

Abstract
Rationale
The importance for Bmp signaling during embryonic stem cell differentiation into myocardial cells has been recognized. The question when and where Bmp signaling in vivo regulates myocardial differentiation has remained largely unanswered.
Objective
To identify when and where Bmp signaling regulates cardiogenic differentiation.
Methods and Results
Here we have observed that in zebrafish embryos, Bmp signaling is active in cardiac progenitor cells prior to their differentiation into cardiomyocytes. Bmp signaling is continuously required during somitogenesis within the anterior lateral plate mesoderm to induce myocardial differentiation. Surprisingly, Bmp signaling is actively repressed in differentiating myocardial cells. We identified the inhibitory Smad6a, which is expressed in the cardiac tissue, to be required to inhibit Bmp signaling and thereby promote expansion of the ventricular myocardium.
Conclusion
Bmp signaling exerts opposing effects on myocardial differentiation in the embryo by promoting as well as inhibiting cardiac growth.
Keywords: BMP, Smad6, Heart, Zebrafish, Bmpr1a
The complex multichambered vertebrate heart is formed from a rapidly expanding primary heart tube due to the combined processes of cardiac differentiation and tissue morphogenesis.
In zebrafish, cardiac progenitor cells are located in the lateral marginal domain at both sides of the organizer of blastula stage embryos.1,2 During gastrulation, the bilateral pools of cardiac progenitor cells migrate dorsal and anterior to form part of the anterior lateral plate mesoderm (ALPM). In the ALPM, several transcription factors (nkx2.5, gata5, and hand2) are expressed that promote cardiac differentiation. The first progenitor cells that are determined to become cardiomyocyte in the ALPM can be identified at the 14-somite stage by expression of the cardiac differentiation marker cardiac myosin, light polypeptide 7 (myl7).3 Around 19 hours postfertilization (hpf) (20-somite stage), these 2 bilateral pools of cardiomyocytes have fused at the midline to form a cardiac disk.4,5 This cardiac disk rotates and extends in an anterior and leftward direction to form a linear heart tube.6 We previously described a continuous growth of the heart tube by the addition of cardiomyocytes to both the arterial and venous pole of the heart tube.7 At the arterial pole, Fgf signaling is required for the growth of the heart.7 At the venous pole we described that the transcription factor Isl1 is present in the differentiating cardiomyocytes, and embryos lacking Isl1 have a small reduction in the number of atrial cardiomyocytes.
Bmp ligands have been implicated in regulating cardiac differentiation in invertebrates as well as vertebrates.8–14 Bmp proteins are synthesized as large preproteins, which are cleaved to release the dimeric C-terminal mature and active region.15 Bmp2 and Bmp4 are closely related, and they can bind to their Bmp type 1 receptors (alk2/8, alk3a, alk3b, and alk6) as homo- or heterodimers.16–18 Interaction of the type 1 receptor with the Bmp type 2 receptor results in the phosphorylation of Smad transcriptional activators. In case of Bmp signaling there are 3 receptor-Smad proteins (Smad1, -5, and -8) that can be phosphorylated and interact with the co-Smad, Smad4. This phosphorylated Smad-protein complex is translocated to the nucleus where it activates gene expression. Secreted antagonists such as Noggin can block the interaction between Bmp ligand and the receptor and thereby prevent Bmp signaling, while the inhibitory Smad protein Smad6 attenuates Bmp signaling in the cytoplasm. Smad6 has been shown to specifically inhibit Bmp signaling but only weakly Tgf-β/activin signaling. Several mechanisms have been proposed for the inhibitory effect of Smad6 on Bmp signaling. Smad6 can bind to Smad1 and interfere with complex formation between Smad1 and Smad4.19 Smad6 can also form a stable complex with the Bmp type I receptors. This interaction can inhibit phosphorylation of Bmp-specific R-Smads and result in inhibition of Bmp signaling.20 Furthermore, Smad6 recruits the E3 ubiquitin ligase Smurf1 (Smad ubiquitin regulatory factor 1) to the signaling receptor complex and enhances the down-regulation of type I receptors through proteasome-dependent degradation.21
In this study, we investigated the role of Bmp signaling in cardiomyocyte differentiation. We observed that in zebrafish embryos, Bmp signaling is active in the ALPM but is absent from the myocardial cells at similar stages. These results raise 2 questions: (i) when is Bmp signaling required for cardiac differentiation and (ii) is the observed inhibition of Bmp signaling in differentiating cardiomyocytes relevant for heart formation? As an answer to the first question, we identified a previously unrecognized continuous requirement for Bmp signaling during cardiac differentiation in the ALPM. As an answer to the second question, we found that Smad6 is required to inhibit Bmp signaling and that the down-regulation of Bmp signaling is required for proper formation of the ventricle and preventing expression of atrial myosin in regions outside the atrium.
Results
Bmp Signaling Is Required and Precedes Cardiac Differentiation
We recently showed that Bmp receptor 1a (alk3a;alk3b) double-mutant embryos (further referred to as alk3 mutants) lack differentiated cardiomyocytes.22 Further examination of the alk3 mutant embryos revealed that the precardiac marker hand2 and the cardiac marker nkx2.5 were lost in alk3 mutant embryos (Figure 1A through 1F), while tbx1 was still expressed in the ALPM (Figure 1G and 1H). Although these results demonstrate that Bmp signaling is required for the induction of cardiac mesoderm, they do not address when and where this takes place. To visualize active Bmp signaling, we used an antibody recognizing phosphorylated Smad1, Smad5, and Smad8 protein (referred to as P-Smad), in combination with the transgene Tg(myl7:GFP) labeling differentiated cardiomyocytes. In agreement with our previous findings,7 we observed that in the ALPM the number of cardiomyocytes expressing myl7:GFP progressively increases from 15 to 20 hpf (Figure 2A through 2E). Interestingly, at the 12 somite stage (15 hpf) a nuclear P-Smad protein was detected in cells near the myl7:GFP-expressing cells, while most myl7:GFP-expressing cells were negative for P-Smad (Figure 2A, B and Online Figure I). This was even more apparent at the 20 somite stage (20 hpf) (Figure 2C through 2E and Online Figure I). At this stage a majority (84%, n=6 embryos) of the myl7:GFP-expressing myocardial cells lacked a nuclear P-Smad1 signal, while nuclear P-Smad signal was clearly present in cells near the myl7:GFP-expressing myocardial cells and in the dorsal part of the neural tube (Figure 2D). Together, these results demonstrate that while Bmp signaling is active in the ALPM, there is very little Bmp signaling activity in the differentiating cardiomyocytes. Since Bmp signaling is required for cardiomyocyte differentiation, we addressed whether Bmp signaling was active in cardiac progenitor cells prior to their differentiation into cardiomyocytes. Therefore we analyzed Tg(Bre:eGFP) embryos in which GFP expression is induced by Bmp signaling due to the presence of a conserved Bmp responsive element (BRE) upstream of the coding sequence for eGFP.23 Since GFP has a half-life time of 24 hours, it is possible to trace cells after the Bmp signal has been turned off. When analyzing Tg- (Bre:eGFP) embryos, we observed that differentiating cardiomyocytes, marked by tropomyosin expression, were positive for Bre:GFP, albeit at lower levels in comparison with cells neighboring the myocardial tissue (Figure 2F through 2H and Online Figure I). Together, these results demonstrate that while cardiomyocytes are exposed to Bmp signaling prior to their differentiation and that the signaling activity is required for proper cardiomyocyte differentiation, Bmp signaling is no longer active in differentiating cardiomyocytes.
