

Abstract
The characterization and potential of mesenchymal stem cells (MSCs) are gender dependent and estrogen influences these properties. This study demonstrated that supplementation with 17β-estradiol (E2) increases the proliferation of bone marrow-MSCs derived from male and female mini-pigs (Mp- and Fp-BMSCs) in a concentration-dependent manner, with 10-12 M E2 suggested as the optimal dose of E2 that led to the greatest improvement in BMSCs proliferation. Supplementation of 10-12 M E2 resulted in down-regulation of β-galactosidase activity and pro-apoptotic activity in both BMSCs, while anti-apoptotic activity was up-regulated in only Fp-BMSCs. Further, E2 increased the osteogenic ability of Fp-BMSCs. Based on these findings, optimal utilization of E2 can improve cellular senescence and apoptosis, as well as in vitro osteogenesis of BMSCs, and could therefore be useful in stem cell therapy, particularly in bone regeneration for adult females.
Keywords: 17 beta-estradiol, gender differences, mesenchymal stem cells, mini-pig, senescence
Introduction
Multipotent mesenchymal stem cells (MSCs) can differentiate into mesenchymal cell lineages (e.g., osteogenic, adipogenic and chondrogenic [17,20]), which makes them a promising candidate for tissue engineering and cellular therapies [20,29]. MSCs derived from bone marrow (BMSCs) are considered a potential cell source in stem cell therapy because of their capacity for self-renewal and differentiation into multiple lineages. However, aged donors do not have a sufficient population of MSCs for efficient autologous transplantation. Although in vitro proliferation can amplify the cell population, excessive duplication can significantly reduce the differentiation potential and tissue formation ability [17]. Therefore, effective enhancement of the differentiation potential of MSCs is necessary for MSC-based tissue engineering.
The cells most commonly used for tissue engineering therapy are BM-MSCs, but these might be considerably affected by gender and estrogen levels [2,7,25,27,31]. Indeed, gender differences have found to have pathophysiological effects and to influence the differentiation ability of stem cells [7,16].
MSCs are novel therapeutic agents for organ protection, and estrogen may enhance the protective function of MSCs by altering cytokine and growth factor production by these cells [8]. Estrogen is a sex steroid hormone that plays multifunctional roles and influences development, metabolism, aging and differentiation in various tissues in both females and males. Moreover, it is known that estrogen is one of the key factors involved in regulation of bone formation and is strongly associated with mitogenesis and proliferation of MSCs [10].
Estrogen receptors have been found to be distributed differently in various tissues of the body based on gender [12]. These differences seem to contribute to the sex dimorphism of steroid functions in these tissues. Gender-based differences in MSCs have been reported for muscle-derived stem cells from mouse and adipose-derived stem cells from humans [2,7], but the regulatory effects of 17β-estradiol (E2) have not been discussed in detail. Therefore, differences in the effects of estrogen on MSCs derived from males and females should be considered, since the sex steroid hormone and regulatory mechanism differ between genders. Pigs have valuable advantages as species
Pigs have valuable advantages as species for stem cell therapy because of their physiologic, genetic, and immunologic similarities to humans [28,30]. In particular, the mini-pig can be used as a preclinical animal model to improve understanding of stem cell physiology and a promising donor species for xenotransplantation by producing size-matched organs [6,28]. Thus, we utilized the mini-pig as an MSC donor to improve our understanding of the effects of stem cell properties and to characterize MSCs by E2 for various clinical applications.
This study was conducted to investigate the effects of E2 based on the variable properties of porcine BMSCs at the cellular and molecular levels to determine how they influence proliferation, cellular senescence, apoptosis and in vitrodifferentiation. Furthermore, differences in the regulatory effects of E2 were evaluated based on the gender of porcine BMSC donors. The results presented herein will help to improve the proliferation and in vitro differentiation potential of MSCs under estrogen regulation.
Materials and Methods
Chemicals and media
All chemicals were purchased from Sigma Chemical Company (USA) and all media was acquired from Gibco (USA), unless otherwise specified. Culture medium for BMSCs consisted of advanced Dulbecco's modified Eagle medium (ADMEM) supplemented with 10% charcoal-stripped fetal bovine serum (FBS), 10 ng/mL basic fibroblast growth factor (bFGF), 1.0% GlutaMax (Gibco) and 1.0% penicillinstreptomycin (10,000 IU and 10,000 µg/mL, respectively; Gibco). The osmolarity of the media was adjusted to 285 ± 5 mOsm/kg, and the pH was adjusted to 7.2.
Isolation and culture of cells
All experiments were authorized by the Animal Center for Biomedical Experimentation at Gyeongsang National University. For isolation of MSCs derived from the bone marrow of male donors (Mp-BMSCs) and female donors (Fp-BMSCs), bone marrow was obtained from three sexually mature 1-year-old male and female mini-pigs (T-type, PWG Micro-pig; PWG Genetics Korea, Korea) under standard surgical procedures. Gelatinous bone marrow was extracted by processing iliac crest bone biopsies and isolation under steroid-free conditions, as previously described [22]. Cells were cultured in medium with 0, 10-6, 10-8, 10-10, 10-12 and 10-14 M 17β-estradiol (Sigma, UK) supplementations at 38.5℃ in a humidified atmosphere of 5% CO2 in air by changing the medium every 3 days. Cells were then re-grown until passage 3 for further experiments.
Analysis of cell surface markers
For phenotyping of Mp- and Fp-BMSCs, cell surface antigens of BMSCs at the third passage were analyzed by flow cytometer (BD FACS Calibur; Becton, Dickinson and Company, USA) analysis in three replicates, as previously described [23]. Samples were labeled with fluorescein isothiocyanate (FITC)-conjugated anti-CD29 (β1-integrin; 1 : 100; BD Biosciences, USA), anti-CD44 (hyaluronate receptor; 1 : 100; BD Biosciences), and anti-CD45 (leukocyte common antigen; 1 : 100; Santa Cruz Biotechnology, USA) antibodies.
