in the form of a TBSS skeleton map with voxel values corresponding to age of nonparametric locally weighted polynomial regression (LOESS) 61 estimated peak FA; see Westlye et al 22 for details. This was divided into 3 ROI myelination tertiles (see Figure 1) defined as early myelinating (age FA peak <25.5), middle myelinating (age FA peak 25.5-29.08), and late myelinating (age FA peak >29.08).
Figure 1.
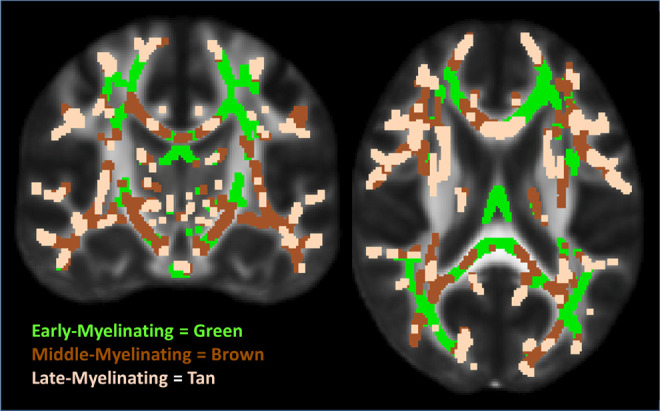
Early-, middle-, and late-myelinating ROIs. ROIs dilated for visualization purposes only. ROIs indicates regions of interest.
Region of interests limited to the TBSS skeleton for the genu, body, and splenium of the CC were created using the Johns Hopkins University (JHU) white matter labels, available as part of the FSL suite. In addition, the genu, body, and splenium ROIs were added together to create one larger CC mask, and then the above-described early-, middle-, and late-myelinating ROIs were applied to that mask. This yielded early-, middle-, and late-myelinating ROIs restricted to the CC (subsequently referred to as CC-early, CC-middle, and CC-late). Average FA was derived for each white matter ROI and was extracted from voxels limited to the TBSS skeleton to reduce the influence of partial volume contamination.
Statistical Analyses
Independent samples t tests were used to examine group differences across CC ROIs. Voxelwise statistics were performed across the TBSS skeleton to test for whole-brain differences in FA across MCI and HC groups. As the primary goal was to evaluate the proportion of voxels significant across the myelination tertile ROIs and not report on the exact localization of differences found, the voxelwise analysis did not control for multiple comparisons. A minimum cluster size of 10 was required and a mask of all significant voxels was saved. Masks for the early-, middle-, and late-myelinating ROIs were then applied to the significant clusters mask to determine what percentage of the total of significant voxels from the voxelwise analyses fell within early-, middle-, and late-myelinating regions. Chi-square analyses were used to compare frequencies of significant voxels across the 3 tertile ROIs. A significance level of P < .05 was used for all analyses.
Results
Participants
The MCI and HC groups were comparable on age (t = −0.56, P = .58), education (t = 0.17, P = .87), sex (χ2 = 0.04, P = .85), ethnicity (χ2 = 3.16, P = .08), Lawton and Brody IADL score (t = 1.83, P = .07), GDS (t = −0.93, P = .36), and MMSE (t = 1.77, P = .08). Due to targeted recruitment, a large percentage (92%) had a self-reported positive family history of AD, and this percentage was comparable across MCI and HC groups (χ2 = 0.05, P = .82; see Table 1).
Table 1.
Demographics and FA by Group.
| HC (n = 79) | MCI (n = 31) | |
|---|---|---|
| Age | 67.81 (9.02) | 68.94 (10.64) |
| Education | 14.94 (2.83) | 14.84 (2.46) |
| Sex, n male/female | 29/50 | 12/19 |
| Ethnicity, n caucasian/AA | 55/24 | 16/15 |
| MMSE | 28.07 (1.65) | 27.40 (1.96) |
| Lawton-Brody IADL | 26.58 (0.96) | 26.16 (1.37) |
| GDS | 4.03 (5.59) | 5.13 (5.64) |
| FA Genu of CC | 0.682 (0.049) | 0.680 (0.041) |
| FA Body of CCa | 0.659 (0.042) | 0.635 (0.044) |
| FA Splenium of CC | 0.760 (0.041) | 0.753 (0.034) |
| FA CC Early | 0.662 (0.040) | 0.654 (0.035) |
| FA CC Middle | 0.695 (0.043) | 0.680 (0.040) |
| FA CC Latea | 0.691 (0.042) | 0.666 (0.046) |
Abbreviations: HC, normal controls; MCI, mild cognitive impairment; AA, African American; MMSE, Mini-Mental Status Examination; FA, fractional anisotropy; CC, corpus callosum; GDS, Geriatric Depression Scale; IADL, instrumental activities of daily living.
a P < .05.
