

Abstract
Using functional magnetic resonance imaging (fMRI), we examined the process of language specialization in the brain by comparing developmental changes in two contrastive orthographies: Chinese and English. In a visual word rhyming judgment task, we found a significant interaction between age and language in left inferior parietal lobule and left superior temporal gyrus, which was due to greater developmental increases in English than in Chinese. Moreover, we found that higher skill only in English children was correlated with greater activation in left inferior parietal lobule. These findings suggest that the regions associated with phonological processing are essential in English reading development. We also found greater developmental increases in English than in Chinese in left inferior temporal gyrus, suggesting refinement of this region for fine-grained word form recognition. In contrast, greater developmental increases in Chinese than in English were found in right middle occipital gyrus, suggesting the importance of holistic visual-orthographic analysis in Chinese reading acquisition. Our results suggest that the brain adapts to the special features of the orthography by engaging relevant brain regions to a greater degree over development.
Keywords: English, Chinese, fMRI, reading development
Introduction
There is extensive evidence that there are dynamic changes in the functional organization of a system that are driven by an adaptation to acquisition of a new skill. Because reading is a uniquely human task, many studies have investigated how neuronal activation is modulated by learning to read. More recently, some have investigated how the nature of the writing system influences the process of learning to read. However, it is not known how differences in structure of writing systems may differentially reorganize the brain networks of reading over development. We wished to test the basic hypothesis that learning to read leads to a growing divergence between different orthographies with increased experience.
Orthographies, such as English, follow the alphabetic principle with semi-regular mapping between graphemes (letters) and phonemes (sounds), thus strongly engaging phoneme-level phonological processing (Goswami, 1993; Nation & Hulme, 1997; Wagner & Torgeson, 1987). Phonemic awareness has been found to be associated with reading performance and to be the best predictor of later reading achievement in English (Anthony, Lonigan, Driscoll, Phillips & Burgess, 2003; Bradley & Bryant, 1983; Muter, Hulme, Snowling & Taylor, 1997). Conversely, non-alphabetic orthographies, such as Chinese, have phonetic radicals that offer some cues to syllabic level pronunciation, yet, do not have systematic mappings from character to syllable. Only 23% of characters can be pronounced correctly following the pronunciation of the phonetic radical (Xing, 2004). As a result, non-alphabetic orthographies may not engage phoneme level phonological processes to the same degree as English (Zhou, 1997). Moreover, phonemic awareness does not appear to be a consistent predictor of reading performance across studies (McBride-Chang, Cho, Liu, Wagner, Shu, Zhou, Cheuk & Muse, 2005a; Shu, Peng & McBride-Chang, 2008). However, there is evidence that sub-lexical processing of the phonetic radical and syllabic level phonological awareness is related to reading acquisition in Chinese (Shu, 2000; Shu et al., 2008). Neuroimaging studies also suggest that the phonological processing regions, such as the left inferior frontal gyrus, and left superior temporal gyrus are heavily involved in English word reading and become more so with skill and age (Turkeltaub, Garaeu, Flowers, Zefirro & Eden, 2003; Dehaene, 2010; Booth, Burman, Meyer, Gitelman, Parrish & Mesulam, 2003a; Booth, Burman, Meyer, Zhang, Gitelman, Parrish & Mesulam, 2004). However, in comparison to English, there is reduced activation in left inferior frontal gyrus and left superior temporal gyrus in Chinese word reading in adults (Bolger, Perfetti &Schneider, 2005; Tan, Spinks, Feng, Siok, Perfetti, Xiong, Fox, Gao & Kalogirou, 2003). One developmental study has also found that there is a developmental decrease in the activation of left superior temporal gyrus in Chinese reading (Cao, Lee, Shu, Yang, Xu, Li & Booth, 2010). This may indicate that the phoneme-level representation plays a less essential role in Chinese reading. In comparison to English, there is also increased brain activation in left middle frontal gyrus in Chinese adult reading (Bolger et al., 2005; Tan, Laird, Karl & Fox, 2005), which may be associated with syllabic level and/or relatively arbitrary mapping between orthography and phonology. This region has also been found to show developmental increase in Chinese reading (Cao, Lee, Shu, Yang, Xu, Li & Booth, 2010). Altogether, the research suggests that English reading development relies heavily on phoneme-level phonological processing, while Chinese reading involves a larger grain size at the syllable level, but that mapping tends to be arbitrary.
