

Abstract
Marine mammals have recently been documented as important facilitators of rapid and efficient nutrient recycling in coastal and offshore waters. Whales enhance phytoplankton nutrition by releasing fecal plumes near the surface after feeding and by migrating from highly productive, high-latitude feeding areas to low-latitude nutrient-poor calving areas. In this study, we measured NH4+ and PO43- release rates from the feces of North Atlantic right whales (Eubalaena glacialis), a highly endangered baleen whale. Samples for this species were primarily collected by locating aggregations of whales in surface-active groups (SAGs), which typically consist of a central female surrounded by males competing for sexual activity. When freshly collected feces were incubated in seawater, high initial rates of N release were generally observed, which decreased to near zero within 24 hours of sampling, a pattern that is consistent with the active role of gut microflora on fecal particles. We estimate that at least 10% of particulate N in whale feces becomes available as NH4+ within 24 hours of defecation. Phosphorous was also abundant in fecal samples: initial release rates of PO43- were higher than for NH4+, yielding low N/P nutrient ratios over the course of our experiments. The rate of PO43- release was thus more than sufficient to preclude the possibility that nitrogenous nutrients supplied by whales would lead to phytoplankton production limited by P availability. Phytoplankton growth experiments indicated that NH4+ released from whale feces enhance productivity, as would be expected, with no evidence that fecal metabolites suppress growth. Although North Atlantic right whales are currently rare (approximately 450 individuals), they once numbered about 14,000 and likely played a substantial role in recycling nutrients in areas where they gathered to feed and mate. Even though the NH4+ released from fresh whale fecal material is a small fraction of total whale fecal nitrogen, and recognizing the fact that the additional nitrogen released in whale urine would be difficult to measure in a field study, the results of this study support the idea that the distinctive isotopic signature of the released NH4+ could be used to provide a conservative estimate of the contribution of the whale pump to primary productivity in coastal regions where whales congregate.
Introduction
It is well established that microbes, zooplankton, and fish are important sources of recycled nitrogen and other nutrients in coastal waters [1, 2]. Recent studies have documented that whales and other air-breathing vertebrates also contribute to primary production through the vertical mixing, horizontal transfer, and recycling of limiting nutrients [3, 4]. Quantitative assessments of these mechanisms make it increasingly clear that marine mammals can provide an important ecosystem service by sustaining productivity in regions where they occur in high densities [5, 6]. Yet the relative importance of the great whales—a group that includes baleen whales and sperm whales—for nutrient recycling in the coastal ocean has been greatly reduced by human activities including commercial hunting and other activities that have increased nutrient release into this environment, such as sewage, atmospheric deposition, and agricultural runoff [4].
In this study, we examined the potential for North Atlantic right whales (Eubalaena glacialis) to pump nutrients to the euphotic zone for phytoplankton production in the Bay of Fundy, a tidally mixed basin. The right whale generally feeds on late-stage copepods (Calanus finmarchicus) during the summer and fall in this area [7, 8]. The bulk of the C. finmarchicus population is in diapause (stage V) at this time, remaining at depths below 100 m of the water column, with actively feeding copepods migrating vertically in the upper 100 m of the water column [7]. Right whales feed in high densities within a relatively well defined area of the bay, approximately 24 x 30 km [9] leaving visible orange-brown feces at the surface that can also be detected by smell [10]. Right whale fecal material is typically clumped, reaching lengths of up to 15 cm. It often floats at the surface, with sufficient cohesion to be collected intact using a dip net.
Surface-active groups, or SAGs, are the most commonly observed surface social behavior of North Atlantic right whales [11]. Typically characterized by a vocalizing, or focal, female, with one or more males socializing at the surface, SAGs present a unique opportunity for the production, distribution, and collection of feces for analysis (Fig 1) [12, 13]. These groups appear to be engaged in courtship, with the focal female swimming on her back, presumably to avoid copulation, and males jostling for this opportunity when she rolls over to breathe. SAGs can last for hours. In addition to these potentially conceptive groups, right whales commonly form SAGs throughout the year, in groups that could have other potential functions such as play, mating practice, or the maintenance of social bonds [13]. During this rigorous activity, feces can readily disassociate into fine particles from the turbulence caused by the interacting whales. Although feces can be observed at the surface while right whales are feeding, SAGs present a convenient opportunity for fecal collection and can function as “hotspots” for the release of nutrients in surface waters.
Fig 1. A surface-active group of at least five North Atlantic right whales in the Bay of Fundy.

Fecal material is suspended in the surface water. Photo courtesy of New England Aquarium.
Several rorqual whales, distinguished by longitudinal folds of skin below their mouth, also occur in the region, including humpback (Megaptera novaeangliae), sei (Balaenoptera borealis), and fin (Balaenoptera physalus) whales. These rorquals feed on zooplankton (typically krill), schooling fish, and occasionally cephalopods. They release feces that can be clumped and positively buoyant when feeding on crustaceans, but tend to be more dispersed, and plume-like when they are consuming fish. These fine particles are readily dispersed by physical mixing.
To examine the ecological role of whales in the Bay of Fundy, we analyzed fecal samples from right whales, plus an occasional humpback, sei, and fin whale, for NH4+ and PO4-3 production rates and isotope ratios. From our previous work with humpback whales on Stellwagen Bank [4], we expected to find elevated levels of NH4+ associated with the right whale fecal material in the Bay of Fundy. Ammonium is of particular ecological significance since it is rapidly and efficiently utilized for phytoplankton growth, resulting in preferential uptake of this nitrogenous nutrient [14]. Phosphorus is essential for the structural and functional components of all organisms; recent studies have found high levels of P in whale feces [15], suggesting that whales may play an important role in distributing these nutrients to surface waters [16]. Micronutrients, such as iron and manganese, have also been found in whale feces [15], though we did not examine them here.