Corpus Callosum
The MCI group demonstrated lower FA than the HC group in the body of the CC (t = 2.69, P = .008); see Table 1. There were no significant group differences in the genu (t = 0.27, P = .79) or splenium (t = 0.83, P = .41). To determine the relative order of myelination of each CC ROI, we applied the myelination tertile ROI map to the JHU CC ROIs. The genu was comprised of predominantly early-myelinating voxels (58.5%; 40.0% middle myelinating and 1.5% late myelinating). The body was comprised of predominantly late-myelinating voxels (47.8%; 37.2% middle myelinating and 15.1% early myelinating). The splenium was comprised of predominantly middle- (50.5%) and early-myelinating (47.8%) voxels (1.8% late myelinating). Average FA extracted from the entire CC for early-, middle-, and late-myelinating tertiles, respectively, showed no significant group differences in the CC-early (t = 0.90, P = .37) or CC-middle (t = 1.66, P = .10) ROIs, but the MCI group showed decreased FA in the CC-late ROI (t = 2.70, P = .008) relative to the HC group.
Whole-Brain Voxelwise Results
Whole-brain voxelwise statistics demonstrated numerous areas of decreased FA in the MCI group relative to the HC group (approximately 15% of all voxels were significant). There were no significant findings for the other contrast (MCI > HC). Chi-square analyses revealed no differences in the frequency of significant voxels across the early-, middle-, and late-myelinating ROIs (χ2 = 0.91, P = .64). These results indicate that group differences are equally likely regardless of order of myelination.
Discussion
The current findings revealed decreased FA in late-myelinating regions within the CC and decreased FA in the body of the CC (comprised predominantly of late-myelinating WM) in individuals with MCI relative to healthy older adults. These results suggest that the pattern of changes in white matter integrity in MCI within the CC supports the retrogenesis model when late-myelinating regions are empirically defined in an independent data set.
In contrast to previous research which considered the genu as a late-myelinating region, given its connections to prefrontal white matter, findings from Lebel et al 36 and the data presented here both demonstrate a general trend for outer regions (eg, genu and splenium) of the CC to myelinate earlier than inner regions (eg, body). Our finding of decreased white matter integrity in the body of the CC in MCI in this study, combined with previous evidence that this region is later myelinating relative to the genu and splenium, suggests that the body is an important region in white matter neurodevelopment and worth assessment in future studies. Our results are consistent with other findings showing decreased white matter integrity in this CC region in MCI and AD. 12,32,37,44,48 The genu and splenium, which did not reveal significant findings in ROI analyses, may be composed of predominantly early- and middle-myelinating fibers. Previous studies have shown varying results within these regions, with some revealing decreased white matter integrity in the anterior CC in patients with MCI versus healthy older adults, 12,32,44,45,47,48,51 while others have revealed no anterior CC abnormalities in MCI 4,46 or in patients with AD. 38,39 Decreased white matter integrity in the splenium is reported to be a fairly consistent finding across studies of AD (see Chua et al for review 62 ); thus, it is somewhat surprising that no differences across MCI and HC groups were found in this ROI in our study. Although the splenium has been frequently implicated in DTI studies of patients with AD, this finding has been less consistent in MCI, with some studies noting lower white matter integrity 4,6,46 and others reporting null results in the posterior CC 12,32,51 ; this inconsistency in the literature may help to explain our results and suggests a need for additional examinations. Further, some studies have also failed to show any differences in white matter integrity in ROIs of the genu and splenium in MCI or AD relative to healthy controls, 43,50,51 consistent with our results.
