

Abstract
Equisetum ramosissimum, a genus of Equisetaceae, is a medicinal plant that can be separated into ethyl acetate (EA), dichloromethane (DM), n-hexane (Hex), methanol (MeOH), and water extracts. EA extract was known to have potent antioxidative properties, reducing power, DPPH scavenging activity, and metal ion chelating activity. This study compared these five extracts in terms of their inhibiting effects on three human malignant melanomas: A375, A375.S2, and A2058. MTT assay presented the notion that both EA and DM extracts inhibited melanoma growth but did not affect the viabilities of normal dermal keratinocytes (HaCaT) or fibroblasts. Western blot analyses showed that both EA and DM extracts induced overexpression of caspase proteins in all three melanomas. To determine their roles in melanogenesis, this study analyzed their in vitro suppressive effects on mushroom tyrosinase. All extracts except for water revealed moderate suppressive effects. None of the extracts affected B16-F10 cells proliferation. EA extract inhibited cellular melanin production whereas DM extract unexpectedly enhanced cellular pigmentation in B16-F10 cells. Data for modulations of microphthalmia-associated transcription factor, tyrosinase, tyrosinase-related protein 1, and tyrosinase-related protein 2 showed that EA extract inhibited protein expression mentioned above whereas DM extract had the opposite effect. Overall, the experiments indicated that the biofunctional activities of EA extract contained in food and cosmetics protect against oxidation, melanoma, and melanin production.
1. Introduction
Mitochondria, chloroplasts, and peroxisomes produce reactive oxygen species (ROS) through respiration and photosynthesis [1, 2]. Studies suggested that the changes in cellular homeostasis caused by high ROS levels can result in oxidative damage [3]. To avoid ROS oxidative injuries, the defense radical scavenging systems used by the human being were separated into enzymatic and nonenzymatic mechanisms. Antioxidant enzymes and substances could reduce oxidative damage by decreasing production of ROS and radicals [2]. These agents include glutathione and catalase, glutathione reductase, superoxide dismutase, and glutathione peroxidase. Others include α-lipoic acid, carotenoids, coenzyme Q10, flavonoids, antioxidative minerals (copper, zinc, manganese, and selenium), and cofactors (folic acid and vitamins A, B1, B2, B6, B12, C, and E). Generally, the above antioxidative materials are applied in synergic ways with each other against various free radical types [1–3].
Malignant melanoma is among the most incursive and life-threatening malignant tumors [4]. Skin cancer can result from exposure to ultraviolet (UV) radiation emitted by the sun and by halogen lamps. Experimental studies of metastatic melanoma are very challenging because systemic treatments are often ineffective and the rapid spread of melanoma cells to retain an intensive property of the cellular spreading which happens later is pathologically confusing [5]. Although melanoma is not a major cause of tumorous symptoms, melanoma is a major cause of death in patients with skin cancer. Treating melanoma is difficult due to its resistance to conventional chemoradiotherapy [6]. No effective therapies for metastatic melanoma are currently available, and effective drugs are urgently needed.
Melanocytes are located in the basal epidermal layer and in the hair follicles. Generally, UV radiation produces pigment by diametric stimulation of melanocytes [7]. Melanin pigmentation has been recognized by many factors; the permeation of sunlight is a well recognized source of melanin pigmentation; specifically, UV radiation causes darkening of the skin and/or sunburn. UV is the most common reason of changes in the visible countenance of human skin. Unusual melanogenesis is a characteristic of many human skin disorders, including abnormal pigmentation, nevi, and melanoma [8].
Many recent studies have investigated the biological functions of natural extracts and their potential applications as health foods, as active ingredients in cosmetics, and as leading compounds in new medicines [9, 10]. Equisetum ramosissimum, a genus of Equisetaceae, is a medicinal plant administered to treat hemorrhage, urethritis, jaundice, and hepatitis [11]. Although the antioxidant activities of E. ramosissimum were identified [12], its biological activities have not been examined. Therefore, this study elucidated the potential protective effects of E. ramosissimum extract against oxidation, melanoma, and melanogenesis.
2. Materials and Methods
2.1. Chemicals and Reagents
Ascorbic acid (vitamin C), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), L-3,4-dihydroxyphenylalanine (L-DOPA), dimethyl sulfoxide (DMSO), 1,1-diphenyl-2-picrylhydrazyl (DPPH), ethanol, ethylenediaminetetraacetic acid (EDTA), ferrous chloride (FeCl2·4H2O), ferric chloride (FeCl3), kojic acid, methanol, potassium ferricyanide (K3Fe(CN)6), 3-tert-butyl-4-hydroxyanisole (BHA), and L-tyrosine were purchased from Sigma-Aldrich Company (St. Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were obtained from Gibco BRL (Gaithersburg, MD, USA). Other chemical buffers and reagents were purchased at the highest available purity and quality.