Over the past three decades, field and laboratory studies have revealed substantial new understandings related to isotopic fractionation, or the relative abundance of stable isotopes, in the marine nitrogen cycle; the application and utility of this approach in studies of marine ecosystems and biogeochemical cycles has been reviewed in detail [17, 18]. Analyses for δ15N of the fecal N and released NH4+ allowed us to explore the utility of this approach in assessing the contribution of right whales to nutrient availability in its feeding grounds.
Materials and Methods
Field sampling was conducted between August 13 and September 2, 2011, under the aegis of the Grand Manan Whale and Seabird Research Station (GMWSRS), Grand Manan Island, New Brunswick, Canada. The research station provided laboratory facilities and an outboard motor boat for sample collection in nearby whale feeding areas.
Sampling
Inshore waters in Grand Manan Basin (~200 m average depth) to the east of Grand Manan Island were sampled from a small boat (Fig 2). Right whales were located by running search patterns augmented by information sharing with researchers from the New England Aquarium (NEA) who were also working in this area. Whales were most often encountered in surface-active groups (SAGs of 5 to 25 individuals), which remained at or very near the sea surface for several hours at a time. Once located, the groups were approached to a distance of about 50 to 100 m, and as the group activity drifted away from its initial location, the boat moved into the area for water sampling and fecal material collection. Turbulence in these sample areas was visible for several minutes after the whales’ departure, and flocculent plumes and clumps of feces were captured. Given the depth of the water column in the area, which is typically more than 100 m, we do not expect that surface-active whales stirred up bottom sediment prior to our sampling.
Fig 2. Whale fecal sampling locations in the Bay of Fundy.
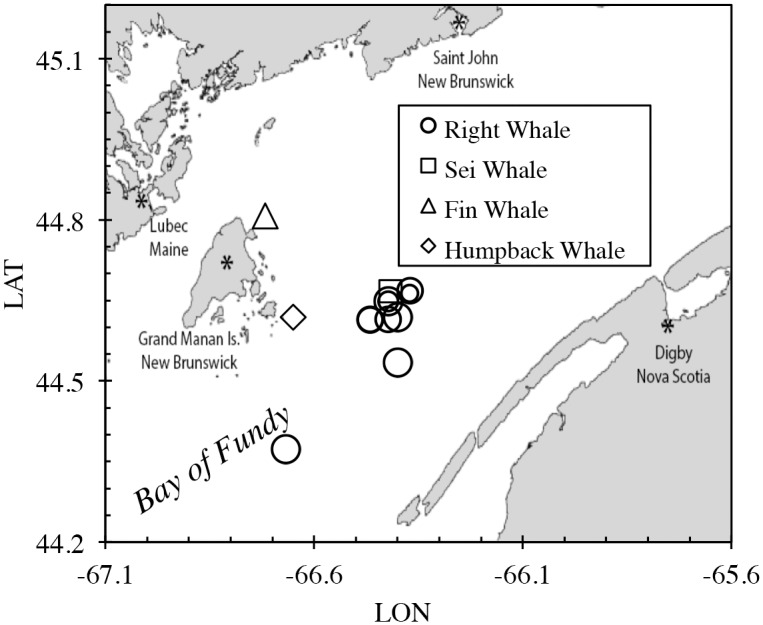
Sampling methods followed the protocols described by Roman and McCarthy [4].
We collected fecal suspensions with a 30-cm-diameter, 150-μm mesh plankton net attached to a 2-m pole. Samples were also collected at the waterline with a wide-mouth plastic bottle and in some cases by scooping globular feces from the surface, which were resuspended in surface seawater. Seawater control samples were collected several km away from recent whale sightings. All sample bottles were immediately placed in an ice chest containing frozen gel blocks and returned to the shore lab as soon as was practicable.
In addition to the right whale samples described above, feces of humpback, fin, and sei whales were collected by New England Aquarium and GMWSRS colleagues working nearby and using similar methods. Colleagues at the University of Alaska Southeast provided archived frozen fecal samples from humpback whales.
Shore-lab processing of fecal suspensions
Fecal samples from the Bay of Fundy were processed at the Grand Manan Whale and Seabird Research Station as soon as they were brought to shore, generally between 5 and 7 hours from the time of collection. Samples were prepared for analyses of phytoplankton nutrient concentrations (NO3-, NH4+, and PO43-), particulate organic nitrogen (PON) concentration, and N stable-isotope ratios for PON and NH4+. Frozen archived samples were resuspended in fresh ambient seawater collected away from whales and analyzed without incubation.
PON concentration and 15N natural abundance (δ15N)
Replicate aliquots of 10–50 ml (depending on visual assessment of particulate density in the sample) were filtered through combusted 25 mm Whatman GF/F glass fiber filters. The filters were then transferred to glass shell vials and placed in a 60°C drying oven for 24 h before capping and storing for later analysis of PON and δ15N on a Europa Scientific model 20/20 continuous flow, elemental analyzer/stable isotope ratio mass spectrometer system at Harvard University [19, 20].
δ15N NH4+ isotope ratio analysis
Aliquots were filtered through combusted 47mm GF/F filters to obtain 200ml of filtrate from each sample. NH4+ isotopic composition (δ15N) was measured according to a modified version of Zhang et al. [21]. Briefly, NH4+ is oxidized to nitrite using hypobromite and then reduced to N2O using acetic acid–buffered sodium azide. Isotope determinations were made at the University of Massachusetts Dartmouth using a GV IsoPrime IRMS, a custom purge-trap sample preparation system, and a CTC PAL autosampler. Reference materials used for calibration were IAEA-N1 and IAEA-N2 and values are reported relative to AIR with reproducibility better than ± 0.5%.
