

Abstract
The fossil record provides the primary source of data for calibrating the origin of clades. Although minimum ages of clades are given by the oldest preserved fossil, these underestimate the true age, which must be bracketed by probabilistic methods based on multiple fossil occurrences. Although most of these methods assume uniform preservation rates, this assumption is unsupported over geological timescales. On geologically long timescales (more than 10 Myr), the origin and cessation of sedimentary basins, and long-term variations in tectonic subsidence, eustatic sea level and sedimentation rate control the availability of depositional facies that preserve the environments in which species lived. The loss of doomed sediments, those with a low probability of preservation, imparts a secular trend to fossil preservation. As a result, the fossil record is spatially and temporally non-uniform. Models of fossil preservation should reflect this non-uniformity by using empirical estimates of fossil preservation that are spatially and temporally partitioned, or by using indirect proxies of fossil preservation. Geologically, realistic models of preservation will provide substantially more reliable estimates of the origination of clades.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
Keywords: stratigraphy, confidence interval, divergence times, clade origin
1. Introduction
The fossil record is a critical source of data for calibrating phylogenies and rates of evolution. Although the oldest fossil of a clade supplies a minimum age for that clade's origin, a suite of fossil occurrences permits probabilistic models of that clade's maximum age [1–6]. The reliability of such probabilistic ages depends on several factors, including establishing the phylogenetic topology, identification of taxa and their assignments to a clade, correlation of strata from different regions, absolute age estimates of fossil-bearing strata, as well as the sampling of the fossil record [7]. This paper focuses on the last of these issues, specifically how the stratigraphic record is assembled and its implications for the preservation and sampling of fossils.
Through a series of modelling and field studies, the stratigraphic distribution of species in local stratigraphic sections is well understood for shorter geological timescales (less than 10 Myr) [8,9]. These studies guide the interpretation of the fossil record over these timescales, including patterns of first and last occurrence, changes in relative abundance and community composition, and morphological changes. In particular, these studies demonstrate the marked non-randomness of fossil occurrences as a result of the processes that govern sedimentation and their effects on the resulting sedimentary record, known as stratigraphic architecture. These studies have led to probabilistic models of fossil occurrence that reflect the underlying controls on the fossil record in local sections [10], and they improve upon ones that assume uniform preservation. These models show, for example, that the difference in age between when a species first appears locally in the fossil record and when it first appeared in the region can often be up to a million years or more [11]. Similarly, the stratigraphic record can cause the last occurrences of species to be clustered at particular layers in a pattern that might lead one to interpret it as a mass extinction, when in fact that clustering is controlled by processes of sedimentation [12]. These modelling studies and numerous field studies that support them show that the fossil record cannot be treated simply at face value as a history of the presence of a species in an area through time [8,9].
Controls on the stratigraphic distribution of fossils are less well understood for larger spatial scales (regional to global) and longer spans of time (more than 10 Myr). Over these large spatial and temporal scales, several additional factors control the accumulation of sediment and the preservation of fossils, and these factors are non-uniform over space and time. Even though patterns of fossil occurrence at these larger scales cannot simply be scaled up from smaller-scale models, such non-uniformity means the fossil record cannot be read at face value over larger spatial and temporal scales.
In particular, the preservation of fossils will be highly non-uniform over the surface of the Earth and through time. This marked non-uniformity requires that probabilistic models for estimating the origin of clades incorporate spatially and temporally variable rates of preservation. This paper addresses the controls on fossil occurrence over large spatial and temporal scales in terrestrial and shallow-marine sedimentary deposits (i.e. shelf, typically up to approx. 100–200 m in depth, but not continental slope and deeper-water environments) and their implications for estimating the time of clade origin.
2. The stratigraphic record as the primary control of the fossil record
Because fossils are found in sedimentary rock, the accumulation of sediments is the primary control on the occurrence of fossils. Whether a particular organism is found in any given sedimentary rock is controlled by the age of the rock, where the organism lived and taphonomic (fossilization) processes. Because most organisms are preserved largely in the habitat in which they lived [13], and because taphonomic processes are strongly tied to sedimentary environment [14], the preservation of an organism is primarily governed by the preservation of strata from the environment in which it lived. Because sedimentary environments shift laterally over time in response to sedimentation and relative changes in sea level, the accumulation of rock in an area is not only a record of changing geological age, but also one of continually changing sedimentary environment (i.e. sedimentary facies, the rock record of that environment). Thus, the fossil record does not simply record the origination and extinction of clades. It is pervasively controlled by whether sediment from the environment in which members of a clade lived was deposited within the biogeographic range of that clade.
Over time scales greater than a few thousand years, the accumulation of sediment is controlled by changes in accommodation, a vertical distance or volume reflecting the movements of the ocean surface caused by changes in the position of sea level, as well as tectonic subsidence, the vertical movements of Earth's crust caused by plate tectonic processes. Accommodation is therefore often thought of as a volume within which sediments can accumulate. Accommodation is secondarily affected by sediment compaction (their compression from the weight of overlying sediment), as well as isostatic adjustment (depression of Earth's surface) to the weight of sediment and water [15–17]).
As this accommodation fills with sediment, water depth or elevation changes by a simple relationship: the change in water depth or elevation equals the change in accommodation minus the thickness of accumulated sediment. Because compaction and isostatic subsidence respond to the changing amounts of overlying sediment and water, changes in water depth or elevation primarily reflect the balance of tectonic subsidence, eustasy (global sea level) and the supply of sediment.
Accommodation must be created for sediment to accumulate and persist over geologically long timescales without being subsequently eroded. Thus, the formation of a sedimentary record requires eustatic sea-level rise or tectonic subsidence. In uplifting regions or regions experiencing a relative sea-level fall, newly deposited sediment tends to be eroded and carried away, and older sediment and rock may also be eroded. Areas undergoing tectonically driven subsidence tend to be large, tens to thousands of kilometres wide, and are known as sedimentary basins. Because the total subsidence in sedimentary basins commonly exceeds several thousand metres [16], far more than the roughly 200 m of variation in eustatic sea level [18], the accumulation of a substantial sedimentary record requires the formation of sedimentary basins [19]. Spatial and temporal variations in the rates of tectonic subsidence, eustasy and sediment supply control the timing and type of sediment accumulation within sedimentary basins.
(a). Extent of sedimentary basins
At any given time, only a limited portion of Earth's surface is occupied by sedimentary basins (approx. 16% today [20]). Other areas that are undergoing uplift and erosion supply sediment to the basins, in addition to carbonate sediments produced by organisms living primarily within marine environments. For example, Neogene (2.58–23 Myr) sediments of North America are limited primarily to coastal areas, where they consist overwhelmingly of shallow-marine deposits (figure 1). Neogene terrestrial deposits on North America are limited mainly to two small areas in the centre of the continent. Thus, shallow-marine and terrestrial biotas have markedly different probabilities of being sampled from North America, particularly when biogeographic distributions of clades are considered. Although there is a good chance that a marine species could be preserved in Neogene sediments from North America, many terrestrial plants and vertebrates, particularly those with small ranges, simply did not live where they could be preserved.
Figure 1.
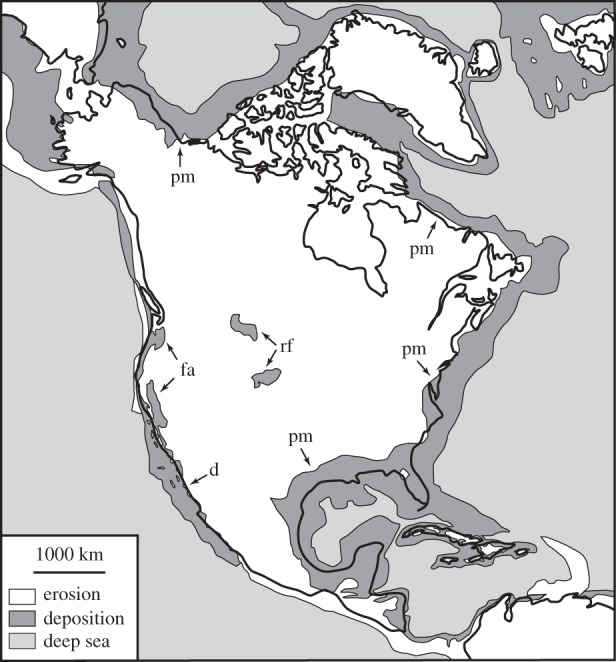
Preserved Neogene sediments from North America, shown in dark grey (terrestrial and shallow marine) and light grey (deep marine). Areas in white have undergone net erosion since the Neogene, and as a result, any sediments that may have been deposited in these areas have since been eroded away. pm, passive margin; rf, retro-foreland; fa, forearc; d, doomed sediments. Adapted from [21].