In contrast to our CC findings, results of the whole-brain voxelwise analysis did not support the retrogenesis model. Decreased white matter integrity in MCI was equally likely across early-, middle-, and late-myelinating regions. It is possible that interhemispheric fibers may be more likely to reflect developmentally based order of myelination because of properties inherent to the CC. For example, the CC is composed predominantly of bidirectional interhemispheric connections conjoined with few crossing fibers and a known, clearly delineated pattern of increasing anisotropy from anterior to posterior. 63,64 Any alterations to CC white matter integrity, including loss of myelination or reduced spatial organization of white matter, may be more easily detected in the CC relative to smaller white matter pathways that contain more crossing fibers and have less well-defined anisotropy profiles. It may also be that other properties of the CC, such as higher signal to noise ratio and higher reliability given its greater homogeneity of fibers, may also help explain the discrepancy between results within the CC and those of the whole-brain analysis.
This study has a number of limitations. First, it will be important for future research to replicate these results. Because follow-up data are unavailable and MCI can reflect a variety of etiologies, it is possible that the results presented herein represent normal variability in white matter morphology that is indicative of normative cognitive strengths and weaknesses, rather than an Alzheimer’s disease trajectory. Reproducing these results in an MCI cohort that later converts to AD would help verify that results represent a pattern of change expected early in the Alzheimer’s pathological process. Second, changes in CC white matter have been shown to correlate with the distribution of Wallerian degeneration at cortical regions following ischemic damage. 65 It is possible then that the findings reported here supporting retrogenesis in the CC reflect a larger, brain-wide pattern of pathogenesis that follows the retrogenesis model that were not detectable in the whole-brain voxel-based white matter analysis. Future studies could use cortical thickness analyses to investigate this hypothesis. Similarly, although decreased FA has been associated with decreased myelin in mouse models, 66 decreased diffusion anisotropy is due to changes in myelin as well as other factors, such as axonal structure and homogeneity of the direction of fiber pathways within a voxel. 67,68 Thus, the myelination tertile ROIs cannot be assumed to only reflect increased myelination with age as FA does not represent a direct measure of myelination and is thus not likely to capture all relevant myelin-related neurodevelopmental or neurodegenerative variability. Third, due to small sample sizes and an effort to limit the number of analyses, we chose not to analyze aMCI and naMCI subgroups separately, given evidence that naMCI frequently represents early Alzheimer’s disease. 69 -71 However, this may make comparisons with other MCI studies more difficult, as samples including only aMCI are more common. Fourth, there may be disparities between the sample used to generate the myelination tertile ROIs 22 and the sample under investigation in this study. The participants from which the myelination tertile ROIs were derived were of above average cognitive functioning and are not necessarily representative of the general population or of the developmental trajectory of the sample used here. In contrast, the neurocognitive performance of participants in this sample was widely distributed and included participants with MCI. Additionally, cohort effects inherent in the cross-sectional design used by Westlye and colleagues 22 may contribute to the current results in that group-wise differences in early development (eg, nutrition and education) could potentially alter white matter developmental trajectory. Longitudinal studies could help clarify the role of environmental and genetic factors on white matter development and degeneration over the life span. Finally, the diffusion data for both the Westlye study and the current study were derived from the center of each pathway based on the processing methodology employed (TBSS). Thin myelinated fibers closer to the cortex or toward the periphery of white matter pathways may be more vulnerable to changes with age or neurodegeneration than the centers of white matter pathways and deeper structures. 72 -74 Therefore, the center-derived DTI methodology used here may predispose findings to be less sensitive to changes in these lateral regions than in regions such as the CC. Alternate methods of projecting anisotropy maps may allow future studies to investigate age-related white matter changes more sensitively in these more thinly myelinated regions. Finally, considering other imaging indices of white matter integrity and myelination in conjunction with DTI metrics may provide a complementary view of the age trajectories of white matter across the human life span and in MCI samples.
In conclusion, these findings support retrogenesis as a mechanism underlying white matter integrity changes within the CC among individuals with MCI. The historical lack of emphasis upon understanding development of brain white matter tracts in the study of callosal changes in AD may explain the lack of consistency that has existed in the literature to date across anterior, posterior, and middle regions of the CC. The current methodology based on empirical data rather than long-standing assumptions about order of myelination provides a novel perspective on interpretation of CC findings that will contribute to improved understanding regarding the presence and pattern of regional callosal involvement in MCI.
