

Abstract
Staphylococcus aureus produces numerous factors that facilitate survival in the human host. S. aureus coagulase (Coa) and von Willebrand factor-binding protein (vWbp) are known to clot plasma through activation of prothrombin and conversion of fibrinogen to fibrin. In addition, S. aureus clumping factor A (ClfA) binds fibrinogen and contributes to platelet aggregation via a fibrinogen- or complement-dependent mechanism. Here, we evaluated the contribution of Coa, vWbp and ClfA to S. aureus pathogenesis in a rabbit model of skin and soft tissue infection. Compared to skin abscesses caused by the Newman wild-type strain, those caused by isogenic coa, vwb, or clfA deletion strains, or a strain deficient in coa and vwb, were significantly smaller following subcutaneous inoculation in rabbits. Unexpectedly, we found that fibrin deposition and abscess capsule formation appear to be independent of S. aureus coagulase activity in the rabbit infection model. Similarities notwithstanding, S. aureus strains deficient in coa and vwb elicited reduced levels of several proinflammatory molecules in human blood in vitro. Although a specific mechanism remains to be determined, we conclude that S. aureus Coa, vWbp and ClfA contribute to abscess formation in rabbits.
Introduction
Staphylococcus aureus remains one of the most prominent human bacterial pathogens worldwide [1, 2]. These Gram-positive cocci cause a wide clinical spectrum of disease and/or syndromes, including endocarditis, bacteremia, pneumonia, toxic shock syndrome, osteomyelitis, and skin and soft tissue infections (SSTIs) [3–5]. The remarkable success of S. aureus as a human pathogen is facilitated by its vast arsenal of virulence factors and an ability to acquire antibiotic resistance readily [5, 6].
Coagulase (Coa) is one of the earliest described virulence factors of S. aureus [7], and is routinely used as a diagnostic tool to differentiate between two major species of Staphylococcus, i.e., coagulase-positive (S. aureus) and coagulase-negative (e.g., S. epidermidis) organisms. Relatively recently, a second S. aureus coagulase was discovered and named von Willebrand factor-binding protein (vWbp) [8]. Coa and vWbp display sequence and structure homology, particularly at the N-terminus [9]. Both proteins insert N-terminal residues into the prothrombin zymogen cleft, which triggers non-proteolytic activation by conformational transformation and formation of a staphylothrombin complex [10, 11]. The C-terminal domain (substrate recognition domain) of coagulase binds fibrinogen, which is transformed into fibrin and subsequently forms a fibrin clot.
Fibrin deposition is a process critical to abscess formation and thereby contributes to host defense against invading S. aureus [12]. The S. aureus coagulases have been linked previously to abscess development in murine systemic [13] and subcutaneous models of infection [14]. Clumping factor A (ClfA), although not a coagulase, is a fibrinogen binding protein that can promote fibrinogen-dependent platelet aggregation and adherence of S. aureus to fibrin [15, 16]. Similar to the coagulases, a role for ClfA in S. aureus abscess formation has been demonstrated in murine models of S. aureus virulence [17–19].
Rabbit models of S. aureus infection were used historically to investigate virulence and host-pathogen interactions, but were replaced largely by mouse infection models. Although neither mouse nor rabbit innate immune systems faithfully recapitulate that of humans, there are characteristics of the rabbit innate immune system—especially those of granulocytes—that seem more closely aligned with those of humans by comparison. A role for coagulases and ClfA has not been reported in a rabbit model of S. aureus SSTI. To that end, we evaluated the role of S. aureus coa, vwb, and clfA in a rabbit skin abscess model.
Materials and Methods
Ethics statement
All animal studies and procedures were approved by the Animal Care and Use Committee at Rocky Mountain Laboratories, National Institute of Allergy and Infectious Diseases (NIAID) under protocols 2011–92 and 2012–027, and conformed to the guidelines of the National Institutes of Health (NIH).
Human venous blood was obtained from healthy donors according to a protocol approved by the Institutional Review Board for Human Subjects, NIAID, NIH. Studies were conducted according to the policies provided in the Declaration of Helsinki, and each volunteer provided written informed consent prior to participation in the study.
Bacterial strains and growth conditions
S. aureus Newman wild-type and isogenic coa (Δcoa), vwb (Δvwb), and coa/vwb (Δcoa/Δvwb) deletion strains, and a clfA transposon mutant strain (ΔclfA), were described previously [13, 20, 21]. Briefly, the pKOR1 allelic replacement system was used to create the S. aureus Δcoa, Δvwb and Δcoa/Δvwb deletion strains [13, 22], and the mariner-based bursa aurealis transposon system was used to construct the ΔclfA strain [21]. All S. aureus strains used for these studies have been phenotypically evaluated for their ability to clot whole blood [13]. We used S. aureus strains Newman, Δcoa, Δvwb, and Δcoa/Δvwb since they were readily available and used previously in murine models of S. aureus abscess formation [13, 21]. Bacteria were cultured in trypticase soy broth (TSB; Difco, Detroit, MI) at 37°C with constant shaking at 225 rpm. Overnight cultures were diluted 1:200 into fresh TSB and grown to early stationary (OD600 ~ 2.0) growth phase prior to use in assays.