Alkaline phosphatase activity
To detect alkaline phosphatase (AP) activity, BMSCs were fixed with 3.7% formaldehyde for 20 min, then stained with the AP chromogen kit (Promega, USA) for 15 to 30 min. Samples were then observed using an inverted microscope (Nikon Diaphot 300; Nikon, Japan).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay following E2 supplementation
A Vybrant MTT Cell Proliferation Assay Kit (Molecular Probes, USA) was used to evaluate the proliferation of BMSCs, following the manufacturer's protocols. Briefly, BMSCs were seeded in sterile 96-well plates at a density of 1 × 103 cells/mL, after which E2 serially diluted in steroid-free media was added in levels to give concentrations of 0, 10-6 M, 10-8 M, 10-10 M, 10-12 M and 10-14 M. At 24 h after seeding, 20 µL (5 mg/mL) of MTT solution was added to each well and the samples were incubated for 4 h at 38.5℃. The supernatant in the wells was then removed, 50 µL of dimethyl sulfoxide (DMSO) was added and the samples were subjected to 10 min of oscillation. Cell proliferation was then quantified using an MTT colorimetric assay and a VersaMax Microplate Reader (Molecular Devices, USA) at an absorbance of 540 nm. After establishing the optimal dose of E2 for improvement of proliferative activity, E2 treated MSCs were evaluated for apoptosis, cellular senescence and in vitro differentiation ability.
β-galactosidase staining
β-galactosidase (β-Gal) activity was used to evaluate cellular senescence using the mammalian β-Gal Assay Kit (Thermo Fisher Scientific, USA), and optical density (OD) was determined using a VersaMax Microplate Reader (Molecular Devices) at a wavelength of 480 nm.
Terminal deoxynucleotidyltransferase (TdT)-mediated dUTP-biotin nick-end labeling (TUNEL) staining
TUNEL staining was performed using an In Situ Cell Death Detection Kit, TMR red (Roche Diagnostics, Germany) according to the manufacturer's instructions. Briefly, cover slips with attached cells were fixed in 3.7% paraformaldehyde for 1 h, then permeabilized in 0.5% Triton X-100 for 1 h at room temperature. The cells were then incubated with TdT enzyme in reaction buffer containing TMR red-labeled dUTP at 37℃ for 1 h. The negative control was incubated in reaction buffer without TdT enzyme. The cells were counterstained with 1 µg/mL 4,6-diamidino 2-phenyindiol (DAPI), and the incidence of apoptosis was evaluated using fluorescence microscopy by counting the fragmented apoptotic bodies in the five sections of each slide.
In vitro differentiation and cytochemical staining
In vitro differentiation of BMSCs into adipocytes, osteocytes, and chondrocytes was evaluated by previously described protocols, with minor modifications [23]. Briefly, cells at 70 to 80% were induced into their mesenchymal lineages under conditions conducive for osteogenic, adipogenic, and chondrogenic differentiation for 21 days. The adipogenic differentiation medium consisted of ADMEM, 10% FBS, 1 µM dexamethasone, 10 µM insulin, 200 µM indomethacin and 500 µM 3-isobutyl-1-methylxanthine. Intracellular lipid droplets within differentiated adipocytes were identified by staining with 0.5% Oil Red O staining. The osteogenic differentiation medium consisted of ADMEM, 10% FBS, 0.1 µM dexamethasone, 50 µM ascorbate-2-phosphate, and 10 mM β-glycerol phosphate. The matrix mineralization and calcium deposits were detected by Alizarin-red S solution staining. For induction of chondrogenic differentiation, cells were cultured in STEMPRO Chondrogenesis Differentiation Kit media with 10% supplement (Invirogen). Chondrogenic differentiation was examined by staining with Alcian Blue 8 GX solution for synthesis of glycosaminoglycans.
Quantitative reverse-transcription polymerase chain reaction (qRT-PCR)
Real-time PCR based on the multiplex method was used to quantify the mRNA levels of genes involved in apoptosis and differentiation (adipocytes, osteocytes, and chondrocytes) in BMSCs cultured with (10-12 M) and without (0 M) E2 supplementation, as previously described [23]. Briefly, total RNA was isolated from the BMSCs using an RNeasy Mini Kit (Qiagen, USA), then quantified using an OPTIZEN 3220 UV BIO spectrophotometer (MECASYS, Korea) in accordance with the manufacturer's instructions. cDNA synthesis for analyzing genes was conducted using 1 µg of total RNA using the Omniscript Reverse Transcription Kit (Qiagen) with oligo dT primer (Invitrogen) at 60℃ for 1 h. Relative qRT-PCR was performed using a LightCyclerTM with FastStart DNA Master SYBR Green I (Roche Diagnostics), which consisted of 2 mM MgCl2, 2 µL SYBR Green, and 0.5 µM each of forward and reverse primers. The amplification program consisted of denaturation at 95℃ for 1 min, followed by 50 PCR cycles at 95℃ for 10 sec, 60℃ for 6 sec and 72℃ for 4 sec, melting curve from 65℃ to 95℃at 0.1℃ per sec and finally cooling at 40℃ for 30 sec. All transcriptional levels of target genes were normalized against a reference gene, hydroxymethylbilane synthase (HMBS). Detailed information regarding each specific primer is presented in Table 1.
Table 1. Primers used for gene specific quantitative reverse-transcription polymerase chain reaction (qRT-PCR).