Footnotes
Authors’ Note: The contents of this article do not represent the views of the US Department of Veterans Affairs or the United States Government.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding: The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Department of Veterans Affairs, Veterans Health Administration, VISN 1 Career Development Award, and the Gilbert Foundation/AFAR. This work was also supported by the Medical Research Service VA Merit Review Awards, Veterans Affairs Clinical Science R&D Career Development Award (1IK2CX000706-01A2), the National Institute of Neurologic Disorders and Stroke (grant number K23NS062148); the National Institute of Nursing Research (grant number R01NR010827), the National Institute on Aging (grant numbers P60AG08812, P01AG004390); the Research Council of Norway (204966/F20), and by the National Institute on Drug Abuse (2T32DA015036).
References
- 1. Brun A, Englund E. A white matter disorder in dementia of the Alzheimer type: a pathoanatomical study. Ann Neurol. 1986;19(3):253–262. [DOI] [PubMed] [Google Scholar]
- 2. Salat DH, Tuch DS, van der Kouwe AJ, et al. White matter pathology isolates the hippocampal formation in Alzheimer's disease. Neurobiol Aging. 2010;31(2):244–256.doi:10.1016/j.neurobiolaging.2008.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Stricker NH, Schweinsburg BC, Delano-Wood L, et al. Decreased white matter integrity in late-myelinating fiber pathways in Alzheimer's disease supports retrogenesis. Neuroimage. 2009;45(1):10–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Delano-Wood L, Bondi MW, Jak AJ, et al. Stroke risk modifies regional white matter differences in mild cognitive impairment. Neurobiol Aging. 2010;31(10):1721–1731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Fellgiebel A, Muller MJ, Wille P, et al. Color-coded diffusion-tensor-imaging of posterior cingulate fiber tracts in mild cognitive impairment. Neurobiol Aging. 2005;26(8):1193–1198. [DOI] [PubMed] [Google Scholar]
- 6. Parente DB, Gasparetto EL, da Cruz LC, Jr, et al. Potential role of diffusion tensor MRI in the differential diagnosis of mild cognitive impairment and Alzheimer's disease. AJR Am J Roentgenol. 2008;190(5):1369–1374. [DOI] [PubMed] [Google Scholar]
- 7. Stricker NH, Salat DH, Foley JM, et al. Decreased white matter integrity in neuropsychologically defined mild cognitive impairment is independent of cortical thinning. J Int Neuropsychol Soc. 2013;19(8):925–937. doi:10.1017/S1355617713000660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Wang L, Goldstein FC, Veledar E, et al. Alterations in cortical thickness and white matter integrity in mild cognitive impairment measured by whole-brain cortical thickness mapping and diffusion tensor imaging. AJNR Am J Neuroradiol. 2009;30(5):893–899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Bartzokis G. Alzheimer's disease as homeostatic responses to age-related myelin breakdown. Neurobiol Aging. 2011;32(8):1341–1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Reisberg B, Franssen EH, Hasan SM, et al. Retrogenesis: clinical, physiologic, and pathologic mechanisms in brain aging, Alzheimer's and other dementing processes. Eur Arch Psychiatry Clin Neurosci. 1999;249(suppl 3):28–36. [DOI] [PubMed] [Google Scholar]
- 11. Bartzokis G. Age-related myelin breakdown: a developmental model of cognitive decline and Alzheimer's disease. Neurobiol Aging. 2004;25(1):5–18; author reply 49-62. [DOI] [PubMed] [Google Scholar]