Nutrient analysis
Filtrate from each sample (~180 ml) was preserved by acidification to pH 2–3 with HCL for subsequent analysis of [NO3-], [NH4+] and [PO43-] on a robotic Westco SmartChem® 200 autoanalyzer (±0.05 to 0.2 μmol l−1 instrument precision depending on analyte), using standard colorimetric chemistries [22] at the Altabet Lab, UMass Dartmouth.
Incubation of fecal suspensions
After the initial samplings described above, the bottles containing fecal suspensions from the Bay of Fundy were placed in a dark temperature-controlled water bath, maintained at the temperature (±1°C) of the surface seawater measured at the collection sites. Samples were drawn at ~12 h intervals to obtain time courses of up to ~80 h from the incubated fecal suspensions and analyzed for particulate and dissolved constituents as described above.
Phytoplankton uptake of whale-derived fecal NH4+
Nitrogen-uptake experiments [23] were conducted to determine if NH4+ released from whale feces would enhance phytoplankton growth. Ambient surface seawater was collected in the Bay of Fundy at a distance of more than 1 km from recent whale activity (ambient NH4+ </ = 0.05 μmol kg-). Two 1-l aliquots of the single surface seawater sample were enriched to 8 μmol kg-1 NH4+ with right whale fecal matter filtrate from samples F1 and F13. A third aliquot without this addition was used as the untreated control. Since the NH4+ concentration was substantially elevated in the experiments with filtrate addition, we expected that the concentration of NH4+ would decline and the concentration of PON would rise relative to the control during the course of the experiment. Moreover, since the δ15N of the added NH4+ was substantially higher (~12‰) than the initial δ15N PON (~6‰), uptake of NH4+ into PON would result in an increase in δ15N PON. An increase in chlorophyll a (Chl a) commensurate with an increase in PON would allow us to infer that the uptake was from phytoplankton assimilation. Initial PON concentrations and δ15N were determined by filtering an aliquot from a control sample onto a GF/F filter as described above. A second aliquot was filtered and kept frozen for subsequent Chl a analysis at Harvard University [22]. (The concentration of Chl a was used as a proxy for phytoplankton biomass. No data are available for phytoplankton species abundances.)
The samples were incubated for 9.5 hours, beginning in midmorning, in a water bath under simulated in situ conditions [20, 24] at 17°C (±1) and with 36% ambient sunlight. The incubations were terminated by filtration, and the filters were dried and stored for later analysis.
Results
Dates, locations, and analytical results for samples collected in 2011, including concentrations of dissolved and particulate constituents of the whale fecal suspensions, are presented in Table 1. Values of δ15N for PON and NH4+ are similar across the range of right whale feces and the three additional species sampled. Concentrations of NH4+ and PO4-3 were orders of magnitude higher than typical values for coastal waters [25–27]. Dissolved NH4+:PO4-3 molar ratios in the right whale fecal suspensions at the initiation of the incubation experiments (Table 1) are quite constrained with a mean of 0.7 (Fig 3). Data for PON and δ15N in right and humpback fecal samples provided by the New England Aquarium and the University of Alaska Southeast are presented in Table 2. The four points clustered near the origin in Fig 3 all have higher PO4-3 than NH4+ concentrations. Given typical Redfield ratios of ~16 N/P for marine plankton and detritus, it is likely that the high PO4-3 concentrations at the initial sampling point reflect a more rapid release for P than for N from the whale fecal material during the time between defecation and the initiation of the incubation experiments on shore. Evidence for this can be seen in the rates of release for NH4+ and PO4-3 between the first and second sampling points during the right whale fecal suspension incubation experiments (Fig 4, showing samples with increased NH4+). For all but one datum point, the release rate for PO4-3 during this interval exceeded that for NH4+.
Table 1. Whale fecal material sampling details and analytical results for dissolved nutrients, PON concentration, and N stable isotope ratios for PON and NH4+ at the first time point, 5–7 h after sampling.
| Sample ID | Date | Lon (deg) | Lat (deg) | [PON] (μMol kg-1) | [NO3-] (μMol kg-1) | [PO43-] (μMol kg-1) | [NH4+] (μMol kg-1) | δ15N PON (‰) | δ15N NH4+ (‰) |
|---|---|---|---|---|---|---|---|---|---|
| F1-RW | 13 Aug 11 | -66.371 | 44.663 | 1161 | 1.91 | 151 | 70 | 9.5 | 12.3 |
| F5-RW | 13 Aug-11 | -66.370 | 44.668 | 206 | 0.32 | 440 | 586 | 9.6 | 13.1 |
| F8-RW | 23 Aug 11 | -66.422 | 44.648 | 2186 | 5.34 | 644 | 657 | 7.9 | 17.3 |
| F11-RW | 24-Aug-11 | -66.465 | 44.615 | 648 | 0.78 | 416 | 364 | 9.7 | 13.4 |
| F12-RW | 24-Aug-11 | -66.465 | 44.614 | 1489 | 0.79 | 534 | 500 | 9.4 | 13.7 |
| F13-RW | 24-Aug-11 | -66.399 | 44.534 | 547 | 0.90 | 128 | 86 | 8.8 | 12.4 |
| F14-RW | 24-Aug-11 | -66.399 | 44.618 | 218 | 0.62 | 74 | 65 | 8.4 | 13.6 |
| F15-RW | 30-Aug-11 | -66.668 | 44.373 | 1371 | 0.92 | * | 316 | 9.3 | 16.1 |
| F16-RW | 2-Sep-11 | -66.422 | 44.648 | 348 | 5.24 | 463 | 793 | 9.5 | 13.1 |
| Mean | 9.1 | 13.9 | |||||||
| SD | 0.62 | 1.6 | |||||||
| F7-HB | 17-Aug-11 | -66.649 | 44.663 | 146 | 6.13 | 61 | 8.8 | 9.0 | 9.0 |
| F6-FW | 13-Aug-11 | -66.717 | 44.807 | 856 | 1.01 | 526 | 67 | 7.1 | 15.7 |
| F18-SW | 18-Sep-11 | -66.417 | 44.667 | 416 | 3.59 | 479 | 737 | 9.8 | 12.0 |
RW is right whale, FW fin whale, HB humpback, SW sei whale.