Rabbit skin and soft tissue infection model
Animal experiments were performed as described [23]. Briefly, bacteria were cultured to early stationary phase of growth and then pelleted by centrifugation. Cells were washed twice with Dulbecco’s phosphate-buffered saline (DPBS; Sigma-Aldrich, St. Louis, MO) and suspended in sterile DPBS at 5 × 109 colony-forming units (CFU)/ml. The S. aureus dose used in this study was determined empirically in rabbits and results in reproducible abscesses that are easily evaluated by gross morphology [23]. S. aureus inocula were verified by enumeration of CFUs on trypticase soy agar plates. Five rabbits (NZW, strain Cr1c:KBL; Western Oregon Rabbit Company, Philomath, OR) were used for each group and each group was infected with a different S. aureus strain. Rabbits were anesthetized and subsequently inoculated with 100 μl of bacterial suspension into the left and right flank (5 rabbits for each strain and thus 10 abscesses per strain), and 100 μl of DPBS was injected into lower right flank for use as a sham infection control. Animals were monitored daily and allowed food and water ad libitum. S. aureus inflammatory lesions were measured daily for 14 days with a caliper as described previously [23]. Experiments were repeated twice using an additional set of two animals per strain to assess S. aureus abscess CFUs on day 2 post infection, and one animal per strain/day was used for histopathology analysis. Animals were humanely euthanized prior to tissue excision in accordance with protocol approved by the Institutional Animal Care and Use Committee.
Histopathology analysis
Abscesses with margins of surrounding tissue were excised and fixed in 10% neutral-buffered formalin for at least 48 hours and processed as described [24]. Tissues sections were stained with hematoxylin-eosin, Masson’s trichrome stain for capsule or Mallory’s phosphotungstic acid-hematoxylin for fibrin visualization [25]. Images of tissue sections were captured using an Olympus model BX-51 microscope and Olympus cellSens Dimension 1.13 software (Olympus, Center Valley, PA).
Quantitative analysis of molecules produced in human whole blood in response to S. aureus
Bacteria at mid-logarithmic growth phase were pelleted by centrifugation, washed twice with Dulbecco’s phosphate-buffered saline (DPBS; Gibco/Life Technologies, Grand Island, NY) and suspended in sterile RPMI 1640 medium buffered with 10 mM HEPES (RPMI/H; Invitrogen/Life Technologies, Grand Island, NY). Bacteria were added to heparinized human blood at a final concentration of 1 × 106 CFU/ml. A 1-ml sample of blood culture was taken immediately to serve as a time zero control and the remaining samples were incubated for 2 h at 37°C with gentle rotation. The blood-bacteria mixture was centrifuged at 1300 × g for 10 min at 25°C to collect plasma for analysis of inflammation molecules. Samples were stored at -80°C until shipped for analysis (Multi-Analyte Profiling (MAP) technology platform (HumanMap® v.2.0; Myriad RBM, Inc., Austin, TX). Data sets were analyzed using a one-way ANOVA and Tukey’s post-test. The complete results of the HumanMap analysis are provided in S1 Table.
Statistical analysis
All statistical analyses were performed using GraphPad Prism version 6.0 (GraphPad Software Inc., San Diego, CA). Data for abscess size were evaluated with a one-way ANOVA and Dunnett’s post-test to correct for multiple comparisons.
Results
SSTIs are among the most common manifestations of S. aureus disease. We previously developed a rabbit SSTI model to assess the relative contribution of USA300 virulence determinants to CA-MRSA pathogenesis [23]. Inasmuch as the S. aureus coagulases and ClfA contribute to fibrin deposition and are linked to abscess formation in murine infection models, we compared the ability of S. aureus Newman wild-type, Δcoa, Δvwb, Δcoa/Δvwb, and ΔclfA strains to cause abscesses in our rabbit SSTI model. Rabbits were infected by subcutaneous inoculation of S. aureus strains and abscess development (lesion size and assessment of gross morphology) was monitored daily for 14 days. All S. aureus strains tested caused formation of typical skin abscesses, as determined by gross morphology [23]. However, there were strain-dependent differences in abscess size (Fig 1A and 1B and S1 Fig). For example, abscesses caused by the Newman wild-type strain were significantly larger than those caused by Δcoa, Δvwb, or Δcoa/Δvwb strains on days 1, 2, 3, and 5 post-infection (P < 0.05, Fig 1A). Although abscesses caused by the ΔclfA strain were smaller than those caused by the wild-type strain (P < 0.05 on day 5), the difference was less pronounced compared to that of the coagulase negative strains (Fig 1A).
Fig 1. Contribution of S. aureus coa, vwb and clfA to formation of rabbit abscesses.