Statistical analysis
One-way analysis of variance (ANOVA) (SPSS ver. 21; IBM, USA) with Duncan's and Tukey's multiple comparisons tests was used to identify differences among treatments. All data are expressed as the mean ± standard deviation (SD), and differences were considered significant when p-values were less than 0.05.
Results
Isolation and cell surface markers of MSCs
MSCs were successfully isolated from the bone marrow of sexually matured male and female mini-pigs, then cultured in vitro as previously described [22]. Homogeneous adherent MSCs with strong AP activity were found among the subcultured cells from the third passage (panel A in Fig. 1).
Fig. 1. Identification of alkaline phosphatase (AP) activity and cell surface markers in the bone marrow-mesenchymal stem cells (MSCs) derived from male and female mini-pigs (Mp- and Fp-BMSCs). (A) Mp- and Fp-BMSCs showed AP activity based on BCIP/NBT staining. (B) Cell surface markers of Mp- and Fp-BMSCs at passage 3 were identified, positive for CD29 and CD44 and negative for CD45 by flow cytometry analysis. Scale bar = 100 µm.

For phenotyping of Mp- and Fp-BMSCs, the cell surface antigen profiles of the MSCs were determined by flow cytometry analysis using specific markers; namely, antibodies against the integrin CD29 (β1-integrin), and matrix receptors CD44 (hyaluronate receptor) and CD45 (leukocyte common antigen). The cell-surface antigen profiles of both BMSCs were similar, and they were strongly positive for mesenchymal markers such as CD29 and CD44, but negative for hematopoietic markers such as CD45 (panel B in Fig. 1).
Effects of E2 on the proliferation of Mp- and Fp-BMSCs
The proliferation rates of Mp- and Fp-BMSCs with various concentrations of E2 (0, 10-6, 10-8, 10-10, 10-12, and 10-14 M) were evaluated by MTT assay (Fig. 2). The proliferation rates of Fp-BMSCs gradually increased with decreasing E2 concentration, peaking at 10-12 M E2. Moreover, the proliferation rate with 10-12 M E2 was significantly (p < 0.05) greater than that with all other concentrations (0, 10-6, 10-8, 10-10, and 10-14 M) of E2. However, the effects of E2 on the proliferation rate of BMSCs differed between Mp-BMSCs and Fp-BMSCs. For Mp-BMSCs, 10-6 and 10-8 M of E2 significantly (p < 0.05) inhibited proliferation, while lower concentrations (10-12 and 10-14 M) significantly (p < 0.05) increased the proliferation rate. Based on the overall results, the optimal dose for proliferative ability was found to be 10-12 M; therefore, this level was used for further evaluations.
Fig. 2. 17β-estradiol (E2) concentration-dependent cell proliferation in Mp- and Fp-BMSCs by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Cells were grown in steroid-free medium supplemented with different concentrations (0, 10-6, 10-8, 10-10, 10-12, and 10-14 M) of E2 for 14 days. The proliferation rates were quantified by measuring the absorbance at 540 nm. The black bars represent Mp-BMSCs, and the white bars represent Fp-BMSCs. The different superscript letters represent significant (p < 0.05) differences among the five replicates.

Effect of E2 on cellular senescence in Mp- and Fp-BMSCs
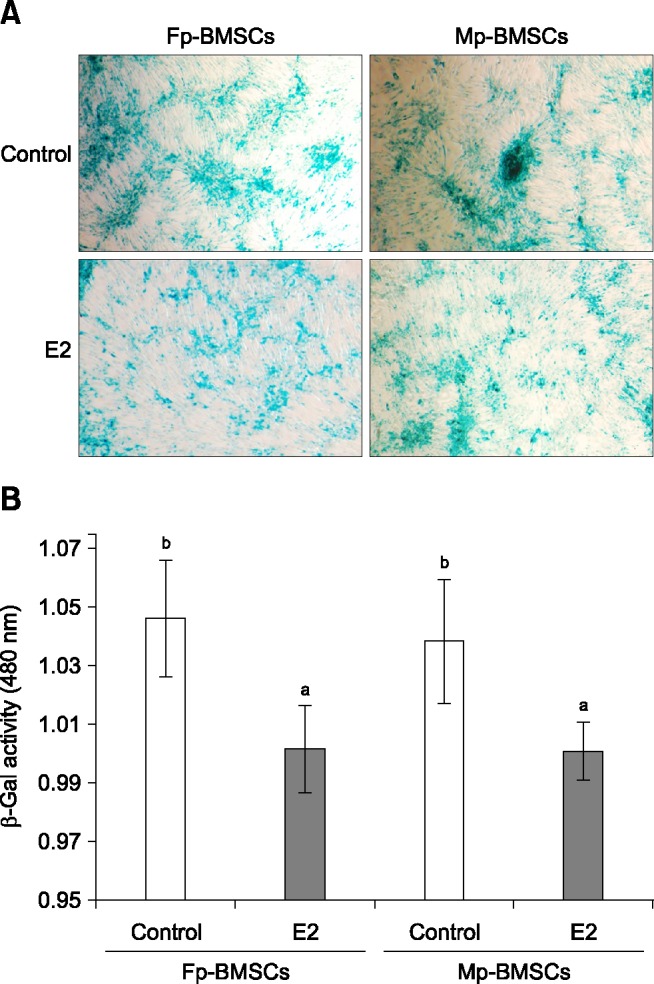
BMSCs cultured with 10-12 M E2 supplementation were compared based on cellular senescence determined from comparison of β-Gal activity between treatment groups and a control. Positive blue staining for β-Gal (associated with senescence) was detected in Mp- and Fp-BMSCs (panel A in Fig. 3), and was quantified by measuring the absorbance at 480 nm (panel B in Fig. 3). There was no difference in the cellular senescence-associated β-Gal activity in the absence or presence of E2 for either BMSCs. However, the quantified cellular senescence was significantly (p < 0.05) lower in Mp- and Fp-BMSCs cultured with 10-12 M E2 relative to those cultured with 0 M E2 (panel B in Fig. 3).