Every continent displays different patterns of sedimentation and erosion, depending on its tectonic setting. These patterns of sedimentation and erosion vary markedly over time, and they reflect the changing tectonic history of a continent. For example, North America was nearly blanketed by shallow-marine deposition during the Ordovician Period (443–485 Myr), and regions of terrestrial deposition were widespread in the Palaeogene (23–66 Myr) [21].
Over shorter time spans, sediment can be deposited more widely than just in subsiding basins. For example, Pleistocene (11 kyr–2.58 Myr) to Holocene (present–11 kyr) sediment is present along many lakes and rivers across North America, even in areas that are undergoing net uplift and erosion. Because of this net uplift, this sedimentary record is ephemeral and unlikely to persist into deep geological time (i.e. more than 1–10 Myr) [20,22], and these deposits can be considered ‘doomed sediments'. This decrease in the quantity of the stratigraphic record backward through time, as well as its accessibility at Earth's surface, are major causes of the pull of the recent, where the quality of fossil record declines with geological age [23,24].
(b). Types of sedimentary basins
Sedimentary basins form from a variety of tectonic processes, and these change systematically over long spans of geological time. Basins differ widely in their size, rates of subsidence and sediment supply, and as a result, have characteristic differences in the facies they contain. Given the primacy of sedimentary basins for preserving a fossil record, and given their markedly different characteristics, it is essential to review the main types of sedimentary basins.
Sedimentary basins are broadly divided into two categories based on the mechanism that causes them to subside [16], each of which will be discussed below. Basins formed by stretching and cooling of the lithosphere (the outer rigid portion of Earth) include rifts, passive margins and cratonic basins. Basins formed by flexure and bending of the lithosphere include two types of foreland basins.
Rift basins form where continental lithosphere is stretched and thinned during continental break-up [25]. Subsidence rates in rift basins are relatively rapid, and vary markedly over distances as short as a few tens of km. Although individual rift basins are on the scale of tens to hundreds of km, rift systems composed of multiple basins may be up to several hundred km wide, and are often much longer. Rift basins commonly begin with a rapid transition from fluvial (river channels and their neighbouring floodplains) to lacustrine (lake) deposits, because the rate of subsidence initially exceeds sediment supply. This is followed by a long shallowing upward to fluvial deposition in their later history as the rate of sedimentation overtakes the rate of subsidence [25]. Unlike most basins, rift basins can form at relatively high elevations (more than 100 m above sea level), especially early in their history. For example, lakes along the east African rift system are at elevations of 500–1500 m, and northern portions of the Rio Grande Rift in the USA are at elevations up to 2300 m. Although failed rifts may remain at relatively high elevations, rift basins that evolve into passive margins eventually subside to near or below sea level and are eventually filled by marine deposits. The elevation of environments within a terrestrial sedimentary basin, as well as the water depths within marine sedimentary basins, are important to consider because of the strong ties of the distribution of biotas to elevation and water depth [9]. A recurring theme will be the difficulty of preserving high-elevation terrestrial deposits, which greatly limits the preservation of some biotas.
Passive margins evolve from the late stage of rifting, and as a result, are necessarily limited to the peripheries of continents, such as the Atlantic coasts of the Americas, Europe and Africa. During rifting and stretching, continental lithosphere is heated from below, and it cools and subsides following rifting. Like oceanic lithosphere, cooling continental lithosphere follows a characteristic exponential subsidence history with progressively slowing subsidence rates over time. Cooling-driven subsidence can persist for up to 600 Myr, with an average basin lifespan of 180 Myr [26,27]. Passive margins are thus one of the longest-lived and most spatially extensive of all sedimentary basins [27,28]. Progressively slowing subsidence of passive margins causes sedimentary environments to build seaward through time, producing a net upward shallowing through much of the basin. Subsidence rates increase seawards on a passive margin, and rivers therefore introduce sediment where subsidence rates are lowest. As a result, terrestrial deposits on passive margins are typically thin and deposited at low elevations (less than 100 m above sea level), and most sediment deposited on passive margins is marine. Subsequent uplift and erosion of the inner edge of a passive margin tends to erode and destroy terrestrial deposits, making the record of passive margins even more dominated by shallow-marine deposits.
Cratonic basins are produced by a small amount of stretching subsidence, followed by a long history of cooling subsidence that lasts for several hundred million years [29,30]. Because their total subsidence is controlled by the amount of stretching [16], subsidence rates in cratonic basins are substantially slower than those in passive margins. Cratonic basins have a characteristic circular shape, can be over 500 km in diameter, and have subsidence rates that decrease outwards from the centre of the basin. Cratonic basins form in the interiors of continents and may form in clusters (e.g. the Williston, Illinois and Michigan Basins of North America). Their subsidence patterns may overlap one another and other basins, such as adjacent passive margins and foreland basins [31]. Cratonic basins are generally filled with marine or low-elevation terrestrial deposits, depending on the position of sea level and the supply of sediment.
Foreland basins form by flexure and bending of the lithosphere, where the lithosphere is laterally compressed, often as a result of continental collision. Compression of the lithosphere creates thrust faults that cause portions of the crust to become stacked upon itself. Mountains formed by this crustal thickening create a vertical load or weight on the lithosphere. The lithosphere is depressed (subsides) under the weight of this load. Because the lithosphere has strength, this subsidence occurs not only under the load, but also adjacent to it, essentially forming a moat called a foreland basin [32]. Foreland basins occur as two distinct types, known as pro-foreland and retro-foreland basins [33,34]. Pro-foreland basins form at the edge of continental lithosphere, whereas retro-foreland basins form in the interior of continents. Although pro-foreland and retro-foreland basins can occur independently of one another, they may form simultaneously during continental collisions, with the pro-foreland basin separated from the retro-foreland basin by a mountain range.
Subsidence rates in a foreland basin reflect the rates of crustal thickening and erosion within the mountain belt. In a pro-foreland basin, the load migrates over time towards the continent interior, and it progressively incorporates and deforms sediment previously deposited in the basin [34]. The preserved pro-foreland basin records the final position of the load and the subsidence generated by it, followed by erosion of the load and filling of the foreland basin with sediment. In contrast, the load in a retro-foreland basin stays in a relatively static position, and the basin therefore records the complete history of loading. This history starts with rapid crustal thickening in the load and therefore rapid subsidence in the basin, followed by slower thickening and erosion of the load, leading to little or no subsidence and rapid sedimentation. This is followed finally by erosion of the load, with final uplift and filling of the basin [35]. As a result, foreland basins typically have two phases, an early phase of rapid deepening that records rapid crustal thickening and loading, and a later phase of net shallowing that records the erosion of the load and a decrease in the rate of subsidence [36].
Foreland basins are laterally limited, typically 100–400 km wide, depending on the rigidity of the lithosphere, and 200–2000 km long, depending on the lateral extent of the mountain load [16]. The entire history of a foreland basin, from initiation to filling, is typically 10–50 Myr, much shorter than the lifespan of a passive margin or cratonic basin [28]. During the early, rapidly subsiding phase of a foreland basin, it may fill with marine sediments if a connection to the open ocean exists (e.g. Persian Gulf), or it may fill with terrestrial sediment if such a connection is lacking (e.g. Tarim Basin, south of the Tien Shan). Terrestrial sedimentation typically occurs in the latter history of foreland basins as erosion of the mountain load causes rapid dispersal of sediment into the basin [36]. Foreland basins may develop in low elevations or at much higher elevations (e.g. 800–2300 m in the Tarim Basin, 500–2000 m in the western USA).
Several other types of basins exist, such as pull-apart, piggyback, forearc and backarc basins [16]. Some of these basins (e.g. pull-apart, piggyback) are less important for the fossil record, owing to their smaller area. For example, 74% of modern basin area comprises passive margins, cratonic basins and foreland basins [20]. Forearc and backarc basins constitute an additional 21% [20], but these basins are less likely to survive into the deep geological record, owing to their tendency to be destroyed through deformation, metamorphism and erosion in collisions with islands or continents [22].
(c). Importance of changes in accommodation for deposition in sedimentary basins
Within a sedimentary basin, the history of deposition and the types of preserved sedimentary environments are driven by changes in accommodation, that is, changes in the rates of subsidence and eustatic sea level change over time. The application of the principles of accommodation and sedimentation to understanding and describing the sedimentary record is known as sequence stratigraphy. All sedimentary basins experience a change in their subsidence rate over time, and eustatic sea level changes continuously, owing to variations in glacial ice volume, groundwater and lakes, ocean temperature, mid-ocean-ridge spreading rates, continental collision and break-up, and sedimentation in ocean basins [18]. In addition, dynamic topography caused by mantle upwelling causes waves of uplift and subsidence to pass through all types of sedimentary basins on timescales of 1–10 Myr, creating relative sea-level variations that may commonly be mistaken for eustatic sea-level changes [37]. As a result, sequence stratigraphic principles that are widely applied over timescales of less than 10 Myr also apply over longer timescales [38–40]. Particularly important is the concept of stacking patterns, which describe how strata (e.g. parasequences and sequences) are spatially arranged. Stacking patterns in marine and terrestrial systems are recognized on different criteria.