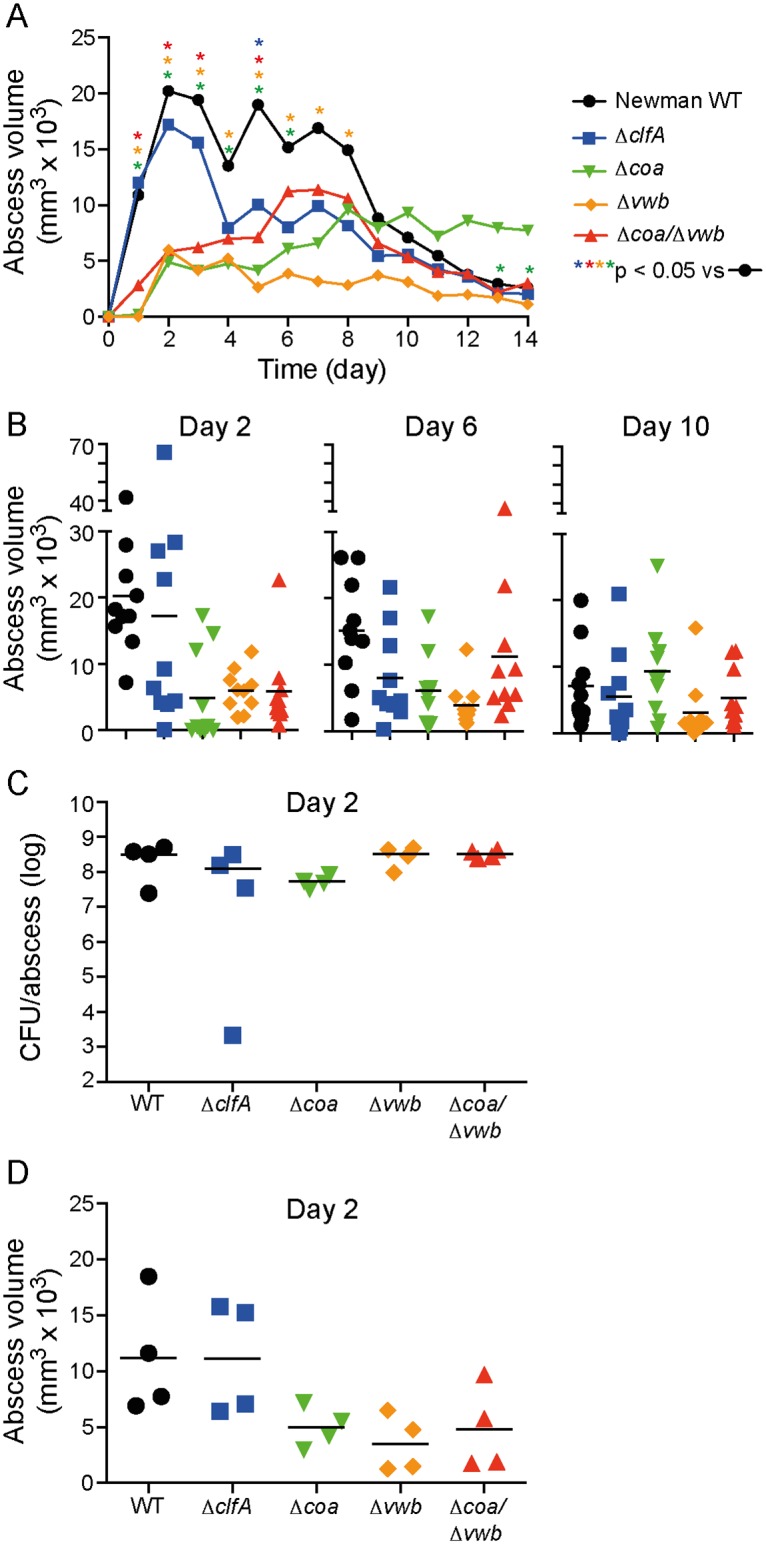
(A) Average abscess volume for rabbits infected subcutaneously with S. aureus Newman wild-type (WT) or isogenic mutant strains as indicated. The volume of 10 abscesses per bacterial strain was measured daily following inoculation. (B) Individual abscesses plotted for selected days are depicted in panel A. (C) A separate set of 2 animals (4 abscesses/strain) was used to determine S. aureus CFU per abscess on day 2 post-infection and (D) the volume of rabbit abscesses. Each symbol represents a data point obtained from a single abscess. P values were calculated using a one-way ANOVA and Dunnett’s post-test.
To determine if abscess size is associated with (or linked directly to) bacterial burden, we performed a second set of experiments to evaluate S. aureus CFUs in rabbit abscesses on day 2 following subcutaneous inoculation with each strain (Fig 1C and 1D). Unexpectedly, we found that CFUs per abscess were similar in all strains tested, indicating that the decreased abscess size (relative to wild-type) for the mutant strains was not due to a decrease in viability or more rapid bacterial clearance in this model. These findings contrast with those reported previously for ΔclfA [26, 27], Δvwb and Δcoa strains [13, 28] in murine abscess models of S. aureus infection. It is possible differences in animal species (mouse versus rabbit) and infection models employed account for the differences in results with bacterial burden.
Inasmuch as S. aureus coagulases and ClfA are involved in fibrin deposition, and since there were no apparent differences in bacterial numbers within abscesses, we next examined histological sections to ascertain differences in abscess fine structure (Fig 2 and Table 1). Abscesses were surgically excised on days 2, 6, and 10 following s.c. inoculation with S. aureus, and histopathology sections were processed and scored (Table 1). We found that all S. aureus strains tested caused formation of structurally discrete abscesses surrounded by a fully developed fibrous capsule by day 10, and that there were limited differences revealed by abscess histopathology—regardless of the S. aureus strain used for infection (Fig 2 and Table 1). Moreover, our data indicate that deposition of fibrin during abscess formation was mostly independent of Coa, vWbp and ClfA activity. Nonetheless, abscesses induced by ΔclfA had relatively weak fibrin deposition that was apparent only in abscesses excised on Day 2 and 10 (Fig 3). These data correspond with those in the mouse SSTI model, where fibrin deposition was apparent in abscesses induced by S. aureus when coagulase activity was blocked by dabigatran [14].
Fig 2. Histopathology of rabbit skin abscess caused by S. aureus.

Histopathology sections represent skin abscesses caused by S. aureus Newman WT (A, B), ΔclfA (C, D) or Δcoa/Δvwb (E, F) strains on day 10 post-infection. Abscess sections were stained with standard Masson’s trichrome stain to enhance fine structure detail of muscle tissues, collagen fibers and fibrin. (A, C and E) original magnification is 20×. (B, D, and F) 200× magnification of selected area (black rectangle) depicted in panels A, C or E.
