

Abstract
A recent shift in legal and social attitudes toward marijuana use has also spawned a surge of interest in understanding the effects of marijuana use on the brain. There is considerable evidence that an adolescent onset of marijuana use negatively impacts white matter coherence. On the other hand, a recent well-controlled study demonstrated no effects of marijuana use on the morphometry of subcortical or cortical structures when users and non-users were matched for alcohol use. Regardless, most studies have involved small, carefully selected samples, so the ability to generalize to larger populations is limited. In an attempt to address this issue, we examined the effects of marijuana use on white matter integrity and cortical and subcortical morphometry using data from the Human Connectome Project (HCP) consortium. The HCP data consists of ultra-high resolution neuroimaging data from a large community sample, including 466 adults reporting recreational marijuana use. Rather than just contrasting two groups of individuals who vary significantly in marijuana usage as typifies prior studies, we leveraged the large sample size provided by the HCP data to examine parametric effects of recreational marijuana use. Our results indicate that the earlier the age of onset of marijuana use, the lower was white matter coherence. Age of onset also also affected the shape of the accumbens, while the number of lifetime uses impacted the shape of the amygdala and hippocampus. Marijuana use had no effect on cortical volumes. These findings suggest subtle but significant effects of recreational marijuana use on brain structure.
Keywords: Marijuana, Morphometry, Connectivity, Human Connectome Project, MRI, DTI
Highlights
-
•
Effects of marijuana use on brain morphology in 466 recreational users were investigated.
-
•
Age of first use impacted white matter integrity.
-
•
Age of first use and lifetime times used impacted shape of the hippocampus and accumbens
-
•
Marijuana use did not affect cortical volume.
1. Introduction
Previous epidemiological studies have revealed strong negative impacts of marijuana use, suggesting that marijuana has similar potential for abuse as other illicit substances (e.g., cocaine), is associated with respiratory illnesses, and leads to cognitive impairment (for a review see ref. Volkow et al., 2014). However, several focused empirical studies have countered these results, finding instead no significant effect of marijuana use on subcortical brain morphometry and only an uncertain effect on cognition (e.g., Block et al., 2000, Pope et al., 2003, Weiland et al., 2015). The past two decades have seen shifts in legal and societal attitudes toward marijuana use, with 23 states and the District of Columbia legalizing medical marijuana and four states legalizing recreational marijuana (Marijuana Resource Center: State Laws Related to Marijuana, 2016); moreover, perceptions of the risk of regular marijuana use have decreased, even amongst adolescents, particularly in Colorado, recreational marijuana is now legal (Schuermeyer et al., 2014). As increases in the potency of marijuana have accompanied these shifts in attitudes (Volkow et al., 2014), it is becoming increasingly important to understand the precise neural effects of long-term marijuana use and the impact of the age of first use.
Adolescence is a sensitive period for brain development with white matter myelination and gray matter pruning, and, critically, an increase in the number of cannabinoid receptors that respond to marijuana (Jacobus and Tapert, 2014). While preliminary studies of the effects of marijuana use on white matter integrity showed no significant effects in adolescents or adults (DeLisi et al., 2006, Gruber and Yurgelun-Todd, 2005), a growing body of research suggests that an adolescent onset of heavy marijuana use can have neurotoxic effects on developing white matter, reflected in decreased white matter coherence as assessed by measures of diffusivity, e.g., fractional anisotropy (FA) and radial diffusivity (RD) (Arnone et al., 2008, Bava et al., 2013, Filbey et al., 2014, Jacobus et al., 2009, Jacobus et al., 2013). Importantly, these effects have been observed longitudinally, suggesting a causation between marijuana use and white matter changes (Bava et al., 2013, Becker et al., 2015, Jacobus et al., 2013). However, most of these studies have relied on small sample sizes (i.e., between 10 and 50 marijuana users, with most below 20), so their ability to generalize to a broader population is limited. Moreover, the majority of these studies all examined the effects of heavy use (e.g., daily use), and much less is known about the effects of casual marijuana use on white matter integrity. As many white matter tracts continue to develop in adolescence and young adulthood (Lebel and Beaulieu, 2011), with maximal change in such development during this time frame (Simmonds et al., 2014), it is important to understand how the age of onset of marijuana use impacts neurodevelopment not only in heavy users but more casual users, especially considering that adolescence is often a time of experimentation with substances of abuse (Schuermeyer et al., 2014).
Studies of the effects of marijuana use on cortical and subcortical morphometrics in humans have typically focused on the amygdala and hippocampus (Rocchetti et al., 2013) and, to a lesser extent, the nucleus accumbens (e.g., Gilman et al., 2014) and orbitofrontal cortex(Churchwell et al., 2010, Filbey et al., 2014, Pagliaccio et al., 2015). These structures are known to have important roles in reward processing and their function/structure is known to be disrupted by drugs of abuse (Berridge and Robinson, 2003). At least some, but far from all, of the evidence suggests an influence of marijuana on brain structure. For example, marijuana users compared to nonusers have been found to have reduced amygdala volume (Churchwell et al., 2010, Schacht et al., 2012), and amygdala volume reductions have been correlated with increased levels of self-reported craving and relapse in consumption after 6-months from detoxification from alcohol dependence (Wrase et al., 2008). On the other hand, a recent meta-analysis of 14 studies of marijuana users compared to nonusers found no summary changes in amygdala volume, but did observe a consistent pattern of reduced hippocampal volume (Rocchetti et al., 2013). In addition, a large number of studies with animals and humans have shown that marijuana affects the structure of the nucleus accumbens (Gilman et al., 2014, Kolb et al., 2006). Hence, there is evidence in the existing literature to suggest the possibility that marijuana influences the structure of these regions, all of which are known to be affected in addiction (Koob and Volkow, 2010).
Nonetheless, a recent well-controlled study by Weiland et al. (2015) found no evidence of an effect of marijuana on the morphometry of these structures. They compared morphometry in a sample of adult and adolescent daily users of marijuana to nonusers (matching the groups for alcohol use), while controlling for other confounding variables of tobacco use, depression, impulsivity, age, and gender. Importantly, they found no group differences in measures of brain morphometry for the nucleus accumbens, amygdala, hippocampus, cerebellum, or 35 cortical regions in each hemisphere. Interestingly, when they simply controlled for the amount of alcohol use, rather than matching users and nonusers, they replicated several findings of Gilman and colleagues. Furthermore, when examining effect size across previous studies, they found that the literature demonstrates a mean lack of effect.
Given the discrepancies in the literature, we wanted to re-examine this issue using a large representative sample. To this end, we analyzed extremely high-quality multi-modal neuroimaging data from 466 participants in the Human Connectome Project (HCP) who reported using marijuana at least once during their lives (Van Essen et al., 2012). The participants in this sample consist of twins and their non-twin siblings who have no history of major psychiatric illness, but vary greatly in terms of race, education, income, BMI, and the degree of recreational drug use. A recent study used this HCP dataset to disentangle causal effects of marijuana use on regional brain volume from shared genetic effects and found that it was mainly shared genetic effects explained differences in bran volumes (Pagliaccio et al., 2015). However, this study did not investigate the effects of marijuana use on white matter integrity or the shape of subcortical regions, which was the focus of the current study. Rather than investigating extremes of marijuana use (i.e., heavy users vs. nonusers) like most previous studies, we leveraged the large sample size to take a parametric approach, examining marijuana use along a spectrum, so as to search more specifically for dose-dependent effects. Nevertheless, a comparison of users and non-users was also performed as a replication of prior work.