Figure 1. Alk3a/Bmpr1a is required for cardiac specification.

A, Illustration representing a dorsal view on a zebrafish embryo at the 12-somite stage (15 hours postfertilization) with bilateral cardiac fields colored in blue. B, Schematic diagram of Bmp signaling. C through H, Expression of nkx2.5 (C and D), hand2 (E and F), and tbx1 (G and H) in wild-type siblings or alk3a/bmpr1a mutant embryos. Embryos at 12-somite stage are shown as dorsal views with anterior to the top.
Figure 2. Restricted activation of Bmp signaling in ALPM.

A, C, and F, Illustrations representing transverse sections through the anterior region of a zebrafish embryo at the 12- or 20-somite stage. B, A single transverse confocal image of a Tg(myl7:eGFP) embryo at 15 hours postfertilization (hpf) (12-somite stage) stained for phosphorylated-Smad1,5,8 (P-Smad). Arrow points to an myl7:eGFP-expressing cell, which is negative for P-Smad. Bar indicates region containing several P-Smad positive cells. D, Single confocal scan of Tg(myl7:eGFP) embryos at 19 hpf (20-somite stage) stained for phosphorylated-Smad1,5,8 (P-Smad). Arrow points to the dorsal part of the neural tube. E, Higher magnification of the myocardium shown in D. G, A single confocal scan of a Tg(bre:GFP) embryo stained for tropomyosin (Tpm) to indicate the myocardium. H, Higher magnification of the myocardium from images shown in G. Corresponding gray-scale images of single-color channels are provided in Online Figure I.
Bmp Signaling Is Required for Cardiomyocyte Differentiation Until Midsegmentation Independent of Mesoderm Specification
To address when Bmp signaling is required to induce cardiac differentiation, we studied the zebrafish lost-a-fin (laf) mutant, encoding a Bmp receptor type I (acvr1l/alk8). While alk3 mutant embryos display severe dorsoventral patterning defects resulting in early embryo lethality, zygotic (Z) laf/alk8 mutant embryos have only mild dorsoventral defects due to the presence of maternal alk8 mRNA in the oocyte during gastrulation.16,17,22 Maternal zygotic (MZ) alk8 mutants, lacking all maternal and zygotic alk8 mRNA, display very severe mesodermal patterning defects and as a consequence die between 16 and 24 hpf.17 As reported previously, zygotic (Z) laf/alk8 mutant embryos have a reduced ventral tailfin together with a smaller atrium in comparison with their wild-type siblings at 48 hours postfertilization (hpf) (Online Figure IIA).12,16,17,24 To investigate when zygotic alk8 mRNA is required to allow atrial cardiomyocytes to form correctly, we re-expressed a functional alk8 gene in Zlaf/alk8 mutant embryos at various developmental time points. For this purpose we used a previously described heat-shock inducible transgenic line driving alk8 expression Tg(hsp70:alk8).25 In agreement with an earlier report, inducing alk8 expression during gastrulation (6 hpf) completely rescued the Zlaf/alk8 phenotypes, including the small atrium (Shin et al25 and data not shown). Interestingly, re-expressing alk8 after gastrulation at 16 hpf (14 somites stage) in Zlaf/alk8 mutant embryos still restored the number of atrial cardiomyocytes to wild-type numbers (Figure 3A and 3B) (wild-type control; 85.6±5 atrial cells versus laf/alk8 mutant; 51.8±4 atrial cells versus laf/alk8 mutant/Tg(hsp70:alk8); 82.6±6 atrial cells, P<0.01), although it no longer rescued the loss of the ventral tailfin (Online Figure IIA). These results suggest that even after gastrulation, Bmp signaling is still required for the correct differentiation of cardiomyocytes. Re-expressing alk8 at 16 hpf (14 somites stage) in Zlaf/alk8 mutant embryos did not, however, restore cardiac looping (Online Figure IIA), indicating that cardiac looping and cardiomyocyte differentiation are 2 distinct processes both requiring Bmp signaling but at different time points during embryo development.
Figure 3. Bmp signaling is required during mid- and late-somite stages for myocardial differentiation.

A, Experimental set-up for temporal rescue of myocardial defect in laf/alk8 mutant embryos. B, Quantification of the number of ventricular and atrial cells in laf/ alk8±, laf/alk8−/−, and laf/alk8−/− Tg(hsp70:alk8) embryos heat-shocked at 16 hours postfertilization (hpf). Bars represent mean ± SEM. **P<0.01. C, Experimental set-up for temporal inhibition of Bmp signaling by heat shock–induced Noggin3 expression. D and F, Quantification of the number of ventricular and atrial cardiomyocytes in wild-type sibling or Tg(hsp70:Noggin3) embryo heat-shocked at 10 hpf (D), 16 hpf (E), and 24 hpf (F), respectively. Bars represent mean ± SEM. **P<0.01.
To address when Bmp signaling is still required to stimulate cardiac differentiation after gastrulation, we blocked Bmp signaling at different postgastrula stages using a previously described heat shock inducible Noggin3 transgenic line.24 The Tg(hsp70:Noggin3) carriers were crossed with Tg(myl7:dsRed-nuc) carriers, and the embryos derived from this cross were heat-shocked at various stages and fixed at 48 hpf to quantify the number of cardiomyocytes in the ventricle and atrium (Figure 3C). After an early block in Bmp signaling from 10 hpf (bud stage), both cardiac chambers were observed to contain fewer cardiomyocytes (wild-type siblings; 122.7±4 ventricular and 84.4±6 atrial cells versus Tg(hsp70:Noggin3) embryos; 84.1±11 ventricular and 24.8±6 atrial cells; n=11, P<0.01) (Figure 3D and Online Figure IIB). In contrast, Tg(hsp70:Noggin3)/Tg(myl7: dsRed-nuc) embryos heat-shocked at 16 hpf (14 somite stage) and fixed at 48 hpf contained normal numbers of ventricular cells (111.8±4 in wild-type siblings versus 105.5±6 ventricular cells in Tg(hsp70:Noggin3)). However, the number of atrial cells was significantly reduced in the Tg(hsp70:Noggin3) embryos heat-shocked at 16 hpf (wild-type siblings; 97.6±5 versus Tg(hsp70:Noggin3); 78.8±2 atrial cells, n=11, P<0.01) (Figure 3E and Online Figure IIB). Finally, Tg(hsp70:Noggin3)/Tg(myl7: dsRed-nuc) embryos heat-shocked at 24 hpf and fixed at 48 hpf contained normal numbers of both ventricular and atrial cardiomyocytes (109.1±3 versus 107.8±9 ventricular cardiomyocytes in wild-type siblings versus Tg(hsp70:Noggin3); 82.5±4 versus 73.6±3 atrial cardiomyocytes in wild-type siblings versus Tg(hsp70:Noggin3)) (Figure 3F and Online Figure IIB). However, looping morphogenesis was affected in these embryos (Online Figure IIB).