- 12. Alves GS, O'Dwyer L, Jurcoane A, et al. Different patterns of white matter degeneration using multiple diffusion indices and volumetric data in mild cognitive impairment and Alzheimer patients. PloS One. 2012;7(12):e52859. doi:10.1371/journal.pone.0052859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Benitez A, Fieremans E, Jensen JH, et al. White matter tract integrity metrics reflect the vulnerability of late-myelinating tracts in Alzheimer's disease. Neuroimage Clin. 2014;4:64–71. doi:10.1016/j.nicl.2013.11.001 S2213-1582(13)00149-6 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fieremans E, Benitez A, Jensen JH, et al. Novel white matter tract integrity metrics sensitive to Alzheimer disease progression. AJNR Am J Neuroradiol. 2013;34(11):2105–2112. doi:ajnr.A3553 [pii] 10.3174/ajnr.A3553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Giedd JN, Blumenthal J, Jeffries NO, et al. Brain development during childhood and adolescence: a longitudinal MRI study. Nat Neurosci. 1999;2(10):861–863. doi:10.1038/13158. [DOI] [PubMed] [Google Scholar]
- 16. Gogtay N, Thompson PM. Mapping gray matter development: implications for typical development and vulnerability to psychopathology. Brain Cogn. 2010;72(1):6–15. doi:10.1016/j.bandc.2009.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Pfefferbaum A, Mathalon DH, Sullivan EV, Rawles JM, Zipursky RB, Lim KO. A quantitative magnetic resonance imaging study of changes in brain morphology from infancy to late adulthood. Arch Neurol. 1994;51(9):874–887. [DOI] [PubMed] [Google Scholar]
- 18. Kinney HC, Brody BA, Kloman AS, Gilles FH. Sequence of central nervous system myelination in human infancy. II. Patterns of myelination in autopsied infants. J Neuropathol Exp Neurol. 1988;47(3):217–234. [DOI] [PubMed] [Google Scholar]
- 19. Yakovlev PI, Lecours AR. The myelogenetic cycles of regional maturation of the brain. In: Minkowski A, ed. Regional Development of the Brain in Early Life. Philadelphia: F. A. Davis Company; 1967:3–70. [Google Scholar]
- 20. Lebel C, Beaulieu C. Longitudinal development of human brain wiring continues from childhood into adulthood. J Neurosci. 2011;31(30):10937–10947. doi:10.1523/JNEUROSCI.5302-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Lebel C, Gee M, Camicioli R, Wieler M, Martin W, Beaulieu C. Diffusion tensor imaging of white matter tract evolution over the lifespan. Neuroimage. 2012;60(1):340–352. doi:10.1016/j.neuroimage.2011.11.094. [DOI] [PubMed] [Google Scholar]
- 22. Westlye LT, Walhovd KB, Dale AM, et al. Life-span changes of the human brain white matter: diffusion tensor imaging (DTI) and volumetry. Cerebral Cortex. 2010;20(9):2055–2068. doi:10.1093/cercor/bhp280. [DOI] [PubMed] [Google Scholar]
- 23. Hasan KM, Halphen C, Sankar A, et al. Diffusion tensor imaging-based tissue segmentation: validation and application to the developing child and adolescent brain. Neuroimage. 2007;34(4):1497–1505. doi:10.1016/j.neuroimage.2006.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Lebel C, Walker L, Leemans A, Phillips L, Beaulieu C. Microstructural maturation of the human brain from childhood to adulthood. Neuroimage. 2008;40(3):1044–1055. doi:10.1016/j.neuroimage.2007.12.053. [DOI] [PubMed] [Google Scholar]
- 25. Sridharan A, Bendlin BB, Gallagher CL, et al. Effect of age and calorie restriction on corpus callosal integrity in rhesus macaques: a fiber tractography study. Neurosci Lett. 2014;569:38–42. doi:10.1016/j.neulet.2014.03.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Sullivan EV, Pfefferbaum A. Diffusion tensor imaging and aging. Neurosci Biobehav Rev. 2006;30(6):749–761. [DOI] [PubMed] [Google Scholar]
- 27. Teipel SJ, Lerche M, Kilimann I, et al. Decline of fiber tract integrity over the adult age range: a diffusion spectrum imaging study. J Magn Reson Imaging. 2014;40(2):348–359. doi:10.1002/jmri.24420. [DOI] [PubMed] [Google Scholar]