2. Materials and methods
2.1. HCP participants
Data analyzed in the current study came from the most recent S900 Release (http://humanconnectome.org/documentation/S900/index.html) from the WU-Minn HCP Consortium (Van Essen et al., 2012). Data were only considered if they had structural (e.g., at least 1 T1w and T2w scan) and diffusion imaging scans, and had complete SSAGA and family information (see below), resulting in 857 possible participants. We further restricted analyses to individuals who had reported using marijuana at least once in their lifetime, resulting in 466 participants in the final sample. An overview of the participant recruitment strategy is described in detail elsewhere (Van Essen et al., 2012). In brief, the HCP aims to “recruit a sample of relatively healthy individuals free of a prior history of significant psychiatric or neurological illnesses. Our goal is to capture a broad range of variability in healthy individuals with respect to behavioral, ethnic, and socioeconomic diversity (p. 2224).” The sample is meant to be representative of the population at large and includes individuals who smoke, are overweight, have subclinical psychiatric symptoms, and—critical for the current study—use recreational drugs. HCP participants are human adult twins (MZ and DZ) and their non-twin siblings, aged 22–35 years.
The data included in this study consisted of individuals from 270 different families, ranging from 1 to 4 members per family, with a mean number of 1.7 members per family. Sibships with individuals having severe neurodevelopmental disorders, documented neuropsychiatric disorders, diabetes, or high blood pressure were excluded, as were twins born before 34 weeks gestation and non-twins born before 37 weeks (Van Essen et al., 2012). Demographic, medical, family history, personality, cognitive, and lifestyle information is collected from each subject over two weeks of phone and in-person interviews as well as through written assessments (e.g. the Semi-Structured Assessment for the Genetics of Alcoholism, SSAGA).
2.2. Marijuana use
Marijuana use was quantified with self-report measures assessed by the Semi-Structured Assessment for the Genetics of Alcoholism (SSAGA). Number of times used was quantified in the SSAGA as 0 (never used), 1 (1–5 uses), 2 (6–10 uses), 3 (11–100 uses), 4 (101–999 uses), or 5 (> 1000 uses). Age of first use in the SSAGA was quantified as follows: 1 (first use at less than 15 years of age), 2 (15 to 17 years of age), 3 (18 to 20 years of age), 4 (≥ 21 years of age), or 5 (never used). Age of first use was reverse scored so that an earlier age of first use was scored more highly, in line with the times used measure.
2.3. Covariates
Age, gender, tobacco and alcohol usage, and years of education were included as covariates in all analyses. Many studies of substance use quantify tobacco use with a “packs per day” measure. As no equivalent measure is available in the SSAGA, we quantified tobacco use using a composite measure calculated from the average of the Z-scores for the following SSAGA measures: “Total times used/smoked ANY TOBACCO in past 7 days”, “Cigarettes per day when smoking regularly”, “Years since respondent smoked last cigarette”, “Years smoked.” In this manner, the cumulative effect of recent and/or past tobacco use could be controlled for. In a similar manner, we quantified alcohol use as a composite measure reflecting frequency of recent and past drinking, calculated from the average of the Z-scores for the following SSAGA measures: “Total drinks in past 7 days”, “Drinks per drinking day in past 12 months”, “Frequency of any alcohol use in past 12 months”, “Drinks per day in heaviest 12-month period”, and “Frequency of any alcohol use, heaviest 12-month period”. Where appropriate, scores were reverse scored such that higher values represented more severe/longer use. A description of these use measures is available here: https://wiki.humanconnectome.org/display/PublicData/HCP+Data+Dictionary+Public-+500+Subject+Release. Years of education was included as a covariate in order to serve as a rough approximation of academic achievement/intelligence. For the DTI data analyses, an index of motion was included as a covariate, as previous work has shown motion to contribute to spurious effects in group analyses (Yendiki et al., 2014). Motion was calculated following Yendiki et al. (2014), with a Total Motion Index being calculated from the average volume-by-volume translation and the average volume-by-volume rotation. The Total Motion Index was defined for each subject i as:
where j = 1, … 2 indexes the 2 motion measures, xij is the value of the j-th motion measure for the i-th subject, and Mj, Qj, and qj are, respectively, the median, upper quartile, and lower quartile of the j-th motion measure over all participants included in a given analysis. Covariates were converted to Z-scores for all of the marijuana use analyses.
While the focus of the current study was the identification of linear effects of marijuana use on brain morphometry, we conducted a group comparison of users and non-users as a confirmatory analysis of prior group studies that have investigated morphometry (for a review, see Jacobus and Tapert, 2014). Users were defined as those who both started using marijuana before the age of 18 and have used marijuana more than 100 times in their lifetime. Non-users were those who reported zero lifetime uses of marijuana. The final sample for this analysis included 247 users and 392 non-users. Age, gender, education, tobacco and alcohol use, and TMI (for DTI analysis) were used as covariates.
2.4. HCP image acquisition and pre-processing
MRI scans were collected using a HCP-customized Siemens 3T Connectome Skyra magnet, as described in detail elsewhere (Uǧurbil et al., 2013, Van Essen et al., 2012). Structural MRI scans were acquired at 0.7 mm isotropic resolution (FOV = 224 mm, matrix = 320, 256 sagittal slices in a single slab) and include a pair of T1-weighted (T1w) and a pair of T2-weighted (T2w) images. Diffusion images were acquired at very-high spatial resolution (1.25 mm isotropic) with a high-angular resolution diffusion imaging (HARDI) approach, incorporating 3 shells of b = 1000, 2000, and 3000 s/mm2 with 270 q-points distributed over the 3 shells. Not all participants had complete diffusion images, with the total number of q-points in the b = 1000 shell (the shell for which diffusion tensor parameters were calculated) ranging from 30 to 90 with a mean of 87.3.
Data downloaded from the HCP for the present study had undergone a minimal preprocessing pipeline, described in detail elsewhere (Glasser et al., 2013). Structural data analyzed in the current study were the result of the PreFreeSurfer and FreeSurfer pipelines. Briefly, T1w and T2w images were corrected for gradient distortion, aligned and averaged (i.e., the pair of T1w images were aligned and averaged together), brain extracted, and corrected for readout distortion. The undistorted T1w and T2w images were then registered together in order to perform bias field correction, and finally, were non-linearly aligned to MNI space. Diffusion data analyzed in the current study were the result of the Diffusion Preprocessing pipeline. Diffusion images underwent b0 intensity normalization, EPI distortion correction with FSL's topup, eddy current and motion correction, and gradient nonlinearity correction.
2.5. Current data analysis
2.5.1. Accounting for family structure
Although the family structure of the HCP and similar studies present unique opportunities for investigating the heritability of brain structures (Pagliaccio et al., 2015, Rentería et al., 2014), the shared variance between family members violates assumptions of independence of observations within the sample. Extensions to permutation methods have been developed that allow for types of designs that contain well-structured non-independence between observations like paired tests and repeated measures (Winkler et al., 2014). More recently, it has been demonstrated that multi-level exchangeability blocks can be defined to allow for permutation tests on voxel-wise brain data with complex sibships such as those contained in the HCP (Winkler et al., 2015). The exchangeability block file used in the current study consisted of separate columns that coded the family type (e.g., two monozygotic twins and a non-twin sibling or a singleton with no relatives enrolled), family ID (i.e., each of the 273 families had a different ID), or sibling type (i.e., participant is a monozygotic twin, a dizygotic twin, a non-twin sibling, or a singleton). Rather than allowing for permutations amongst all individuals, permutations were constrained at both the whole-block level (e.g., swapping labels between members of like family types, such as families with two monozygotic twins and a non-twin sibling) or the within-block level (e.g., swapping labels between two monozygotic twins). In this manner, heritability—or more specifically the non-independence due to heritability—is treated as a nuisance variable, but without directly modeling the heritability.
All voxel-wise analyses involving related individuals were carried out using multi-level block permutation methods in FSL's PALM tool version 94a (Winkler et al., 2015), an extension of permutation methods for the General Linear Model (Winkler et al., 2014). Nested exchangeability blocks were defined which restricted permutations to the same family type, which allowed us to account for family structure without directly modeling these complicated repeated-measures factors. Voxel-wise statistics were corrected at the cluster-level using Threshold-Free Cluster Enhancement (TFCE) (Smith and Nichols, 2009, Winkler et al., 2014) to a Family-Wise Error (FWE) rate of p < 0.05, with 5000 permutations.