In conclusion, these data demonstrate that in the embryo proper, there is a continuous requirement for Bmp signaling activity to induce cardiomyocyte differentiation. However, atrial and ventricle cardiomyocytes require a Bmp signal at different stages. While progenitor cells that form the myocardium of the ventricle require activation of Bmp signaling from gastrulation until early segmentation stages (bud stage), those that form the myocardium of the atrium require activation of Bmp signaling from early through late segmentation stages (12–15 somites). This observation is consistent with our previous observation that cardiomyocyte differentiation is initiated in the ventricle and continues in the atrium.7
Bmp Signaling via Alk8 Is Dispensable for Cell Proliferation in the ALPM
Previous studies in chick and mouse embryos showed that inhibiting Bmp signaling stimulates cell proliferation in the ALPM.26–29 To analyze whether cell proliferation was affected in the ALPM of Zlaf/alk8 mutant embryos, we pulse-labeled the embryos from 16 to 19 hpf with BrdU and quantified the number of BrdU positive cells. Expression of nkx2.5 was used to mark the cardiac mesoderm. By counting all BrdU positive cells in ALPM of serial sections (from anterior nkx2.5 positive cells until the notochord appeared in the sections), no difference in the number of BrdU positive cells was observed between laf/alk8 mutant and their wild-type sibling embryos (wild-type sibling 412±6 versus laf/alk8 mutant 420±63) (Figure 4A through 4E). Similar results were obtained with the Tg(hsp70:Noggin3) embryos in which Bmp signaling was inhibited at 16 hpf (data not shown).
Figure 4. Cell proliferation in ALPM is unaffected in laf/alk8 mutant embryos.
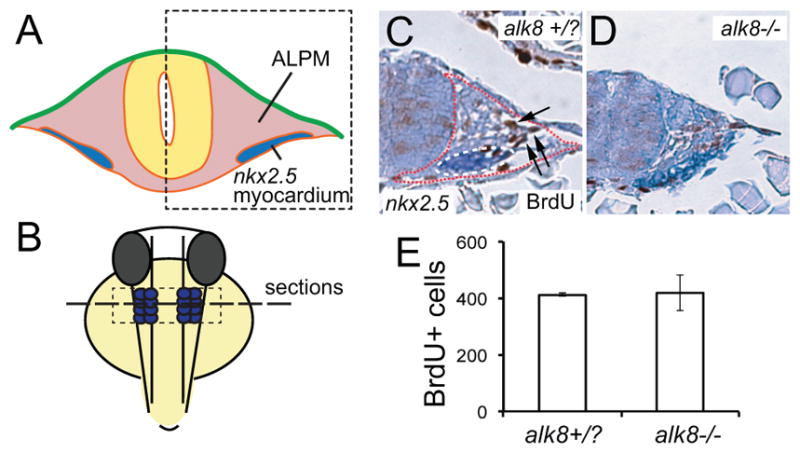
A and B, Illustrations indicating the location and orientation of the transverse sections shown in C and D. C and D, Transverse sections of wild-type sibling (+/− and +/−) (C) and laf/alk8 mutant (−/−) (D) embryo stained for nkx2.5 expression (blue staining) and BrdU (brown), counterstained with hematoxilin. Region of ALPM is indicated by red dotted line, and arrows point to BrdU positive nuclei. E, Quantification of BrdU positive nuclei in serial sections of the ALPM (indicated by presence of nkx2.5 positive cells and absence of notochord) of wild-type siblings and laf/alk8 mutant embryos (n=3). Bars represent ± SEM.
Bmp Signaling Induces tbx2b and tbx20 Expression
Since we observed no effect on cell proliferation in the ALPM upon reducing Bmp signaling, we wondered whether Bmp signaling could affect cardiac differentiation by regulating the expression of cardiogenic factors. The T-box factors tbx20 and tbx2 play independent roles during cardiomyocyte differentiation in mouse embryos.30 In addition, both tbx20 and tbx2 are direct transcriptional targets of Bmp signaling.30,31 To address when BMP-dependent regulation of tbx2b and tbx20 occurs during cardiac differentiation, we reduced Bmp signaling at the mid- to late-somite stage. We observed a reduced expression of both tbx20 and tbx2b in the cardiac region as a consequence of noggin3 expression at the 15-somite stage (Figure 5A through 5D). Quantitative RT-PCR analysis on cardiac explants revealed that tbx20 and tbx2b were significantly down-regulated, 3.5- fold and 5-fold, respectively, as a response to Noggin3 expression during late segmentation stages (Figure 5E and 5F). These results demonstrate that Bmp signaling at midsomite stages is required to induce tbx20 and tbx2b expression, 2 transcription factors that regulate myocardial differentiation.
Figure 5. Overexpression of noggin3 reduces expression of cardiac differentiation markers.

A through D, In situ hybridization for tbx20 and tbx2b at 24 hours postfertilization in wildtype siblings and Tg(hsp70:noggin3) embryos heat-shocked at 15-somite stage. White dotted line in C outlines the linear heart tube. E, Schematic representation of manual dissection of the cardiac region from 25-somite stage embryos subjected to RT-qPCR. F, Quantification of RT-qPCR results showing a down-regulation of the cardiac transcription factors tbx2b and tbx20 in 25-somite hearts of Tg(hsp70:noggin3) embryos heat-shocked at 18-somite stage. **P>0.01. ***P>0.001.
Bmp Signaling in the Cardiac Field Is Inhibited by the Inhibitory Smad6
We observed that cardiomyocytes were exposed to Bmp signaling prior to differentiation and that Bmp signaling was lost in differentiating cardiomyocytes (Figure 1). This raised the question of whether the reduced Bmp signaling observed in the differentiating myocardium is relevant for normal heart formation and, if so, how this is regulated. A possible explanation for the low Bmp signaling activity in cardiomyocytes would be that in cardiomyocytes, Bmp ligands are a limiting factor. To test this possibility we overexpressed bmp2b in the entire embryo including the myl7-expressing cardiac cells using the heat-shock inducible Tg(hsp70:bmp2b) transgene. We observed that cardiomyocytes located medially in the cardiac field, which in wild-type embryos lack P-Smad (Figure 6A through 6D′), were positive for P-Smad in heat-shocked Tg(hsp70:Bmp2b) embryos (Figure 6E through 6H′, indicated by the white arrows). P-Smad levels in these cardiomyocytes, however, were still low when compared with other cell types such as those in the ventral neural tube (Figure 6E through 6H′, indicated by the red arrowhead). This suggested that either medial cardiomyocytes lack the appropriate receptors to respond to the Bmp signal or they may contain an endogenous inhibitory factor. Since expression of alk3a, alk3b, and alk8 was reported to be ubiquitous at the relevant stages,16,17,22 we addressed the involvement of an inhibitor. The inhibitory Smad6 protein can repress Bmp signaling by competing with Smad1 for Smad4 binding or by preventing Smad1,5,8 phosphorylation.19–21 In zebrafish, smad6 is duplicated, and we observed that smad6a was expressed in 2 bilateral populations of cells at the 15-somite stage (Figure 7A and 7B). At later stages when the cardiac fields have fused at the midline, we observed smad6a expression in the cardiac disk (Figure 7C). When the heart tube has formed at 24 hpf, we observed smad6a expression in the linear heart tube predominantly in the future ventricle and in a region posterior of the arterial pole (Figure 7D). To test the function of Smad6a during cardiac differentiation, we generated 2 nonoverlapping antisense morpholino oligos (MOs) complementary to the 5′UTR and translation start site of the smad6a transcript. Injection of these smad6a MOs did not affect overall embryo morphology, but we observed ectopic phospho-Smad1,5,8 signals specifically in the medial differentiating cardiomyocytes (Figure 7 E through 7H′ for ATG MO and data not shown) in comparison with embryos injected with a controlMO(Figure 7A through 7D). In wild-type embryos, only 16% of myl7:GFP expressing cardiomyocytes were P-Smad positive, whereas in smad6a morphants (MO injected), this percentage was increased to 62% (n=6 for control, n=5 for MO injected). Together, these results demonstrate that the inhibitory Smad6a is expressed during myocardial differentiation, during which it is required to prevent Smad phosphorylation and thereby activation of the Bmp signaling pathway.