Table 1. Contribution of coa, vwb and clfA to structure and development of the abscess.
| Strain | WT | ΔclfA | Δcoa | Δvwb | Δcoa/Δvwb | PBS ctrl | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days post-infection | d2 | d6 | d10 | d2 | d6 | d10 | d2 | d6 | d10 | d2 | d6 | d10 | d2 | d6 | d10 | d6 | d10 |
| Capsule granulation tissue | 0/2 | 2/2 | 2/2 | 0/2 | 2/2 | 2/2 | 0/2 | 2/2 | 0/2 | 0/2 | 2/2 | 2/2 | 0/2 | 1/2 | 1/2 | 0/2 | 0/2 |
| Fibrous capsule | 0/2 | 0/2 | 2/2 | 0/2 | 2/2 | 2/2 | 0/2 | 0/2 | 2/2 | 0/2 | 0/2 | 2/2 | 0/2 | 0/2 | 2/2 | 0/2 | 0/2 |
| Epithelialization | 0/2 | 2/2 | 1/2 | 0/2 | 0/2 | 1/2 | 0/2 | 1/2 | 0/2 | 0/2 | 1/2 | 0/2 | 0/2 | 0/2 | 1/2 | 0/2 | 0/2 |
| Vasculitis/vascular necrosis | 2/2 | 2/2 | 2/2 | 1/2 | 0/2 | 0/2 | 2/2 | 2/2 | 0/2 | 2/2 | 2/2 | 1/2 | 2/2 | 2/2 | 0/2 | 0/2 | 0/2 |
| Thrombosis | 2/2 | 2/2 | 1/2 | 0/2 | 0/2 | 0/2 | 2/2 | 2/2 | 0/2 | 1/2 | 2/2 | 0/2 | 2/2 | 2/2 | 0/2 | 0/2 | 0/2 |
| Coagulative necrosis | 2/2 | 2/2 | 1/2 | 0/2 | 0/2 | 2/2 | 2/2 | 1/2 | 1/2 | 1/2 | 2/2 | 0/2 | 0/2 | 2/2 | 0/2 | 0/2 | 0/2 |
| Extracellular bacteria | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 0/2 | 0/2 |
| Intracellular bacteria | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 1/2 | 1/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 0/2 | 0/2 |
| Compiled score | 10/16 | 14/16 | 13/16 | 5/16 | 8/16 | 11/16 | 9/16 | 11/16 | 7/16 | 8/16 | 13/16 | 9/16 | 8/16 | 11/16 | 8/16 | 0/16 | 0/16 |
Abscesses were scored based on the presence (1) or absence (0) of a chosen feature based on 6 histopathology sections of two abscesses (3 each) for each bacterial strain. 0/2 = present in neither abscess; 1/2 = present in 1 of 2 abscesses; 2/2 = present in both abscesses.
Fig 3. Fibrin deposition in rabbit skin abscess caused by S. aureus Newman.

Representative sections of rabbit skin abscesses on Day 2 (A, D, G, K, N), Day 6 (B, E, H, L, O) and Day 10 (C, F, J, M, P) post infection. Abscesses from rabbits infected with S. aureus Newman WT (A-C), ΔclfA (D-F), Δcoa (G-J), Δvwb (K-M) and Δcoa/Δvwb (N-P). Tissue sections were stained with Mallory’s phosphotungstic acid-hematoxylin stain for visualization of fibrin (black arrows). Magnification is 200×. Inset image is the abscess at 20× (black rectangle).
Abscesses caused by the Newman wild-type strain scored categorically highest on each day based on histopathology features, with few exceptions (Table 1). One notable distinction was that abscesses from rabbits infected with the ΔclfA strain had no evidence of thrombosis and weak vascular necrosis in the majority of sections analyzed. Although present at the earlier time points, vascular necrosis was also limited in tissue samples from abscess caused by coagulase deficient strains (Fig 4 and Table 1).
Fig 4. Vasculitis caused by S. aureus Newman WT.
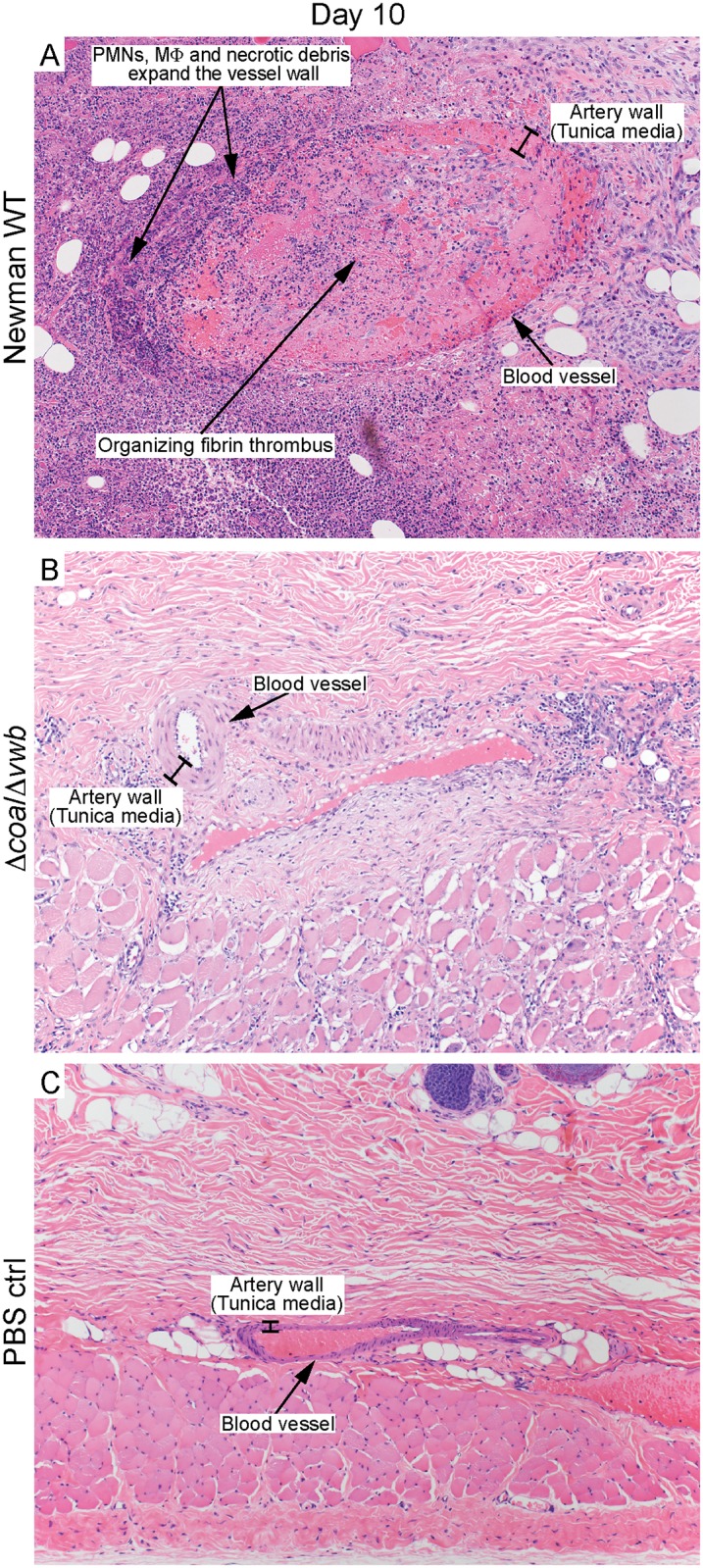
Histopathology sections of rabbit abscesses depicting vascular necrosis caused by S. aureus Newman WT (A), and an intact artery within a Δcoa/Δvwb induced abscess (B) or PBS control (C) on day 10 post infection. Original magnification is 100×.