In marine systems, four stacking patterns are now recognized: retrogradational, aggradational, progradational and degradational (figure 2) [17,40,41]. Each represents the balance between the rate of accommodation and the rate of sedimentation, and each has characteristic and predictable expressions in the sedimentary record. Retrogradational stacking develops when accommodation exceeds sedimentation. As a result, stratal units (such as parasequences or sequences) are stacked upwards and landwards through time, creating a net upward deepening. Aggradational stacking forms when the rates of accommodation and sedimentation are equal. In aggradational stacking, stratal units are placed vertically on top of the preceding unit, with no net landwards or seawards displacement, producing a trend of no net upwards shallowing or deepening. Progradational stacking occurs when the sedimentation is faster than accommodation, provided that the rate of accommodation is positive. Progradationally stacked units are stacked upwards and seawards through time, causing a net upward shallowing. Degradational stacking is similar to progradational stacking but occurs when the rate of accommodation is negative, which requires uplift or eustatic sea-level fall. In degradational stacking, each successive unit is placed seawards and downwards along the depositional profile.
Figure 2.
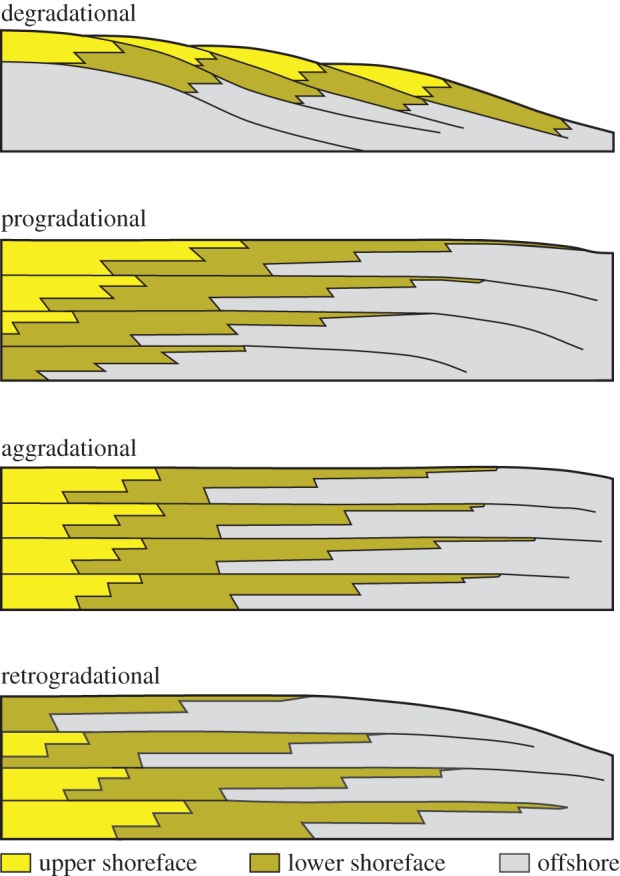
Idealized cross section, showing the four types of stacking patterns. In each cross section, landward is to the left and seaward is to the right, modified from [34]. Rates of accommodation are negative for degradational stacking becoming increasingly positive from progradational to retrogradational stacking. Although stacking for upper shoreface, lower shoreface and offshore facies is shown, any three laterally adjacent marine environments from relatively shallow to relatively deep could be substituted.
In terrestrial systems, stacking patterns are also based on the relative rates of accommodation and sedimentation (figure 3). Where the accommodation is faster than sedimentation, fluvial systems tend to be dominated by floodplain deposits with isolated fluvial channels [42] or by lacustrine systems [43]. Where the sedimentation exceeds accommodation, terrestrial deposits tend to consist of stacked fluvial channels with minor floodplain deposits.
Figure 3.
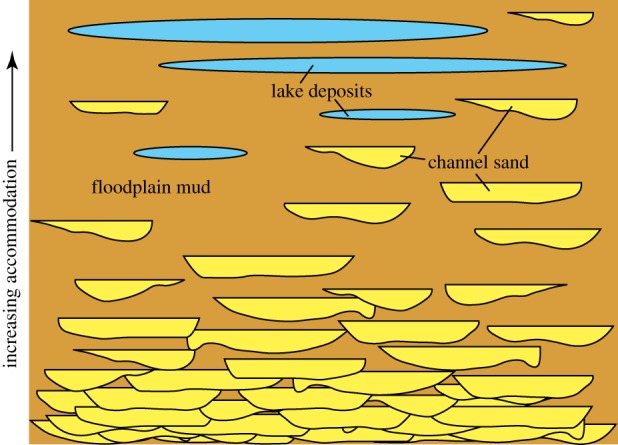
Idealized cross section, showing the change in architecture of terrestrial (river channel, floodplain, lake) deposits as the rate of accommodation increases. Adapted from [9].
Changes in subsidence rate over the history of a sedimentary basin control its long-term stacking patterns. For example, subsidence is faster earlier in the history of rifts, passive margins, cratonic basins and foreland basins than later, and as a result, these basins tend to have initial retrogradational stacking, followed by increasingly progradational stacking [36,44]. Foreland basins also oscillate between periods in which the rate of crustal thickening and loading exceed the rate of mountain erosion and those in which the rate of erosion is greater than the rate of loading. Consequently, foreland basins vary over timescales of 10 Myr with alternating retrogradational and progradational stacking where marine facies are present [45]. Where filled with fluvial sediment, foreland basins alternate between floodplain-dominated intervals with isolated channel deposits and periods characterized by stacked fluvial channel deposits [42,46], that is, between deposits like those at the top and the bottom of figure 3.
Because sea-level change is cyclical over a wide range of timescales [18], complex patterns of stacking can arise (figure 4). For example, a short-term rapid relative rise in sea level superimposed on a long-term slow relative rise in sea level can generate small-scale retrogradational stacking within large-scale progradational stacking. This hierarchical arrangement produces what are known as sequence sets. As a result, a particular facies may be encountered repeatedly with a short vertical stratigraphic interval, but separated from other occurrences of the facies by thick intervals of dissimilar facies. In addition, the presence of hiatuses (periods of non-deposition or erosion) at sequence boundaries and sequence-set boundaries will cause these facies intervals to be even more widely spaced in geological time.
Figure 4.

Highly stylized cross section of sequence sets, illustrating progradational sequence stacking in the lowstand sequence set (LSS) and highstand sequence set (HSS), retrogradational stacking in the transgressive sequence set (TSS), and degradational stacking in the falling-stage sequence set (FSS), modified from [38]. Each sequence set is composed of multiple depositional sequences (sensu [40]).
3. Principles for interpreting the antiquity of clades
Given the unlikelihood of preserving and sampling the oldest member of a clade, as well as the time that must elapse between genetic divergence and acquisition of a preservable character that allows the clade to be diagnosed [7], the oldest sampled member of a clade will always underestimate the age of the clade. Estimating a clade's true antiquity, therefore, generally requires an extrapolation or a probabilistic estimation based on rates of fossil preservation. Estimates of these rates should reflect the fossilization process, the structure and nature of the stratigraphic record over long spans of geological time, as well as the effects of uneven sampling by palaeontologists.
(a). Preservation varies among taxa
Variations in preservation among taxa reflect not only the presence or absence of hard parts (shell, bone, wood, etc.), but also their chemical composition, shape and size [14,47,48]. Organisms lacking hard parts generally have their preservation limited to relatively few deposits with extraordinary preservation [49], which hinders efforts to develop quantitative models of fossil occurrence. For organisms with hard parts, differences in preservation necessitate separate estimates of preservation rate for different clades. A common approach for inferring whether a normally preservable organism is absent within a stratigraphic interval is to use a taphonomic control taxon, that is, another taxon that commonly co-occurs in taphonomically similar deposits [50]. This approach can and should be extended to taxa that are not only taphonomically similar, but also environmentally similar, as organisms may have similar probabilities of fossilization yet lived in different environments.
(b). Preservation varies among depositional environments
Clades are most likely to be found in the environment in which they lived [9]. Although transport out of habitat has been reported, this is often because the circumstances are so extraordinary, such as ankylosaurs preserved in marine deposits [51], or nautiloids carried thousands of kilometres by ocean currents [52]. Even so, post-mortem transport is the exception rather than the rule [13]. Estimates of the preservation rate of a clade should therefore reflect the occurrence of the depositional environment in which the clade lived. For example, regarding all marine strata as a sampling opportunity is likely to be misleading, because clades are generally confined to a narrow range of environments, particularly at lower taxonomic levels [9].