Figure 6. Bmp signaling is inhibited in the cardiac field.

A through H′, Single confocal scan of Tg(myl7:eGFP) embryos stained for eGFP in green and P-Smad in red at 25-somite stage. A through D, Wild-type embryo with enlargement of the myocardium in A′ through D′. E through H, Representative Tg(hsp70:bmp2b) embryo heat-shocked at 18-somite stage with enlargement of the myocardium shown in E′ through H′. Red arrowhead points to strong P-Smad signal in neural tube. White arrows point to myocardial cells with intermediate level of P-Smad staining. Scale bars represent 50 μm. Scale bars in enlargements represent 10 μm.
Figure 7. The smad6a expression in the cardiac field is required to inhibit Smad1,5,8 phosphorylation.

A through D, In situ hybridization for Smad6a at 15-somite stage in lateral (A) and dorsal (B through D) views, 23-somite stage and 24 hours postfertilization; h, linear heart tube; e, eye. E through L′, Single confocal scan of Tg(myl7:eGFP) embryos stained for eGFP in green and P-Smad in red at 25-somite stage. One half of the ALPM is shown with the midline to the left. The tissue is counterstained with DAPI indicating nuclei. E through H, Embryo injected with control morpholino (MO) with enlargement of myocardium shown in E′ through H′. I through L, Representative Smad6a atg MO injected embryo with enlargement of the myocardium shown in I′ through L′. White arrows point to P-Smad positive myocardial cells. Scale bars represent 50 μm. Scale bars in enlargements represent 10 μm.
Ectopic Bmp Signaling Results in a Hypoplastic Ventricle and Ectopic Expression of Atrial Myosin
To address the biological relevance of Bmp inhibition in the cardiac field during cardiac differentiation, we analyzed the heart tubes of Tg(hsp70:bmp2b) embryos heat-shocked at the 14- or 18-somite stages as well the heart tubes of smad6a morphant embryos at 48 hpf (Figure 8A through 8C). In both the Tg(hsp70:bmp2b) and the smad6a morphant embryos, we observed that cardiac looping was compromised and the size of the ventricle was reduced. To further investigate the effect of ectopic Bmp signaling on cardiac growth, we quantified the number of atrial and ventricle myocytes in those hearts. Surprisingly, we observed that the number of ventricular cardiomyocytes in heat-shocked Tg(hsp70:bmp2b) embryos as well as in smad6a morphant embryos was significantly reduced (wild-type 150.0±9; Tg(hsp70:bmp2b) 122.3±9; smad6a morphants 89.9±4 ventricular cells; P<0.05 and P<0.01) (Figure 8E), while the number of atrial cardiomyocytes was not significantly different from wild-type siblings on ectopic Bmp signaling in Tg(hsp70:bmp2b) embryos or smad6a morphants (wild-type 123.8±6; Tg(hsp70:bmp2b) 111.0±8; smad6a morphants 107.6±6) (Figure 8E). The reduction in the number of ventricle myocytes was most prominent in the smad6a morphant embryos. To address whether Bmp ligands are limiting Bmp signaling in the absence of the inhibitory Smad6, we combined heat shock–induced bmp2b expression with the knock-down of smad6a. However, when we induced ectopic bmp2b expression at the 14-somite stage in smad6a morphants, we did not observe an additional reduction of the number of cells in the ventricle (smad6a morphants; 89.9±4 versus Tg(hsp70:bmp2b)/smad6a morphants; 84.2±7) (Figure 8D and 8E). Interestingly, we did observe the appearance of cells located within the ventricle or arterial pole that expressed the atrial myosin Myh6 (wild-type 0.0±0 cells in n=5 embryos; Tg(hsp70:bmp2b) 1.8±0.5 cells in n=5 of 6 embryos; smad6a morphants 1±0.3 cells in n=5 of 7 embryos; Tg(hsp70:bmp2b)/smad6a MO 8.2±1.8 cells in n=5 of 6 embryos) (Figure 8D through D″). Since we had observed that Bmp signaling was required to induce tbx20 and tbx2b expression in the differentiating myocardium, we next analyzed whether Bmp activity was also sufficient to induce the expression of these cardiac transcription factors. Indeed, we observed ectopic expression of tbx20 in the region posterior to the arterial pole and enhanced expression of tbx2b at places where tbx2b is normally already expressed, including the myocardium. Together, these results demonstrate that inhibition of Bmp signaling by Smad6 is required to allow efficient myocardial differentiation.
Figure 8. Ectopic activation of Bmp signaling reduces cardiomyocyte numbers.

A through D″, Reconstruction of confocal z-stacks of the hearts of a representative uninjected wild-type sibling (A), a Tg(hsp70:bmp2b) heat-shocked at 18 somites (B), a Smad6a MO injected embryo (C), and a Tg(hsp70:bmp2b) embryo injected with Smad6a MO (D through D″). All embryos shown are Tg(myI7:DsRed2-Nuc) and are stained with α-DsRed antibody (red) and an α-S46/Myh6 antibody (green). Scale bars in A represent 50 μm. E, Quantification of the total number of cardiomyocytes in ventricle or atrium of control embryos, Tg(hsp70:bmp2b) heat-shocked at 18 somites, smad6a MO injected embryos, and a Tg(hsp70:bmp2b)/smad6a MO injected embryos at 48 hours postfertilization (hpf), heat-shocked at 18 hpf. Bars represent mean ± SEM *P<0.05. **P<0.01 in comparison with control embryos. F and G, Dorsal views of in situ hybridization for tbx20 and tbx2b expression in Tg(hsp70:bmp2b) embryos heat-shocked at 15 somites and analyzed at 24 hpf. Wild-type controls for panels F and G are presented in Figure 5A (tbx20) and 5C (tbx2b). Arrow in F points to ectopic posterior expression domain. H and I, Lateral views of in situ hybridization for tbx2b expression in wild-type and Tg(hsp70:bmp2b) embryos heat-shocked at 15 somites and analyzed at 36 hpf. Arrows indicate position of the heart; h, heart; e, eye.
Discussion
Our genetic analysis demonstrates that Bmp signaling activity is strictly controlled during cardiomyocyte differentiation, since it is required in the cardiac progenitor cells to induce cardiac differentiation but dispensable and even deleterious once differentiation is initiated. While Bmp signaling is active in cardiac progenitor cells to induce their differentiation, Smad6a actively inhibits Bmp signaling once differentiation is initiated. Inhibition of Bmp signaling in the myocardium is required to allow restricted levels and localization of tbx20 and tbx2b expression and proper differentiation of the cardiac chambers (Online Figure III).