Nevertheless, as a further control for heritability, we conducted follow-up analyses by randomly selecting one user from each family such that there were no siblings within the subsample, resulting in 272 marijuana users. Identical analyses were performed using the full user sample and the unrelated subsample. These analyses were conducted using FSL's randomise package, which performs permutation methods, without using exchangeability blocks. Following the PALM analyses, 5000 permutations were performed. Likewise, voxelwise results were corrected using Threshold-Free Cluster Enhancement to a FWE-corrected p < 0.05.
2.5.2. White matter integrity
We further processed the pre-processed diffusion images using FSL's FDT toolbox and Tract-Based Spatial Statistics (TBSS; Smith et al., 2006). Briefly, the diffusion tensor model (Pierpaoli and Basser, 1996) was fit at each voxel yielding maps for fractional anisotropy (FA; fraction of anisotropic diffusion), axial diffusivity (AD; diffusivity along the principal axis), radial diffusivity (RD; average of diffusion along the 2 secondary axes), and mean diffusivity (MD; mean of all diffusivity axes). These maps were then nonlinearly aligned to the FMRIB 1 mm FA template (in the same space as the MNI152 standard brain) and skeletonized. This last step creates a skeleton which represents the centers of all fiber bundles that are generally common to all participants in the study. The skeleton was thresholded at FA < 0.3. The skeleton and nonlinear registration parameters were generated using the FA maps, and these transformations were subsequently applied to the other diffusion parameter maps. Voxel-wise statistics were then performed in this skeleton space.
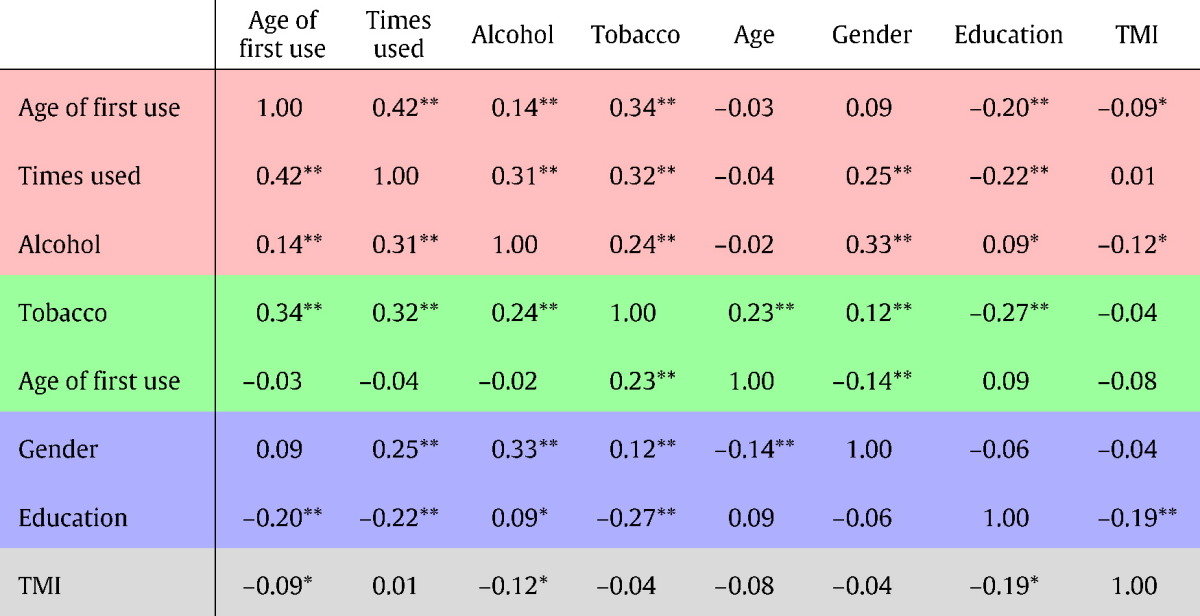
The effect of marijuana use on white matter was assessed with a linear parametric analysis across level of marijuana use. Participants reported a wide-range of levels of marijuana use in terms of age of first use as well as the number of times used over the lifetime, as shown in Fig. 1. Age of first use and times used were included as factors of interest and, as described above, all analyses included alcohol and tobacco use, age, gender, years of education, and Total Motion Index as covariates. For a table of correlations between all covariates see Table 1.
Fig. 1.

Histograms depicting self-report measures of marijuana use including age of first use (A) and times used in lifetime (B).
Table 1.
Correlation matrix for imaging covariates. Those rows shaded in red represent marijuana use measures, those shaded in green represent other substance use covariates, those shaded in blue represent demographic covariates, and the gray row represents the diffusion Total Motion Index (TMI) covariate that was only used for white matter analyses. * denotes a correlation with a puncorr < 0.05; ** denotes a correlation with a puncorr < 0.01.
2.5.3. Volumetric analyses
Analyses of voxelwise gray matter morphometry were carried out with FSL-VBM (Douaud et al., 2007) an optimized VBM protocol (Good et al., 2001) carried out with FSL tools (Smith et al., 2004). First, structural images were brain-extracted and gray matter-segmented before being registered to the MNI 152 standard space using non-linear registration (Andersson et al., 2007). The resulting images were averaged and flipped along the x-axis to create a left-right symmetric, study-specific gray matter template. Second, all native gray matter images were non-linearly registered to this study-specific template and “modulated” to correct for local expansion (or contraction) due to the non-linear component of the spatial transformation. The modulated gray matter images were then smoothed with an isotropic Gaussian kernel with a sigma of 3 mm. Finally, voxelwise GLM was applied using permutation-based non-parametric testing, correcting for multiple comparisons across space, using Threshold-Free Cluster Enhancement (TBSS; Smith and Nichols, 2009).
Following Weiland et al. (2015), we also performed a multivariate analysis on the effects of marijuana use on subcortical and cortical volumes and cortical thickness extracted with FreeSurfer. These data were downloaded in table format from https://db.humanconnectome.org/ and were the result of the HCP FreeSurfer preprocessing pipeline (Glasser et al., 2013). Rather than analyzing whether marijuana showed a multivariate effect across all 35 cortical regions contained in this table as did Weiland et al. (2015), we chose an a priori approach, focusing on prefrontal regions and subcortical regions where marijuana has been shown to have significant effects (e.g., Bellis et al., 2005, Churchwell et al., 2010). Regions of interest included 15 prefrontal cortical regions (in each the left and right hemispheres): medial and lateral orbitofrontal cortex, caudal anterior cingulate, caudal middle frontal, inferior frontal gyrus (3 subregions: pars orbitalis, pars opercularis, pars triangularis), rostral middle frontal, superior frontal, and frontal pole. Subcortical regions included nucleus accumbens, hippocampus, cerebellum cortex and white matter, thalamus, and amygdala. White matter volumes were included for the anterior and mid anterior corpus callosum.
2.5.4. Subcortical shape
Shape analyses of subcortical ROIs (hippocampus, amygdala, accumbens) were carried out using FSL's FIRST tool (Patenaude et al., 2011). FIRST allows for a model-based segmentation and registration of anatomical images, where volumetric labels are parameterized as surface meshes. Models for each subcortical structure are based on a training set of manually traced images. Vertex locations from each participant were projected onto the surface of the average shape (transformed to MNI space), resulting in scalar projection values. This approach normalizes for brain size. The same models of marijuana use generated for the white matter analyses were used here. Again, PALM was used to perform multi-level block permutation analyses. In order to identify which, if any, subregion of the hippocampus, amygdala, or nucleus accumbens exhibited a shape difference, we referred to the Jülich histological atlas (Amunts et al., 2005) contained in FSL. Because FIRST only reports changes at the surface of the structure and does not contain information about which (if any) deep levels of a structure are impacted, these subregion labels represent approximations.
3. Results
3.1. Demographics and substance measures
Descriptive statistics for age, gender, race, and education level are reported in Table 2. Histograms of the distribution of marijuana use measures are shown in Fig. 1. Summarizing these data, non-twins were more likely to have used marijuana than twins and men were more likely to have used marijuana than women, but marijuana use did not differ by race, ethnicity, or education level. Those reporting marijuana use (compared to those with no lifetime history of marijuana use) showed higher levels of tobacco use (t(858) = 9.5, p < 0.001), alcohol use (χ2(5) = 183.2, p < 0.001), and illicit drug use (χ2(5) = 151.4, p < 0.001).