2.2. Plant Material Extraction and Isolation
Two authors of this study (Dr. Chieh-Chih Shih and Professor Zhi-Hong Wen) prepared the extracts as follows. First, methanol (2.2 L) was requested in two extraction procedures of E. ramosissimum from a powder consisting of ground E. ramosissimum plant, and the extract was then refluxed for 30 minutes. After a filtering procedure, the extract was concentrated to obtain methanol (MeOH) crude extract, 9.52 g. The crude extract was added to distilled water (200 mL) and then partitioned with n-hexane (Hex, 1.95 g), dichloromethane (DM, 0.67 g), and ethyl acetate (EA, 0.26 g), which left an aqueous layer (H2O, 6.44 g). All fractions were concentrated, freeze-dried, dissolved in DMSO to obtain a stock solution (500 mg/mL), and then diluted with DMEM to the required concentrations.
2.3. Assays of Antioxidant Effects
2.3.1. Reducing Power Assay
Assays of the reducing power of the crude extracts were performed as described in the literature [13]. Briefly, dissimilar concentrations of each extract were blended with 85 μL of 67 mM sodium phosphate buffer (pH 6.8) and 2.5 μL of 20% K3Fe(CN)6. The admixture was kept at 50°C for 20 min. After addition of 160 μL 10% TCA, the admixture was centrifuged for 10 min at 3,000 g. The supernatant (75 μL) was mixed with 2% FeCl3 (25 μL), and absorbance was read with a spectrophotometer (BioTek Co., Winooski, VT, USA) at 700 nm with a BHA solution as a positive control. High absorbance was interpreted as a high capacity for metal ion reduction.
2.3.2. DPPH• Radical Scavenging Ability Assay
DPPH is a stable free radical with a violet color. Reaction of DPPH• with an antioxidant provides hydrogen, which results in decreased absorbance at 517 nm. DPPH assay was performed as described previously, with some minor modification [13]. Various concentrations of E. ramosissimum extracts were added to 100 μL of aqueous stable DPPH• (60 μM) solution and allowed to stand at room temperature for 60 min. Vitamin C was used as a positive control. Low absorbance was interpreted as a high DPPH scavenging ability. The calculation for free radical scavenging activity (%) is done as follows:
| (1) |
2.3.3. Metal Chelating Activity
The chelating properties of ferrous ions (Fe2+) were explored using a method described previously [14]. Briefly, various concentrations of extract were dissolved in DMSO and added to a 10 μL solution of FeCl2·4H2O (2 mM). Next, 20 μL ferrozine (5 mM) was added, and the admixture was shaken vigorously for 10 minutes. The absorbance was 562 nm. EDTA was used as a positive control, and the formula employed to calculate metal chelating activity is as follows:
| (2) |
2.4. Cell Line Cultures
Human and rat melanoma cell lines were obtained from Bioresource Collection and Research Center (Taiwan): A375 (BCRC number 60039), A375.S2 (BCRC number 60263), A2058 (BCRC number 60240), and B16-F10 (BCRC number 60031). Human fibroblasts were separated from the foreskin primary culture (Institutional Review Board, KMUH-IRB-990269). Cells were cultured in DMEM supplemented with 10% FBS and 1% antibiotics. Human skin keratinocytes, HaCaT cells, were cultured in Keratinocyte-SFM (Gibco, USA) supplemented with bovine pituitary extract and human recombinant epidermal growth factor. All cell lines were cultured in 5% CO2 at 37°C.
2.5. MTT Assay of Cell Viability
The influences of extracts on cell development were estimated with MTT assay [15]. Cells were seeded at 8 × 103 cells/well in 96-well plates and incubated for 24 h before addition of extracts. After 24 h, MTT solution was dispensed into each well. After 2 h, the culture medium was discarded, and DMSO was added to each well. The absorbance of the formazan salt was 595 nm, and the cell viability was computed as follows:
| (3) |
2.6. Western Blot Analysis
This analysis was performed as described previously with some minor modifications [16]. 1 × 106 cells were treated with extracts or with the vehicle control for 24 h, and the cells were then harvested and lysed with RIPA lysis buffer. Equal amounts of protein were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and next transferred into a polyvinylidene fluoride membrane. The membranes were incubated with corresponding primary antibodies and afterwards incubated with secondary antibodies corresponding with the primary antibodies. The signals were visualized with a chemiluminescence detection kit (Amersham, Piscataway, NJ, USA).