The temporal distinct effects of Bmp signaling on cardiomyocyte number provide insights into the cellular mechanism by which the size of the vertebrate heart is controlled. Our analysis of mutant embryos deficient for Bmpr1a demonstrated that Bmp signaling is required to induce expression of nkx2.5 and hand2 in the ALPM, corroborating previous data from mouse mutants in which Bmpr1a was deleted in the lateral mesoderm by using MesP1-cre.8 During gastrulation a gradient of Bmp signaling along the dorsoventral axis patterns the mesoderm. In both Drosophila and zebrafish, Bmp signaling during gastrulation is required to specify the initial pool of cardiac progenitor cells that express nkx2.5.9,10,12 Our data demonstrate that beyond gastrulation, Bmp signaling is still required to promote cardiac differentiation. This conclusion differs from a previous report suggesting that Bmp signaling after gastrulation is dispensable for cardiac differentiation.12 In the latter study, the authors used different methods to perturb Bmp signaling than were used in our study, which could have been less potent in inhibiting Bmp signaling. Indeed, it was reported that Tg(Hsp70l:dnBmpr1b-eGFP) line and the chemical Bmp inhibitor Dorsomorphin are both less potent than Tg(hsp70:noggin3) line used in this study in recapitulating a full loss of Bmp signaling.24,32,33 We therefore believe that this difference in experimental setup used to perturb Bmp signaling provides a likely explanation for the different outcome of the 2 studies.
Our results are consistent with a model in which initial Bmp signaling activity in the ALPM is required to induce cardiac differentiation by the induction of transcription factors such as nkx2.5, hand2, tbx20, and tbx2, while during myocardial differentiation a more restricted and confined activation of the Bmp pathway is required to allow compartmentalization of the myocardium into distinct chamber and primitive myocardium (Online Figure III). We observed that inhibiting Bmp signaling by ectopic Noggin expression at mid- to late-somite stages resulted in the down-regulation of the cardiac transcription factors tbx20 and tbx2b. Bmp signaling strictly regulates the expression of both tbx20 and tbx2 during myocardial differentiation, and it was suggested that these transcription factors are direct targets of this pathway.30,31 Our results from the Tg(Bre:GFP) embryo experiment are consistent with such a cell-autonomous role for Bmp signaling during myocardial differentiation. Tbx20 and tbx2 regulate the compartmentalization of the myocardium into primitive and chamber myocardium. Mice homozygous mutant for tbx20 establish a small heart tube that fails to undergo looping morphogenesis and to initiate chamber formation.34–37 The transcriptional repressor tbx2 was shown to regulate AVC formation by preventing chamber differentiation.38,39
We and others previously showed that Bmp signaling is required for normal looping morphogenesis.11,22,24,40,41 Here we observed that while cardiomyocyte differentiation was rescued, the looping defect observed in alk8 mutant hearts was not rescued by re-expressing wild-type alk8 at mid-somite stages. In addition we observed abnormal looping morphogenesis in embryos in which Noggin was expressed at 24 hpf, at which time cardiomyocyte differentiation was unaffected. These results suggest that Bmp signaling independent from its role during myocardial differentiation controls looping morphogenesis. Corroborating such a model is the observation that Bmp2/4 is expressed in the looped heart tube, where it is required to confine tbx2 expression to the AVC.42,43
Inhibition of Bmp Signaling by Smad6 Is Required for Myocardial Differentiation
Our results demonstrate that Smad6a is expressed in the cardiac field and linear heart tube to inhibit Bmp signaling. Interestingly, Smad6 is also expressed in the myocardium of the chick heart at stages 10 and 15, where it is regulated by Bmp signaling.28 In addition, in human embryonic stem cells, Smad6 expression is up-regulated during cardiac differentiation (personal communication, S. Braam, R. Passier, and C. Mummery), suggesting that the role for Smad6 during cardiac differentiation has been conserved in higher vertebrates.
Smad6 is not the only negative regulator of Bmp signaling in the myocardium since other mechanisms have been identified. First, a detailed analysis of the mouse nkx2.5 mutant demonstrated that nkx2.5 negatively regulates phosph-Smad1,5,8 levels in the cardiac field during myocardial expansion, possibly by repressing Bmp2 expression.27 Second, it was reported that Bmp signaling in the myocardium is negatively regulated by a direct interaction of tbx20 with phosphorylated Smad1 and Smad5.30 Since Bmp signaling regulates expression of nkx2.5, tbx20, and Smad6 in the myocardium, they may be part of a concerted feedback inhibition loop to control and fine-tune Bmp signaling activity. While Bmp signaling is initially activated broadly in the cardiac mesoderm, it becomes confined to AVC, where it induces the transcriptional repressor tbx2, which is required to maintain a primitive myocardium. Ectopic expression of tbx2 in the entire myocardium of the early mouse heart results in small and unlooped hearts,44 illuminating the necessity to prevent premature activation of tbx2 by Bmp signaling in the heart tube and prospective chambers.
Our results potentially clarify some of the seemingly contradictory results obtained from in vitro experiments. In cardiac differentiation assays using embryonic and induced pluripotent stem cells, both stimulating and inhibitory effects of Bmp growth factors have been reported.45–48 Interestingly, recent results were reported showing that myocardial differentiation is most efficient when embryoid bodies are incubated with a combination of Bmp and activin for 2 days, after which they are cultured in the presence of dorsomorphin and SB431542 to inhibit Bmp and Nodal signaling, respectively.48 Thus, our observations described here contribute to a better understanding of how cardiogenic differentiation is regulated in vivo. A good understanding of the in vivo regulation will allow optimizing this process in vitro in the future.
Online Methods
Zebrafish lines
To identify cardiomyocytes we used the Tg(myl7:EGFP)twu26 1 and Tg(-5.1myl7:nDsRed2)f2 2 lines. Mutants used in this study were lin/bmpr1aahu4087, bmpr1absa0028 and laf/acvr1ltm110b. To re-express alk8 we used Tg(hsp70:alk8) 3, to down regulate bmp signaling we used Tg(hsp70I:Nog3)fr14 4 and to overexpress Bmp2b we used Tg(hsp70:Bmp2b)fr13 4. To detect Bmp signaling activity we used Tg(Bre:GFP)p77 5.
Cell counts
To count cardiomyocytes at various stages we used the Tg(myl7:DsRed2-nuc) line stained using an α-DsRed antibody (Clontech, 1:100) and an α-Myh6 antibody (s46 from the Hybridoma bank 1:10) and a nuclear counter stain with DAPI. Images were acquired by confocal microscopy with a Leica CLSM confocal microscope. 3-D reconstruction and cell counting were performed using Volocity software (Improvision).
Histological methods
Whole mount immunofluorescence was performed according to 6. Vibratome sections were used to visualize pSmad in the lateral plate mesoderm. Embryos were embedded in 3% agarose in PBS and 100 μm sections were stained O/N with primary antibody and for 4 h with secondary antibody.
Agarose slices were subsequently counterstained O/N with DAPI (Invitrogen, 1:2500). Antibodies used in this study: α-DsRed (Clontech, 1:200), α-Myh6 antibody (Hybridoma bank, s46, 1:10), α-eGFP (Chemokine, 1:200) and α-phospho-Smad1, 5, 8 (Cell Signaling, 1:200).
Heat-shock experiments
Heat-shock experiments were performed according to 4. Embryos collected from crosses between heterozygous transgenic carriers and wild-type fish were heat-shocked by transferring the embryos to E3 medium preheated to 37 °C and incubation at 37 °C for 30 min.
BrdU incorporation
BrdU labeling was performed by soaking the embryos in embryo medium containing 5 mg/ml BrdU (Roche) for 3 hours. α-BrdU (Roche) antibody labeling was performed on 10-μm thick paraffin sections which were then stained with 3,3′-Diaminobenzidine (DAB).