The acute inflammatory response associated with S. aureus SSTI is triggered at least in part by production of pro-inflammatory signaling molecules and rapid recruitment of immune cells. To gain insight into the role of S. aureus coagulases and ClfA in the induction of host inflammation, we utilized a multi-analyte profiling approach to measure immune molecule production in human blood in response to S. aureus Newman wild-type, ΔclfA, Δcoa, Δvwb, and Δcoa/Δvwb. We performed the protein profiling experiments using human blood because there is paucity of reagents available to analyze a comprehensive panel of rabbit immune mediators. We also demonstrated previously that human and rabbit blood incubated with S. aureus have similar proinflammatory cytokine gene transcription profiles [29, 30]. As anticipated, several proinflammatory molecules were upregulated in blood samples incubated with S. aureus for 2 h compared to control blood lacking bacteria, including interleukin (IL)-8, myeloperoxidase (MPO), tumor necrosis factor (TNFα), and vascular endothelial growth factor (VEGF) (Fig 5 and S1 Table). There were also notable differences in levels of proinflammatory molecules elicited by S. aureus mutant and wild-type strains tested. For example, there was reduced expression of key proinflammatory mediators (IL-1β, MPO, PAI-1 and ENA-78) in human blood incubated with the S. aureus Δcoa/Δvwb strain compared to the wild-type Newman strain (Fig 5). If this phenomenon can be extended to host responses in tissues, it could provide an explanation in part for diminished pathology caused by the mutant strains in the rabbit SSTI model of infection, albeit this hypothesis requires further investigation.
Fig 5. S. aureus Newman causes increased production of proinflammatory molecules in human whole blood.

S. aureus was cultured in human heparinized blood for 2 h. Accumulation of proinflammatory molecules in plasma was evaluated by quantitative, multiplexed immunoassays (HumanMAP v2.0; Myriad RBM) as described in Materials and Methods section. Data represents average of 3 donors with one-way ANOVA and Tukey’s post-test used to determine statistical significance. *P < 0.05 for the selected pairs; # p < 0.05 compared to uninfected blood sample (ctrl).
Discussion
The rabbit is historically the classical animal model for studying S. aureus pathogenesis [31, 32] and has been used to model a diversity of diseases and syndromes such as endocarditis, pneumonia, sepsis, and toxemia [33–37]. We recently developed a rabbit model of skin and soft tissue infection [23] to study the contribution of S. aureus leukotoxins to abscess formation. In vitro studies indicate that susceptibility of rabbit cells to several S. aureus secreted leukotoxins and hemolysins approximates that of human cells more closely than those of murine origin [38, 39]. For example, mouse leukocytes are less susceptible (7–10 fold) to the cytolytic effects of PVL compared with human or rabbit leukocytes, and purified PVL has been tested directly in a rabbit skin infection model [40].