Logographic orthographies such as Chinese also differ from alphabetic orthographies in the structural characteristics of the script. English is a left-right linear layout of letters, whereas Chinese character forms are complex visual-spatial configurations in a two-dimensional square. Behavioral studies suggest that visual-orthographic skills constitute a more important predictor in Chinese reading acquisition than phonological skills (Ho, Chan, Chung, Lee & Tsang, 2007; McBride-Chang, Chow, Zhong, Burgess & Hayward, 2005b). Orthographic skills have also been shown to be important in English reading acquisition especially for reading speed and fluency (Frith, 1986; O’Brien, Wolf, Miller, Lovett & Morris, 2011). In terms of neural correlates, neuroimaging studies have suggested differential engagement of visual processing regions across writing systems. Even though some studies suggest that Chinese and English orthographic processing equally engage the left occipito-temporal cortex (Sakurai, Momose, Iwata, Sudo, Ohtomo & Kanazawa, 2000; Liu, Dunlap, Fiez & Perfetti, 2007), some studies suggest that English orthographic processing shows a greater engagement of this region (Tan et al., 2005; Kim, Byun, Lee, Gaillard, Xu & Theodor, 2011). The interactive account argues that the ventral occipito-temporal cortex receives top-down predictions from higher-level linguistic representation regions as well as bottom-up basic visual inputs (Olulade, Flowers, Napoliello & Eden, 2013; Price & Devlin, 2011). The semi-regular mapping between graphemes and phonemes in English may cause greater top-down prediction from associated phonology especially at the phoneme level in comparison to the more arbitrary mapping between orthography and phonology in Chinese. Therefore the left occipito-temporal cortex may be more involved in English than in Chinese reading. A number of studies have demonstrated that Chinese shows greater involvement of similar regions in the right hemisphere (Bolger et al., 2005; Liu & Perfetti, 2003; Tan et al., 2005). Developmental studies have also found a developmental increase in the involvement of the left occipito-temporal cortex in English reading (Booth, Burman, Meyer, Zhang, Choy, Gitelman, Parrish & Mesulam, 2003b; Shaywitz, Skudlarski, Holahan, Marchione, Constable, Fulbright, Zelterman, Lacadie & Shaywitz, 2007) and a developmental increase in the involvement of the right middle occipital gyrus in Chinese reading (Cao et al., 2010; Cao, Peng, Liu, Jin, Fan, Deng & Booth, 2009). Altogether, studies suggest that orthographic processing is important in both English and Chinese reading, however, due to the nature of the visual forms and the mapping between orthography and phonology, English orthographic processing appears to show greater involvement of the left occipito-temporal cortex, whereas Chinese visual word processing appears to show greater engagement of the right middle occipital gyrus (Tan et al., 2005). This differential engagement may be due to the demands on holistic visuo-spatial processing of Chinese characters, and the detailed processing of individual letters in English.
The purpose of the current study was to investigate how the nature of the writing system influences the development of the reading network. We wished to examine whether reading acquisition, particularly, the process of mapping orthography to phonology was marked by the growing divergence between alphabetic orthographies (i.e. English) versus non-alphabetic orthographies (i.e. Chinese) in brain regions involved in orthographic and phonological processing.
Methods
Participants
The current study included one hundred and three participants (n=103). Participants included Chinese-speaking adults (n=20; ages 19 – 28 years; mean age = 21.5), Chinese-speaking children (n=30; ages 8.5 – 12.5 years; mean age = 10.8), English-speaking adults (n=24; ages 18 – 34 years; mean age = 21.0), and English-speaking children (n=29; ages 7.6–13.8; mean age = 10.4). The Chinese and English groups were controlled for age and gender (see Table 1). All participants including children performed above chance level on the visual rhyming task with their accuracy above 60%. Most participants (83 out of 103) were also tested on an auditory rhyming judgment task as a measure of their phonological ability (see Table 1). Chinese-speaking adults were undergraduate and graduate students from Beijing Normal University, and Chinese-speaking children were from a public elementary school in Hebei province. Chinese children and adults all learned English as a second language. English-speaking adults and children were from Chicago metropolitan area. All participants met the following criteria: (1) native speakers of their language (English or Mandarin Chinese), (2) right-handed, (3) free of neurological disease or psychiatric disorders, (4) no attention deficit hyperactivity disorder, and (5) no learning disability. The Institutional Review Board at Northwestern University and Beijing Normal University approved the informed consent procedures.
Table 1.
Sample size (N) as well as means and standard deviations for each group for age, accuracy and reaction time on the visual rhyming task in the scanner, and accuracy on an auditory rhyming outside of the scanner.