Recovery of cardiac tissue and RNA extraction
Whole intact hearts from homozygous Tg(myl7:EGFP)twu34 and Tg(myl7:EGFP)twu34/ Tg(hsp70l:nog3)fr14 embryos were separated from the body proper by fluorescence guided manual dissection. About 300 hearts were retrieved per genotype and stored in RNAlater (Ambion) for further processing. Total RNA was extracted with the RNeasy Micro Kit (Qiagen) according to manufacturer’s instructions. Obtained RNA was assessed for yield and quality with a ND-1000 spectrophotometer (Nanodrop Technologies) and a 2100 Bioanalyzer RNA 6000 Nano chip (Agilent Technologies), respectively. RNA was stored at −80°C
RT-qPCR
For each sample, 100ng of total RNA were reverse transcribed with the Omniscript RT Kit (Qiagen) according to manufacturer’s instructions. Quantitative real-time PCR was carried out on the iQ5 detection system (BioRad) in iQ 96-Well PCR Plates (BioRad). Efficiencies were calculated with REST2009 software (Qiagen). Cqs were determined automatically by the iQ5 software v2.0.148.60623 (BioRad) via the PCR Base Line Subtracted Curve Fit mode. gnb2l1 was chosen as a stable reference gene determined by the geNorm algorithm. Differential gene expression was determined by an efficiency corrected ΔΔCq method. Statistical significance of gene regulation compared to the wild-type control was assessed using Student’s t-test.
Supplementary Material
Novelty and Significance.
What Is Known?
In embryos in which bone morphogenetic protein (BMP) signaling activity is compromised, cardiomyocyte specification is reduced.
Both stimulation and inhibition of BMP signaling activity can enhance cardiomyocyte differentiation of embryonic stem cells.
What New Information Does This Article Contribute?
Cardiac progenitor cells located in the lateral plate mesoderm of the embryo are exposed to and require a BMP signal to initiate cardiomyocyte differentiation.
Once cardiomyocyte differentiation is initiated, BMP signaling is inhibited due to the expression of Smad6 in the cardiomyocytes.
Ectopic BMP signaling activity induces the expression of Tbx2 and Tbx20 and prevents cardiac chamber formation.
After the first report that in Drosophila embryos, Dpp signaling (related to vertebrate BMP and TGF-β) is required to induce cardiac mesoderm, it was recognized that in vertebrates also, BMP signaling induces cardiomyocyte specification and differentiation. However, it is unclear when and where in the vertebrate embryo BMP signaling is active during myocardial differentiation. Here we demonstrate that, cardiomyocyte specification in the lateral plate mesoderm is completely absent in zebrafish mutant embryos that lack BMP signaling. We show that cardiac progenitor cells are exposed to and require BMP signaling to initiate differentiation; however, once differentiation is intiated, BMP signaling is actively repressed by the expression of Smad6, which is required to allow cardiac chamber differentiation. We also show that activating BMP signaling in the myocardium results in the ectopic expression of Tbx2 and Tbx20 and prevents cardiac chamber formation. From these results we conclude that Bmp signaling activity is strictly controlled during cardiomyocyte differentiation. It is required in the cardiac progenitor cells to induce cardiac differentiation, but it is dispensable or even deleterious once differentiation is initiated. Understanding the signals that are required for cardiomyocyte differentiation in the embryo proper will facilitate optimization of in vitro protocols for cardiomyocyte differentiation of multipotent, embryonic stem cells.
Acknowledgments
We gratefully acknowledge Dr. Mullins for providing the Tg(hsp70:alk8) line, and Bilge San and the Hubrecht Imaging Center for technical assistance. We gratefully thank Robert Kelly and members of the Bakkers laboratory for valuable discussions during the preparation of this manuscript.
Sources of Funding
Work in Jeroen Bakkers’ laboratory was supported by EU FP7 FP7-NMP-2007-214539 (BioScent). Salim Abdelilah-Seyfried was supported by a Heisenberg fellowship by the Deutsche Forschungsgemeinschaft.
Non-standard Abbreviations and Acronyms
- ALPM
anterior lateral plate mesoderm
- AVC
atrioventricular canal
- Bmp
bone morphogenetic protein
- BrdU
5-bromo-2′-deoxyuridine
- BRE
Bmp responsive element
- hpf
hours postfertilization
- MO
Morpholino oligo
- RT-PCR
reverse transcriptase, polymerase chain reaction
Footnotes
The online-only Data Supplement is available with this article at http://circres.ahajournals.org/lookup/suppl/doi:10.1161/CIRCRESAHA.111.261172/-/DC1.
Disclosures
None.
References
- 1.Stainier DY, Lee RK, Fishman MC. Cardiovascular development in the zebrafish: I. Myocardial fate map and heart tube formation. Development. 1993;119:31–40. doi: 10.1242/dev.119.1.31. [DOI] [PubMed] [Google Scholar]
- 2.Keegan BR, Meyer D, Yelon D. Organization of cardiac chamber progenitors in the zebrafish blastula. Development. 2004;131:3081–3091. doi: 10.1242/dev.01185. [DOI] [PubMed] [Google Scholar]
- 3.Yelon D, Horne SA, Stainier DY. Restricted expression of cardiac myosin genes reveals regulated aspects of heart tube assembly in zebrafish. Dev Biol. 1999;214:23–37. doi: 10.1006/dbio.1999.9406. [DOI] [PubMed] [Google Scholar]
- 4.Stainier DY. Zebrafish genetics and vertebrate heart formation. Nat Rev Genet. 2001;2:39–48. doi: 10.1038/35047564. [DOI] [PubMed] [Google Scholar]
- 5.Glickman NS, Yelon D. Cardiac development in zebrafish: coordination of form and function. Semin Cell Dev Biol. 2002;13:507–513. doi: 10.1016/s1084952102001040. [DOI] [PubMed] [Google Scholar]
- 6.Smith KA, Chocron S, von der Hardt S, de Pater E, Soufan A, Bussmann J, Schulte-Merker S, Hammerschmidt M, Bakkers J. Rotation and asymmetric development of the zebrafish heart requires directed migration of cardiac progenitor cells. Dev Cell. 2008;14:287–297. doi: 10.1016/j.devcel.2007.11.015. [DOI] [PubMed] [Google Scholar]
- 7.De Pater E, Clijsters L, Marques SR, Lin Y-F, Garavito-Aguilar ZV, Yelon D, Bakkers J. Distinct phases of cardiomyocyte differentiation regulate growth of the zebrafish heart. Development. 2009;136:1633–1641. doi: 10.1242/dev.030924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klaus A, Saga Y, Taketo MM, Tzahor E, Birchmeier W. Distinct roles of wnt/beta-catenin and bmp signaling during early cardiogenesis. Proc Natl Acad Sci U S A. 2007;104:18531–18536. doi: 10.1073/pnas.0703113104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Frasch M. Induction of visceral and cardiac mesoderm by ectodermal dpp in the early drosophila embryo. Nature. 1995;374:464–467. doi: 10.1038/374464a0. [DOI] [PubMed] [Google Scholar]