2.7. Mushroom Tyrosinase Measurement
Mushroom tyrosinase activity was evaluated as described previously with some minor modifications [17]. Samples were incubated with mushroom tyrosinase (25 U/mL), and L-tyrosine (2 mM) in phosphate buffer (pH 6.8) was added. The mixtures were then kept at 37°C for 30 minutes. Kojic acid was used as a positive control. Tyrosinase inhibitory activity was determined by the following equation:
| (4) |
where A is the optical density (OD490) without testing extract; B is OD490 with tyrosinase and without testing extract; C is OD490 with testing extract; and D is OD490 with tyrosinase and without testing extract.
2.8. Melanin Quantification Assessment
This analysis was demonstrated as described in the literature with some minor modifications [17]. Cell pellets were liquefied with 1.0 N NaOH, warmed to 80°C for 1 hour, and centrifuged at 10,000 g for 10 minutes. The quantity of melanin was determined by extracting the supernatant and running it through the spectrophotometer, which gave a result of 475 nm.
2.9. Statistical Analysis
Biofunctional assays of the E. ramosissimum extracts in each platform were performed in triplicate. Results were expressed as means ± SD. Analysis of variance was used for data analysis. A p value less than 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Antioxidant Activity of E. ramosissimum
Free radicals have roles in signal travel and in physiological, metabolic, and immune reactions. Even though free radicals are needed for normal healthy biochemical processes in the body, they have severe negative health effects [18, 19]. One of the study aims was to test antioxidative properties, which were examined by ferric reducing powers, DPPH radical scavenging capacities, and metal chelating power activities.
First, a simple, rapid, and reliable test was used to measure Fe(III)-ferricyanide complex synthesis. In this test, the extracts reducing properties of E. ramosissimum were indicated by changes in the color of the solution (from light yellow to different shades of green and blue). Table 1 presented the notion that the reducing power of EA extract resulted in stronger dose-dependent suppressive effects compared to the other four extracts and showed the highest scavenging of 0.87 ± 0.23 at 200 μg/mL.
Table 1.
Antioxidant activities of E. ramosissimum extracts, including reducing power, DPPH free radical scavenging activity, and ferrous ion chelating power.
| Extracts (μg/mL) | Reducing power (OD700) | DPPH scavenging capacity (%) | Chelating activity (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 50 | 100 | 200 | 50 | 100 | 200 | 50 | 100 | 200 | |
| EA | 0.60 ± 0.06 | 0.79 ± 0.06 | 0.87 ± 0.23 | <10.0 | 15.23 ± 2.76 | 43.41 ± 7.68 | 20.47 ± 3.57 | 34.69 ± 5.03 | 44.56 ± 1.32 |
| DM | 0.59 ± 0.03 | 0.65 ± 0.07 | 0.68 ± 0.04 | <10.0 | <10.0 | <10.0 | 15.57 ± 1.39 | 17.49 ± 0.13 | 20.19 ± 1.72 |
| Hex | 0.56 ± 0.11 | 0.65 ± 0.10 | 0.68 ± 0.03 | <10.0 | <10.0 | <10.0 | 17.98 ± 4.50 | 20.71 ± 0.33 | 22.53 ± 6.07 |
| MeOH | 0.58 ± 0.01 | 0.67 ± 0.13 | 0.73 ± 0.02 | <10.0 | 10.69 ± 1.66 | 15.60 ± 1.28 | 10.63 ± 1.65 | 18.89 ± 1.19 | 20.43 ± 0.07 |
| Water | 0.62 ± 0.03 | 0.65 ± 0.10 | 0.70 ± 0.01 | <10.0 | <10.0 | 14.87 ± 0.73 | 10.77 ± 0.79 | 13.33 ± 4.55 | 13.52 ± 0.59 |
| Vitamin Ca | — | — | — | — | 85.55 ± 0.48 | — | — | — | — |
| BHAb | — | 1.64 ± 0.29 | — | — | — | — | — | — | — |
| EDTAc | — | — | — | — | — | — | — | 95.49 ± 0.05 | — |
All statistics are presented as average values ± SD; n = 3. aVitamin C (100 mM) was utilized as a positive control for DPPH assay; bEDTA (100 mM) was used as a positive control for analysis of metal chelating ability; cBHA (100 mM) was applied as a positive control for analysis of reducing power. Assays not performed in this study were indicated by dashes.
The second oxidation inhibitory assay was DPPH radical scavenging test. As antioxidants stabilize DPPH radicals, the color of DPPH solution changes from violet to yellow as diphenylpicrylhydrazine is formed. Table 1 illustrated the results for the five extracts, and the comparisons presented the notion that the EA extract had the strongest radical scavenging effects in a dose-dependent manner.