Inasmuch as S. aureus Coa, vWbp and ClfA are linked previously to abscess formation following murine systemic infection, we employed the rabbit skin and soft tissue infection model to assess the role of coagulase in development of subcutaneous abscesses. Our data indicate that all three of these molecules contribute to the formation of S. aureus abscesses in the experimental rabbit infection model. However, we found that the direct contribution of S. aureus Coa and vWbp to capsule formation and fibrin deposition was limited (Figs 2 and 3) compared to that reported for the S. aureus murine kidney model [13]. There are a couple of potential explanations for differences observed between the infection models. First, there are significant differences in the host proinflammatory response to invading pathogens between mice and rabbits, and as an example, IL-8 is a factor critical for neutrophil recruitment in humans and rabbits but is absent in mice [41, 42]. While it is evident that proinflammatory mediators play a critical role in formation of S. aureus abscesses [43–46], it is unlikely that species-specific production of proinflammatory molecules contribute to the differences reported for the role of coagulases on abscess structure between the models. On the other hand, it is possible that the role of S. aureus coagulases on abscess structure differs depending on the anatomical location of the abscess, rather than the animal species tested. Renal abscesses form as a result of systemic infection, during which disseminated bacteria within host blood accumulate in blood filtration organs such as the kidney or liver. S. aureus commonly accumulates in the renal arcuate arteries and causes infarcts [47, 48]. The combination of bacteria and tissue damage elicits neutrophil and other immune cell infiltration, and facilitates formation of a mature abscess. By contrast, invading S. aureus are recognized early during SSTI by local keratinocytes and resident skin monocytes, which initiate cytokine signaling to promote immune cell recruitment [49]. This triggers influx of neutrophils to the infection site to initiate the process of abscess formation [12, 50]. The influx of neutrophils also contributes to increased vascular permeability at the site of inflammation [51, 52]. Since coagulases and clumping factor A function primarily through binding or modifying fibrinogen—one of the most abundant plasma glycoproteins [53]—it is possible that limited access to fibrinogen in subcutaneous tissue reduces the role of coagulases and/or ClfA in formation of the SSTI abscess compared to the kidney. Indeed, consistent with our findings in rabbits, a S. aureus strain deficient for coa and vwb formed smaller subcutaneous abscesses in murine SSTI, and inhibition of the staphylothrombin complex by dabigatran treatment did not prevent deposition of fibrin and fibrinogen within the S. aureus wild-type abscess capsule [14]. However, in that study, abscess structure was not assessed directly by histopathology following infection with the S. aureus Δcoa/Δvwb deletion strain. Nevertheless, more work is needed to determine if there are variations in organ-specific immune response and/or bacterial response that may influence abscess development. Collectively, the data obtained from our rabbit infection model confirm previous findings that Coa, vWbp and ClfA are involved in the pathogenesis of S. aureus SSTI and contribute to the host proinflammatory response to infection.
Supporting Information
Scatter plot of abscess volumes from data shown in Fig 1A. Rabbits were infected subcutaneously with S. aureus Newman wild-type (WT) or isogenic mutant strains. The volume of 10 abscesses per bacterial strain was measured for 14 days following inoculation. Each symbol represents a data point obtained from a single abscess.
(TIF)
S. aureus was cultured in human heparinized blood up to 2 h. Accumulation of proinflammatory molecules in plasma was evaluated by quantitative, multiplexed immunoassays (HumanMAP v2.0; Myriad RBM) as described in Materials and Methods section. Data represents average of 3 donors ±SEM.
(DOCX)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
Support was provided by Intramural Research Program of the National Institute of Allergy and Infectious Diseases, National Institutes of Health.
References
- 1.Moran GJ, Krishnadasan A, Gorwitz RJ, Fosheim GE, McDougal LK, Carey RB, et al. Methicillin-resistant S. aureus infections among patients in the emergency department. N Engl J Med. 2006;355: 666–74. [DOI] [PubMed] [Google Scholar]
- 2.Boucher H, Miller LG, Razonable RR. Serious infections caused by methicillin-resistant Staphylococcus aureus. Clin Infect Dis. 2010;51: S183–S97. 10.1086/653519 [DOI] [PubMed] [Google Scholar]
- 3.Fridkin SK, Hageman JC, Morrison M, Sanza LT, Como-Sabetti K, Jernigan JA, et al. Methicillin-resistant Staphylococcus aureus disease in three communities. N Engl J Med. 2005;352: 1436–44. [DOI] [PubMed] [Google Scholar]
- 4.Landrum ML, Neumann C, Cook C, Chukwuma U, Ellis MW, Hospenthal DR, et al. Epidemiology of Staphylococcus aureus blood and skin and soft tissue infections in the US military health system, 2005–2010. JAMA. 2012;308: 50–9. 10.1001/jama.2012.7139 [DOI] [PubMed] [Google Scholar]
- 5.Lowy FD. Staphylococcus aureus Infections. N Engl J Med. 1998;339: 520–32. [DOI] [PubMed] [Google Scholar]
- 6.Kobayashi SD, Musser JM, DeLeo FR. Genomic analysis of the emergence of vancomycin-resistant Staphylococcus aureus. MBio. 2012;3: e00170–12. 10.1128/mBio.00170-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Loeb L. The influence of certain bacteria on the coagulation of the blood. J Med Res. 1903;10: 407–19. [PMC free article] [PubMed] [Google Scholar]
- 8.Bjerketorp J, Nilsson M, Ljungh Å, Flock J-I, Jacobsson K, Frykberg L. A novel von Willebrand factor binding protein expressed by Staphylococcus aureus. Microbiology. 2002;148: 2037–44. [DOI] [PubMed] [Google Scholar]