* for no data.
Fig 3. Regression analysis of dissolved NH4+ and PO43- concentrations in whale feces filtrate at initial sampling.
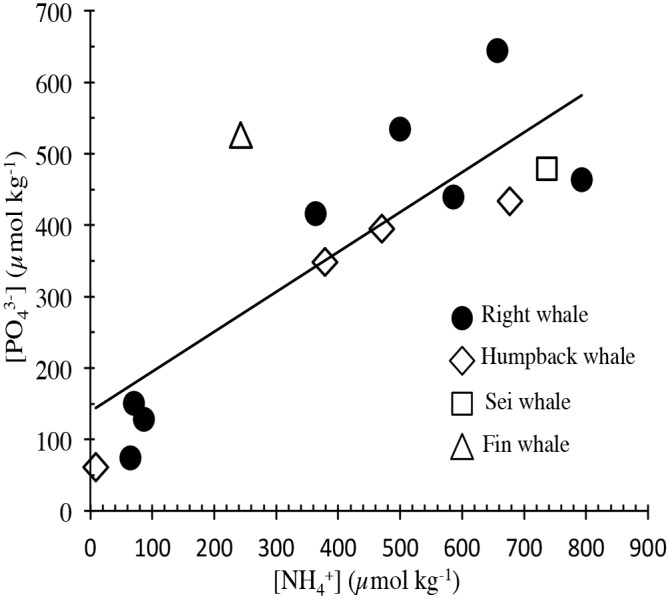
For all species combined, R2 = 0.73.
Table 2. N analyses on frozen archived samples provided by colleagues from the New England Aquarium (NEA), and the University of Alaska, Sitka (UAS).
BOF is Bay of Fundy; SeC Seymour Canal, AK; SiS Sitka Sound, AK;
| Sample ID | Source | Locale | Date | Lon* (deg) | Lat* (deg) | [PON] (μMol kg-1) | N % | δ15N PON (‰) | |
|---|---|---|---|---|---|---|---|---|---|
| RW1-08 | NEA | BOF | 8/08* | -66.7 | 44.4 | 146 | 6.2 | 13.4 | |
| RW2-08 | NEA | BOF | 8/08* | -66.7 | 44.4 | 856 | 7.5 | 12.5 | |
| RW3-08 | NEA | BOF | 8/08* | -66.7 | 44.4 | 416 | 8.0 | † | |
| F-19-HB | UAS | SeC | 10/12/08 | -134.1 | 57.8 | 326 | † | 9.5 | |
| F20-HB | UAS | SeC | 11/15/08 | -134.1 | 57.8 | 333 | 3.5 | 9.4 | |
| F-21-HB | UAS | SiS | 6/22/08 | -135.6 | 56.9 | 550 | † | 9.6 | |
*approx. date or location;
† no data.
Fig 4. Regression analysis of release rates for NH4+ and PO43- for right whale feces.

Rates were calculated for the period between the initial sampling and the first time point during the fecal suspension incubation experiments, R2 = 0.63.
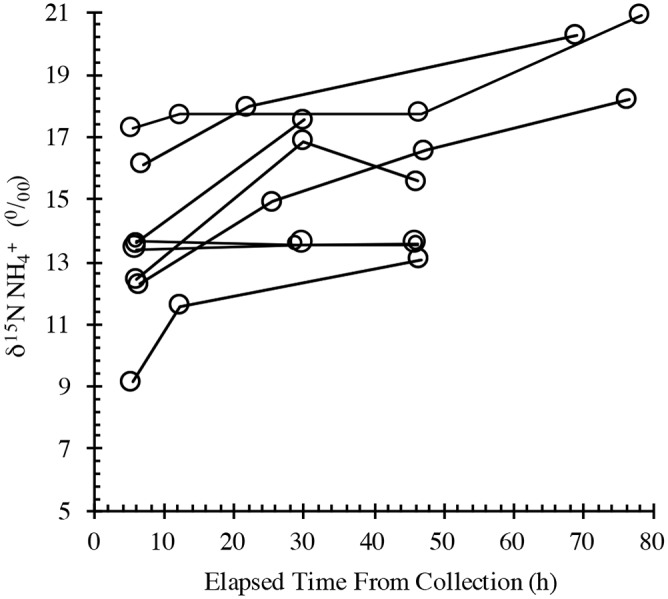
The NH4+ release experiments in this study were run longer than the ~24h time period used by Roman and McCarthy [4], who found that NH4+ generally increased to the end point of the incubation. To determine whether these release rates were sustained over a longer period, we extended the time courses to > 48h. Ammonium was again observed to increase in the whale fecal suspensions during the first 24h of incubation, but with only a few exceptions, NH4+ either leveled off or declined beyond 30 hours (Fig 5). The time course for δ15N of right whale fecal PON and the NH4+ that was released during the incubation experiments is shown in Fig 6. In comparison with the NH4+ concentration data, the NH4+ δ15N data trended toward higher values in more of the experiments after the 24-hr time point.
Fig 5. Time courses for NH4+ concentration in fecal suspensions from the Bay of Fundy right whale.

Fig 6. Time courses of δ15N NH4+ in fecal suspensions from the Bay of Fundy right whale.