Fig. 3. Analysis of cellular senescence in Mp- and Fp-BMSCs following E2 supplementation using β-galactosidase (β-Gal) staining. (A) Detection of senescence-associated β-Gal staining of Mp- and Fp-BMSCs cultured in the control (0 M) and E2 (10-12 M). (B) Cell senescence was quantified in Mp- and Fp-BMSCs by measuring senescence-associated β-Gal staining at an absorbance of 405 nm. The white and black bars represent cells cultured in the control and E2, respectively. The different superscript letters represent significant (p < 0.05) differences. 200× (A).
Effect of E2 on the apoptosis of Mp- and Fp-BMSCs
Expression of apoptosis-related genes of Mp- and Fp-BMSCs in the absence and presence of 10-12 M E2 was determined by TUNEL staining (Fig. 4) and qRT-PCR (panel A and B in Fig. 5). We evaluated the ratio of apoptosis by using the combined criteria of TUNEL positivity and nuclear changes to quantify fragmented apoptotic bodies in five sections of each slide. In the 15 sections of each BMSC, both Mp- and Fp-BMSCs had 5% to 46% TUNEL-positive apoptotic bodies in the absence of E2. In contrast, TUNEL-positive apoptotic bodies were 4% to 25% in both BMSCs in the presence of E2. The mRNA levels of the anti-apoptosis (Bcl2 and Birc) and pro-apoptosis (Bax, Bak, and Hsp90) genes were quantitatively analyzed. In the absence of E2, the expression levels of Bcl2 and Birc were significantly (p < 0.05) different between Mp- and Fp-BMSCs (panel A in Fig. 5). However, upon comparison with the un-treated control, the expression of Bcl2 and Birc was significantly (p < 0.05) up-regulated in Fp-BMSCs in the presence of E2 (10-12 M), but no significant (p < 0.05) difference was observed in Mp-BMSCs (panel A in Fig. 5).
Fig. 4. Detection of apoptosis in Mp- and Fp-BMSCs following E2 (10-12 M) supplementation using terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. Nuclei were counterstained with 4,6-diamidino 2-phenyindiol (DAPI; blue), and fragmented apoptotic bodies (arrow) were stained with TUNEL dye (red). Scale bars = 100 µm.

Fig. 5. Quantitative mRNA analysis of anti- and pro-apoptosis genes in Mp- and Fp-BMSCs following E2 supplementation. Cells were cultured in the control (0 M) and E2 (10-12 M) until they reached confluence. Expression of the anti-apoptosis related Bcl2 and Birc (A) and the pro-apoptosis related Bax, Bak and Hsp90 (B) was determined by qRT-PCR. Each experiment was performed in triplicate, and HMBS was used as the internal control gene. Different superscript letters represent significant (p < 0.05) differences among different cells.

The mRNA levels of the pro-apoptosis genes Bax and Hsp90 also differed significantly (p < 0.05) between Mp- and Fp-BMSCs in the absence of E2. However, the expression of Bak did not differ between BMSCs. In the presence (10-12 M) of E2, the expression of pro-apoptosis genes, Bax, Bak, and Hsp90, was significantly (p < 0.05) down-regulated in both BMSCs compared with the absence (0 M) of E2.
Effect of E2 on the differentiation ability of Mp- and Fp-BMSCs
In vitro differentiation of Mp- and Fp-BMSCs was induced with various concentrations of E2 (0, 10-10, 10-12, and 10-14 M) for 21 days. Cytochemical staining verified that BMSCs from males and females progressed toward differentiation into adipocytes, osteocytes, and chondrocytes, as confirmed by the cytoplasmic accumulation of lipid vacuoles, the deposition of calcified extracellular matrix, and synthesis of glycosaminoglycans by Oil red O, Alizarin red S, and Alcian blue 8GX staining, respectively. In vitro differentiation into adipocytes, osteocytes, and chondrocytes was induced by various concentrations of E2 (0, 10-10, 10-12, and 10-14 M), and both BMSCs stained positive for these cell types with all E2 concentrations (Fig. 6). The mRNA levels of adipocyte-, osteocyte-, and chondrocyte-related genes were comparatively quantified between Mp- and Fp-BMSCs cultured in the absence and presence (10-12 M) of E2 (Figs. 7, 8, 9).
Fig. 6. Cytochemical staining of Mp- and Fp-BMSCs in which differentiation was induced with various concentrations of E2. (A) Adipogenic induction in the E2-supplemented BMSCs was determined by Oil-red O staining of neutral lipid vacuoles, which showed intracellular accumulation. (B) Osteogenic induction in E2-supplemented BMSCs was determined by Alizarin-red S solution staining for identification of the mineralized matrix. (C) Chondrogenic induction in the E2-supplemented BMSCs was determined by Alcian blue 8GX solution staining for synthesis of glycosaminoglycans. The concentrations of E2 used were 0, 10-10, 10-12, and 10-14 M in the adipogenic, osteogenic, and chondrogenic induction medium. Scale bars = 100 µm.

Fig. 7. Relative quantification of adipogenic differentiation-related genes in Mp- and Fp-BMSCs following supplementation of E2 (10-12 M). Adipogenic differentiation was induced for 21 days and was demonstrated by the expression of adipocyte protein 2 (AP2), lipoprotein lipase (LPL), and fatty acid binding protein (FABP) by qRT-PCR. Five replicates were used, and HMBS was employed as an internal control gene. Error bars with asterisks indicate a significant (p < 0.05) differences between the undifferentiated control and differentiated cells.