Within the oxidative conditions, ferrozine can develop complexes with Fe2+ to be measured quantitatively. In the presence of chelating materials, complex constructions are dislocated, which lightens the red color of the complex. Table 1 demonstrated the data for the last evaluation of antioxidative properties. The extracts showed low-to-moderate Fe2+ scavenging activities at concentrations of 50–200 μg/mL, and EA extract possessed the highest value of 44.56 ± 1.32 at 200 μg/mL.
3.2. Cytotoxicity of E. ramosissimum on Human Melanoma Cells and Normal Cells
Melanoma is a malignant tumor that starts in a certain type of skin cell and is activated when the abnormal cells in the affected part of the body begin to proliferate in an uncontrolled manner [20]. Metastatic malignant melanomas are highly resistant to existing therapies and have a very poor prognosis, and thus new treatment strategies are urgently needed. In early stages of the development of new chemoprotective substances, the main considerations are normal cell allergic responses, sensitivity and potential reactions, and toxic side effects [9]. MTT method was applied to evaluate the cytotoxic effectivenesses of E. ramosissimum extracts on normal human skin cells, including epidermal keratinocytes (HaCaT) and dermal fibroblasts in Figure 1. These two cells were treated with various concentrations (0 to 100 μg/mL) to compare dose-dependent impacts. In both cells, high doses (100 μg/mL) of the five E. ramosissimum extracts had minor effects, and all cellular viabilities exceeded 65% after a 24-hour treatment. That is, the E. ramosissimum extracts had no severe discernible toxic effects on human normal cells.
Figure 1.

Effects of E. ramosissimum extracts on viability of normal human cells according to MTT assay. Suppression of cell viability was measured in (a) HaCaT and (b) fibroblast cells cultured with 5, 10, 25, 50, and 100 μg/mL EA, DM, Hex, MeOH, and water extracts. The results for the control group cultured without extracts were shown on the left (gray line). All experimental data were presented as average values ± SD; n = 3; ∗ p < 0.05.
After establishing the fact that the E. ramosissimum extracts did not cause major injuries to normal cells, the melanoma platforms were used to investigate their antiproliferative effects. Cell line A375 was from a malignant melanoma of a 54-year-old female; and cell line A375.S2 comprised cells which were differentiated from A375 cells; and cell line A2058 comprised highly invasive melanoma cells [21]. In Figure 2, we demonstrated the cytotoxicities of the three cell lines, notably, and high doses (50 and 100 μg/mL) of the EA, DM, and Hex extracts from E. ramosissimum had much larger influences on A375 and A375.S2 cells compared to the A2058 cells. For example, after 24 h treatment with 50 and 100 μg/mL EA extract, the viabilities of the A375 and A375.S2 cells decreased to 30% whereas the viability of the A2058 cells only decreased to 50%. MeOH extract had no effect on the A2058 cells, and the water extract had no apparent cytotoxic effect on the three melanoma cell types. According to our statistical data, the E. ramosissimum extracts have little harmful effects on normal skin cells, and these extracts, particularly EA, DM, and Hex extracts, actually inhibit melanoma cellular proliferation.
Figure 2.

Effects of E. ramosissimum extracts on viability of human melanoma cells according to MTT assay. Suppression of cell viability was measured in (a) A375, (b) A375.S2, and (c) A2058 cells cultured with 5, 10, 25, 50, and 100 μg/mL EA, DM, Hex, MeOH, and water. The results for the control group cultured without extract were demonstrated on the left (gray line). All experimental data were presented as average values ± SD; n = 3; ∗ p < 0.05.
3.3. Effects of EA and DM Extracts on Caspase Proteins in Melanoma Cells
Caspase is a family of cysteine-aspartic proteases that mediates type I programmed cell death (apoptosis), and more than 10 family members have been identified so far [22]. The activation of caspase-associated proteins is essential for apoptosis induced by various apoptotic stimuli. These changes include blebbing, cell shrinkage, nuclear fragmentation, chromatin condensation, and chromosomal DNA fragmentation; however, the failure of cancer cell apoptosis is a major contributor to tumor development and autoimmune disease [23].