Nutrient analyses on the filtrates from the whale-fecal incubations yielded undetectable concentrations of NO2-, and the concentrations of NO3- ranged from about 1 to 5 μmol kg-1, which are typical for the surface waters of the central basin during late summer [25–27]. Subsequent samplings from the same whale fecal incubations used for the NH4+ and PO43- release experiments revealed no measureable increase in NO2-, or NO3- concentrations (data not shown).
In the uptake studies, it was evident that NH4+ derived from whale feces could support phytoplankton growth unhindered to any obvious degree by other metabolites associated with the fecal material (Fig 7). Assimilation of NH4+ (and likely PO43-, though these rates were not measured) produced from this feces resulted in an approximate doubling of phytoplankton biomass as measured by PON and Chl a levels during the 10-hr experiment. (The control sample received no nutrient augmentation, and its Chl a concentration declined during the incubation.) As would be expected in a nutrient-enrichment experiment, the decrease in NH4+ concentrations, from about 8 to 4 μmol kg-1, approximately matched an increase in PON concentrations from about 4.5 to 8.5 μmol kg-1. Similarly, the Chl a concentration doubled from 0.6 to 1.3 μg l-1. These trends and the similar ratios of PON to Chl a concentrations, 6–7 μmol PON kg-1 per μg Chl a l-1, at the initiation and conclusion of this experiment, together suggest that the suspended PON consisted mostly of phytoplankton. The δ15N of PON increased as would be expected with uptake of the more highly 15N-enriched NH4+ derived from whale feces. The δ15N of residual NH4+ also increased, an expected result of the isotope fractionation associated with the preferential assimilation of the lighter N during phytoplankton uptake.
Fig 7. Uptake experiment using natural 15NH4+ tracer obtained from right whale fecal matter suspension filtrates.

Change in NH4+ concentration over incubation time (a); change in PON (b); change in Chl a (c), changes in δ15N for NH4+ and PON (d).
Discussion
Though long overlooked, the effect of marine mammals and other top predators on primary productivity and phytoplankton ecology is an emerging area of research [6, 15, 28]. It is interesting to note that both in our prior study, using humpback whales feeding on sand lance in Massachusetts Bay, and in this one, with right whales feeding on copepods, the yield of NH4+ as a function of fecal nitrogen concentration in our experiments is similar: ~4–5 nmol N (μmol fecal N)-1 hr-1. In other words about ten percent of the nitrogen in fresh feces is released as NH4+ day-1 (Fig 8). We would expect this NH4+ to diffuse and assimilate rapidly into the planktonic community of the euphotic zone. Whales can maintain prey aggregations by increasing nutrients in feeding areas when concentrations decline [4].
Fig 8. Rates of NH4+released vs. PON concentration for fecal suspensions.
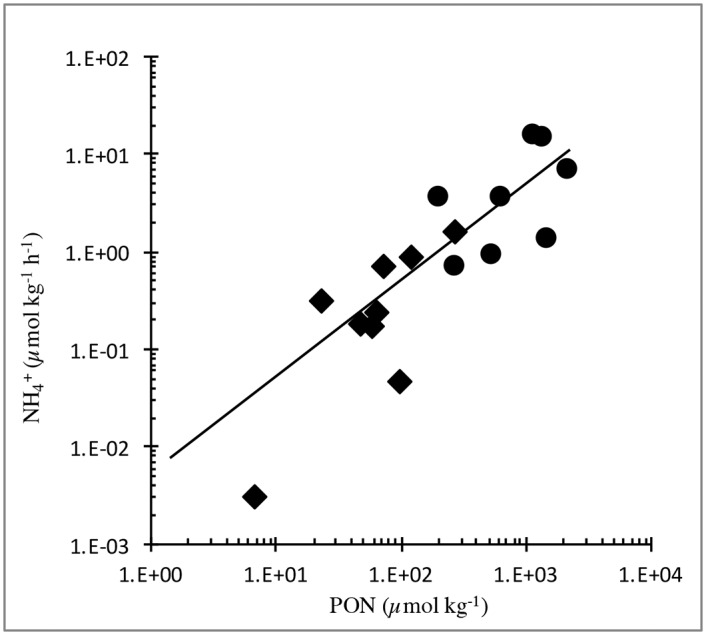
Data for initial ~20h of incubation, combining results for right whales (circles •, this study) and published data for humpback whales on Stellwagen Bank (diamonds ◆) [4].
Right whale fecal material was relatively 15N enriched with average δ15N values of ~ 9‰ (s.d = 0.6, n = 9; Table 1) as compared to the oceanic average near 5‰ [29] and surface PON observed in this study (~5.8‰; Fig 7d). This enrichment is likely due to whale feeding at higher trophic levels relative to phytoplankton as δ15N increases by 3 to 4‰ per trophic step [30]. Even considering, though the data are few, other whale species consuming different food in different regions, this degree of enrichment appears to hold (Table 1). It follows that NH4+ released from whale fecal material would be 15N enriched as confirmed by average values for the first time point of the release experiments of ~14‰ (s.d. = 1.6‰). Further 15N enrichment in the released NH4+, however, was not expected as fractionation results in isotope depletion of the product. Instead it would appear that NH4+ is preferentially released from components of the fecal material with higher than average δ15N for this material. Increasing δ15N for released NH4+ during incubation (Fig 6) suggests this process intensifies with time.
Regardless of its cause, the high δ15N of released NH4+ provides a strong signal for natural isotope tracing of the importance of this N source for phytoplankton. Qualitatively, uptake of 15N enriched NH4+ readily accounts for the ~2‰ increase in PON in δ15N during the course of the incubation experiment (Fig 7d). Mass balance calculation shows that the added PON had a δ15N of ~ 9‰ similar to the average δ15N of NH4+ for the first time point of the release experiments (Table 1). However, this value is about 2‰ lower than the initial δ15N NH4+ in the incubation experiments, a likely consequence of isotope fractionation during phytoplankton uptake [31]. Such fractionation is consistent with the increase in δ15N NH4+ as its concentration is drawn down during the incubation. Beyond confirming that NH4+ release from whale fecal material can support phytoplankton growth in controlled experiments, these results prove the feasibility of using this isotope signal in field studies designed to ascertain the contribution of nitrogen supplied from whale feces to local phytoplankton productivity.