Fig. 8. Relative quantification of osteogenic differentiation-related genes in Mp- and Fp-BMSCs following supplementation of E2 (10-12 M). Osteogenic differentiation was induced for 21 days and was demonstrated by the expression of osteonectin (ON), Runt-related transcription factor 2 (RUNX2), and biglycan (BG) by qRT-PCR. Five replicates were used, and HMBS was employed as an internal control gene. Error bars with asterisks indicate a significant (p < 0.05) difference between the undifferentiated control and differentiated cells. O, osteogenic differentiated cells.

Fig. 9. Relative quantification of chondrogenic differentiation-related genes in Mp- and Fp-BMSCs following supplementation with E2 (10-12 M). Chondrogenic differentiation was induced for 21 days and was evidenced by the expression of aggrecan (ACAN), collagen type2 (COL2), and collagen type10, alpha 1 (COL10A1) by qRT-PCR. Five replicates were used, and HMBS was employed as an internal control gene. Error bars with asterisks indicate a significant (p < 0.05) difference between the undifferentiated control and differentiated cells. C, chondrogenic differentiated cells.

Adipocyte-related genes, adipocyte fatty acid binding protein (AP2), lipoprotein lipase (LPL), and fatty acid binding protein (FABP) in differentiated Fp-BMSCs and Mp-BMSCs showed increased expression levels compared to undifferentiated BMSCs cultured in the absence and presence (10-12 M) of E2, respectively (Fig. 7). However, the mRNA levels of AP2, LPL, and FABP in Fp-BMSCs were significantly (p < 0.05) lower in the presence of E2 than in its absence after adipogenic differentiation. In contrast, the mRNA levels of these three genes in Mp-BMSCs were significantly (p < 0.05) higher in the presence of E2 than in its absence.
In the presence or absence of 10-12 M E2, the mRNA levels of osteonectin (ON), runt-related transcription factor 2 (RUNX2), and biglycan (BG) were significantly (p < 0.05) up-regulated following induction of osteogenic differentiation compared with un-differentiated BMSCs (Fig. 8). However, the mRNA levels of these three genes in osteogenic differentiation-induced Fp-BMSCs were significantly (p < 0.05) higher in the presence of E2 than in its absence. Following induction of osteogenic differentiation of Mp-BMSCs, the mRNA levels of these three genes were not significantly (p < 0.05) different in the presence and absence of E2.
The mRNA levels of the chondrocyte-related genes, aggrecan (ACAN), collagen type 2 (COL2), and collagen type X alpha 1 (COL10A1), were significantly (p < 0.05) up-regulated in both BMSCs in the presence and absence of 10-12 M E2 after induction of chondrogenic differentiation (Fig. 9). Following induction of chondrogenic differentiation in both BMSCs, the mRNA levels of these three genes were higher in the presence of E2 than in its absence, but the difference was not significant.
Discussion
The present study investigated the regulatory effects of E2 in in vitro cultivation and induction of differentiation according to the gender of BMSC donors. The pig is known to be physiologically and genetically similar to humans, making it a promising donor species for xenotransplantation [6]. Moreover, generation of BMSCs from pigs has been extensively reported by many researchers [11,23], and they have been found to be morphologically and immune-phenotypically similar to human MSCs [29]. However, the gender-specific effects of cell proliferation, cellular senescence, apoptosis and in vitro differentiation ability on E2 have not yet been reported in mini-pig or pig-derived MSCs yet.
In this study, cell proliferation of PBMCs varies depending on the E2 concentrations, with the proliferation rates of Mp- and Fp-BMSCs gradually increasing with decreases in E2 concentration and peaking at 10-12 M-10-14 M and 10-12 M, respectively. These regulatory patterns are suggested for its strong mitogenic effect of E2, which could be explained by the biphasic action of steroids, in which they lose their function and even inhibit cell proliferation of human- and rat-derived BMSCs at high concentrations [15,16]. Moreover, the range of E2 doses that significantly increase the proliferation of Fp-BMSCs (10-6 M-10-14 M) is wider than that for Mp-BMSCs (10-12 M-10-14 M). E2 plays a role in increasing the cell proliferation in Mp- and Fp-BMSCs, which is to other species-derived BMSCs, but the effective doses of E2 vary by species or gender of the donors. Female rat-derived BMSCs showed increased cell proliferation in response to 10-10 M-10-12 M E2, whereas male rat-derived BMSCs were not affected by E2 [16]. In addition, among the BMSCs derived from different physiological statuses of rats, ovariectomized female rat-derived BMSCs showed increasing colony-forming unit-fibroblast (CFU-F) activity and number of cells in response to treatment with 10-7 M E2 [3]. Therefore, the dose-dependent effects of E2 on MSCs from female donors could depend on the physiological status of the female donor. Further, E2 appears to have a proliferative effect on BMSCs derived from male and female donors.
The results of the present study indicated that 10-12 M E2 was the optimal concentration for improving cell proliferation in both BMSCs, where this optimal concentration of E2 could play important roles in other cellular processes such as cellular senescence and apoptosis. Cell proliferation and apoptosis are opposite cellular processes [13]; however, both are physiologically associated and regulated for important cellular senescence, known as a permanent cell proliferation arrest. Moreover, cellular senescence and apoptosis are increased by long-term culture or physiological aging of donor MSCs, where proliferation ability decreases depending on the cell type. Stress responses in males and females are reportedly different, which may be due to differences in apoptosis ability in both genders [19], where E2 can increase the resistance to oxidative stress [4].