Table 2.
Table of demographic information from the full sample on 857 participants with imaging data in the HCP S900 data release. Significant χ2 tests are indicated by an asterisk, reflecting p < 0.05.
| Non-users | Users | χ2 statistic | |
|---|---|---|---|
| Gender | 6.0* | ||
| Female | 27.6% | 28.4% | |
| Male | 18.3% | 25.7% | |
| Ethnicity | 3 | ||
| Hispanic/Latino | 3.1% | 5.2% | |
| Not Hispanic/Latino | 42.1% | 48.6% | |
| Unknown or not reported | 0.6% | 0.3% | |
| Race | 6.6 | ||
| Am. Indian/Alaskan Nat. | 0.1% | 0.1% | |
| Asian/Nat. Hawaiian/other Pacific Is. | 2.3% | 2.9% | |
| Black or African Am. | 6.7% | 9.7% | |
| More than one | 0.8% | 1.5% | |
| Unknown or not reported | 0.3% | 1.3% | |
| White | 35.5% | 38.7% | |
| Education | 10 | ||
| < 12 | 1.0% | 2.6% | |
| 12 | 6.0% | 8.4% | |
| 13 | 2.1% | 3.8% | |
| 14 | 5.6% | 7.1% | |
| 15 | 3.1% | 2.7% | |
| 16 | 20.9% | 21.4% | |
| > 16 | 7.0% | 8.3% | |
| Twin status | 4.2* | ||
| Not twin | 21.3% | 29.0% | |
| Twin | 24.5% | 25.2% |
3.2. White-matter Tract-Based Spatial Statistics
We then examined the effects of marijuana use on white matter diffusion parameters (FA, AD, RD, and, MD). The group comparison showed no significant effects, possibly suggesting that the frequency of marijuana use in the HCP sample is not severe enough to replicate previous studies, which largely focused on comparisons of non-users and daily marijuana users. In line with this possibility, there were no linear effects of the number of times used on white matter coherence in users.
As shown in Fig. 2, age of first use had a positive association with FA as well as a negative association with RD, such that an earlier age of first use was associated with lower FA and greater RD in a large cluster of right hemisphere white matter. These tracts primarily subsisted of the Superior Longitudinal Fasciculus (SLF), Inferior Longitudinal Fasciculus (ILF), and Forceps Major and Minor. The SLF connects the prefrontal cortex and parietal cortex and is involved in executive functions (Depue et al., 2016, Smolker et al., 2014), and the ILF connects the temporal and occipital cortices, has been shown to affected by adolescent marijuana abuse (Bava et al., 2013, Jacobus et al., 2009). The Forceps Major and Minor are extensions of the corpus callosum connecting the left and right occipital and frontal lobes, respectively. Thus, even though most of the effects on FA and RD were found in the right hemisphere, communication between the left and right hemispheres may be impacted by marijuana age of onset.
Fig. 2.

Effects of age of first use of marijuana on Tract-Based Spatial Statistics from the full sample of 466 marijuana users. Hot colors depict a positive relationship between age of first use and FA, i.e., earlier age of onset associated with decreased FA, and cold colors depict a negative relationship between age of first use and RD, i.e., earlier age of onset associated with increased RD. Earlier marijuana use was associated with decreased white matter coherence in the right Superior Longitudinal Fasciculus, lateral prefrontal white matter, anterior and posterior corpus callosum (extending to the Forceps Minor and Major), as well as the right Inferior Longitudinal Fasciculus. White numbers reflect MNI coordinates in the Z (axial slices), and Y (coronal slices) planes.
When examining the subset of unrelated marijuana users, we confirmed the negative effect of an earlier age of first use on FA and RD in the SLF, as shown in Fig. 3. Increased RD has been related to demyelination in neurodegenerative diseases such as multiple sclerosis (Klawiter et al., 2011), suggestive of a neurotoxic effect with early age of first use of marijuana. These findings taken together, and in line with previous studies (Becker et al., 2015, Jacobus and Tapert, 2014), suggest that an earlier age of onset is associated with decreased coherence of white matter. Moreover, these results suggest that marijuana use has an impact on the development of these white matter tracts during adolescence.
Fig. 3.

Effects of age of first use of marijuana on Tract-Based Spatial Statistics from the restricted sample of 277 unrelated marijuana users. Hot colors depict a positive relationship between age of first use and FA, i.e., earlier age of onset associated with decreased FA, and cold colors depict a negative relationship between age of first use and RD, i.e., earlier age of onset associated with increased RD. Earlier marijuana use was associated with decreased white matter coherence in the right Superior Longitudinal Fasciculus, lateral prefrontal white matter, and the anterior corpus callosum extending to the Forceps Minor. White numbers reflect MNI coordinates in the Z (axial slices), and Y (coronal slices) planes.
3.3. Volumetric analyses
The VBM analysis in the full sample and the subsample of unrelated users showed no significant results, nor in the group comparison. Likewise, the multivariate analysis of a priori cortical and subcortical ROI volumes revealed no significant effects of marijuana use with respect to either age of first use (F(20,522) = 0.50, p = 0.97, ηp2 = 0.02) or times used over the lifetime (F(40,1052) = 0.88, p = 0.69, ηp2 = 0.03). These findings are in line with those presented by Weiland et al. (2015), demonstrating no significant effects of marijuana use on cortical brain morphometry, coupled with minimal effect sizes.
3.4. Shape analyses of subcortical structures
We then investigated whether the shape of the hippocampus, amygdala, or accumbens was impacted by marijuana use. For the full sample of users, the results are shown in Fig. 4. There was a trend (pFWE < 0.08) for a significant negative relationship between the number of times used and scalar values in the left amygdala (Fig. 4B). Participants with a higher number of times used showed more inward deflection in the superficial group of the left amygdala. In the right nucleus accumbens, there was a significant positive relationship between age of first use and shape, such that participants with an earlier age of onset of use showed greater outward deflection (Fig. 4B). This finding suggests that a later age of first use is related to more outward deflection (but not overall volume) of the right anterior nucleus accumbens. In the left hippocampus cornu ammonis and posterior hippocampus subiculum, a greater number of times used was associated with greater inward deflection (Fig. 4C).
Fig. 4.

Effects of marijuana use on subcortical shape in the full sample of users. (A) Trend for significant negative relationship between number of times used and scalar values in the left amygdala. Participants with a higher number of times used showed more inward deflection in the superficial group of the left amygdala. FWE-corrected p < 0.08. Saggital slice (left), Coronal slice (middle), Axial slice (right) (B) Significant positive relationship between age of first use and scalar values in the right nucleus accumbens. Participants with a earlier age of first use showed more inward deflection in the right posterior ventral accumbens. FWE-corrected p < 0.05. Saggital slice (left), Coronal slice (middle), Axial slice (right) (C) Significant negative association between times used marijuana and scalar values in the left hippocampus. Participants with greater number of lifetime uses showed more inward deflection in the anterior hippocampus cornu ammonius and the posterior hippocampus subiculum. FWE-corrected p < 0.05.
The effects of the number of times used marijuana in the restricted sample closely mirrored those in the full sample. In the left superficial amygdala there was a trend for participants with a higher number of times used to show more inward deflection (Fig 5B). Likewise, the same pattern was observed in the left hippocampus cornu ammonis (Fig. 5C). For age of first use, however, there was the opposite pattern (significant to pFWE < 0.05), albeit in the left posterior nucleus accumbens (Fig. 5B), suggesting that a later age of first age is associated with a more inward deflection of the left posterior nucleus accumbens. This latter finding is similar to that observed by Gilman et al. (2014) who found that more frequent use was associated with an inward deflection of the nucleus accumbens.
Fig. 5.