The association of marine taxa with particular environments often weakens at higher taxonomic levels. For example, marine invertebrate genera display a strong association to specific environments over timescales approaching 10 Myr [53–57]. Families and orders of marine invertebrates commonly show a broader distribution among environments than genera at any one time, as well as a drift to deeper-water environments over timescales of tens to hundreds of millions of years [50,58–61].
The occurrence of terrestrial taxa is likewise tied to depositional environments, with temperature, elevation and moisture being the dominant controls [62–65]. Terrestrial floras and faunas also undergo evolutionary changes in their preferred habitat and biome [66]. They may also display strong differences in preservation among associated depositional environments, which can exert a powerful control on their preservation and abundance [48]. For example, articulated and well-preserved vertebrate fossils are more common in floodplain and palaeosol (soil) deposits, whereas vertebrate fossils in river channel deposits are more commonly disarticulated, dissociated and abraded [67,68]. Similarly, floodplain deposits are more likely to contain leaves and in situ wood, whereas channel deposits are more likely to contain transported logs, limbs and roots [65,69].
The preferred habitat of some organisms may not be an environment in which sediment deposition generally occurs. For example, rocky coasts are rarely preserved [70], such that the fossil record of marine organisms requiring hard substrates is largely limited to hardgrounds (surfaces of seafloor cementation that develop within depositional areas) and cases in which they attach to other shells [71–73]. For terrestrial settings, large areas of modern continents are non-depositional. For such clades, preservation rates may be low, or even effectively zero, and decreasing with geological age, owing to the loss of doomed sediments.
(c). Preservation varies among sedimentary basins
Preservation of a clade varies among sedimentary basins, owing to the biogeographic distribution of the clade, as well as the size, longevity and preserved environments of basins. The concept of sedimentary basins, that is, that sedimentation is geographically focused in limited areas, is of particular importance for determining the origin of clades, because clades are generally rare and geographically restricted early in their history [74–76]. Owing to sedimentary basins, repeated sampling at any one time will be limited to relatively few areas, with a necessarily limited coverage of biomes and biogeographic provinces.
As a result of their widely varying sizes, different types of sedimentary basins offer greatly different opportunities for sampling clades, particularly when clades are confined to few biogeographic provinces. Given the limited number of basins, the fossil record will consist of repeated sampling opportunities from relatively few biogeographic provinces, and this structure will be more pronounced for terrestrial basins, especially upland basins.
Differences in basin longevity also play a role. Cooling-driven basins (passive margins, cratonic basins) typically last an order of magnitude longer than flexural basins (foreland) and stretching basins (rifts) [28]. As a result, cooling-driven basins will offer more persistent opportunities for sampling clades and biogeographic provinces than will be possible in flexural basins or rifts.
Basins differ systematically in their preserved sedimentary environments, leading to substantial differences in the nature of the marine and terrestrial fossil records. Marine deposition occurs in many flexural basins, but dominates the substantially larger and longer-lived cooling basins. In contrast, terrestrial deposition is most common in stretching and flexural basins. Within terrestrial systems, there are marked differences in the preservation of low-elevation and high-elevation deposits. Low-elevation settings are more likely to be preserved within pro-foreland, retro-foreland, rift, and cratonic basins. Of these, only cratonic basins are likely to have geologically sustained deposition. High-elevation provinces and biomes are less likely to be preserved and are largely limited to some rift and foreland basins.
(d). Preservation varies through time
Preservation of clades varies through time for biological and stratigraphic reasons. Biologically, clades tend to be rare and geographically restricted initially, then expand through the first half of their history in both abundance and geographical range; and then contract through the latter half of their history prior to their extinction [74–77]. Stratigraphically, preservation varies through time, partly from the formation and cessation of sedimentary basins, but also through temporal variations in subsidence, eustatic and sedimentation rates.
The formation and cessation of sedimentary basins provides the first-order control on fossil preservation, with temporal variations in subsidence rate and eustatic sea level providing a second-order control. As a result, fossil preservation will not only be clustered geographically, it will be clustered through time. One might hope that such clustering might somehow cancel out on the global scale, leading to globally uniform rates of preservation. This is unlikely to be true for three reasons.
First, the initiation of particular types of sedimentary basins is non-uniform and correlated over time. The roughly 400 Myr cycle of supercontinent formation and break-up, known as the Wilson cycle [78], produces systematic variations in the types of basins forming over geological time [28]. For example, supercontinent break-up leads to the formation of long chains of rift basins of similar geological age, succeeded by a pair of passive margins on opposite sides of the newly formed ocean basin. Similarly, cratonic basins formed in two global pulses, one coinciding with the break-up of Rodinia in the Late Proterozoic to Early Cambrian and the other coinciding with the break-up of Pangaea in the Late Permian to Triassic [29,79]. In contrast, times of supercontinent assembly generate a series of collisions between arcs (volcanic island chains), islands and continents, and this results in a protracted interval in which pro-foreland and retro-foreland basins form. The Wilson cycle is also significant in that long-term sea level is strongly correlated, with times of supercontinent assembly characterized by low eustatic sea level and times of supercontinent break-up corresponding to high positions of eustatic sea level [80].
Second, basins of a similar type and age will share similar subsidence and therefore depositional histories, such as the pair of passive margins on either side of the Atlantic Ocean or the trio of North American cratonic basins initiated in the Cambrian. Because basins tend to have declining rates of subsidence over time, basins of the same age will follow a similar retrogradational to progradational history. As a result, basins of similar type and age will tend to have a similar stratigraphic architecture and facies. Likewise, long-term eustatic sea-level changes will cause basins to have correlated stacking patterns, resulting in parallel variations in preservation rate. Within any individual basin, long-term stacking patterns will systematically control the availability of particular sedimentary environments, causing the sampling of any particular environment to be highly non-uniform through geological time.
Third, the progressive loss of doomed sediments over time systematically changes the rate of fossil preservation. For example, caves form in areas that are undergoing net erosion, yet sediment may be washed into caves and remain over relatively short time scales. Although ancient cave fills can be preserved, even ones bearing fossils, cave-fill deposits are overwhelmingly most common in Pleistocene to Holocene records [81], where they may be an important source of fossils. Many modern river, lake and pond deposits are also examples of doomed sediment where they are not accumulating in subsiding basins. Although these doomed sediments may be palaeontologically important records of the most recent history of clades, they are progressively lost with time, diminishing their importance in older geological intervals.
Doomed sediments are not limited to surficial terrestrial deposits. Sedimentary basins that lie along convergent continental margins have a lower probability of surviving subsequent continental collision, metamorphism and erosion. For example, forearc basins older than 10 kyr have less than a 20% probability of surviving a 10-fold increase in age, compared with passive margin sediments, whose probability is 60–70% [22]. Deep-sea sediments also have a steadily decreasing probability of preservation with age [82,83]. Half of the oceanic lithosphere is less than 85 Ma, and all overlying sediments are therefore younger. No oceanic lithosphere today is older than 180 Ma, creating an upper limit to the age of deep-sea sediment, except in the rare cases where oceanic lithosphere has been uplifted and emplaced onto a continent and not subsequently metamorphosed.
Doomed sediments are important because they impose an irreversible decline in the probability of preservation backward through time. Importantly, however, doomed sediments are predictably more limited to certain terrestrial records, the deep sea and particular types of basins along convergent margins. Whether they are important in any particular study of the fossil record will depend on the antiquity, biogeography and environment of a clade.
(e). Sampling varies with exposure and collection
Even though sedimentary basins may preserve a wide range of sedimentary environments of any given age, only a limited set of these can be sampled in most cases. Rocks of any given age are accessible in only a small portion of most sedimentary basins where erosion was sufficient to bring them to the surface but not enough to remove them altogether. As a result, it is often not possible to sample coeval shallower-water strata in more landward areas where uplift has caused wholesale erosion and destruction of the rock record [84]. Likewise, coeval deeper-water strata may be buried in seaward locations, making their fossils largely inaccessible, except in drill cores.
The correspondence between subsequent structural deformation and the original sedimentary basin will dictate how extensively rocks of a given age can be sampled within a sedimentary basin. Where subsequent deformation is unrelated to the mechanism of basin formation, more complete sampling of the basin is possible. For example, the latest Cretaceous to Early Palaeogene mountain building in the western USA uplifted older passive margin and foreland basin deposits in a complex pattern that provides widespread opportunities for sampling rocks of a given age. In contrast, uplift on the edges of many Mesozoic to Cenozoic passive margins, such as along the Atlantic Ocean, is parallel to the basin margin. As a result, rocks of a given age are exposed only in narrow belts largely parallel to the original depositional strike, that is, parallel to the ancient shoreline. For example, Cretaceous rocks in New Jersey are exposed in a belt only 25 km wide, and that of the entire Palaeogene and Neogene is only 50 km wide. Because both of these belts are largely parallel to depositional strike, exposed rocks of a given age in New Jersey contain a narrow range of sedimentary environments.