We observed that 10-12 M E2 was able to improve proliferation, which was able to inhibit cellular senescence in both gender-derived BMSCs from mini-pigs during in vitro culture. We also conducted TUNEL staining, which demonstrated that the ratio of apoptosis decreased in both confluent BMSCs in the presence of E2. However, it is not possible to statistically quantify the ratio of apoptotic body fragmentation, despite the strong correlation between TUNEL positivity and nuclear fragmentation and condensation, which are characteristics of apoptosis. Hence, we quantitatively analyzed the mRNA levels of apoptosis-related genes in the Mp- and Fp-BMSCs following E2 supplementation. In the present study, the mRNA levels of apoptosis-related genes were gender-dependently regulated in both growth-arrested confluent BMSCs in response to supplementation with 10-12 M E2. In addition, the basal mRNA levels of apoptosis- and stress-related genes were found to differ significantly between Mp- and Fp-BMSCs in the absence of E2 (with the exception of Bak). These differences in mRNA levels between BMSCs indicated that the expression of apoptosis-related genes was specifically dependent on the donor gender. Bcl2 is the best characterized gene of the anti-apoptotic Bcl2 family that has been associated with the prevention of cell death initiation [21]. The regulatory functions of Bcl2 showed up-regulated expression in female rat-derived MSCs in response to 10-7 M E2 supplementation. Moreover, increased apoptosis was observed in anti-apoptotic Bcl2-knockdown MSCs in contrast to normal MSCs following E2 supplementation [3], while the pro-apoptotic Bcl2 family members Bax and Bak showed a regulatory function of apoptosis during development and tissue homeostasis [24]. According to these results, the regulatory effects of E2 on the anti-apoptotic Bcl2 gene of Fp-BMSCs were similar to those of previous studies. These data suggest an important role for E2 in inhibiting cellular senescence, pro-apoptosis and stress in growth-arrested confluent Mp- and Fp-BMSCs. E2 also possesses the regulatory ability to enhance the anti-apoptosis genes, Bcl2 and Birc, in Fp-BMSCs. In contrast, E2 was not able to increase the anti-apoptosis genes in Mp-BMSCs, despite the effects of E2, which can play a significant role in regulation to reduce cellular senescence and improve cell proliferation.
Previous studies have shown gender differences in the differentiation ability of stem cells [2,7,16]. In skeletal muscle-derived stem cells from male mice, the capacity for osteogenic differentiation and bone regeneration was found to be greater than that of female-derived stem cells [7]. Further, in humans, male adipose-derived stem cells were found to have greater osteogenic potential than stem cells from females [2]. Hence, the osteogenic potential of female-derived MSCs is lower than that of male-derived MSCs, despite the higher prevalence of bone and cartilage diseases in women. Estrogen is one of the positive key regulators of bone mass in vivo [29], which improves bone formation by inhibiting bone resorption [27]. Recent studies reported that estrogen plays a reciprocal function in adipogenesis and osteogenesis of osteoblast cells and mouse BMSCs [9,14]. However, differences in the in vitro regulatory effects of E2 between BMSCs derived from different genders are not well known.
In this study, adipogenic, osteogenic, and chondrogenic Mp- and Fp-BMSCs showed strong positive staining for specific cell lineages following supplementation with various concentrations of E2 (0, 10-10, 10-12, and 10-14 M). Interestingly, 10-12 M E2 decreased the levels of AP2, LPL, and FABP in adipogenic-induced Fp-BMSCs. However, the mRNA levels of these genes were more elevated than in Mp-BMSCs. Conversely, E2 was shown to increase the adipogenic ability in Mp-BMSCs. The osteogenic differentiation ability in Fp-BMSCs was increased in the presence of E2 (10-12 M), but no distinct influence was found in Mp-BMSCs. Thus, the osteogenesis-related ON, RUNX2, and BG genes were up-regulated by supplementation with E2 in Fp-BMSCs. As a result, E2 differently regulates in vitro adipogenic and osteogenic differentiation in BMSCs derived from different genders. In the present study, E2 was found to improve or effectively regulate the osteogenic differentiation potential and to inhibit the adipogenic differentiation of Fp-BMSCs in vitro. Similar to the results observed Fp-BMSCs, E2 supplementation has been reported to improve the osteogenic potential of mouse and human female bone marrow stromal cells and osteoblasts by inhibiting adipogenesis [9,14].
In chondrogenic differentiation, E2 supplementation improved the capacity of BMSCs derived from males and females, but was not significantly enhanced, despite the effective up regulation of ACAN, COL10A1, and COL2. Similarly, E2 did not significantly influence in vitro chondrogenic differentiation in rabbit MSCs [1] or bovine articular chondrocytes [5]. Conversely, E2 was reported to be a strong negative predictor of cartilage regeneration after BMSC- and chondrocyte-based transplantation [25]. In BMSCs from young men, E2 supplementation showed dose-dependent inhibition of 3D chondrogenesis and reduced COL2 deposition, with 10-8 M E2 being found to be the most effective concentration [18]. However, E2 has been shown to influence cartilage under certain physiological and pathological conditions, even though cartilage is known to be a sex hormone-sensitive tissue. Furthermore, estrogen replacement therapy reduced the loss of articular cartilage in postmenopausal women [26]. Therefore, it is difficult to find a correlation between the in vivo effects of E2 and its vitro modulation of chondrogenesis in BMSCs.