Effects of marijuana use on subcortical shape in the restricted sample of unrelated users and the group comparison of users and non-users. (A) Trend for a significant negative relationship between number of times used and scalar values in the left amygdala. Participants with a higher number of times used showed more inward deflection in the superficial group of the left amygdala. FWE-corrected p < 0.09. (B) Significant negative relationship between age of first use and scalar values in the left nucleus accumbens. Participants with a earlier age of first use showed more inward deflection in the right posterior ventral accumbens. FWE-corrected p < 0.05. (C) Significant negative association between times used marijuana and scalar values in the left hippocampus. Participants with greater number of lifetime uses showed more inward deflection in the anterior hippocampus cornu ammonius. FWE-corrected p < 0.05. (D) Significant group effect between users and non-users on shape in the right amygdala. Users showed greater outward deflection of the superficial group of the amygdala (left image) and the laterobasal amygdala (right image) compared to non-users. FWE-corrected p < 0.05. (E) Significant group effect between users and non-users on shape in the left hippocampus (view of the medial surface, with right posterior hemisphere cropped). Users showed greater outward deflection of the hippocampus subiculum relative to non-users. FWE-corrected p < 0.05.
When comparing users and non-users, significant effects on shape were discovered in the right amygdala and left hippocampus. There were two clusters in the right amygdala, one in the dorsal superficial group and another in the ventral laterobasal group, that showed more outward deflection in users compared to non-users. Similarly, in the left ventral anterior hippocampus (subiculum), users showed more outward deflection compared to non-users. This region was more ventral and anterior to the cluster in the subiculum that showed a negative association with times used in users. These group results are somewhat surprising, as the correlational analyses within users showed inward deflections in more heavy users. However, the left and right nucleus accumbens also showed opposite patterns of inward and outward deflections with greater use, respectively. Thus, one possibility is that marijuana has differential effects in the left and right hemispheres. Nevertheless, Gilman et al. (2014) found more inward deflections for heavy users in both left and right nucleus accumbens and the right amygdala.
4. Discussion
Despite a trend of de-criminalization and softening societal views, the scientific literature on the effects of marijuana on the brain has not yet reached a consensus. Work with adolescents has consistently shown that heavy marijuana use leads to disruptions in the integrity of white matter (e.g., Becker et al., 2015, Jacobus and Tapert, 2014). However, work investigating the effects of marijuana on the morphometry of gray matter and subcortical regions has yielded inconsistent findings, with several papers even providing strong evidence that heavy marijuana has no effects on brain morphometry (Weiland et al., 2015). The current study took a broad look at this question using a multi-modal neuroimaging approach on data from almost 500 participants reporting varying levels of recreational marijuana use. These data from the HCP include the highest quality neuroimaging data publically available, with almost twice the resolution for structural images (Van Essen et al., 2012), and using state of the art diffusion imaging (Sotiropoulos et al., 2013, Uǧurbil et al., 2013).
This large dataset enabled us to investigate parametric associations between marijuana use and cortical and subcortical brain morphometry and white matter integrity. An advantage of this dataset is that it represents a representative community sample, with great variability in race, ethnicity, gender, education level, mental health symptoms, and drug use (marijuana and otherwise). Moreover, such ‘big data’ sets like the HCP are becoming more and more commonplace (e.g., Nooner et al., 2012), and it will be important to mine these datasets to discover new insights about the functions and organization of the human brain.
Importantly, this data set provided information on age of first use providing information on whether or not use occurred during early adolescence. Adolescence is a time of critical brain development for white matter tracts, with white matter volume increasing into young adulthood (Lebel and Beaulieu, 2011). Heavy marijuana use during this critical period may impact development (Bava et al., 2010), and specifically, a number of frontal and association white matter tracts show decreased white matter integrity with heavy adolescent use (Filbey et al., 2014, Jacobus et al., 2009, Jacobus et al., 2013). Recently, Becker et al. (2015) found that over the course of two years, young adults with an adolescent onset of marijuana use showed reduced longitudinal development of FA in key frontal, central, and parietal white matter tracts. The current findings are largely consistent with those reports. We found that an earlier age of first use was associated with decreased FA and increased RD in long-range tracts including the forceps minor, Superior Longitudinal Fasciculus, and Inferior Longitudinal Fasciculus. Given that the average age of participants in the current study was 29.2, the effects of marijuana on white matter appear to be long lasting, persisting 10–20 years after the first use of marijuana. Future, studies should examine the effects of duration of use on the trajectory of brain development.
With regards to shape, a recent meta-analysis has suggested that the subcortical structure most consistently implicated in showing an effect of marijuana use is the hippocampus (Rocchetti et al., 2013). Consistent with that finding, we observed that anterior aspects of the hippocampus showed shape differences due to the number of times marijuana has been used. This finding is interesting as anterior and posterior subregions of the hippocampus have dissociable roles in memory, with the anterior portion (in particular the cornu ammonis) being involved more in memory encoding and the posterior portion being involved more in memory retrieval (Bartsch et al., 2010, Eldridge et al., 2005, Zeineh et al., 2003); however, an exploratory analysis revealed no connection between hippocampal or amygdala shape and memory performance in these participants (not reported here for the sake of brevity).
Gilman et al. (2014) used a similar multi-modal approach and found that gray matter density (measured by VBM), shape (measured by FIRST), and volume (measured by FreeSurfer) of the left nucleus accumbens was significantly different for recreational users and controls. More specifically, in users, the left and right accumbens showed a negative relationship between shape and marijuana use frequency, such that the accumbens showed more inward deflections with more frequent recent use, as well as greater volume. However, in an attempt to replicate these findings, Weiland et al. (2015) found that the results of Gilman et al. (2014) held only when participants were not matched in terms of alcohol use. When matched for alcohol use, marijuana users and controls showed no significant effects in terms of cortical or subcortical morphometry. Moreover, when Weiland et al. (2015) examined the effect sizes observed in 11 prior studies, they found a net zero effect for the effects of marijuana on brain morphometry (primarily examined in terms of volume). Notably the results of the current study take into consideration the potentially confounding factors of alcohol and tobacco use as well as gender, age, and years of education.
A recent paper using some of the same sample from the HCP (S500 release) examined how genetic vs. environmental factors might contribute to brain volume as a function of marijuana use. They found that marijuana use was associated with smaller volume of the left amygdala and right ventral striatum (Pagliaccio et al., 2015). However, their analyses suggested that the results for the amygdala are likely driven by shared genetic factors as compared to environmental factors as both marijuana user twins and their non-user twin showed reduced volume compared to concordant non-user twins. While the current study did not directly examine the role of genetics and shared environment in mediating the effects of marijuana on the brain, our analyses accounted for family structure. When accounting for family structure, we found no effects of marijuana on the amygdala, and a trend for a more inward deflection of the right accumbens, but no accompanying difference in volume. However, the analysis of Pagliaccio et al. (2015) was limited in sample size for certain sibships, in particular, monozygotic twins discordant for marijuana use. With the upcoming complete HCP dataset consisting of 1200 participants, it will be important to update the analyses of Pagliaccio and colleagues, as well as adding subcortical shape as a measure, to see if a causal relationship arises with a larger sample (potentially three-times larger).
4.1. Limitations
As the current study is not longitudinal but rather cross-sectional, it cannot speak to whether the use of marijuana causes changes in neural structures. Such an analysis will require longitudinal data as is to-be-collected by the new national ABCD (Adolescent Brain and Cognitive Development) study funded by NIH. Nonetheless, the current study can provide hints as to potential effects of marijuana use due to its large sample size and because family structure was controlled for in the analyses. In addition, despite the 100-fold increase in the number of marijuana users from most studies and the high-quality of imaging data, the data on marijuana use history from the HCP is relatively sparse. Whereas alcohol and tobacco were assessed in terms of recent and past use, questions of marijuana use were restricted to self-report measures of the age of first use and the number of times used in the lifetime. As such, it was not possible to accurately examine the effects of duration of use or more specifically, the effects of time since last use. As noted in the Materials and methods Section, the age of first use and number of times used data was coarsely coded using relatively arbitrary ranges. In particular, the number of times used score presented widely different categories for participants to select, ranging from 1–5 times used to “more than 1000.” Moreover, no data was available regarding the recency of this use.