- 10.Kishimoto Y, Lee KH, Zon L, Hammerschmidt M, Schulte-Merker S. The molecular nature of zebrafish swirl: bmp2 function is essential during early dorsoventral patterning. Development. 1997;124:4457–4466. doi: 10.1242/dev.124.22.4457. [DOI] [PubMed] [Google Scholar]
- 11.Zhang H, Bradley A. Mice deficient for bmp2 are nonviable and have defects in amnion/chorion and cardiac development. Development. 1996;122:2977–2986. doi: 10.1242/dev.122.10.2977. [DOI] [PubMed] [Google Scholar]
- 12.Marques SR, Yelon D. Differential requirement for bmp signaling in atrial and ventricular lineages establishes cardiac chamber proportionality. Dev Biol. 2009;328:472–482. doi: 10.1016/j.ydbio.2009.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schultheiss TM, Burch JB, Lassar AB. A role for bone morphogenetic proteins in the induction of cardiac myogenesis. Genes Dev. 1997;11:451–462. doi: 10.1101/gad.11.4.451. [DOI] [PubMed] [Google Scholar]
- 14.Schlange T, Andree B, Arnold HH, Brand T. Bmp2 is required for early heart development during a distinct time period. Mech Dev. 2000;91:259–270. doi: 10.1016/s0925-4773(99)00311-1. [DOI] [PubMed] [Google Scholar]
- 15.Nakayama T, Cui Y, Christian JL. Regulation of bmp/dpp signaling during embryonic development. Cell Mol Life Sci. 2000;57:943–956. doi: 10.1007/PL00000736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bauer H, Lele Z, Rauch GJ, Geisler R, Hammerschmidt M. The type i serine/threonine kinase receptor alk8/lost-a-fin is required for bmp2b/7 signal transduction during dorsoventral patterning of the zebrafish embryo. Development. 2001;128:849–858. doi: 10.1242/dev.128.6.849. [DOI] [PubMed] [Google Scholar]
- 17.Mintzer KA, Lee MA, Runke G, Trout J, Whitman M, Mullins MC. Lost-a-fin encodes a type i bmp receptor, alk8, acting maternally and zygotically in dorsoventral pattern formation. Development. 2001;128:859–869. doi: 10.1242/dev.128.6.859. [DOI] [PubMed] [Google Scholar]
- 18.Little SC, Mullins MC. Bone morphogenetic protein heterodimers assemble heteromeric type i receptor complexes to pattern the dorsoventral axis. Nat Cell Biol. 2009;11:637–643. doi: 10.1038/ncb1870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hata A, Lagna G, Massague J, Hemmati-Brivanlou A. Smad6 inhibits bmp/smad1 signaling by specifically competing with the smad4 tumor suppressor. Genes Dev. 1998;12:186–197. doi: 10.1101/gad.12.2.186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Goto K, Kamiya Y, Imamura T, Miyazono K, Miyazawa K. Selective inhibitory effects of smad6 on bone morphogenetic protein type i receptors. JBiol Chem. 2007;282:20603–20611. doi: 10.1074/jbc.M702100200. [DOI] [PubMed] [Google Scholar]
- 21.Murakami G, Watabe T, Takaoka K, Miyazono K, Imamura T. Cooperative inhibition of bone morphogenetic protein signaling by smurf1 and inhibitory smads. Mol Biol Cell. 2003;14:2809–2817. doi: 10.1091/mbc.E02-07-0441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Smith KA, Noel E, Thurlings I, Rehmann H, Chocron S, Bakkers J. Bmp and nodal independently regulate lefty1 expression to maintain unilateral nodal activity during left–right axis specification in zebrafish. PLoS Genet. 2011;7:e1002289. doi: 10.1371/journal.pgen.1002289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Alexander C, Zuniga E, Blitz IL, Wada N, Le Pabic P, Javidan Y, Zhang T, Cho KW, Crump JG, Schilling TF. Combinatorial roles for bmps and endothelin 1 in patterning the dorsal–ventral axis of the craniofacial skeleton. Development. 2011;138:5135–5146. doi: 10.1242/dev.067801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chocron S, Verhoeven MC, Rentzsch F, Hammerschmidt M, Bakkers J. Zebrafish bmp4 regulates left–right asymmetry at two distinct developmental time points. Dev Biol. 2007;305:577–588. doi: 10.1016/j.ydbio.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 25.Shin D, Shin CH, Tucker J, Ober EA, Rentzsch F, Poss KD, Hammerschmidt M, Mullins MC, Stainier DY. Bmp and fgf signaling are essential for liver specification in zebrafish. Development. 2007;134:2041–2050. doi: 10.1242/dev.000281. [DOI] [PubMed] [Google Scholar]
- 26.Waldo KL, Kumiski DH, Wallis KT, Stadt HA, Hutson MR, Platt DH, Kirby ML. Conotruncal myocardium arises from a secondary heart field. Development. 2001;128:3179–3188. doi: 10.1242/dev.128.16.3179. [DOI] [PubMed] [Google Scholar]
- 27.Prall OW, Menon MK, Solloway MJ, Watanabe Y, Zaffran S, Bajolle F, Biben C, McBride JJ, Robertson BR, Chaulet H, Stennard FA, Wise N, Schaft D, Wolstein O, Furtado MB, Shiratori H, Chien KR, Hamada H, Black BL, Saga Y, Robertson EJ, Buckingham ME, Harvey RP. An nkx2–5/bmp2/smad1 negative feedback loop controls heart progenitor specification and proliferation. Cell. 2007;128:947–959. doi: 10.1016/j.cell.2007.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tirosh-Finkel L, Zeisel A, Brodt-Ivenshitz M, Shamai A, Yao Z, Seger R, Domany E, Tzahor E. Bmp-mediated inhibition of fgf signaling promotes cardiomyocyte differentiation of anterior heart field progenitors. Development. 2010;137:2989–3000. doi: 10.1242/dev.051649. [DOI] [PubMed] [Google Scholar]
- 29.Hutson MR, Zeng XL, Kim AJ, Antoon E, Harward S, Kirby ML. Arterial pole progenitors interpret opposing fgf/bmp signals to proliferate or differentiate. Development. 2010;137:3001–3011. doi: 10.1242/dev.051565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Singh R, Horsthuis T, Farin HF, Grieskamp T, Norden J, Petry M, Wakker V, Moorman AFM, Christoffels VM, Kispert A. Tbx20 interacts with smads to confine tbx2 expression to the atrioventricular canal. Circ Res. 2009;105:442–452. doi: 10.1161/CIRCRESAHA.109.196063. [DOI] [PubMed] [Google Scholar]
- 31.Mandel EM, Kaltenbrun E, Callis TE, Zeng X-XI, Marques SR, Yelon D, Wang D-Z, Conlon FL. The bmp pathway acts to directly regulate tbx20 in the developing heart. Development. 2010;137:1919–1929. doi: 10.1242/dev.043588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pyati UJ, Webb AE, Kimelman D. Transgenic zebrafish reveal stage-specific roles for bmp signaling in ventral and posterior mesoderm development. Development. 2005;132:2333–2343. doi: 10.1242/dev.01806. [DOI] [PubMed] [Google Scholar]