Overall, our findings support those of previous studies that investigated rat BMSCs [16] and gender-based studies that suggest that E2 not only regulates osteogenic differentiation and maintains bone formation markers [31], but also improves cellular senescence and apoptosis in BMSCs from female donors. The outcomes of the present study strongly indicate that E2 has the potential for use as an effective modulator to improve the capacity of MSCs from females, especially with cellular aging and bone regeneration. The results presented herein also provide crucial information for improving and optimizing MSCs for future clinical applications in adult or aged donors. Because of differences in the responses to proliferation, apoptosis rate and adipo/osteogenic capacities between male and female-derived BMSCs, it is necessary to find novel biomarkers and gender-based therapeutic approaches. The in vitro results of this study will help improve differentiation potentials and quality of BMSCs for future in vivo applications.
Acknowledgments
We gratefully acknowledge financial support from the Next-Generation BioGreen 21, Rural Development Administration (grant No. PJ007969) and National Research Foundation (NRF) of Korea (grant No. NRF-2015R1D1A1A 01056639).
Footnotes
Conflict of Interest: There is no conflict of interest.
References
- 1.Ab-Rahim S, Selvaratnam L, Kamarul T. The effect of TGFβ-1 and β-estradiol on glycosaminoglycan and type II collagen distribution in articular chondrocyte cultures. Cell Biol Int. 2008;32:841–847. doi: 10.1016/j.cellbi.2008.03.016. [DOI] [PubMed] [Google Scholar]
- 2.Aksu AE, Rubin JP, Dudas JR, Marra KG. Role of gender and anatomical region on induction of osteogenic differentiation of human adipose-derived stem cells. Ann Plast Surg. 2008;60:306–322. doi: 10.1097/SAP.0b013e3180621ff0. [DOI] [PubMed] [Google Scholar]
- 3.Ayaloglu-Butun F, Terzioglu-Kara E, Tokcaer-Keskin Z, Akcali KC. The effect of estrogen on bone marrow-derived rat mesenchymal stem cell maintenance: inhibiting apoptosis through the expression of Bcl-xL and Bcl-2. Stem Cell Rev. 2012;8:393–401. doi: 10.1007/s12015-011-9292-0. [DOI] [PubMed] [Google Scholar]
- 4.Baba T, Shimizu T, Suzuki Y, Ogawara M, Isono K, Koseki H, Kurosawa H, Shirasawa T. Estrogen, insulin, and dietary signals cooperatively regulate longevity signals to enhance resistance to oxidative stress in mice. J Biol Chem. 2005;280:16417–16426. doi: 10.1074/jbc.M500924200. [DOI] [PubMed] [Google Scholar]
- 5.Claassen H, Schluter M, Schunke M, Kurz B. Influence of 17β-estradiol and insulin on type II collagen and protein synthesis of articular chondrocytes. Bone. 2006;39:310–317. doi: 10.1016/j.bone.2006.02.067. [DOI] [PubMed] [Google Scholar]
- 6.Cooper DK, Gollackner B, Sachs DH. Will the pig solve the transplantation backlog? Annu Rev Med. 2002;53:133–147. doi: 10.1146/annurev.med.53.082901.103900. [DOI] [PubMed] [Google Scholar]
- 7.Corsi KA, Pollett JB, Phillippi JA, Usas A, Li G, Huard J. Osteogenic potential of postnatal skeletal muscle-derived stem cells is influenced by donor sex. J Bone Miner Res. 2007;22:1592–1602. doi: 10.1359/jbmr.070702. [DOI] [PubMed] [Google Scholar]
- 8.Crisostomo PR, Wang Y, Markel TA, Wang M, Lahm T, Meldrum DR. Human mesenchymal stem cells stimulated by TNF-α, LPS, or hypoxia produce growth factors by an NF-κB but not JNK-dependent mechanism. Am J Physiol Cell Physiol. 2008;294:C675–C682. doi: 10.1152/ajpcell.00437.2007. [DOI] [PubMed] [Google Scholar]
- 9.Dang Z, Löwik CWGM. The balance between concurrent activation of ERs and PPARs determines daidzein-induced osteogenesis and adipogenesis. J Bone Miner Res. 2004;19:853–861. doi: 10.1359/JBMR.040120. [DOI] [PubMed] [Google Scholar]
- 10.Eriksen EF, Colvard DS, Berg NJ, Graham ML, Mann KG, Spelsberg TC, Riggs BL. Evidence of estrogen receptors in normal human osteoblast-like cells. Science. 1988;241:84–86. doi: 10.1126/science.3388021. [DOI] [PubMed] [Google Scholar]
- 11.Faast R, Harrison SJ, Beebe LFS, Mcilfatrick SM, Ashman RJ, Nottle MB. Use of adult mesenchymal stem cells isolated from bone marrow and blood for somatic cell nuclear transfer in pigs. Cloning Stem Cells. 2006;8:166–173. doi: 10.1089/clo.2006.8.166. [DOI] [PubMed] [Google Scholar]
- 12.Fadini GP, de Kreutzenberg S, Albiero M, Coracina A, Pagnin E, Baesso I, Cignarella A, Bolego C, Plebani M, Nardelli GB, Sartore S, Agostini C, Avogaro A. Gender differences in endothelial progenitor cells and cardiovascular risk profile: the role of female estrogens. Arterioscler Thromb Vasc Biol. 2008;28:997–1004. doi: 10.1161/ATVBAHA.107.159558. [DOI] [PubMed] [Google Scholar]
- 13.Guo M, Hay BA. Cell proliferation and apoptosis. Curr Opin Cell Biol. 1999;11:745–752. doi: 10.1016/s0955-0674(99)00046-0. [DOI] [PubMed] [Google Scholar]