Additionally, while tobacco and alcohol were controlled for using scores selected to best represent the impact of chronic, co-morbid substance use, it is possible that alternative metrics would change the representation of variance due to these substances. As it stands, the alcohol and tobacco use scores used in this presented significant co-variance with age of first use and times used, highlighting both the need to control for these factors and the importance of a data-set large enough to separate the effects of each variable.
While participants were excluded from the HCP for major psychiatric or neurological illness (i.e., illness requiring treatment) participants underwent a psychiatric screening as part of the SSAGA, and psychiatric symptoms were assessed with the NIH Toolbox and the Achenbach Adult Self-Report questionnaire (Achenbach et al., 2005). Examining the interactions of psychiatric symptoms and marijuana use was beyond the scope of the current study, but future studies should examine these effects. Previous studies have shown that adolescent marijuana use is co-morbid with a number of psychopathologies including childhood trauma (Sartor et al., 2013), depression (Diego et al., 2003), attention-deficit hyperactivity disorder (Molina and Pelham, 2003), and psychosis (Arseneault et al., 2004, Forti et al., 2015, Malone et al., 2010). Furthermore, the HCP contains information about parental psychopathology. While much of the psychiatric information available in the HCP has less information than a targeted study of psychopathology, there is enough information for future studies to assess multivariate effects of marijuana use variables and co-morbid psychopathology and other substance use.
Lastly, while the advanced imaging analyses used in this study provide powerful ways to non-invasively understand the anatomical changes occurring with a brain, they are limited in that they cannot speak to the mechanisms whereby marijuana use might influence brain structure. Specifically, they cannot elucidate the microscopic changes responsible for the more macroscopic GM and WM impacts (Mori and Zhang, 2006). For example, while shape changes of the accumbens and hippocampus might reflect inflammation, marijuana has been found to have anti-inflammatory properties (Burstein and Zurier, 2009). Macroscopic morphological changes could be caused by neuronal loss or changes in cytoarchitecture such as neuronal size, dendritic spine density, dendritic length, or synaptic protein levels (Kubicki et al., 2005). As such, morphometry studies can strongly inform where such changes are occurring, but cannot pinpoint the microscopic causes of these structural changes. It is important to note that the two major components of marijuana, Δ-9-tetrahydrocannabinol (THC) and cannabidiol (CBD), have opposite effects behaviorally, symptomatically, and in terms of functional activation of all of the regions-of-interest for the current study (Rocchetti et al., 2013). With legalization of marijuana comes more accurate assays of THC and CBD concentrations, and thus, future research can and should focus on examining whether THC and CBD have dissociable effects on brain morphometry (for a similar argument, see Weiland et al., 2015).
5. Conclusion & future directions
Examined from multiple perspectives including white matter integrity, subcortical shape, and brain volume, our parametric analyses suggest that an early onset of marijuana use may be associated with subtle changes in brain regions implicated as being altered in substance abuse. These findings provide for the possibility that marijuana use during adolescence, which is a time of rapid brain development, might, at least in some individuals, have long-lasting effects, independent of the genetic effects suggested by a recent analysis of cortical volume on a somewhat overlapping sample (Pagliaccio et al., 2015). The results of the current study show the utility of analyzing data from data sets with large Ns, as they can reveal subtle effects that might otherwise appear to be contradictory with smaller sample sizes. The current study contains 75% of the target sample of 1200 participants for the HCP, and thus, represents a preliminary analysis that should be re-examined with the full data release.
Financial disclosures
All of the authors report no biomedical financial interests or potential conflicts of interest.
Acknowledgements
This work was supported by the National Institute on Drug Abuse (F32DA034412 to JMO; 1U01DA041120 to MTB). Data were provided by the Human Connectome Project, WU-Minn Consortium (Principal Investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University. This work utilized the Janus supercomputer, which is supported by the National Science Foundation (award number CNS-0821794) and the University of Colorado Boulder. The Janus supercomputer is a joint effort of the University of Colorado Boulder, the University of Colorado Denver and the National Center for Atmospheric Research.
References
- Achenbach T.M., Krukowski R.A., Dumenci L., Ivanova M.Y. Assessment of adult psychopathology: meta-analyses and implications of cross-informant correlations. Psychol. Bull. 2005;131:361–382. doi: 10.1037/0033-2909.131.3.361. [DOI] [PubMed] [Google Scholar]
- Amunts K., Kedo O., Kindler M., Pieperhoff P., Mohlberg H., Shah N.J., Habel U., Schneider F., Zilles K. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps. Anat. Embryol. (Berl.) 2005;210:343–352. doi: 10.1007/s00429-005-0025-5. [DOI] [PubMed] [Google Scholar]
- Andersson J.L.R., Jenkinson M., Smith S.M. 2007. Non-Linear Registration aka Spatial Normalisation. [Google Scholar]
- Arnone D., Barrick T.R., Chengappa S., Mackay C.E., Clark C.A., Abou-Saleh M.T. Corpus callosum damage in heavy marijuana use: preliminary evidence from diffusion tensor tractography and tract-based spatial statistics. NeuroImage. 2008;41:1067–1074. doi: 10.1016/j.neuroimage.2008.02.064. [DOI] [PubMed] [Google Scholar]
- Arseneault L., Cannon M., Witton J., Murray R.M. Causal association between cannabis and psychosis: examination of the evidence. Br. J. Psychiatry. 2004;184:110–117. doi: 10.1192/bjp.184.2.110. [DOI] [PubMed] [Google Scholar]
- Bartsch T., Schönfeld R., Müller F.J., Alfke K., Leplow B., J A., G D., Koch J.M. Focal lesions of human hippocampal. Science (80-.) 2010;328:1412–1416. doi: 10.1126/science.1188160. [DOI] [PubMed] [Google Scholar]
- Bava S., Jacobus J., Mahmood O., Yang T.T., Tapert S.F. Neurocognitive correlates of white matter quality in adolescent substance users. Brain Cogn. 2010;72:347–354. doi: 10.1016/j.bandc.2009.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bava S., Jacobus J., Thayer R.E., Tapert S.F. Longitudinal changes in white matter integrity among adolescent substance users. Alcohol. Clin. Exp. Res. 2013;37:E181–E189. doi: 10.1111/j.1530-0277.2012.01920.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker M.P., Collins P.F., Lim K.O., Muetzel R.L., Luciana M. Longitudinal changes in white matter microstructure after heavy cannabis use. Dev. Cogn. Neurosci. 2015;16:23–35. doi: 10.1016/j.dcn.2015.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellis M.D., Narasimhan A., Thatcher D.L., Keshavan M.S., Soloff P., Clark D.B. Prefrontal cortex, thalamus, and cerebellar volumes in adolescents and young adults with adolescent-onset alcohol use disorders and comorbid mental disorders. Alcohol. Clin. Exp. Res. 2005;29:1590–1600. doi: 10.1097/01.alc.0000179368.87886.76. [DOI] [PubMed] [Google Scholar]
- Berridge K.C., Robinson T.E. Parsing reward. Trends eurosci. 2003;26:507–513. doi: 10.1016/S0166-2236(03)00233-9. [DOI] [PubMed] [Google Scholar]