- 28. Walhovd KB, Westlye LT, Amlien I, et al. Consistent neuroanatomical age-related volume differences across multiple samples. Neurobiol Aging. 2011;32(5):916–932. doi:10.1016/j.neurobiolaging.2009.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Bartzokis G, Cummings JL, Sultzer D, Henderson VW, Nuechterlein KH, Mintz J. White matter structural integrity in healthy aging adults and patients with Alzheimer disease: a magnetic resonance imaging study. Arch Neurol. 2003;60(3):393–398. [DOI] [PubMed] [Google Scholar]
- 30. Bartzokis G, Lu PH, Geschwind DH, et al. Apolipoprotein E affects both myelin breakdown and cognition: implications for age-related trajectories of decline into dementia. Biol Psychiatry. 2007;62(12):1380–1387. doi:S0006-3223(07)00318-6 [pii] 10.1016/j.biopsych.2007.03.024. [DOI] [PubMed] [Google Scholar]
- 31. Delano-Wood L, Abeles N, Sacco JM, Wierenga CE, Horne NR, Bozoki A. Regional white matter pathology in mild cognitive impairment: differential influence of lesion type on neuropsychological functioning. Stroke. 2008;39(3):794–799. doi:10.1161/STROKEAHA.107.502534. [DOI] [PubMed] [Google Scholar]
- 32. Di Paola M, Di Iulio F, Cherubini A, et al. When, where, and how the corpus callosum changes in MCI and AD: a multimodal MRI study. Neurology. 2010;74(14):1136–1142. doi:74/14/1136 [pii] 10.1212/WNL.0b013e3181d7d8cb. [DOI] [PubMed] [Google Scholar]
- 33. Di Paola M, Spalletta G, Caltagirone C. In vivo structural neuroanatomy of corpus callosum in Alzheimer's disease and mild cognitive impairment using different MRI techniques: a review. J Alzheimers Dis. 2010;20(1):67–95. doi: R06Q23414651653 H [pii] 10.3233/JAD-2010-1370. [DOI] [PubMed] [Google Scholar]
- 34. Hanyu H, Asano T, Sakurai H, et al. Diffusion-weighted and magnetization transfer imaging of the corpus callosum in Alzheimer's disease. J Neurol Sci. 1999;167(1):37–44. [DOI] [PubMed] [Google Scholar]
- 35. Head D, Buckner RL, Shimony JS, et al. Differential vulnerability of anterior white matter in nondemented aging with minimal acceleration in dementia of the Alzheimer type: evidence from diffusion tensor imaging. Cerebral Cortex. 2004;14(4):410–423. [DOI] [PubMed] [Google Scholar]
- 36. Lebel C, Caverhill-Godkewitsch S, Beaulieu C. Age-related regional variations of the corpus callosum identified by diffusion tensor tractography. NeuroImage. 2010;52(1):20–31. doi: S1053-8119(10)00359-9 [pii] 10.1016/j.neuroimage.2010.03.072. [DOI] [PubMed] [Google Scholar]
- 37. Preti MG, Baglio F, Lagana MM, et al. Assessing corpus callosum changes in Alzheimer's disease: comparison between tract-based spatial statistics and atlas-based tractography. PloS One. 2012;7(4):e35856. doi:10.1371/journal.pone.0035856 PONE-D-11-15104 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Duan JH, Wang HQ, Xu J, et al. White matter damage of patients with Alzheimer's disease correlated with the decreased cognitive function. Surg Radiol Anat. 2006;28(2):150–156. [DOI] [PubMed] [Google Scholar]
- 39. Naggara O, Oppenheim C, Rieu D, et al. Diffusion tensor imaging in early Alzheimer's disease. Psychiatry Res. 2006;146(3):243–249. [DOI] [PubMed] [Google Scholar]
- 40. Takahashi S, Yonezawa H, Takahashi J, Kudo M, Inoue T, Tohgi H. Selective reduction of diffusion anisotropy in white matter of Alzheimer disease brains measured by 3.0 Tesla magnetic resonance imaging. Neurosci Lett. 2002;332(1):45–48. [DOI] [PubMed] [Google Scholar]
- 41. Teipel SJ, Stahl R, Dietrich O, et al. Multivariate network analysis of fiber tract integrity in Alzheimer's disease. NeuroImage. 2007;34(3):985–995. [DOI] [PubMed] [Google Scholar]