- 33.Yu PB, Hong CC, Sachidanandan C, Babitt JL, Deng DY, Hoyng SA, Lin HY, Bloch KD, Peterson RT. Dorsomorphin inhibits bmp signals required for embryogenesis and iron metabolism. Nat Chem Biol. 2008;4:33–41. doi: 10.1038/nchembio.2007.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Singh MK, Christoffels VM, Dias JM, Trowe MO, Petry M, Schuster-Gossler K, Burger A, Ericson J, Kispert A. Tbx20 is essential for cardiac chamber differentiation and repression of tbx2. Development. 2005;132:2697–2707. doi: 10.1242/dev.01854. [DOI] [PubMed] [Google Scholar]
- 35.Stennard FA, Costa MW, Lai D, Biben C, Furtado MB, Solloway MJ, McCulley DJ, Leimena C, Preis JI, Dunwoodie SL, Elliott DE, Prall OW, Black BL, Fatkin D, Harvey RP. Murine t-box transcription factor tbx20 acts as a repressor during heart development, and is essential for adult heart integrity, function and adaptation. Development. 2005;132:2451–2462. doi: 10.1242/dev.01799. [DOI] [PubMed] [Google Scholar]
- 36.Takeuchi JK, Mileikovskaia M, Koshiba-Takeuchi K, Heidt AB, Mori AD, Arruda EP, Gertsenstein M, Georges R, Davidson L, Mo R, Hui CC, Henkelman RM, Nemer M, Black BL, Nagy A, Bruneau BG. Tbx20 dose-dependently regulates transcription factor networks required for mouse heart and motoneuron development. Development. 2005;132:2463–2474. doi: 10.1242/dev.01827. [DOI] [PubMed] [Google Scholar]
- 37.Cai CL, Zhou W, Yang L, Bu L, Qyang Y, Zhang X, Li X, Rosenfeld MG, Chen J, Evans S. T-box genes coordinate regional rates of proliferation and regional specification during cardiogenesis. Development. 2005;132:2475–2487. doi: 10.1242/dev.01832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Habets PE, Moorman AF, Clout DE, van Roon MA, Lingbeek M, van Lohuizen M, Campione M, Christoffels VM. Cooperative action of tbx2 and nkx2.5 inhibits anf expression in the atrioventricular canal: implications for cardiac chamber formation. Genes Dev. 2002;16:1234–1246. doi: 10.1101/gad.222902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Harrelson Z, Kelly RG, Goldin SN, Gibson-Brown JJ, Bollag RJ, Silver LM, Papaioannou VE. Tbx2 is essential for patterning the atrioventricular canal and for morphogenesis of the outflow tract during heart development. Development. 2004;131:5041–5052. doi: 10.1242/dev.01378. [DOI] [PubMed] [Google Scholar]
- 40.Chen JN, van Eeden FJ, Warren KS, Chin A, Nusslein-Volhard C, Haffter P, Fishman MC. Left–right pattern of cardiac bmp4 may drive asymmetry of the heart in zebrafish. Development. 1997;124:4373–4382. doi: 10.1242/dev.124.21.4373. [DOI] [PubMed] [Google Scholar]
- 41.Schilling TF, Concordet JP, Ingham PW. Regulation of left–right asymmetries in the zebrafish by shh and bmp4. Dev Biol. 1999;210:277–287. doi: 10.1006/dbio.1999.9214. [DOI] [PubMed] [Google Scholar]
- 42.Ma L, Lu MF, Schwartz RJ, Martin JF. Bmp2 is essential for cardiac cushion epithelial–mesenchymal transition and myocardial patterning. Development. 2005;132:5601–5611. doi: 10.1242/dev.02156. [DOI] [PubMed] [Google Scholar]
- 43.Verhoeven MC, Haase C, Christoffels VM, Weidinger G, Bakkers J. Wnt signaling regulates atrioventricular canal formation upstream of bmp and tbx2. Birth Defects Res A Clin Mol Teratol. 2011;91:435–440. doi: 10.1002/bdra.20804. [DOI] [PubMed] [Google Scholar]
- 44.Christoffels VM, Hoogaars WMH, Tessari A, Clout DEW, Moorman AFM, Campione M. T-box transcription factor tbx2 represses differentiation and formation of the cardiac chambers. Dev Dyn. 2004;229:763–770. doi: 10.1002/dvdy.10487. [DOI] [PubMed] [Google Scholar]
- 45.Yuasa S, Itabashi Y, Koshimizu U, Tanaka T, Sugimura K, Kinoshita M, Hattori F, Fukami S-I, Shimazaki T, Okano H, Ogawa S, Fukuda K. Transient inhibition of bmp signaling by noggin induces cardiomyocyte differentiation of mouse embryonic stem cells. Nature Biotechnology. 2005;23:607–611. doi: 10.1038/nbt1093. [DOI] [PubMed] [Google Scholar]
- 46.Hao J, Daleo MA, Murphy CK, Yu PB, Ho JN, Hu J, Peterson RT, Hatzopoulos AK, Hong CC. Dorsomorphin, a selective small molecule inhibitor of bmp signaling, promotes cardiomyogenesis in embryonic stem cells. PloS One. 2008;3:e2904. doi: 10.1371/journal.pone.0002904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Takei S, Ichikawa H, Johkura K, Mogi A, No H, Yoshie S, Tomotsune D, Sasaki K. Bone morphogenetic protein-4 promotes induction of cardiomyocytes from human embryonic stem cells in serum-based embryoid body development. AJP: Heart and Circulatory Physiology. 2009;296:H1793–H1803. doi: 10.1152/ajpheart.01288.2008. [DOI] [PubMed] [Google Scholar]
- 48.Kattman SJ, Witty AD, Gagliardi M, Dubois NC, Niapour M, Hotta A, Ellis J, Keller G. Stage-specific optimization of activin/nodal and bmp signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell. 2011;8:228–240. doi: 10.1016/j.stem.2010.12.008. [DOI] [PubMed] [Google Scholar]
References
- 1.Huang CJ, Tu CT, Hsiao CD, Hsieh FJ, Tsai HJ. Germ-line transmission of a myocardium-specific gfp transgene reveals critical regulatory elements in the cardiac myosin light chain 2 promoter of zebrafish. Dev Dyn. 2003;228:30–40. doi: 10.1002/dvdy.10356. [DOI] [PubMed] [Google Scholar]
- 2.Mably JD, Mohideen MA, Burns CG, Chen JN, Fishman MC. Heart of glass regulates the concentric growth of the heart in zebrafish. Curr Biol. 2003;13:2138–2147. doi: 10.1016/j.cub.2003.11.055. [DOI] [PubMed] [Google Scholar]
- 3.Shin D, Shin CH, Tucker J, Ober EA, Rentzsch F, Poss KD, Hammerschmidt M, Mullins MC, Stainier DY. Bmp and fgf signaling are essential for liver specification in zebrafish. Development. 2007;134:2041–2050. doi: 10.1242/dev.000281. [DOI] [PubMed] [Google Scholar]
- 4.Chocron S, Verhoeven MC, Rentzsch F, Hammerschmidt M, Bakkers J. Zebrafish bmp4 regulates left-right asymmetry at two distinct developmental time points. Dev Biol. 2007;305:577–588. doi: 10.1016/j.ydbio.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 5.Alexander C, Zuniga E, Blitz IL, Wada N, Le Pabic P, Javidan Y, Zhang T, Cho KW, Crump JG, Schilling TF. Combinatorial roles for bmps and endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton. Development. 2011;138:5135–5146. doi: 10.1242/dev.067801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Smith KA, Chocron S, von der Hardt S, de Pater E, Soufan A, Bussmann J, Schulte-Merker S, Hammerschmidt M, Bakkers J. Rotation and asymmetric development of the zebrafish heart requires directed migration of cardiac progenitor cells. Dev Cell. 2008;14:287–297. doi: 10.1016/j.devcel.2007.11.015. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.