Compounding these limitations to sampling, collection effort by palaeontologists is greatly uneven, with some regions receiving intense study and others receiving little or none, despite the availability of fossiliferous rock. Databases can exacerbate this, particularly in their early stages [85]; for example, currently 78% of vertebrate collections and 74% of marine invertebrate collections in the Paleobiology Database are from North America and Europe.
As a result of exposure and collection effort, the number of sampling opportunities for any given geological time interval can be significantly less than what is available in the fossil record, making the described fossil record of any given age dependent on a few regions. Consequently, patterns of exposure and study by palaeontologists make it less likely that sampling will be sufficient to even out heterogeneities in fossil preservation.
4. Realistic models of fossil preservation
The simplest and most straightforward model of fossil preservation assumes that preservation is constant through time and space, that is, a stationary Poisson process. Although such a model was a reasonable and necessary assumption when confidence intervals on fossil ranges were first developed [86,87], it is clear now that preservation is non-uniform on both short [8,9] and long timescales.
Preservation of fossils varies among depositional environments, among depositional basins, and through time. Sampling also varies, owing to limited exposure and variations in collection effort. The net effect is that fossil occurrences will be highly clumped both spatially and temporally. Such clumping was noted in the earliest studies of gap distributions in the fossil record [86,88,89]. Clumping typically has been detected as departures from exponential gap distributions, often with the Kolmogorov–Smirnov (K–S) test, widely criticized for its low statistical power [90,91]. Given the potential for type 2 errors with the K–S test, non-random gap distributions are probably more common than is generally reported. Unfortunately, models assuming uniform preservation continue to be used widely [4,6,92–94].
Rather than start with a geologically unrealistic model of uniform preservation, evaluated with low-power statistical tests, models of preservation should start with an informed prior, namely that fossil preservation is non-uniform in space and time [5,10,95]. At small spatial and temporal scales (less than 10 Myr), such models have been useful for understanding a wide range of patterns in the fossil record [8–12]. Theoretical models for larger spatial and temporal scales do not yet exist, but could be developed from the principles described here. For now, empirical models that capture the stratigraphic and geographical structure of fossil occurrences are readily developed, even with a limited understanding of the facies and basins in which clades are found. To be useful, such empirical models need not capture all aspects of sedimentary environments, basins and temporal variations in fossil recovery, as incorporating even some of these would still improve upon models that assume uniform preservation.
Several approaches to non-uniform preservation have been adopted. Perhaps the most straightforward is to use databases to partition the data and develop per-locality sampling rate distributions that capture environmental, geographical and temporal variations in fossil recovery [5]. For example, Wagner & Marcot [5] calculated separate lognormal rates of recovery with the data binned by stage and continent, and using lepidosauromorphs (lizards, snakes, etc.) as a taphonomic/ecological control taxa for mammals, the subject of their study. They found that these distributed rates perform ‘vastly better’ than the best uniform sampling rates. Partitioning their data by depositional basins rather than continents would probably give superior rates, given that it would capture the underlying basin-scale controls on fossil occurrence. Likewise, partitioning the data by depositional environment may also yield improved results, but such an approach might be hindered by a lack of consistent environmental interpretations as well as problems of overpartitioning, leading to bins with insufficient data. Nonetheless, the approach of Wagner & Marcot [5] shows that considering environmental, geographic and temporal variations in sampling rate is superior to assuming uniform preservation.
Partitioning the data and using taphonomic/ecological control groups to estimate sampling probabilities can also be used to develop sampling functions for fossilized birth–death models [3,6,96], time-scaled phylogenies [4] and confidence intervals on fossil ranges [95]. It is unclear whether this approach is feasible for capture–mark–recapture methods that simultaneously estimate speciation, extinction and sampling rates [97], given that it may be difficult or even impossible to extract the large amounts of data from the fossil record needed to estimate the parameters of these methods.
Indirect proxies of preservation could also be used to constrain sampling probabilities. One possibility is rock-outcrop area (the area over which rocks of a given age are exposed at the Earth's surface), which correlates with fossil occurrences and diversity [98–103]. For example, Sansom et al. [103] used rock outcrop area and sea level to constrain the divergence times of jawless vertebrates in the lower Palaeozoic. They showed that ghost clades are explained substantially better by using confidence intervals on the oldest members of a clade that incorporate non-constant preservation. Other possible indirect proxies include the number of stratigraphic units [97], or the number of gap-bound sedimentary packages [100,104].
In many cases, indirect proxies may be more available than details on sedimentary environment, basin type and sequence stratigraphic architecture that form the direct controls on the occurrence of fossils. Indirect proxies should be used with caution as they can give a misleading picture of potential fossil recovery where not all strata are ‘sampling opportunities' [5], that is, collections from which a clade of interest could have been sampled had they been present. For example, Early Palaeozoic jawless vertebrates have a strong affinity for near-shore terrigenous sandy substrates [103,105–107]. Estimates of outcrop area in general will incorporate many more environments than such near-shore ones, thereby overestimating the opportunities for fossil recovery and artificially shortening confidence intervals on the origin of clades. Indeed, when Sansom et al. [103] accounted for habitat, the length of their confidence intervals increased, better reflecting the probability of recovery of these jawless vertebrates. If indirect proxies such as outcrop area, number of stratigraphic units and gap-bound packages were tailored to particular habitats, biomes and provinces, they could produce improved estimates of fossil recovery.
Parametrized distributions have also been used to describe variable sampling probabilities through time and among taxa [108]. For example, Silvestro et al. [108] fit a generalized beta distribution for each taxon that reflected the tendency for species to be rare at the beginning and end of their range. This was used with a birth–death model to study the diversification of a clade, but it could also be used to estimate the age of clade origin. The accuracy of this approach is affected by strong deviations from their preservation model, but only when those deviations are consistent throughout the dataset. Given the shared history of sedimentary basins, such consistency of deviations is probably the norm in the fossil record.
Exceptional fossilization (e.g. preservation of soft parts) poses a significant problem for estimating the age of clade origin [4], because such occurrences are often unique or nearly so. Considerably more examples of exceptional fossilization must be found before it is possible to develop a parametrized or empirical model of their occurrence through time, among basins and among depositional environments. Where the oldest-known member of a clade is found in a bed with exceptional preservation, it may not be possible to say more than that the minimum age of the clade is the age of that bed, and that the true age of the clade is almost certainly far older [86,87].
5. Conclusion
Although it has been well understood that the fossil record is highly structured and non-uniform over relatively short timescales (less than 10 Myr), it is also structured and non-uniform over geologically long timescales (more than 10 Myr). Not only is the availability of suitable facies a consideration on these longer timescales, the geographical distribution and types of sedimentary basins are particularly important in that they determine the sampling of provinces and biomes. Changes in eustatic sea level tend to produce correlated patterns of fossil occurrence among sedimentary basins. Long-term cycles of supercontinent formation and break-up also create correlated non-uniform patterns of basin initiation and cessation. In addition, some sedimentary environments and basins are ‘doomed sediments’, unlikely to survive into the ancient past, and these impose a secular trend on the fossil record. Deep-marine, shallow-marine, low-elevation terrestrial and high-elevation terrestrial settings are all controlled by separate factors and are expected to show discordant patterns of fossil occurrence through the Phanerozoic. Finally, exposure of sedimentary basins is non-uniform, as is study by palaeontologists.
These variations are highly structured, and empirical models can be readily developed to improve significantly on the false assumption of constant fossil preservation through time. It may also be possible to develop a theoretical model to capture the spatial and temporal variation in the fossil record, based on an understanding of basin formation and long-term variations in rates of accommodation and sedimentation. It is time to abandon the model of uniform preservation and adopt the geologically realistic case of spatially and temporally non-uniform preservation. Such an approach will produce more reliable estimates of the time of origination of clades.
Acknowledgements
I thank P. Donoghue and Ziheng Yang for the invitation to participate in the Royal Society discussion meeting, ‘Dating species divergences using rocks and clocks'. I also appreciate discussions with M. Patzkowsky, S. Danise and P. Monarrez, as well as the helpful comments on the manuscript by S. Peters, an anonymous reviewer, and guest editor P. Donoghue.
Competing interests
I have no competing interests.
Funding
Travel support was provided by the Royal Society.