Caspase-9 initiates an apoptotic cascade by cleaving and activates caspase-3 [24]. However, the maturation of caspase-9 requires autocatalytic cleavage by apoptosomes released by damaged mitochondria [25]. The cleavage of caspase-3 activates caspase-6 and caspase-7; the protein itself is processed and activated by caspase-8, caspase-9, and caspase-10. The activation of caspase-3 induces cellular apoptosis and proteolysis in specific substrates [26]. Figure 3 showed how caspase affected the apoptotic process induced by EA and DM extracts of E. ramosissimum in the three human melanoma cells. Remarkable alterations caused by molecular proteins associated with apoptosis included increased proteolysis induced by caspase-3 and caspase-9. A low concentration (10 μg/mL) of EA extract induced stimulated enzymes in A375, A375.S2, and A2058 cells. Although DM extract also triggered caspase-3 and caspase-9, its effect was smaller compared to a similar dose of EA extract. Notably, 50 μg/mL DM extract was needed to trigger caspase-3 and caspase-9 in A2058 cells. The cellular proteins caspase-3 and caspase-9 have important roles in the regulation of nuclear DNA damage caused by apoptosis and in the decomposition of organelles. Our experiments demonstrated that EA induced caspase protein changes, and similar results were also shown from DM extract, but lower.
Figure 3.

Expressions of caspase proteins in (a) A375, (b) A375.S2, and (c) A2058 cells after treatment with 10, 50, and 100 μg/mL EA and DM extracts for 24 h. Band darkness indicated relative protein expressions in comparison with GAPDH.
3.4. Mushroom Tyrosinase Inhibition
In mammals, the rate-limiting enzyme tyrosinase is the most important enzyme of pigment biosynthesis reactions. Other enzymes only adjust to differences in synthesis of eumelanin and pheomelanin [10, 27, 28]. To determine whether the extracts inhibited melanin synthesis by suppressing tyrosinase, in vitro tyrosinase activity was tested in mushroom type. In Table 2, it was shown that all extracts except for water had dose-dependent (5–100 μg/mL) inhibiting effects on the mushroom tyrosinase system. At a concentration of 100 μg/mL, the repressive effect of EA extract was slightly lower than that of kojic acid. At this concentration, EA had the strongest inhibiting effect (approximately 40%).
Table 2.
Mushroom tyrosinase inhibition by different concentrations of E. ramosissimum extracts.
| Concentrations (μg/mL) | Mushroom tyrosinase inhibition (%) | ||
|---|---|---|---|
| 5 | 50 | 100 | |
| EA | 22.42 ± 0.25 | 24.40 ± 4.90 | 38.93 ± 3.09 |
| DM | 20.69 ± 4.53 | 21.30 ± 1.10 | 23.79 ± 3.84 |
| Hex | 20.54 ± 0.64 | 22.33 ± 0.69 | 23.82 ± 4.25 |
| MeOH | 18.54 ± 2.70 | 20.45 ± 0.36 | 23.27 ± 1.67 |
| Water | 17.88 ± 3.46 | 19.37 ± 0.12 | 23.82 ± 1.24 |
| Kojic acida | — | — | 30.23 ± 5.68 |
All statistics are presented as average values ± SD; n = 3. aKojic acid (100 μg/mL) was applied as a positive control. Assays not performed in this study were indicated by dashes.
3.5. Melanin Content of B16-F10 Cells
Melanin is the source of skin color and can protect the skin from UV radiation damage which induces DNA mutations. Despite its protective functions, superabundance of melanin negatively affects the skin, which then causes social problems [29, 30]. To demonstrate the skin whitening effects of E. ramosissimum and its inhibiting effects on melanogenesis, melanin alterations were measured after treatments with E. ramosissimum extracts. In Figure 4(a), it was demonstrated that the five fraction extracts did not substantially harm B16 cell viability. In Figure 4(b), we presented the notion that the EA extract had the ability to decrease melanin production by about 32% at 200 µg/mL in a dose-dependent trend from concentrations of 5 to 200 µg/mL. In contrast, 200 μg/mL DM extract substantially augmented melanin production (112%), and the effects of DM extract were dose-dependent. Other extracts, including Hex, MeOH, and water, did not substantially change melanin production, even at the maximum experimental dose of 200 μg/mL. These experimental results suggested that the EA extract had potential applications as a whitening agent in cosmetic products whereas the DM extract could be used as a skin darkening agent.
Figure 4.

(a) Effects of E. ramosissimum extracts on viability of B16-F10 cells according to MTT assay and (b) effects of E. ramosissimum extracts on melanin content quantification of all extracts were processed with 5, 50, 100, and 200 μg/mL, respectively. The results for the control group cultured without extracts were shown on the left (gray line). All experimental results were presented as average values ± SD; n = 3; ∗ p < 0.05.