Our findings support a previous study in the Southern Hemisphere showing high concentrations of phosphorous relative to carbon in whale feces [15] and expand upon this work in documenting high rates of PO43- release during incubations with whale fecal material. It is possible that other soluble bioactive constituents in these samples—such as Fe, other trace metals, or organic compounds—could have stimulated phytoplankton activity. Pygmy blue whale feces have been found to increase the photosynthetic rate and growth of three marine phytoplankton species [32]. In that study, feces were air dried and crushed into a fine powder before their use, whereas our study used filtrate from right whale fecal matter. As with the previous study, it is not surprising that added nutrients and micronutrients would stimulate phytoplankton growth. Just as important, however, we found no evidence that fecal metabolites inhibited growth.
Phytoplankton Chl a content can change in response to irradiance level, but the light used for these incubations (36% surface light) is unlikely to have promoted a strong adaptive response during the experiment. Likely explanations for the ~30% decline in Chl a in the control bottle include cell lyses and consumption by small microzooplankton grazers [33]. Although no copepods or other macroplankton were visibly observed in any of the bottles, no effort was made to physically exclude grazers from these incubation bottles. If microzooplankton grazing did cause a reduction of phytoplankton in the control experiments, we would also expect that they would have consumed Chl a in the enriched experiments. Excluding all microzooplankton from such experiments is impractical. Future experiments like these, however, could estimate grazing by microzooplankton using the Landry dilution technique [34].
Chl a effectively doubled in the augmented experimental bottle, or more than doubled when compared to the control bottle. The decrease in NH4+ during the incubation approximately balances the increase in PON, which would be consistent with assimilation of NH4+ by phytoplankton and retention in the particulate pool. Analyses were not made for PO43- or particulate organic phosphorous, but it is likely that the concentrations of dissolved and particulate phosphorus changed relative to one another, and consistent with phytoplankton assimilation, in a manner similar to nitrogen.
Filtration of the whale fecal material (GFF filter, nominal porosity ~0.7 μm) for this enrichment experiment would not have removed bacterioplankton. Thus, it is possible that some of the conversion of NH4+ to PON was attributable to free-living bacterioplankton that passed through the GFF filter and particle-associated bacteria in the original ocean water. To the degree that this occurred, it would have contributed to an elevated PON/Chl a ratio in enrichment experiments. Consumption of phytoplankton by microzooplankton would also be consistent with this observation.
Approximately 30% of Chl-a biomass sinks out of the euphotic zone in the Bay of Fundy [35], and the bulk sediment flux consists mostly of algal cells and fecal pellets. Curiously, this benthic community is relatively impoverished, with low rates of macrobenthic production. As right whales in the area have frequently been observed with mud on their heads and bodies while feeding [36], the bioturbation they facilitate could be another function that has come close to vanishing with the near extinction of these whales.
Our study complements recent efforts in the Southern Hemisphere and in the southern Gulf of Maine to quantify the impact of great whales on marine ecosystems [3, 4, 6]. In the Southern Ocean, the iron content of whale feces is orders of magnitude higher than the background levels of Antarctic seawater [5]. Such high levels of iron in the surface layer could act as a fertilizer, and the krill-baleen whale system likely played a crucial role in the iron cycle of the Southern Hemisphere, especially before commercial hunting depleted many populations of whales.
In the North Atlantic, right whales appeared to be close to extinction since the 1930s [37]. Spatially explicit estimates suggest that there were 9,075 to 21,328 right whales in the North Atlantic before commercial whaling [38]. At that time, foraging whales and surface-active groups would have been more common, and perhaps larger in size and more extensive in duration—field observations indicate that the longer a SAG remains active, the larger it tends to be, as more whales approach the group over time [12]. This spatial and temporal cohesion suggests that SAGs can act like whale pump hotspots. Because these groups are highly visible and dynamic, they can also be useful for researchers. Researchers in the Auckland Islands relied on breeding aggregations to collect feces from southern right whales for micronutrient analysis [39], and earlier studies in the Bay of Fundy used samples collected during right whale courtship activity or when whales fluked up to dive [40].
The right whale population in the northwest Atlantic is comprised of approximately 450 individuals, and right whales are nearly extinct in the eastern North Atlantic. Naturally, their ecological impact has been greatly reduced since their populations were reduced from possibly more than 20,000. Before whaling and the rise in atmospheric deposition, right whales and other mysticetes played an important role in enhancing nitrogen and phosphorous on a local and global scale [4, 16]. Production of phytoplankton stocks that support copepods and other prey consumed by whales will benefit most immediately from the release of excreta in nutrient-limited waters, especially in the summer [4].
If efforts to restore populations of this once economically and ecologically important species are successful, the right whale’s influence on ecosystem processes might increase throughout its range: not only on its summer feeding areas of the northern Gulf of Maine and southwestern Scotian Shelf, but also in the winter calving grounds off Florida and Georgia and spring feeding areas along the Great South Channel and Cape Cod Bay. As numbers rise, we are sure to learn new ecological roles for this once common species. As consumers of copepods and other phytoplankton grazers, right whales could reduce grazing pressure on planktonic microalgae while adding nutrients through the whale pump; such a tritrophic mutualism has been proposed for seabirds (such as albatrosses, petrels, and shearwaters) and phytoplankton in the Southern Hemisphere [28]. Understanding the foraging behavior of right whales will not only help inform management of this endangered whale but also increase our understanding of vertical and horizontal nutrient transfer by this species.