- 9.McAdow M, DeDent AC, Emolo C, Cheng AG, Kreiswirth BN, Missiakas DM, et al. Coagulases as determinants of protective immune responses against Staphylococcus aureus. Infect Immun. 2012;80: 3389–98. 10.1128/IAI.00562-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Friedrich R, Panizzi P, Fuentes-Prior P, Richter K, Verhamme I, Anderson PJ, et al. Staphylocoagulase is a prototype for the mechanism of cofactor-induced zymogen activation. Nature. 2003;425: 535–9. [DOI] [PubMed] [Google Scholar]
- 11.Kroh HK, Panizzi P, Bock PE. Von Willebrand factor-binding protein is a hysteretic conformational activator of prothrombin. Proc Natl Acad Sci U S A. 2009;106: 7786–91. 10.1073/pnas.0811750106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kobayashi SD, Malachowa N, DeLeo FR. Pathogenesis of Staphylococcus aureus abscesses. Am J Pathol. 2015;185: 1518–27. 10.1016/j.ajpath.2014.11.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cheng AG, McAdow M, Kim HK, Bae T, Missiakas DM, Schneewind O. Contribution of coagulases towards Staphylococcus aureus disease and protective immunity. PLoS Pathog. 2010;6: e1001036 10.1371/journal.ppat.1001036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vanassche T, Verhaegen J, Peetermans WE, Van Ryn J, Cheng A, Schneewind O, et al. Inhibition of staphylothrombin by dabigatran reduces Staphylococcus aureus virulence. J Thromb Haemost. 2011;9: 2436–46. 10.1111/j.1538-7836.2011.04529.x [DOI] [PubMed] [Google Scholar]
- 15.O’Brien L, Kerrigan SW, Kaw G, Hogan M, Penadés J, Litt D, et al. Multiple mechanisms for the activation of human platelet aggregation by Staphylococcus aureus: roles for the clumping factors ClfA and ClfB, the serine—aspartate repeat protein SdrE and protein A. Mol Microbiol. 2002;44: 1033–44. [DOI] [PubMed] [Google Scholar]
- 16.McDevitt D, Francois P, Vaudaux P, Foster TJ. Identification of the ligand-binding domain of the surface-located fibrinogen receptor (clumping factor) of Staphylococcus aureus. Mol Microbiol. 1995;16: 895–907. [DOI] [PubMed] [Google Scholar]
- 17.Hair PS, Echague CG, Sholl AM, Watkins JA, Geoghegan JA, Foster TJ, et al. Clumping factor A interaction with complement factor i increases C3b cleavage on the bacterial surface of Staphylococcus aureus and decreases complement-mediated phagocytosis. Infect Immun. 2010;78: 1717–27. 10.1128/IAI.01065-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hair PS, Ward MD, Semmes OJ, Foster TJ, Cunnion KM. Staphylococcus aureus clumping factor A binds to complement regulator factor I and increases factor I cleavage of C3b. J Infect Dis. 2008;198: 125–33. 10.1086/588825 [DOI] [PubMed] [Google Scholar]
- 19.Higgins J, Loughman A, Van Kessel KPM, Van Strijp JAG, Foster TJ. Clumping factor A of Staphylococcus aureus inhibits phagocytosis by human polymorphonuclear leucocytes. FEMS Microbiol Lett. 2006;258: 290–6. [DOI] [PubMed] [Google Scholar]
- 20.Baba T, Bae T, Schneewind O, Takeuchi F, Hiramatsu K. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: polymorphism and evolution of two major pathogenicity islands. J Bacteriol. 2008;190: 300–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.McAdow M, Kim HK, DeDent AC, Hendrickx APA, Schneewind O, Missiakas DM. Preventing Staphylococcus aureus sepsis through the inhibition of its agglutination in blood. PLoS Pathog. 2011;7: e1002307 10.1371/journal.ppat.1002307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bae T, Schneewind O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid. 2006;55: 58–63. [DOI] [PubMed] [Google Scholar]
- 23.Kobayashi SD, Malachowa N, Whitney AR, Braughton KR, Gardner DJ, Long D, et al. Comparative analysis of USA300 virulence determinants in a rabbit model of skin and soft tissue infection. J Infect Dis. 2011;204: 937–41. 10.1093/infdis/jir441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kennedy AD, Wardenburg JB, Gardner DJ, Long D, Whitney AR, Braughton KR, et al. Targeting of alpha-hemolysin by active or passive immunization decreases severity of USA300 skin infection in a mouse model. J Infect Dis. 2010;202: 1050–8. 10.1086/656043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Carson FL. Hladik C. Histotechnology: A self-instructional text. 3rd ed American Society for Clinical Pathology Press; 2009. [Google Scholar]
- 26.Cheng AG, Kim HK, Burts ML, Krausz T, Schneewind O, Missiakas DM. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 2009;23: 3393–404. 10.1096/fj.09-135467 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kwiecinski J, Jin T, Josefsson E. Surface proteins of Staphylococcus aureus play an important role in experimental skin infection. APMIS. 2014;122: 1240–50. 10.1111/apm.12295 [DOI] [PubMed] [Google Scholar]
- 28.Loof TG, Goldmann O, Naudin C, Mörgelin M, Neumann Y, Pils MC, et al. Staphylococcus aureus-induced clotting of plasma is an immune evasion mechanism for persistence within the fibrin network. Microbiology. 2015;161: 621–7. 10.1099/mic.0.000019 [DOI] [PubMed] [Google Scholar]