- 42. Teipel SJ, Wegrzyn M, Meindl T, et al. Anatomical MRI and DTI in the diagnosis of Alzheimer's disease: a European multicenter study. J Alzheimers Dis. 2012;31(suppl 3):S33–S47. doi:10.3233/JAD-2012-112118. [DOI] [PubMed] [Google Scholar]
- 43. Fellgiebel A, Wille P, Muller MJ, et al. Ultrastructural hippocampal and white matter alterations in mild cognitive impairment: a diffusion tensor imaging study. Dement Geriatr Cogn Disord. 2004;18(1):101–108. [DOI] [PubMed] [Google Scholar]
- 44. Liu J, Yin C, Xia S, et al. White matter changes in patients with amnestic mild cognitive impairment detected by diffusion tensor imaging. PloS One. 2013;8(3):e59440. doi:10.1371/journal.pone.0059440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Scrascia F, Curcio G, Ursini F, et al. Relationship among diffusion tensor imaging, EEG activity, and cognitive status in mild cognitive impairment and Alzheimer's disease patients. J Alzheimers Dis. 2014;38(4):939–950. doi:10.3233/JAD-130788. [DOI] [PubMed] [Google Scholar]
- 46. Ukmar M, Makuc E, Onor ML, Garbin G, Trevisiol M, Cova MA. Evaluation of white matter damage in patients with Alzheimer's disease and in patients with mild cognitive impairment by using diffusion tensor imaging. La Radiologia medica. 2008;113(6):915–922. doi:10.1007/s11547-008-0286 -1. [DOI] [PubMed] [Google Scholar]
- 47. Wang PN, Chou KH, Chang NJ, et al. Callosal degeneration topographically correlated with cognitive function in amnestic mild cognitive impairment and Alzheimer's disease dementia. Hum Brain Map. 2014;35(4):1529–1543. doi:10.1002/hbm.22271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Xie S, Xiao JX, Gong GL, et al. Voxel-based detection of white matter abnormalities in mild Alzheimer disease. Neurology. 2006;66(12):1845–1849. [DOI] [PubMed] [Google Scholar]
- 49. Bozzali M, Falini A, Franceschi M, et al. White matter damage in Alzheimer's disease assessed in vivo using diffusion tensor magnetic resonance imaging. J Neurol Neurosurg Psychiatry. 2002;72(6):742–746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bozzao A, Floris R, Baviera ME, Apruzzese A, Simonetti G. Diffusion and perfusion MR imaging in cases of Alzheimer's disease: correlations with cortical atrophy and lesion load. AJNR Am J Neuroradiol. 2001;22(6):1030–1036. [PMC free article] [PubMed] [Google Scholar]
- 51. Stahl R, Dietrich O, Teipel SJ, Hampel H, Reiser MF, Schoenberg SO. White matter damage in Alzheimer disease and mild cognitive impairment: assessment with diffusion-tensor MR imaging and parallel imaging techniques. Radiology. 2007;243(2):483–492. [DOI] [PubMed] [Google Scholar]
- 52. Holm L, Cassidy JD, Carroll LJ, Borg J. Summary of the WHO collaborating centre for neurotrauma task force on mild traumatic brain injury. J Rehabil Med. 2005;37(3):137–141. [DOI] [PubMed] [Google Scholar]
- 53. Jak AJ, Bondi MW, Delano-Wood L, et al. Quantification of five neuropsychological approaches to defining mild cognitive impairment. Am J Geriatr Psychiatry. 2009;17(5):368–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Pedraza O, Clark JH, O'Bryant SE, et al. Diagnostic validity of age and education corrections for the Mini-Mental State Examination in older African Americans. J Am Geriatr Soc. 2012;60(2):328–331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Yesavage JA, Brink TL, Rose TL, et al. Development and validation of a geriatric depression screening scale: a preliminary report. J Psychiatr Res. 1982;17(1):37–49. [DOI] [PubMed] [Google Scholar]
- 56. Lawton MP, Brody EM. Assessment of older people: self-maintaining and instrumental activities of daily living. Gerontologist. 1969;9(3):179–186. [PubMed] [Google Scholar]