3.6. Expression of Melanogenesis-Related Proteins in B16-F10 Cells
A well known role of microphthalmia-associated transcription factor (MITF) in melanogenesis is normal synthesis of melanin in melanocytes. Expression of MITF increases melanin production by facilitating biosynthesis of tyrosinase, tyrosinase-related protein 1 (TRP-1), and TRP-2 [31]. Expression of MITF is also significant for stabilizing tyrosinase protein and modulating its catalytic activity [8, 32]. To elucidate the mechanisms of its activity, expressions of melanogenesis-related proteins were analyzed by western blot. Figure 5 presented the notion that EA extract concentrations ranging from 10 to 100 μg/mL reduced expressions of MITF, tyrosinase, TRP-1, and TRP-2 in a dose-dependent manner. Conversely, DM extract promoted expressions of MITF, tyrosinase, TRP-1, and TRP-2 in B16-F10 cells in a dose-dependent manner. Both of these phenomena were consistent with the data for melanin content shown in Figure 4(b).
Figure 5.

Expressions of tyrosinase, TRP-1, TRP-2, and MITF after treatment with E. ramosissimum extracts. B16-F10 cells were treated with EA and DM extracts at concentrations of 10, 50, and 100 μg/mL for 24 h. Protein expressions were shown in comparison with GAPDH.
4. Conclusion
In summary, the experiments in this study demonstrated that the most beneficial of the five fraction extracts of E. ramosissimum was EA because of its multiple biofunctional properties (Figure 6). The experimental outcomes showed that, by acting as an antioxidant ingredient and electron donor, EA extract discontinued or terminated free radical chain reactions. In human melanoma, EA and DM extracts affected the viabilities of melanoma cells and showed low toxicity in both normal human cells, HaCaT cells and fibroblasts. To understand the mechanisms of cell death, we performed western blot analyses of protein expressions in melanoma cells, which pointed out that both extracts induced caspase-3 and caspase-9, both of which have vital roles in apoptosis. Research evaluations of the potential use of EA extract as a whitening agent illustrated that it inhibited mushroom tyrosinase activity and the synthesis of melanin; in contrast, DM extract increased the quantity of melanin. Western blot analyses showed that EA and DM extracts decreased and increased melanin content, respectively, by regulating MITF, tyrosinase, Trp-1, and Trp-2. Whereas this study established the biological functions of E. ramosissimum, our future studies will further investigate the components and mechanisms of these compounds.
Figure 6.
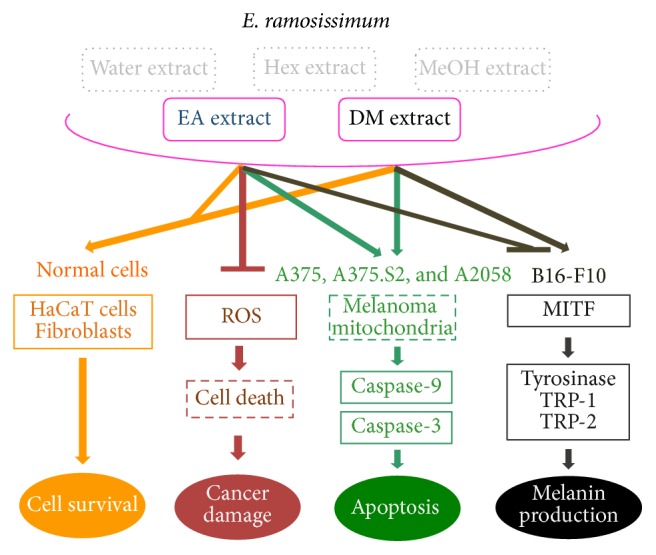
Schematic diagram of biofunctions of E. ramosissimum extracts in human skin cells, including normal cell survival, apoptotic pathways of melanoma, and melanogenesis.
Acknowledgments
This work was supported by grants from the Ministry of Science and Technology, Taiwan (MOST104-2622-E-037-001; MOST104-2622-E-037-003-CC2; MOST104-2221-E-037-005-MY2; and MOST104-2628-E-037-001-MY3), from the Center for Stem Cell Research, Kaohsiung Medical University, Kaohsiung, Taiwan (KMU-TP104G00 and KMUTP104G02-05), from Kaohsiung Medical University, Taiwan (KMU-DK105005), from Kaohsiung Medical University, NSYSU-KMU Joint Research Project, Taiwan (NSYSUKMU105-P 007), and from the Center for Infectious Disease and Cancer Research, Kaohsiung Medical University, Taiwan (KMU-TP104E18).
Competing Interests
The authors have no competing interests regarding the publication of this study.
Authors' Contributions
Pin-Hui Li and Yu-Pin Chiu equally contributed to this study.