- Block R.I., O'Leary D.S., Ehrhardt J.C., Augustinack J.C., Ghoneim M.M., Arndt S., Hall J.a. Effects of frequent marijuana use on brain tissue volume and composition. Neuroreport. 2000;11:491–496. doi: 10.1097/00001756-200002280-00013. [DOI] [PubMed] [Google Scholar]
- Burstein S.H., Zurier R.B. Cannabinoids, endocannabinoids, and related analogs in inflammation. AAPS J. 2009;11:109–119. doi: 10.1208/s12248-009-9084-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churchwell J.C., Lopez-Larson M.P., Yurgelun-Todd D.A. Altered frontal cortical volume and decision making in adolescent cannabis users. Front. Psychol. 2010;1 doi: 10.3389/fpsyg.2010.00225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeLisi L.E., Bertisch H.C., Szulc K.U., Majcher M., Brown K., Bappal A., Ardekani B.A. A preliminary DTI study showing no brain structural change associated with adolescent cannabis use. Harm Reduct. J. 2006;3:17. doi: 10.1186/1477-7517-3-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depue B.E., Orr J.M., Smolker H.R., Naaz F., Banich M.T. The organization of right prefrontal networks reveals common mechanisms of inhibitory regulation across cognitive, emotional, and motor processes. Cereb. Cortex. 2016;26:1634–1646. doi: 10.1093/cercor/bhu324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diego M.A., Field T.M., Sanders C.E. Academic performance, popularity, and depression predict adolescent substance use. Adolescence. 2003;38:35–42. [PubMed] [Google Scholar]
- Douaud G., Smith S.M., Jenkinson M., Behrens T.E.J., Johansen-Berg H., Vickers J., James S., Voets N., Watkins K., Matthews P.M., James A. Anatomically related grey and white matter abnormalities in adolescent-onset schizophrenia. Brain. 2007;130:2375–2386. doi: 10.1093/brain/awm184. [DOI] [PubMed] [Google Scholar]
- Eldridge L.L., Engel S.A., Zeineh M.M., Bookheimer S.Y., Knowlton B.J. A dissociation of encoding and retrieval processes in the human hippocampus. J. Neurosci. 2005;25:3280–3286. doi: 10.1523/JNEUROSCI.3420-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filbey F.M., Aslan S., Calhoun V.D., Spence J.S., Damaraju E., Caprihan A., Segall J. Long-term effects of marijuana use on the brain. Proc. Natl. Acad. Sci. U. S. A. 2014;111:16913–16918. doi: 10.1073/pnas.1415297111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forti M.D., Marconi A., Carra E., Fraietta S., Trotta A., Bonomo M., Bianconi F., Gardner-sood P., Connor J.O., Russo M., Stilo S.A., Marques T.R., Mondelli V., Dazzan P., Pariante C., David A.S., Gaughran F., Atakan Z., Iyegbe C., Powell J., Morgan C., Lynskey M., Murray R.M. Proportion of patients in south London with first-episode psychosis attributable to use of high potency cannabis: a case-control study. Lancet Psychiatry. 2015;366:1–6. doi: 10.1016/S2215-0366(14)00117-5. [DOI] [PubMed] [Google Scholar]
- Gilman J.M., Kuster J.K., Lee S., Lee M.J., Kim B.W., Makris N., van der Kouwe A., Blood A.J., Breiter H.C. Cannabis use is quantitatively associated with nucleus accumbens and amygdala abnormalities in young adult recreational users. J. Neurosci. 2014;34:5529–5538. doi: 10.1523/JNEUROSCI.4745-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glasser M.F., Sotiropoulos S.N., Wilson J.A., Coalson T.S., Fischl B., Andersson J.L.R., Xu J., Jbabdi S., Webster M., Polimeni J.R., Van Essen D.C., Jenkinson M. The minimal preprocessing pipelines for the human connectome project. NeuroImage. 2013;80:105–124. doi: 10.1016/j.neuroimage.2013.04.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Good C.D., Johnsrude I.S., Ashburner J., Henson R.N., Friston K.J., Frackowiak R.S. A voxel-based morphometric study of ageing in 465 normal adult human brains. NeuroImage. 2001;14:21–36. doi: 10.1006/nimg.2001.0786. [DOI] [PubMed] [Google Scholar]
- Gruber S.A., Yurgelun-Todd D.A. Neuroimaging of marijuana smokers during inhibitory processing: a pilot investigation. Cogn. Brain Res. 2005;23:107–118. doi: 10.1016/j.cogbrainres.2005.02.016. [DOI] [PubMed] [Google Scholar]
- Jacobus J., McQueeny T., Bava S., Schweinsburg B.C., Frank L.R., Yang T.T., Tapert S.F. White matter integrity in adolescents with histories of marijuana use and binge drinking. Neurotoxicol. Teratol. 2009;31:349–355. doi: 10.1016/j.ntt.2009.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobus J., Squeglia L.M., Infante M.A., Bava S., Tapert S.F. White matter integrity pre- and post-marijuana and alcohol initiation in adolescence. Brain Sci. 2013;3:396–414. doi: 10.3390/brainsci3010396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobus J., Tapert S.F. Effects of cannabis on the adolescent brain. Curr. Pharm. Des. 2014;20:2186–2193. doi: 10.2174/13816128113199990426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klawiter E.C., Schmidt R.E., Trinkaus K., Liang H.F., Budde M.D., Naismith R.T., Song S.K., Cross A.H., Benzinger T.L. Radial diffusivity predicts demyelination in ex vivo multiple sclerosis spinal cords. NeuroImage. 2011;55:1454–1460. doi: 10.1016/j.neuroimage.2011.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolb B., Gorny G.G., Limebeer C.L., Parker L.A. Chronic treatment with delta-9-tetrahydrocannabinol alters the structure of neurons in the nucleus accumbens shell and medial prefrontal cortex of rats. Synapse. 2006;60:429–436. doi: 10.1002/syn.20313. [DOI] [PubMed] [Google Scholar]
- Koob G.F., Volkow N.D. Neurocircuitry of addiction. Neuropsychopharmacology. 2010;35:217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubicki M., Park H., Westin C.F., Nestor P.G., Mulkern R.V., Maier S.E., Niznikiewicz M., Connor E.E., Levitt J.J., Frumin M., Kikinis R., Jolesz F.a., McCarley R.W., Shenton M.E. DTI and MTR abnormalities in schizophrenia: analysis of white matter integrity. NeuroImage. 2005;26:1109–1118. doi: 10.1016/j.neuroimage.2005.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebel C., Beaulieu C. Longitudinal development of human brain wiring continues from childhood into adulthood. J. Neurosci. 2011;31:10937–10947. doi: 10.1523/JNEUROSCI.5302-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malone D.T., Hill M.N., Rubino T. Adolescent cannabis use and psychosis: epidemiology and neurodevelopmental models. Br. J. Pharmacol. 2010;160:511–522. doi: 10.1111/j.1476-5381.2010.00721.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marijuana Resource Center: State Laws Related to Marijuana [WWW document] 2016. https://www.whitehouse.gov/ondcp/state-laws-related-to-marijuana (URL) (accessed 3.5.16)
- Molina B.S.G., Pelham W.E., Jr. Childhood predictors of adolescent substance use in a longitudinal study of children with ADHD. J. Abnorm. Psychol. 2003;112:497–507. doi: 10.1037/0021-843x.112.3.497. (10.1037/0021-843X.112.3.497) [DOI] [PubMed] [Google Scholar]
- Mori S., Zhang J. Principles of diffusion tensor imaging and its applications to basic neuroscience research. Neuron. 2006;51:527–539. doi: 10.1016/j.neuron.2006.08.012. [DOI] [PubMed] [Google Scholar]