References
- 1.Foote M, Hunter JP, Janis CM, Sepkoski JJ. 1999. Evolutionary and preservational constraints on origins of biologic groups: divergence times of eutherian mammals. Science 283, 1310–1314. ( 10.1126/science.283.5406.1310) [DOI] [PubMed] [Google Scholar]
- 2.Tavaré S, Marshall CR, Will O, Soligo C, Martin RD. 2002. Using the fossil record to estimate the age of the last common ancestor of extant primates. Nature 416, 726–729. ( 10.1038/416726a) [DOI] [PubMed] [Google Scholar]
- 3.Stadler T. 2010. Sampling-through-time in birth—death trees. J. Theor. Biol. 267, 396–404. ( 10.1016/j.jtbi.2010.09.010) [DOI] [PubMed] [Google Scholar]
- 4.Bapst DW. 2013. A stochastic rate-calibrated method for time-scaling phylogenies of fossil taxa. Methods Ecol. Evol. 4, 724–733. ( 10.1111/2041-210X.12081) [DOI] [Google Scholar]
- 5.Wagner PJ, Marcot JD. 2013. Modelling distributions of fossil sampling rates over time, space and taxa: assessment and implications for macroevolutionary studies. Methods Ecol. Evol. 4, 703–713. ( 10.1111/2041-210X.12088) [DOI] [Google Scholar]
- 6.Heath TA, Huelsenbeck JP, Stadler T. 2014. The fossilized birth-death process for coherent calibration of divergence-time estimates. Proc. Natl Acad. Sci. USA 111, E2957–E2966. ( 10.1073/pnas.1319091111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Donoghue PCJ, Benton MJ. 2007. Rocks and clocks: calibrating the tree of life using fossils and molecules. Trends Ecol. Evol. 22, 424–431. ( 10.1016/j.tree.2007.05.005) [DOI] [PubMed] [Google Scholar]
- 8.Holland SM. 2000. The quality of the fossil record - a sequence stratigraphic perspective. In Deep time: paleobiology's perspective (eds Erwin DH, Wing SL), pp. 148–168. Lawrence, KS: The Paleontological Society. [Google Scholar]
- 9.Patzkowsky ME, Holland SM. 2012. Stratigraphic paleobiology. Chicago, IL: University of Chicago Press. [Google Scholar]
- 10.Holland SM. 2003. Confidence limits on fossil ranges that account for facies changes. Paleobiology 29, 468–479. ( 10.1666/0094-8373(2003)029%3C0468:CLOFRT%3E2.0.CO;2) [DOI] [Google Scholar]
- 11.Holland SM, Patzkowsky ME. 2002. Stratigraphic variation in the timing of first and last occurrences. Palaios 1, 134–146. ( 10.1669/0883-1351(2002)017%3C0134:SVITTO%3E2.0.CO;2) [DOI] [Google Scholar]
- 12.Holland SM, Patzkowsky ME. 2015. The stratigraphy of mass extinction. Palaeontology 58, 903–924. ( 10.1111/pala.12188) [DOI] [Google Scholar]
- 13.Kidwell SM. 2013. Time-averaging and fidelity of modern death assemblages: building a taphonomic foundation for conservation palaeobiology. Palaeontology 56, 487–522. ( 10.1111/pala.12042) [DOI] [Google Scholar]
- 14.Brett CE, Baird GC. 1986. Comparative taphonomy: a key to paleoenvironmental interpretation based on fossil preservation. Palaios 1, 207–227. ( 10.2307/3514686) [DOI] [Google Scholar]
- 15.Pitman WC. 1978. Relationship between eustasy and stratigraphic sequences of passive margins. Geol. Soc. Am. Bull. 89, 1389–1403. ( 10.1130/0016-7606(1978)89%3C1389:RBEASS%3E2.0.CO;2) [DOI] [Google Scholar]
- 16.Allen PA, Allen JR. 2005. Basin analysis: principles and applications. New York, NY: Wiley-Blackwell. [Google Scholar]
- 17.Catuneanu O. 2006. Principles of sequence stratigraphy. New York, NY: Elsevier. [Google Scholar]
- 18.Miller KG, et al. 2005. The Phanerozoic record of global sea-level change. Science 310, 1293–1298. ( 10.1126/science.1116412) [DOI] [PubMed] [Google Scholar]
- 19.Jervey MT. 1988. Quantitative geological modelling of siliciclastic rock sequences and their seismic expression. Soc. Econ. Paleont. Mineral. Spec. Pub. 42, 47–69. [Google Scholar]
- 20.Nyberg B, Howell JA. 2015. Is the present the key to the past? A global characterization of modern sedimentary basins. Geology 43, 643–646. ( 10.1130/G36669.1) [DOI] [Google Scholar]
- 21.Blakey R. 2015. North America key time-slices paleotectonic and sedimentation maps. See http://cpgeosystems.com/namkeypaleogeography.html.
- 22.Sadler PM. 2009. Models of time-averaging as a maturation process: how soon do sedimentary sections escape reworking? Pal. Soc. Short Courses Pal. 14, 188–209. [Google Scholar]
- 23.Raup DM. 1972. Taxonomic diversity during the Phanerozoic. Science 177, 1065–1071. ( 10.1126/science.177.4054.1065) [DOI] [PubMed] [Google Scholar]
- 24.Peters SE, Foote M. 2001. Biodiversity in the Phanerozoic: a reinterpretation. Paleobiology 27, 583–601. ( 10.1666/0094-8373(2001)027%3C0583:BITPAR%3E2.0.CO;2) [DOI] [Google Scholar]
- 25.Withjack MO, Schlische RW, Olsen PE. 2002. Rift-basin structure and its influence on sedimentary systems. Soc. Econ. Paleont. Mineral. Spec. Pub. 73, 4–81. [Google Scholar]
- 26.McKenzie DP. 1978. Some remarks on the development of sedimentary basins. Earth Planet. Sci. Lett. 40, 25–32. ( 10.1016/0012-821X(78)90071-7) [DOI] [Google Scholar]
- 27.Bradley DC. 2008. Passive margins through earth history. Earth Sci. Rev. 91, 1–26. ( 10.1016/j.earscirev.2008.08.001) [DOI] [Google Scholar]
- 28.Woodcock NH. 2004. Life span and fate of basins. Geology 32, 685–688. ( 10.1130/G20598.1) [DOI] [Google Scholar]
- 29.Allen PA, Armitage JJ. 2012. Cratonic basins. In Syntectonic basin development, active to ancient: recent advances (eds Busby C, Azor A), pp. 602–620. Oxford, UK: Wiley-Blackwell. [Google Scholar]
- 30.Armitage JJ, Allen PA. 2010. Cratonic basins and the long-term subsidence history of continental interiors. J. Geol. Soc. Lond. 167, 61–70. ( 10.1144/0016-76492009-108) [DOI] [Google Scholar]
- 31.Beaumont C, Quinlan G, Hamilton J. 1988. Orogeny and stratigraphy: numerical models of the Paleozoic in the eastern interior of North America. Tectonics 7, 389–416. ( 10.1029/TC007i003p00389) [DOI] [Google Scholar]
- 32.DeCelles PG, Giles KA. 1996. Foreland basin systems. Basin Res. 8, 105–124. ( 10.1046/j.1365-2117.1996.01491.x) [DOI] [Google Scholar]
- 33.Dickinson WR. 1974. Tectonics and sedimentation. Soc. Econ. Paleont. Mineral. Spec. Pub. 22, 1–27. ( 10.2110/pec.74.22.0001) [DOI] [Google Scholar]
- 34.Naylor M, Sinclair HD. 2008. Pro- vs. retro-foreland basins. Basin Res. 20, 285–303. ( 10.1111/j.1365-2117.2008.00366.x) [DOI] [Google Scholar]
- 35.Flemings PB, Jordan TE. 1989. A synthetic stratigraphic model of foreland basin development. J. Geophys. Res. 94, 3851–3866. ( 10.1029/JB094iB04p03851) [DOI] [Google Scholar]
- 36.Heller PL, Angevine CL, Winslow NS, Paola C. 1988. Two-phase stratigraphic model of foreland basin sequences. Geology 16, 501–504. ( 10.1130/0091-7613(1988)016%3C0501:TPSMOF%3E2.3.CO;2) [DOI] [Google Scholar]
- 37.Petersen KD, Nielsen SB, Clausen OR, Stephenson R, Gerya T. 2010. Small-scale mantle convection produces stratigraphic sequences in sedimentary basins. Science 329, 827–830. ( 10.1126/science.1190115) [DOI] [PubMed] [Google Scholar]
- 38.Mitchum RM, Van Wagoner JC. 1991. High-frequency sequences and their stacking patterns: sequence-stratigraphic evidence of high-frequency eustatic cycles. Sediment. Geol. 70, 131–160. ( 10.1016/0037-0738(91)90139-5) [DOI] [Google Scholar]