References
- 1.Apel K., Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology. 2004;55:373–399. doi: 10.1146/annurev.arplant.55.031903.141701. [DOI] [PubMed] [Google Scholar]
- 2.Gutiérrez-Uzquiza Á., Arechederra M., Bragado P., Aguirre-Ghiso J. A., Porras A. p38α mediates cell survival in response to oxidative stress via induction of antioxidant genes: effect on the p70S6K pathway. Journal of Biological Chemistry. 2012;287(4):2632–2642. doi: 10.1074/jbc.m111.323709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Richardson C., Yan S., Vestal C. G. Oxidative stress, bone marrow failure, and genome instability in hematopoietic stem cells. International Journal of Molecular Sciences. 2015;16(2):2366–2385. doi: 10.3390/ijms16022366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kim K.-N., Ahn G., Heo S.-J., et al. Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environmental Toxicology and Pharmacology. 2013;35(1):39–46. doi: 10.1016/j.etap.2012.10.002. [DOI] [PubMed] [Google Scholar]
- 5.Ascenso A., Pedrosa T., Pinho S., et al. The effect of lycopene preexposure on UV-B-irradiated human keratinocytes. Oxidative Medicine and Cellular Longevity. 2016;2016:15. doi: 10.1155/2016/8214631.8214631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lee C.-C., Chiou L.-Y., Wang J.-Y., et al. Functional ginger extracts from supercritical fluid carbon dioxide extraction via in vitro and in vivo assays: antioxidation, antimicroorganism, and mice xenografts models. The Scientific World Journal. 2013;2013:8. doi: 10.1155/2013/210845.210845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ghosh R., Guha D., Bhowmik S., Karmakar S. Antioxidant enzymes and the mechanism of the bystander effect induced by ultraviolet C irradiation of A375 human melanoma cells. Mutation Research. 2013;757(1):83–90. doi: 10.1016/j.mrgentox.2013.06.022. [DOI] [PubMed] [Google Scholar]
- 8.Liu W.-S., Kuan Y.-D., Chiu K.-H., et al. The extract of Rhodobacter sphaeroides inhibits melanogenesis through the MEK/ERK signaling pathway. Marine Drugs. 2013;11(6):1899–1908. doi: 10.3390/md11061899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lee C.-C., Chen Y.-T., Chiu C.-C., Liao W.-T., Liu Y.-C., David Wang H.-M. Polygonum cuspidatum extracts as bioactive antioxidaion, anti-tyrosinase, immune stimulation and anticancer agents. Journal of Bioscience and Bioengineering. 2015;119(4):464–469. doi: 10.1016/j.jbiosc.2014.09.008. [DOI] [PubMed] [Google Scholar]
- 10.Lee M.-G., Kuo S.-Y., Yen S.-Y., et al. Evaluation of Cinnamomum osmophloeum Kanehira extracts on tyrosinase suppressor, wound repair promoter, and antioxidant. The Scientific World Journal. 2015;2015:7. doi: 10.1155/2015/303415.303415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Oh H., Kim D.-H., Cho J.-H., Kim Y.-C. Hepatoprotective and free radical scavenging activities of phenolic petrosins and flavonoids isolated from Equisetum arvense. Journal of Ethnopharmacology. 2004;95(2-3):421–424. doi: 10.1016/j.jep.2004.08.015. [DOI] [PubMed] [Google Scholar]
- 12.Paulsamy S., Moorthy D., Nandakumar K., et al. Evaluation of in vitro antioxidant potential of methanolic extracts of the ferns, Actiniopteris radiata (Sw) Link and Equisetum ramosissimum Desf. International Journal of Research and Development in Pharmacy & Life Sciences. 2013;2(3):451–455. [Google Scholar]
- 13.Wang H.-M., Chou Y.-T., Hong Z.-L., et al. Bioconstituents from stems of Synsepalum dulcificum Daniell (Sapotaceae) inhibit human melanoma proliferation, reduce mushroom tyrosinase activity and have antioxidant properties. Journal of the Taiwan Institute of Chemical Engineers. 2011;42(2):204–211. doi: 10.1016/j.jtice.2010.05.008. [DOI] [Google Scholar]
- 14.Wang H.-M., Pan J.-L., Chen C.-Y., et al. Identification of anti-lung cancer extract from Chlorella vulgaris C-C by antioxidant property using supercritical carbon dioxide extraction. Process Biochemistry. 2010;45(12):1865–1872. doi: 10.1016/j.procbio.2010.05.023. [DOI] [Google Scholar]
- 15.Wang H.-M., Chen C.-Y., Wen Z.-H. Identifying melanogenesis inhibitors from Cinnamomum subavenium with in vitro and in vivo screening systems by targeting the human tyrosinase. Experimental Dermatology. 2011;20(3):242–248. doi: 10.1111/j.1600-0625.2010.01161.x. [DOI] [PubMed] [Google Scholar]