- 57. Behrens TE, Woolrich MW, Jenkinson M, et al. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magn Reson Med. 2003;50(5):1077–1088. [DOI] [PubMed] [Google Scholar]
- 58. Smith SM, Jenkinson M, Johansen-Berg H, et al. Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. Neuroimage. 2006;31(4):1487–1505. [DOI] [PubMed] [Google Scholar]
- 59. Smith SM, Jenkinson M, Woolrich MW, et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage. 2004;23(suppl 1):S208–S219. [DOI] [PubMed] [Google Scholar]
- 60. Leritz EC, Salat DH, Milberg WP, et al. Variation in blood pressure is associated with white matter microstructure but not cognition in African Americans. Neuropsychology. 2012;24(2):199–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Cleveland WS, Devlin SJ. Locally weighted regression: an approach to regression analysis by local ditting. J Am Stat Assoc. 1988;83(403):596–610. [Google Scholar]
- 62. Chua TC, Wen W, Slavin MJ, Sachdev PS. Diffusion tensor imaging in mild cognitive impairment and Alzheimer's disease: a review. Curr Opin Neurol. 2008;21(1):83–92. [DOI] [PubMed] [Google Scholar]
- 63. Aboitiz F, Scheibel AB, Fisher RS, Zaidel E. Fiber composition of the human corpus callosum. Brain Res. 1992;598(1-2):143–153. [DOI] [PubMed] [Google Scholar]
- 64. Hofer S, Frahm J. Topography of the human corpus callosum revisited-Comprehensive fiber tractography using diffusion tensor magnetic resonance imaging. NeuroImage. 2006;32(3):989–994. [DOI] [PubMed] [Google Scholar]
- 65. de Lacoste MC, Kirkpatrick JB, Ross ED. Topography of the human corpus callosum. J Neuropathol Exp Neurol. 1985;44(6):578–591. [DOI] [PubMed] [Google Scholar]
- 66. Harsan LA, Poulet P, Guignard B, et al. Brain dysmyelination and recovery assessment by noninvasive in vivo diffusion tensor magnetic resonance imaging. J Neurosci Res. 2006;83(3):392–402. doi:10.1002/jnr.20742. [DOI] [PubMed] [Google Scholar]
- 67. Madler B, Drabycz SA, Kolind SH, Whittall KP, MacKay AL. Is diffusion anisotropy an accurate monitor of myelination? Correlation of multicomponent T2 relaxation and diffusion tensor anisotropy in human brain. Magn Reson Imaging. 2008;26(7):874–888. doi:10.1016/j.mri.2008.01.047. [DOI] [PubMed] [Google Scholar]
- 68. Song SK, Sun SW, Ramsbottom MJ, Chang C, Russell J, Cross AH. Dysmyelination revealed through MRI as increased radial (but unchanged axial) diffusion of water. NeuroImage. 2002;17(3):1429–1436. [DOI] [PubMed] [Google Scholar]
- 69. Fischer P, Jungwirth S, Zehetmayer S, et al. Conversion from subtypes of mild cognitive impairment to Alzheimer dementia. Neurology. 2007;68(4):288–291. [DOI] [PubMed] [Google Scholar]
- 70. Rountree SD, Waring SC, Chan WC, Lupo PJ, Darby EJ, Doody RS. Importance of subtle amnestic and nonamnestic deficits in mild cognitive impairment: prognosis and conversion to dementia. Dement Geriatric Cogn Disord. 2007;24(6):476–482. [DOI] [PubMed] [Google Scholar]
- 71. Schneider JA, Arvanitakis Z, Leurgans SE, Bennett DA. The neuropathology of probable Alzheimer disease and mild cognitive impairment. Ann Neurol. 2009;66(2):200–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Bartzokis G, Lu PH, Mintz J. Quantifying age-related myelin breakdown with MRI: novel therapeutic targets for preventing cognitive decline and Alzheimer's disease. J Alzheimers Dis. 2004;6(6 suppl):S53–S539. [DOI] [PubMed] [Google Scholar]
- 73. Marner L, Nyengaard JR, Tang Y, Pakkenberg B. Marked loss of myelinated nerve fibers in the human brain with age. J Comp Neurol. 2003;462(2):144–152. doi:10.1002/cne.10714. [DOI] [PubMed] [Google Scholar]
- 74. Sandell JH, Peters A. Effects of age on nerve fibers in the rhesus monkey optic nerve. J Comp Neurol. 2001;429(4):541–553. [DOI] [PubMed] [Google Scholar]