- 39.Posamentier HW, Allen GP, James DP. 1992. High resolution sequence stratigraphy : the East Coulee Delta, Alberta. J. Sediment. Petrol. 62, 310–317. [Google Scholar]
- 40.Van Wagoner JC, Mitchum RM, Campion KM, Rahmanian VD. 1990. Siliciclastic sequence stratigraphy in well logs, cores, and outcrops. Tulsa, OK: American Association of Petroleum Geologists. [Google Scholar]
- 41.Neal J, Abreu V. 2009. Sequence stratigraphy hierarchy and the accommodation succession method. Geology 37, 779–782. ( 10.1130/G25722A.1) [DOI] [Google Scholar]
- 42.Bridge JS, Leeder MR. 1979. A simulation model of alluvial stratigraphy. Sedimentology 26, 617–644. ( 10.1111/j.1365-3091.1979.tb00935.x) [DOI] [Google Scholar]
- 43.Keighley D, Flint S, Howell JA, Moscariello A. 2003. Sequence stratigraphy in lacustrine basins: a model for part of the Green River Formation (Eocene), southwest Uinta Basin, Utah, U.S.A. J. Sediment. Res. 73, 987–1006. ( 10.1306/050103730987) [DOI] [Google Scholar]
- 44.Bowman SA, Vail PR. 1999. Interpreting the stratigraphy of the Baltimore Canyon section, offshore New Jersey with PHIL, a stratigraphic simulator. Soc. Econ. Paleont. Mineral. Spec. Pub. 62, 117–138. [Google Scholar]
- 45.Kamola DL, Huntoon JE. 1995. Repetitive stratal patterns in a foreland basin sandstone and their possible tectonic significance. Geology 23, 177–180. ( 10.1130/0091-7613(1995)023%3C0177:RSPIAF%3E2.3.CO;2) [DOI] [Google Scholar]
- 46.Shanley KW, McCabe PJ, Hettinger RD. 1992. Tidal influence in Cretaceous fluvial strata from Utah, USA: a key to sequence stratigraphic interpretation. Sedimentology 39, 905–930. ( 10.1111/j.1365-3091.1992.tb02159.x) [DOI] [Google Scholar]
- 47.Kidwell SM, Flessa KW. 1996. The quality of the fossil record: populations, species, and communities. Annu. Rev. Earth Planet. Sci. 24, 433–464. ( 10.1146/annurev.earth.24.1.433) [DOI] [Google Scholar]
- 48.Behrensmeyer AK, Kidwell SM, Gastaldo RA. 2000. Taphonomy and paleobiology. Paleobiology 26, 103–147. ( 10.1666/0094-8373(2000)26%5B103:TAP%5D2.0.CO;2) [DOI] [Google Scholar]
- 49.Bottjer DJ, Etter W, Hagadorn JW, Tang CM. 2002. Exceptional fossil preservation: a unique view on the evolution of marine life. New York, NY: Columbia University Press. [Google Scholar]
- 50.Bottjer DJ, Jablonski D. 1988. Paleoenvironmental patterns in the evolution of post-Paleozoic benthic marine invertebrates. Palaios 3, 540–560. ( 10.2307/3514444) [DOI] [Google Scholar]
- 51.Coombs WP, Demére TA. 1996. A Late Cretaceous nodosaurid ankylosaur (Dinosauria: Ornithischia) from marine sediments of coastal California. J. Paleontol. 70, 311–326. [Google Scholar]
- 52.Saunders WB, Spinosa C. 1979. Nautilus movement and distribution in Palau, Western Caroline Islands. Science 204, 1199–1201. ( 10.1126/science.204.4398.1199) [DOI] [PubMed] [Google Scholar]
- 53.Ziegler AM, Cocks RM, Bambach RK. 1968. The composition and structure of Lower Silurian marine communities. Lethaia 1, 1–27. ( 10.1111/j.1502-3931.1968.tb01724.x) [DOI] [Google Scholar]
- 54.Boucot AJ. 1981. Principles of benthic marine paleoecology. New York, NY: Academic Press. [Google Scholar]
- 55.Ludvigsen R, Westrop SR, Pratt BR, Tuffnell PA, Young GA. 1986. Dual biostratigraphy: zones and biofacies. Geosci. Can. 13, 139–154. [Google Scholar]
- 56.Holland SM, Patzkowsky ME. 2004. Ecosystem structure and stability: Middle Upper Ordovician of central Kentucky, USA. Palaios 19, 316–331. ( 10.1669/0883-1351(2004)019%3C0316:ESASMU%3E2.0.CO;2) [DOI] [Google Scholar]
- 57.Holland SM, Zaffos A. 2011. Niche conservatism along an onshore-offshore gradient. Paleobiology 37, 270–286. ( 10.1666/10032.1) [DOI] [Google Scholar]
- 58.Sepkoski JJ, Sheehan PM. 1983. Diversification, faunal change, and community replacement during the Ordovician radiation. In Biotic interactions in Recent and fossil benthic communities (eds Tevesz MJS, McCall PL), pp. 673–718. New York, NY: Plenum. [Google Scholar]
- 59.Jablonski D, Sepkoski JJ, Bottjer DJ, Sheehan PM. 1983. Onshore-offshore patterns in the evolution of Phanerozoic shelf communities. Science 222, 1123–1125. ( 10.1126/science.222.4628.1123) [DOI] [PubMed] [Google Scholar]
- 60.Sepkoski JJ, Miller AI. 1985. Evolutionary faunas and the distribution of Paleozoic benthic communities in space and time. In Phanerozoic diversity patterns (ed. Valentine JW.), pp. 153–190. Princeton, NJ: Princeton University Press. [Google Scholar]
- 61.Jablonski D, Bottjer DJ. 1990. Onshore-offshore trends in marine invertebrate evolution. In Causes of evolution: a paleontologic perspective (eds Ross RM, Allmon WD), pp. 21–75. Chicago, IL: University of Chicago Press. [Google Scholar]
- 62.Whittaker RH. 1967. Gradient analysis of vegetation. Biol. Rev. 49, 207–264. ( 10.1111/j.1469-185X.1967.tb01419.x) [DOI] [PubMed] [Google Scholar]
- 63.Ohmann JL, Spies TA. 1998. Regional gradient analysis and spatial pattern of woody plant communities in Oregon forests. Ecol. Monogr. 68, 151–182. ( 10.1890/0012-9615(1998)068%5B0151:RGAASP%5D2.0.CO;2) [DOI] [Google Scholar]
- 64.DiMichele WA, Cecil CB, Montañez IP, Falcon-Lang HJ. 2010. Cyclic changes in Pennsylvanian paleoclimate and effects on floristic dynamics in tropical Pangaea. Int. J. Coal Geol. 83, 329–344. ( 10.1016/j.coal.2010.01.007) [DOI] [Google Scholar]
- 65.Gastaldo RA, Demko TM. 2010. The relationship between continental landscape evolution and the plant-fossil record: long term hydrologic controls on preservation. Top. Geobiol. 32, 249–285. ( 10.1007/978-90-481-8643-3_7) [DOI] [Google Scholar]
- 66.Donoghue MJ, Edwards EJ. 2014. Biome shifts and niche evolution in plants. Annu. Rev. Ecol. Evol. S. 45, 547–572. ( 10.1146/annurev-ecolsys-120213-091905) [DOI] [Google Scholar]
- 67.Hunt AE, Newell AJ. 1996. Taphofacies and early dinosaur evolution: an example from the Bull Canyon Formation (Upper Triassic: Norian), east-central New Mexico. New Mex. Geol. 18, 55. [Google Scholar]
- 68.Rogers RR, Kidwell SM. 2000. Associations of vertebrate skeletal concentrations and discontinuity surfaces in continental and shallow marine records: a test in the Cretaceous of Montana. J. Geol. 108, 131–154. ( 10.1086/314399) [DOI] [PubMed] [Google Scholar]
- 69.Greenwood DR. 1991. The taphonomy of plant macrofossils. In The processes of fossilization (ed. Donovan SK.), pp. 141–169. New York, NY: Columbia University Press. [Google Scholar]
- 70.Johnson ME, Skinner DF, MacLeod KG. 1988. Ecological zonation during the carbonate transgression of a Late Ordovician rocky shore (northeastern Manitoba, Hudson Bay, Canada). Palaeogeogr. Palaeoclimatol. Palaeoecol. 65, 93–114. ( 10.1016/0031-0182(88)90115-0) [DOI] [Google Scholar]
- 71.Kidwell SM, Jablonski D. 1983. Taphonomic feedback: ecological consequences of shell accumulation. In Biotic interactions in recent and fossil benthic communities (eds Tevesz MJS, McCall PL), pp. 195–248. New York, NY: Plenum. [Google Scholar]
- 72.Kidwell SM. 1991. Taphonomic feedback (live/dead interactions) in the genesis of bioclastic beds: keys to reconstructing sedimentary dynamics. In Cycles and events in stratigraphy (eds Einsele G, Ricken W, Seilacher A), pp. 268–282. Berlin, Germany: Springer. [Google Scholar]