- 16.Lin C.-L., Chen R.-F., Chen J. Y.-F., et al. Protective effect of caffeic acid on paclitaxel induced anti-proliferation and apoptosis of lung cancer cells involves NF-κb pathway. International Journal of Molecular Sciences. 2012;13(5):6236–6245. doi: 10.3390/ijms13056236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen X., Zhang B., Yuan X., et al. Isoliquiritigenin-induced differentiation in mouse melanoma B16F0 cell line. Oxidative Medicine and Cellular Longevity. 2012;2012:11. doi: 10.1155/2012/534934.534934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pan X. H., Zhang X. L., Sun H. L., Zhang J., Yan M., Zhang H. Autophagy inhibition promotes 5-fluorouraci-induced apoptosis by stimulating ROS formation in human non-small cell lung cancer A549 cells. PLoS ONE. 2013;8(2) doi: 10.1371/journal.pone.0056679.e56679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Godic A., Poljšak B., Adamic M., Dahmane R. The role of antioxidants in skin cancer prevention and treatment. Oxidative Medicine and Cellular Longevity. 2014;2014:6. doi: 10.1155/2014/860479.860479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ma D.-L., Liu L.-J., Leung K.-H., et al. Antagonizing STAT3 dimerization with a rhodium(III) complex. Angewandte Chemie—International Edition. 2014;53(35):9178–9182. doi: 10.1002/anie.201404686. [DOI] [PubMed] [Google Scholar]
- 21.Liu M., Douthwaite S. Resistance to the macrolide antibiotic tylosin is conferred by single methylations at 23S rRNA nucleotides G748 and A2058 acting in synergy. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(23):14658–14663. doi: 10.1073/pnas.232580599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nakagawa T., Zhu H., Morishima N., et al. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-β . Nature. 2000;403(6765):98–103. doi: 10.1038/47513. [DOI] [PubMed] [Google Scholar]
- 23.Harrington H. A., Ho K. L., Ghosh S., Tung K. C. Construction and analysis of a modular model of caspase activation in apoptosis. Theoretical Biology and Medical Modelling. 2008;5, article 26 doi: 10.1186/1742-4682-5-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wu P.-F., Chiu C.-C., Chen C.-Y., Wang H.-M. D. 7-Hydroxydehydronuciferine induces human melanoma death via triggering autophagy and apoptosis. Experimental Dermatology. 2015;24(12):930–935. doi: 10.1111/exd.12805. [DOI] [PubMed] [Google Scholar]
- 25.Pang Y., Bai X. C., Yan C., et al. Structure of the apoptosome: mechanistic insights into activation of an initiator caspase from Drosophila . Genes & Development. 2015;29(3):277–287. doi: 10.1101/gad.255877.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lo S.-C., Wang Y., Weber M., Larson J. L., Scearce-Levie K., Sheng M. Caspase-3 deficiency results in disrupted synaptic homeostasis and impaired attention control. The Journal of Neuroscience. 2015;35(5):2118–2132. doi: 10.1523/jneurosci.3280-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gruber J. V., Holtz R. Examining the impact of skin lighteners in vitro. Oxidative Medicine and Cellular Longevity. 2013;2013:7. doi: 10.1155/2013/702120.702120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Neagu E., Paun G., Albu C., Radu G.-L. Assessment of acetylcholinesterase and tyrosinase inhibitory and antioxidant activity of Alchemilla vulgaris and Filipendula ulmaria extracts. Journal of the Taiwan Institute of Chemical Engineers. 2015;52:1–6. doi: 10.1016/j.jtice.2015.01.026. [DOI] [Google Scholar]
- 29.Brenner M., Hearing V. J. The protective role of melanin against UV damage in human skin. Photochemistry and Photobiology. 2008;84(3):539–549. doi: 10.1111/j.1751-1097.2007.00226.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chen C.-Y., Lin L.-C., Yang W.-F., Bordon J., Wang H.-M. D. An updated organic classification of tyrosinase inhibitors on melanin biosynthesis. Current Organic Chemistry. 2015;19(1):4–18. doi: 10.2174/1385272819666141107224806. [DOI] [Google Scholar]
- 31.Hoek K. S., Schlegel N. C., Eichhoff O. M., et al. Novel MITF targets identified using a two-step DNA microarray strategy. Pigment Cell and Melanoma Research. 2008;21(6):665–676. doi: 10.1111/j.1755-148X.2008.00505.x. [DOI] [PubMed] [Google Scholar]
- 32.Lee J.-Y. Down-regulation of MITF, TRP-1, TRP-2, and tyrosinase expressions by compounds isolated from Pruni persicae Flos in murine B16F10 melanoma. Journal of the Korean Society for Applied Biological Chemistry. 2014;57(6):827–834. doi: 10.1007/s13765-014-4288-5. [DOI] [Google Scholar]