- 29.Malachowa N, Kobayashi SD, Sturdevant DE, Scott DP, DeLeo FR. Insights into the Staphylococcus aureus-host interface: global changes in host and pathogen gene expression in a rabbit skin infection model. PLoS ONE. 2015;10: e0117713 10.1371/journal.pone.0117713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Miller LS, Cho JS. Immunity against Staphylococcus aureus cutaneous infections. Nat Rev Immunol. 2011;11: 505–18. 10.1038/nri3010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rogers DE. Studies on bacteriemia: I. Mechanisms relating to the persistence of bacteriemia in rabbits following the intravenous injection of staphylococci. J Exp Med. 1956;103: 713–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rogers DE, Melly MA. Studies on bacteriemia: II. Further observations on the granulocytopenia induced by the intravenous injection of staphylococci. J Exp Med. 1957;105: 99–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cheung AL, Eberhardt KJ, Chung E, Yeaman MR, Sullam PM, Ramos M, et al. Diminished virulence of a sar-/agr- mutant of Staphylococcus aureus in the rabbit model of endocarditis. J Clin Invest. 1994;94: 1815–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lin Y-C, Anderson MJ, Kohler PL, Strandberg KL, Olson ME, Horswill AR, et al. Proinflammatory exoprotein characterization of toxic shock syndrome Staphylococcus aureus. Biochemistry. 2011;50: 7157–67. 10.1021/bi200435n [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Spaulding AR, Lin Y-C, Merriman JA, Brosnahan AJ, Peterson ML, Schlievert PM. Immunity to Staphylococcus aureus secreted proteins protects rabbits from serious illnesses. Vaccine. 2012;30: 5099–109. 10.1016/j.vaccine.2012.05.067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Strandberg KL, Rotschafer JH, Vetter SM, Buonpane RA, Kranz DM, Schlievert PM. Staphylococcal superantigens cause lethal pulmonary disease in rabbits. J Infect Dis. 2010;202: 1690–7. 10.1086/657156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Croisier-Bertin D, Piroth L, Charles P-E, Larribeau A, Biek D, Ge Y, et al. Ceftaroline versus ceftriaxone in a highly penicillin-resistant pneumococcal pneumonia rabbit model using simulated human dosing. Antimicrob Agents Chemother. 2011;55: 3557–63. 10.1128/AAC.01773-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bernheimer AW. Staphylococcal alpha toxin. Ann N Y Acad Sci. 1965;128: 112–23. [DOI] [PubMed] [Google Scholar]
- 39.Wiseman GM. The hemolysins of Staphylococcus aureus. Bacteriol Rev. 1975;39: 317–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cribier B, Prévost G, Couppie P, Finck-Barbançon V, Grosshans E, Piémont Y. Staphylococcus aureus leukocidin: A new virulence factor in cutaneous infections? An epidemiological and experimental study. Dermatology. 1992;185:175–80. [DOI] [PubMed] [Google Scholar]
- 41.Neves F, Abrantes J, Almeida T, de Matos AL, Costa PP, Esteves PJ. Genetic characterization of interleukins (IL-1α, IL-1β, IL-2, IL-4, IL-8, IL-10, IL-12A, IL-12B, IL-15 and IL-18) with relevant biological roles in lagomorphs. Innate Immun. 2015;21: 787–801. 10.1177/1753425915606209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mestas J, Hughes CCW. Of mice and not men: Differences between mouse and human immunology. J Immunol. 2004;172: 2731–8. [DOI] [PubMed] [Google Scholar]
- 43.Kielian T, Bearden ED, Baldwin AC, Esen N. IL-1 and TNF-α play a pivotal role in the host immune response in a mouse model of Staphylococcus aureus-induced experimental brain abscess. J Neuropathol Exp Neurol. 2004;63: 381–96. [DOI] [PubMed] [Google Scholar]
- 44.Holley MM, Kielian T. Th1 and Th17 cells regulate innate immune responses and bacterial clearance during central nervous system infection. J Immunol. 2012;188: 1360–70. 10.4049/jimmunol.1101660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cho JS, Guo Y, Ramos RI, Hebroni F, Plaisier SB, Xuan C, et al. Neutrophil-derived IL-1β is sufficient for abscess formation in immunity against Staphylococcus aureus in mice. PLoS Pathog. 2012;8: e1003047 10.1371/journal.ppat.1003047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chan LC, Chaili S, Filler SG, Barr K, Wang H, Kupferwasser D, et al. Nonredundant roles of interleukin-17A (IL-17A) and IL-22 in murine host defense against cutaneous and hematogenous infection due to methicillin-resistant Staphylococcus aureus. Infect Immun. 2015;83: 4427–37. 10.1128/IAI.01061-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.De Navasquez S. Experimental pyelonephritis in the rabbit produced by staphylococcal infection. J Pathol Bacteriol. 1950;62: 429–36. [DOI] [PubMed] [Google Scholar]
- 48.Freedman LR. Experimental pyelonephritis VI. observations on susceptibility of the rabbit kidney to infection by a virulent strain of Staphylococcus aureus. Yale J Biol Med. 1960;32: 272–9. [PMC free article] [PubMed] [Google Scholar]
- 49.Miller LS, Pietras EM, Uricchio LH, Hirano K, Rao S, Lin H, et al. Inflammasome-mediated production of IL-1β is required for neutrophil recruitment against Staphylococcus aureus in vivo. J Immunol. 2007;179: 6933–42. [DOI] [PubMed] [Google Scholar]
- 50.Krishna S, Miller LS. Host—pathogen interactions between the skin and Staphylococcus aureus. Curr Opin Microbiol. 2012;15: 28–35. 10.1016/j.mib.2011.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gautam N, Maria Olofsson A, Herwald H, Iversen LF, Lundgren-Akerlund E, Hedqvist P, et al. Heparin-binding protein (HBP/CAP37): A missing link in neutrophil-evoked alteration of vascular permeability. Nat Med. 2001;7: 1123–7. [DOI] [PubMed] [Google Scholar]
- 52.Edens HA Parkos CA. Neutrophil transendothelial migration and alteration in vascular permeability: focus on neutrophil-derived azurocidin. Curr Opin Hematol. 2003;10: 25–30. [DOI] [PubMed] [Google Scholar]
- 53.Weisel JW, Fibrinogen and fibrin. In: Fibrous Proteins: Coiled-Coils, Collagen and Elastomers. Adv Protein Chem. 2005;70: 247–99. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Scatter plot of abscess volumes from data shown in Fig 1A. Rabbits were infected subcutaneously with S. aureus Newman wild-type (WT) or isogenic mutant strains. The volume of 10 abscesses per bacterial strain was measured for 14 days following inoculation. Each symbol represents a data point obtained from a single abscess.
(TIF)
S. aureus was cultured in human heparinized blood up to 2 h. Accumulation of proinflammatory molecules in plasma was evaluated by quantitative, multiplexed immunoassays (HumanMAP v2.0; Myriad RBM) as described in Materials and Methods section. Data represents average of 3 donors ±SEM.
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.