- 73.Tomas˘ových A, Fürsich FT, Wilmsen M. 2006. Preservation of autochthonous shell beds by positive feedback between increased hardpart-input rates and increased sedimentation rates. J. Geol. 114, 287–312. ( 10.1086/501220) [DOI] [Google Scholar]
- 74.Miller AI. 1997. A new look at age and area: the geographic and environmental expansion of genera during the Ordovician Radiation. Paleobiology 23, 410–419. ( 10.1017/S0094837300019813) [DOI] [PubMed] [Google Scholar]
- 75.Foote M, Crampton JS, Beu AG, Marshall BA, Cooper RA, Maxwell PA, Matcham I. 2007. Rise and fall of species occupancy in Cenozoic fossil mollusks. Science 318, 1131–1134. ( 10.1126/science.1146303) [DOI] [PubMed] [Google Scholar]
- 76.Liow LH, Stenseth NC. 2007. The rise and fall of species: implications for macroevolutionary and macroecological studies. Proc. R. Soc. B 274, 2745–2752. ( 10.1098/rspb.2007.1006) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jones KE, Sechrest W, Gittleman JL. 2005. Age and area revisited: identifying global patterns and implications for conservation. In Phylogeny and conservation (eds Purvis A, Gittleman JL, Brooks TM), pp. 141–165. New York, NY: Cambridge University Press. [Google Scholar]
- 78.Wilson JT. 1966. Did the Atlantic close and then re-open? Nature 211, 676–681. ( 10.1038/211676a0) [DOI] [Google Scholar]
- 79.Klein G, Hsui AT. 1987. Origin of cratonic basins. Geology 15, 1094–1098. ( 10.1130/0091-7613(1987)15%3C1094:OOCB%3E2.0.CO;2) [DOI] [Google Scholar]
- 80.Meyers SR, Peters SE. 2011. A 56 million year rhythm in North American sedimentation during the Phanerozoic. Earth Planet. Sci. Lett. 303, 174–180. ( 10.1016/j.epsl.2010.12.044) [DOI] [Google Scholar]
- 81.Plotnick RE, Kenig F, Scott AC. 2015. Using the voids to fill the gaps: caves, times, and stratigraphy. Geol. Soc. Lond. Spec. Pub. 404, 233–250. ( 10.1144/SP404.5) [DOI] [Google Scholar]
- 82.Parsons B. 1982. Causes and consequences of the relation between area and age of the ocean floor. J. Geophys. Res. 87, 289–302. ( 10.1029/JB087iB01p00289) [DOI] [Google Scholar]
- 83.Rowley DB. 2002. Rate of plate creation and destruction: 180 Ma to present. Geol. Soc. Am. Bull. 114, 927–933. ( 10.1130/0016-7606(2002)114%3C0927:ROPCAD%3E2.0.CO;2) [DOI] [Google Scholar]
- 84.Smith AB, Gale AS, Monks NEA. 2001. Sea-level change and rock-record bias in the Cretaceous: a problem for extinction and biodiversity studies. Paleobiology 27, 241–253. ( 10.1666/0094-8373(2001)027%3C0241:SLCARR%3E2.0.CO;2) [DOI] [Google Scholar]
- 85.Valentine JW, Jablonski D, Krug AZ, Berke SK. 2012. The sampling and estimation of marine paleodiversity patterns: implications of a Pliocene model. Paleobiology 39, 1–20. ( 10.1666/0094-8373-39.1.1) [DOI] [Google Scholar]
- 86.Strauss D, Sadler PM. 1989. Classical confidence intervals and Bayesian probability estimates for ends of local taxon ranges. Math. Geol. 21, 411–427. ( 10.1007/BF00897326) [DOI] [Google Scholar]
- 87.Marshall CR. 1990. Confidence intervals on stratigraphic ranges. Paleobiology 16, 1–10. [Google Scholar]
- 88.Paul CRC. 1982. The adequacy of the fossil record. In Problems of phylogenetic reconstruction (eds Joysey KA, Friday AE), pp. 75–117. New York, NY: Academic Press. [Google Scholar]
- 89.McKinney ML. 1986. Biostratigraphic gap analysis. Geology 14, 36–38. ( 10.1130/0091-7613(1986)14%3C36:BGA%3E2.0.CO;2) [DOI] [Google Scholar]
- 90.Thode HJ. 2002. Testing for normality. New York, NY: Marcel Dekker. [Google Scholar]
- 91.Razali NM, Wah YB. 2011. Power comparisons of Shapiro–Wilk, Komogorov–Smirnov, Lilliefors and Anderson–Darling tests. J. Stat. Mod. Anal. 2, 21–33. [Google Scholar]
- 92.Marshall CR. 2008. A simple method for bracketing absolute divergence times on molecular phylogenies using multiple fossil calibration points. Am. Nat. 171, 726–742. ( 10.1086/587523) [DOI] [PubMed] [Google Scholar]
- 93.Wang SC, Zimmerman AE, McVeigh BS, Everson PJ, Wong H. 2012. Confidence intervals for the duration of a mass extinction. Paleobiology 38, 265–277. ( 10.1666/11016.1) [DOI] [Google Scholar]
- 94.Alroy J. 2014. A simple Bayesian method of inferring extinction. Paleobiology 40, 584–607. ( 10.1666/13074) [DOI] [Google Scholar]
- 95.Marshall CR. 1997. Confidence intervals on stratigraphic ranges with nonrandom distributions of fossil horizons. Paleobiology 23, 165–173. [Google Scholar]
- 96.Stadler T, Künert D, Bonhoeffer S, Drumond AJ. 2013. Birth–death skyline plot reveals temporal changes of epidemic spread in HIV and hepatitis C virus (HCV). Proc. Natl Acad. Sci. USA 110, 228–233. ( 10.1073/pnas.1207965110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Liow LH, Finarelli JA. 2014. A dynamic global equilibrium in carnivoran diversification over 20 million years. Phil. Trans. R. Soc. B 281, 20132312 ( 10.1098/rspb.2013.2312) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Raup DM. 1976. Species diversity in the Phanerozoic: an interpretation. Paleobiology 4, 1–15. [Google Scholar]
- 99.Smith AB. 2001. Large-scale heterogeneity of the fossil record: implications for Phanerozoic biodiversity studies. Phil. Trans. R. Soc. B 356, 351–367. ( 10.1098/rstb.2000.0768) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Peters SE. 2006. Macrostratigraphy of North America. J. Geol. 114, 391–412. ( 10.1086/504176) [DOI] [Google Scholar]
- 101.Hannisdal B, Peters SE. 2011. Phanerozoic Earth system evolution and marine biodiversity. Science 334, 1121–1124. ( 10.1126/science.1210695) [DOI] [PubMed] [Google Scholar]
- 102.Smith AB, Benson RBJ. 2013. Marine diversity in the geological record and its relationship to surviving bedrock area, lithofacies diversity, and original marine shelf area. Geology 41, 171–174. ( 10.1130/G33773.1) [DOI] [Google Scholar]
- 103.Sansom RS, Randle E, Donoghue PCJ. 2015. Discriminating signal from noise in the fossil record of early vertebrates reveals cryptic evolutionary history. Proc. R. Soc. B 282, 20142245 ( 10.1098/rspb.2014.2245) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Peters SE, Heim NA. 2010. The geological completeness of paleontological sampling in North America. Paleobiology 36, 61–79. ( 10.1666/0094-8373-36.1.61) [DOI] [Google Scholar]
- 105.Allulee JL, Holland SM. 2005. The sequence stratigraphic and environmental context of primitive vertebrates: harding Sandstone, Upper Ordovician, Colorado, USA. Palaios 20, 518–533. ( 10.2110/palo.2003.p03-120) [DOI] [Google Scholar]
- 106.Liu HP, McKay RM, Young JN, Witzke BJ, McVey KJ, Liu X. 2006. A new Lagerstätte from the Middle Ordovician St. Peter Formation in northeast Iowa, USA. Geology 34, 969–972. ( 10.1130/G22911A.1) [DOI] [Google Scholar]
- 107.Davies NS, Sansom IJ. 2009. Ordovician vertebrate habitats: a Gondwanan perspective. Palaios 24, 717–722. ( 10.2110/palo.2009.p09-040r) [DOI] [Google Scholar]
- 108.Silvestro D, Schnitzler J, Liow LH, Antonelli A, Salamin N. 2014. Bayesian estimation of speciation and extinction from incomplete fossil occurrence data. Syst. Biol. 63, 349–367. ( 10.1093/sysbio/syu006) [DOI] [PMC free article] [PubMed] [Google Scholar]