- Nooner K.B., Colcombe S.J., Tobe R.H., Mennes M., Benedict M.M., Moreno A.L., Panek L.J., Brown S., Zavitz S.T., Li Q., Sikka S., Gutman D., Bangaru S., Schlachter R.T., Kamiel S.M., Anwar A.R., Hinz C.M., Kaplan M.S., Rachlin A.B., Adelsberg S., Cheung B., Khanuja R., Yan C., Craddock R.C., Calhoun V., Courtney W., King M., Wood D., Cox C.L., Kelly A.M.C., Di Martino A., Petkova E., Reiss P.T., Duan N., Thomsen D., Biswal B., Coffey B., Hoptman M.J., Javitt D.C., Pomara N., Sidtis J.J., Koplewicz H.S., Castellanos F.X., Leventhal B.L., Milham M.P. The NKI-Rockland sample: a model for accelerating the pace of discovery science in psychiatry. Front. Neurosci. 2012;6:1–11. doi: 10.3389/fnins.2012.00152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagliaccio D., Barch D.M., Bogdan R., Wood P.K., Lynskey M.T., Heath A.C., Agrawal A. Shared predisposition in the association between cannabis use and subcortical brain structure. JAMA Psychiatry. 2015;63130:1–8. doi: 10.1001/jamapsychiatry.2015.1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patenaude B., Smith S.M., Kennedy D.N., Jenkinson M. A Bayesian model of shape and appearance for subcortical brain segmentation. NeuroImage. 2011;56:907–922. doi: 10.1016/j.neuroimage.2011.02.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierpaoli C., Basser P.J. Toward a quantitative assessment of diffusion anisotropy. Magn. Reson. Med. 1996;36:893–906. doi: 10.1002/mrm.1910360612. [DOI] [PubMed] [Google Scholar]
- Pope H.G., Gruber A.J., Hudson J.I., Cohane G., Huestis M.A., Yurgelun-Todd D.A. Early-onset cannabis use and cognitive deficits: what is the nature of the association? Drug Alcohol Depend. 2003;69:303–310. doi: 10.1016/s0376-8716(02)00334-4. [DOI] [PubMed] [Google Scholar]
- Rentería M.E., Hansell N.K., Strike L.T., McMahon K.L., de Zubicaray G.I., Hickie I.B., Thompson P.M., Martin N.G., Medland S.E., Wright M.J. Genetic architecture of subcortical brain regions: common and region-specific genetic contributions. Genes Brain Behav. 2014;13:821–830. doi: 10.1111/gbb.12177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocchetti M., Crescini A., Borgwardt S., Caverzasi E., Politi P., Atakan Z., Fusar-Poli P. Is cannabis neurotoxic for the healthy brain? A meta-analytical review of structural brain alterations in non-psychotic users. Psychiatry Clin. Neurosci. 2013;67:483–492. doi: 10.1111/pcn.12085. [DOI] [PubMed] [Google Scholar]
- Sartor C.E., Waldron M., Duncan A.E., Grant J.D., Mccutcheon V.V., Nelson E.C., Madden P.A.F., Bucholz K.K., Heath A.C. Childhood sexual abuse and early substance use in adolescent girls: the role of familial influences. Addiction. 2013;108:993–1000. doi: 10.1111/add.12115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacht J.P., Hutchison K.E., Filbey F.M. Associations between cannabinoid receptor-1 (CNR1) variation and hippocampus and amygdala volumes in heavy cannabis users. Neuropsychopharmacology. 2012;37:2368–2376. doi: 10.1038/npp.2012.92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuermeyer J., Salomonsen-Sautel S., Price R.K., Balan S., Thurstone C., Min S.J., Sakai J.T. Temporal trends in marijuana attitudes, availability and use in Colorado compared to non-medical marijuana states: 2003–11. Drug Alcohol Depend. 2014;140:145–155. doi: 10.1016/j.drugalcdep.2014.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmonds D.J., Hallquist M.N., Asato M., Luna B. Developmental stages and sex differences of white matter and behavioral development through adolescence: a longitudinal diffusion tensor imaging (DTI) study. NeuroImage. 2014;92:356–368. doi: 10.1016/j.neuroimage.2013.12.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith S.M., Jenkinson M., Johansen-Berg H., Rueckert D., Nichols T.E., Mackay C.E., Watkins K.E., Ciccarelli O., Cader M.Z., Matthews P.M., Behrens T.E.J. Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. NeuroImage. 2006;31:1487–1505. doi: 10.1016/j.neuroimage.2006.02.024. [DOI] [PubMed] [Google Scholar]
- Smith S.M., Jenkinson M., Woolrich M.W., Beckmann C.F., Behrens T.E.J., Johansen-Berg H., Bannister P.R., De Luca M., Drobnjak I., Flitney D.E., Niazy R.K., Saunders J., Vickers J., Zhang Y., De Stefano N., Brady J.M., Matthews P.M. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage. 2004;23(Suppl. 1):S208–S219. doi: 10.1016/j.neuroimage.2004.07.051. [DOI] [PubMed] [Google Scholar]
- Smith S.M., Nichols T.E. Threshold-free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. NeuroImage. 2009;44:83–98. doi: 10.1016/j.neuroimage.2008.03.061. [DOI] [PubMed] [Google Scholar]
- Smolker H.R., Depue B.E., Reineberg A.E., Orr J.M., Banich M.T. Individual differences in regional prefrontal grey matter morphometry and fractional anisotropy are associated with different constructs of executive function. Brain Struct. Funct. 2014;220:1291–1306. doi: 10.1007/s00429-014-0723-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotiropoulos S.N., Jbabdi S., Xu J., Andersson J.L.R., Moeller S., Auerbach E.J., Glasser M.F., Hernandez M., Sapiro G., Jenkinson M., Feinberg D.A., Yacoub E., Lenglet C., Van Essen D.C., Ugurbil K., Behrens T.E.J. Advances in diffusion MRI acquisition and processing in the human connectome project. NeuroImage. 2013;80:125–143. doi: 10.1016/j.neuroimage.2013.05.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uǧurbil K., Xu J., Auerbach E.J., Moeller S., Vu A.T., Duarte-Carvajalino J.M., Lenglet C., Wu X., Schmitter S., Van de Moortele P.F., Strupp J., Sapiro G., De Martino F., Wang D., Harel N., Garwood M., Chen L., Feinberg D.A., Smith S.M., Miller K.L., Sotiropoulos S.N., Jbabdi S., Andersson J.L.R., Behrens T.E.J., Glasser M.F., Van Essen D.C., Yacoub E. Pushing spatial and temporal resolution for functional and diffusion MRI in the human connectome project. NeuroImage. 2013;80:80–104. doi: 10.1016/j.neuroimage.2013.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Essen D.C., Ugurbil K., Auerbach E.J., Barch D.M., Behrens T.E.J., Bucholz R., Chang A., Chen L., Corbetta M., Curtiss S.W., Della Penna S., Feinberg D.A., Glasser M.F., Harel N., Heath A.C., Larson-Prior L., Marcus D., Michalareas G., Moeller S., Oostenveld R., Petersen S.E., Prior F., Schlaggar B.L., Smith S.M., Snyder A.Z., Xu J., Yacoub E. The human connectome project: a data acquisition perspective. NeuroImage. 2012;62:2222–2231. doi: 10.1016/j.neuroimage.2012.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow N.D., Baler R.D., Compton W.M., Weiss S.R.B. Adverse health effects of marijuana use. N. Engl. J. Med. 2014;370:2219–2227. doi: 10.1056/NEJMra1402309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiland B.J., Thayer R.E., Depue B.E., Sabbineni A., Bryan A.D., Hutchison K.E. Daily marijuana use is not associated with brain morphometric measures in adolescents or adults. J. Neurosci. 2015;35:1505–1512. doi: 10.1523/JNEUROSCI.2946-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winkler A.M., Ridgway G.R., Webster M.a., Smith S.M., Nichols T.E. Permutation inference for the general linear model. NeuroImage. 2014;92:381–397. doi: 10.1016/j.neuroimage.2014.01.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winkler A.M., Webster M.A., Vidaurre D., Nichols T.E., Smith S.M. Multi-level block permutation. NeuroImage. 2015;123:253–268. doi: 10.1016/j.neuroimage.2015.05.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wrase J., Makris N., Braus D.F., Mann K., Smolka M.N., Kennedy D.N., Caviness V.S., Hodge S.M., Tang L., Albaugh M., Ziegler D.A., Davis O.C., Kissling C., Schumann G., Breiter H.C., Heinz A. Amygdala volume associated with alcohol abuse relapse and craving. Am. J. Psychiatry. 2008;165:1179–1184. doi: 10.1176/appi.ajp.2008.07121877. [DOI] [PubMed] [Google Scholar]
- Yendiki A., Koldewyn K., Kakunoori S., Kanwisher N., Fischl B. Spurious group differences due to head motion in a diffusion MRI study. NeuroImage. 2014;88:79–90. doi: 10.1016/j.neuroimage.2013.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeineh M.M., Engel S.A., Thompson P.M., Bookheimer S.Y. Dynamics of the hippocampus during encoding and retrieval of face-name pairs. Science (80-.) 2003;299:577–580. doi: 10.1126/science.1077775. [DOI] [PubMed] [Google Scholar]