It should be noted that major environmental changes appear to be underway in our study area. Sea surface temperatures in the Gulf of Maine have increased faster than 99% of the global ocean [41], and data collected by tags on basking sharks in 2012, the year following our study, indicated that the Bay of Fundy has undergone rapid subsurface warming [42]. There have been fewer right whales observed in the bay in recent years, perhaps a result of this temperature shift and changes in primary productivity and copepod abundances. Whether right whales will continue to thrive in this area, or discover new feeding grounds, remains to be seen.
Acknowledgments
We thank Annabel Beichman, Andrew Westgate, Laurie Murison, Jennifer Larkum, Jared Juckiewicz, and Zach Siders for support in collecting samples in the Bay of Fundy. Samples were collected under Fisheries and Oceans Canada license #325842. Jan Straley and Roz Rolland supplied additional fecal samples.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by the U.S. Marine Mammal Commission and the Museum of Comparative Zoology, Harvard University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. It was also supported by a Sarah and Daniel Hrdy Fellowship in Conservation Biology at Harvard University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. http://www.oeb.harvard.edu/employment/hrdy.html). http://www.mcz.harvard.edu/).
References
- 1.Longhurst AR, Harrison WG. The biological pump: Profiles of plankton production and consumption in the upper ocean. Prog Oceanogr. 1989;22:47–123. [Google Scholar]
- 2.Turner JT. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog Oceanogr. 2015;130:205–248. [Google Scholar]
- 3.Lavery TJ, Roudnew B, Seymour J, Mitchell JM, Smetacek V, Nicol S. Whales sustain fisheries: Blue whales stimulate primary production in the Southern Ocean. Mar Mamm Sci. 2014. 10.1111/mms.12108 [DOI] [Google Scholar]
- 4.Roman J, McCarthy JJ. The whale pump: marine mammals enhance primary productivity in a coastal basin. PLoS ONE. 2010;5:e13255 10.1371/journal.pone.0013255 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nicol S, Bowie A, Jarman S, Lannuzel D, Meiners KM, van der Merwe P. Southern Ocean iron fertilization by baleen whales and Antarctic krill. Fish Fish. 2010;11:203–209. 10.1111/j.1467-2979.2010.00356.x [DOI] [Google Scholar]
- 6.Roman J, Estes J, Morrisette L, Smith C, Costa D, McCarthy J, et al. Whales as marine ecosystem engineers. Front Ecol Environ. 2014: 10.1890/130220 [DOI] [Google Scholar]
- 7.Baumgartner MF, Cole TVN, Campbell RG, Teegraden GJ, Durbin EG. Associations between North Atlantic right whales and their prey, Calanus finmarchicus, over diel and tidal time scales. Mar Ecol Prog Ser. 2003;264:155–166. [Google Scholar]
- 8.Murison LD, Gaskin DE. The distribution of right whales and zooplankton in the Bay of Fundy, Canada. Can J Zool. 1989;67:1411–1420. [Google Scholar]
- 9.Brown MW, Allen JM, Kraus SD. The designation of seasonal right whale conservation areas in the waters of Atlantic Canada In: Shackell NL, Willison JHM, editors. Marine Protected Areas and Sustainable Fisheries. Montreal: McGill-Queens University Press; 1995. p. 90–98. [Google Scholar]
- 10.Rolland RM, Hamilton PK, Kraus SD, Davenport B, Gillett RM, Wasser SK. Faecal sampling using detection dogs to study reproduction and health in North Atlantic right whales (Eubalaena glacialis). J Cetacean Res Manage. 2006;8:121–125. [Google Scholar]
- 11.Parks SE, Tyack PL. Sound production by North Atlantic right whales (Eubalaena glacialis) in surface-active groups. J Acoust Soc Am. 2005;117:3297–3306. [DOI] [PubMed] [Google Scholar]
- 12.Kraus SD, Hatch JJ. Mating strategies in the North Atlantic right whale (Eubalaena glacialis). J Cetacean Res Manag. 2001;2:237–244. [Google Scholar]
- 13.Parks SE, Brown MW, Conger LA, Hamilton PK, Knowlton AR, Kraus SD, et al. Occurrence, composition, and potential functions of North Atlantic right whale (Eubalaena glacialis) surface-active groups. Mar Mamm Sci. 2007;23(4):868–887. 10.1111/j.1748-7692.2007.00154.x [DOI] [Google Scholar]
- 14.McCarthy JJ. The kinetics of nutrient utilization. In Physiological bases of phytoplankton ecology. Can J Fish Aquat Sci Bull. 1981;210:211–233. [Google Scholar]
- 15.Ratnarajah L, Bowie AR, Lannuzel D, Meiners KM, Nicol S. The biogeochemical role of baleen whales and krill in Southern Ocean nutrient cycling. PLoS ONE. 2014;9(12):e114067 10.1371/journal.pone.0114067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Doughty CE, Roman J, Faurby S, Wolf A, Haque A, Bakker ES, et al. Global nutrient transport in a world of giants. Proc Natl Acad Sci. 2015;113:868–873. 10.1073/pnas.1502549112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Altabet MA. Isotopic tracers of the marine nitrogen cycle: Present and past In: Volkman J, editor. Marine Organic Matter: Chemical and Biological Markers The Handbook of Environmental Chemistry. Berlin: Springer-Verlag; 2006. p. 251–293. [Google Scholar]
- 18.Montoya JP. Natural abundance of 15N in marine planktonic ecosystems In: Michener R, Lajtha K, editors. Stable isotopes in ecology and environmental science, 2nd edition Malden, MA: Blackwell; 2008. p. 176–201. [Google Scholar]
- 19.Barrie A, Workman C. An automated analytical system for nutritional investigations using 15N tracers. Spectrosc Int J. 1984;3:439–447. [Google Scholar]
- 20.McCarthy JJ, Garside C, Nevins JL. Nitrogen dynamics during the Arabian Sea Northeast monsoon. Deep Sea Res Pt II. 1999;46(8–9):1623–1664. [Google Scholar]
- 21.Zhang L, Altabet MA, Wu T, Hadas O. Sensitive measurement of NH4+15N/14N (δ15N) at natural abundance levels in fresh and saltwaters. Anal Chem. 2007;79:5297–5303. [DOI] [PubMed] [Google Scholar]
- 22.Parsons TR, Maita Y, Lalli CM. A Manual of Chemical and Biological Methods for Seawater Analysis. NY: Pergamon Press; 1984. [Google Scholar]
- 23.McCarthy JJ, Yilmaz A, Coban-Yildiz Y, Nevins JL. Nitrogen cycling in the offshore waters of the Black Sea. Estuar Coast Shelf Sci. 2007;74:493–514. [Google Scholar]
- 24.Lohrenz SE, Wiesenburg DA, Rein CR, Arnone RA, Taylor CD, Knauer GA, et al. A comparison of in situ and simulated in situ methods for estimating oceanic primary production. J Plankton Res. 1992;14(2):201–221. [Google Scholar]
- 25.Martin JL, LeGresley MM, Strain PM. Plankton monitoring in the Western Isles region of the Bay of Fundy during 1999–2000. Canadian Technical Report of Fisheries and Aquatic Sciences, 2629. 2006.