- 14.Heim M, Frank O, Kampmann G, Sochocky N, Pennimpede T, Fuchs P, Hunziker W, Weber P, Martin I, Bendik I. The phytoestrogen genistein enhances osteogenesis and represses adipogenic differentiation of human primary bone marrow stromal cells. Endocrinology. 2004;145:848–859. doi: 10.1210/en.2003-1014. [DOI] [PubMed] [Google Scholar]
- 15.Hong L, Colpan A, Peptan IA. Modulations of 17-β estradiol on osteogenic and adipogenic differentiations of human mesenchymal stem cells. Tissue Eng. 2006;12:2747–2753. doi: 10.1089/ten.2006.12.2747. [DOI] [PubMed] [Google Scholar]
- 16.Hong L, Sultana H, Paulius K, Zhang G. Steroid regulation of proliferation and osteogenic differentiation of bone marrow stromal cells: a gender difference. J Steroid Biochem Mol Biol. 2009;114:180–185. doi: 10.1016/j.jsbmb.2009.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jaiswal N, Haynesworth SE, Caplan AI, Bruder SP. Osteogenic differentiation of purified, culture expanded human mesenchymal stem cells in vitro. J Cell Biochem. 1997;64:295–312. [PubMed] [Google Scholar]
- 18.Jenei-Lanzl Z, Straub RH, Dienstknecht T, Huber M, Hager M, Grässel S, Kujat R, Angele MK, Nerlich M, Angele P. Estradiol inhibits chondrogenic differentiation of mesenchymal stem cells via nonclassic signaling. Arthritis Rheum. 2010;62:1088–1096. doi: 10.1002/art.27328. [DOI] [PubMed] [Google Scholar]
- 19.Jog NR, Caricchio R. Differential regulation of cell death programs in males and females by Poly (ADP-ribose) polymerase-1 and 17β estradiol. Cell Death Dis. 2013;4:e758. doi: 10.1038/cddis.2013.251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Johnstone B, Hering TM, Caplan AI, Goldberg VM, Yoo JU. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp Cell Res. 1998;238:265–272. doi: 10.1006/excr.1997.3858. [DOI] [PubMed] [Google Scholar]
- 21.Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c from mitochondria: a primary site for Bcl-2 regulation of apoptosis. Science. 1997;275:1132–1136. doi: 10.1126/science.275.5303.1132. [DOI] [PubMed] [Google Scholar]
- 22.Lee SL, Kang EJ, Maeng GH, Kim MJ, Park JK, Kim TS, Hyun SH, Lee ES, Rho GJ. Developmental ability of miniature pig embryos cloned with mesenchymal stem cells. J Reprod Dev. 2010;56:256–262. doi: 10.1262/jrd.09-196a. [DOI] [PubMed] [Google Scholar]
- 23.Lee WJ, Hah YS, Ock SA, Lee JH, Jeon RH, Park JS, Lee SI, Rho NY, Rho GJ, Lee SL. Cell source-dependent in vivo immunosuppressive properties of mesenchymal stem cells derived from the bone marrow and synovial fluid of minipigs. Exp Cell Res. 2015;333:273–288. doi: 10.1016/j.yexcr.2015.03.015. [DOI] [PubMed] [Google Scholar]
- 24.Lindsten T, Ross AJ, King A, Zong WX, Rathmell JC, Shiels HA, Ulrich E, Waymire KG, Mahar P, Frauwirth K, Chen Y, Wei M, Eng VM, Adelman DM, Simon MC, Ma A, Golden JA, Evan G, Korsmeyer SJ, MacGregor GR, Thompson CB. The combined functions of proapoptotic Bcl-2 family members Bak and Bax are essential for normal development of multiple tissues. Mol Cell. 2000;6:1389–1399. doi: 10.1016/s1097-2765(00)00136-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nejadnik H, Hui JH, Feng Choong EP, Tai BC, Lee EH. Autologous bone marrow-derived mesenchymal stem cells versus autologous chondrocyte implantation: an observational cohort study. Am J Sports Med. 2010;38:1110–1116. doi: 10.1177/0363546509359067. [DOI] [PubMed] [Google Scholar]
- 26.Parker D, Hwa SY, Sambrook P, Ghosh P. Estrogen replacement therapy mitigates the loss of joint cartilage proteoglycans and bone mineral density induced by ovariectomy and osteoarthritis. APLAR J Rheumatol. 2003;6:116–127. [Google Scholar]
- 27.Qu Q, Perälä-Heape M, Kapanen A, Dahllund J, Salo J, Väänänen HK, Härkänen P. Estrogen enhances differentiation of osteoblasts in mouse bone marrow culture. Bone. 1998;22:201–209. doi: 10.1016/s8756-3282(97)00276-7. [DOI] [PubMed] [Google Scholar]
- 28.Sachs DH. The pig as a potential xenograft donor. Vet Immunol Immunopathol. 1994;43:185–191. doi: 10.1016/0165-2427(94)90135-x. [DOI] [PubMed] [Google Scholar]
- 29.Shukla D, Box GN, Edwards RA, Tyson DR. Bone marrow stem cells for urologic tissue engineering. World J Urol. 2008;26:341–349. doi: 10.1007/s00345-008-0311-y. [DOI] [PubMed] [Google Scholar]
- 30.Su J, Chen X, Huang Y, Li W, Li J, Cao K, Cao G, Zhang L, Li F, Roberts AI, Kang H, Yu P, Ren G, Ji W, Wang Y, Shi Y. Phylogenetic distinction of iNOS and IDO function in mesenchymal stem cell-mediated immunosuppression in mammalian species. Cell Death Differ. 2014;21:388–396. doi: 10.1038/cdd.2013.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Syed F, Khosla S. Mechanisms of sex steroid effects on bone. Biochem Biophys Res Commun. 2005;328:688–696. doi: 10.1016/j.bbrc.2004.11.097. [DOI] [PubMed] [Google Scholar]