- 26.Townsend DW, Pettigrew NR, Thomas AC. Offshore blooms of the red tide dinoflagellate, Alexandrium sp., in the Gulf of Maine. Cont Shelf Res. 2001;21:347–369. [Google Scholar]
- 27.Townsend DW, Pettigrew NR, Thomas AC. On the nature of Alexandrium fundyense blooms in the Gulf of Maine. Deep Sea Res Pt II. 2005;52:2603–2630. [Google Scholar]
- 28.Savoca MS, Nevitt GA. Evidence that dimethyl sulfide facilitates a tritrophic mutualism between marine primary producers and top predators. Proc Natl Acad Sci. 2014;111:4157–4161. 10.1073/pnas.1317120111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sigman DM, Altabet MA, Michener R, McCorkle DC, Fry B, Holmes RM. Natural abundance-level measurement of the nitrogen isotopic composition of oceanic nitrate: an adaptation of the ammonia diffusion method. Mar Chem. 1997;57:227–242. [Google Scholar]
- 30.Vander Zanden MJ, Cabana G, Rasmussen JB. Comparing trophic position of freshwater fish calculated using stable nitrogen isotope ratios (δ15N) and literature dietary data. Can J Fish Aquat Sci. 1997;54:1142–1158. [Google Scholar]
- 31.Waser NAD, Harrison PJ, Nielsen B, Calvert SE, Tirpin DH. Nitrogen isotope fractionation during the uptake and assimilation of nitrate, nitrite, ammonium, and urea by a marine diatom. Limnol Oceanogr. 1998;43:215–224. [Google Scholar]
- 32.Smith LV, McMinn A, Martin A, Nicol S, Bowie AR, Lannuzel D, et al. Preliminary investigation into the stimulation of phytoplankton photophysiology and growth by whale faeces. J Exp Mar Biol Ecol. 2013;446:1–9. [Google Scholar]
- 33.Calbet A, Landry MR. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol Oceanogr. 2004;49:51–57. [Google Scholar]
- 34.Latasa M, Landry MR, Schlüter L, Bidigare RR. Pigment specific growth and grazing rates of phytoplankton in the central equatorial Pacific. Limnol Oceanogr. 1997;42:289–298. [Google Scholar]
- 35.Emerson CW, Roff JC, Wildlish DJ. Pelagic-benthic energy coupling at the mouth of the Bay of Fundy. Ophelia. 1986;(26):165–180. [Google Scholar]
- 36.Kraus SD, Hamilton PK. Ocean floor encounters by right whales ehance risk of entanglement in fishing gear. In review. 2016.
- 37.Kraus SD, Brown MD, Caswell H, Clark CW, Fujiwara M, Hamilton PK, et al. North Atlantic right whales in crisis. Science. 2005;309:561–562. [DOI] [PubMed] [Google Scholar]
- 38.Monsarrat S, Pennino MG, Smith TD, Reeves RR, Meynard CN, Kaplan DM, et al. A spatially explicit estimate of the pre-whaling abundance of the endangered North Atlantic right whale. Conserv Biol. 2015; 10.1111/cobi.12664 [DOI] [PubMed] [Google Scholar]
- 39.Wing SR, Jack L, Shatova O, Leichter JJ, Barr D, Frew RD, et al. Seabirds and marine mammals redistribute bioavailable iron in the Southern Ocean. Mar Ecol Prog Ser. 2014;510:1–13. [Google Scholar]
- 40.Rolland RM, Hunt KE, Kraus SD, Wasser SK. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. Gen Comp Endocrinol. 2005;142:308–317. [DOI] [PubMed] [Google Scholar]
- 41.Pershing AJ, Alexander MA, Hernandez CM, Kerr LA, Le Bris A, Mills KE, et al. Slow adaptation in the face of rapid warming leads to collapse of the Gulf of Maine cod fishery. Science. 2015;350:809–812. 10.1126/science.aac9819 [DOI] [PubMed] [Google Scholar]
- 42.Koopman HN, Westgate AJ, Siders ZA, Cahoon LB. Rapid subsurface ocean warming in the Bay of Fundy as measured by free-swimming basking sharks. Oceanogr. 2014;27:14–16. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.