| English | N | Age | Visual rhyming accuracy | Visual rhyming reaction time | Auditory rhyming accuracy | |
|---|---|---|---|---|---|---|
| Children | 29 | 10.4 (1.3) | 84% (10%) | 1391 (252) | 80% (11%) | |
| Adults | 24 | 21.0 (3.5) | 95% (4%) | 984 (291) | 85% (6%) | |
| Chinese | ||||||
| Children | 30 | 10.8 (1.4) | 77% (10%) | 1668 (271) | 73% (15%) | |
| Adults | 20 | 21.5 (2.2) | 94% (3%) | 1228 (324) | 90% (9%) |
Tasks
During scanning, participants performed a rhyming judgment task to sequentially presented visual word pairs interspersed with perceptual control and fixation baseline trials. Pairs of word stimuli either rhymed or did not rhyme. Participants were asked to respond as quickly and as accurately as possible, using their right index finger for a yes (rhyme) response and their right middle finger for a no (non-rhyme) response. The presentation of all words was 800 ms, with a 200 ms interval between the two words. A red fixation cross appeared on the screen right after the offset of the second word, indicating the need to make a response. The response interval duration was variable (2200, 2600, or 2800 ms), such that each trial lasted for either 4000, 4400, or 4800 ms. Fixation baseline trials (48) required the participant to press the “yes” button when a fixation cross at the center of the screen turned from black to red. Perceptual trials (24) were also included as part of a larger study, but were not of interest in the present experiment. These trials required participants to determine whether two sequentially presented symbol patterns matched or mismatched by pressing the “yes” or “no” button. The timing for the perceptual control and fixation baseline trials was the same as for the lexical trials. The order of presentation of lexical trials, perceptual trials and fixation trials was optimized for event-related designs by OptSeq ((http://surfer.nmr.mgh.harvard.edu/optseq/).
In English, all words were monosyllabic without homophones. There were 24 trials in each of 4 conditions, and included two non-conflicting conditions, such that the words in each pair had similar orthographic and phonological endings (O+P+: e.g., gate–hate), or different orthographic and phonological endings (O−P−: e.g., press–list), and two conflicting conditions, such that words had similar orthographic but different phonological endings (O+P−, e.g., pint–mint), or different orthographic but similar phonological endings (O−P+, e.g., jazz–has). All words were matched across conditions for written word frequency in children (Zeno, 1996) and the sum of their written bigram frequency (English Lexicon Project, http://elexicon.wustl.edu).
In Chinese, all words consisted of two characters without homophones at the word level. Two-character words were used in the task in order to be consistent with the auditory rhyming task outside of the scanner. As with the English stimuli, there were 24 trials in each of 4 conditions, two non-conflicting and two conflicting. Similar orthography was defined as sharing a phonetic radical for the second character of the word. The two non-conflicting conditions included one with similar orthography and phonology (O+P+, e.g. 弥瞛 /bu3/, 纯朴 /pu3/) and one with different orthography and phonology (O−P−, e.g. 损坏 /huai4/, 学科 /ke1/). The two conflicting conditions included one with similar orthography and different phonology (O+P−, e.g. 翻琯 /yi4/, 濷择 /ze2/) and another with different orthography and similar phonology (O−P+, e.g. 环保 /bao3/, 大炮/ pao4/). Rhyming in the Chinese task was defined as the second character of the first and second word sharing the same rhyme. Participants were told to ignore the tone, however, in order to avoid the possible influence of tone on the rhyming judgment, we balanced the number of trials with the same tone and different tone. In half of the trials of the 4 lexical conditions (rhyming and non-rhyming), the second character of the first and the second word had the same tone (e.g. 弥瞛/bu3/, 纯朴/pu3/), and in the other half they had different tones (e.g. 濒捕/bu3/, 缈绑/pu2/). The two-character words were chosen from the Chinese textbooks used in grade 1 to grade 3 and the youngest Chinese children in the current study were in grade 3. The Chinese words were matched on several variables across conditions including adult written frequency (Beijing Language and Culture University, 1990), number of strokes, word familiarity in third-graders, and word familiarity in fifth-graders. In an independent study on 100 children who were the same age as our fMRI child participants, we made sure that all characters used in the study were known by using a word familiarity measure on a 7-point scale (Mean=5.77, range = 3.07–6.94). The second characters of words were also matched on adult written frequency (Beijing Language and Culture University, 1990) and number of strokes.
MRI data acquisition
Children were given a practice session in a scanner simulator. Different stimuli were used in the practice and scanning sessions. Participants lay in the scanner with their head position secured with foam padding. An optical response box was placed in the participant’s dominant right hand and a compression alarm ball placed in the left hand. The head coil was positioned over the participant’s head so that they could effectively use the mirror to view the projection screen at the rear of the scanner. Images for the Chinese-speaking participants were acquired using a 3.0 Tesla Siemens scanner (Siemens Healthcare, Erlangen, Germany) at Beijing Normal University and images for the English-speaking participants were acquired using an identical scanner at Northwestern University. Identical scanning protocols were used at the two locations. A number of studies have suggested that variance of sites is small and the reproducibility is similar between and within sites (Sutton, Goh, Hebrank, Welsh, Chee & Park, 2008; Gountouna et al., 2010). In the current study, we were interested in the interaction between age and language, therefore any site variation was less likely to play a role in our findings. In order to insure site variation was a minor issue, in a separate model, we included “site” as a covariate when examining the interaction between age and language. Gradient echo localizer images were acquired to determine the placement of the functional slices. For the functional images, a susceptibility weighted single-shot EPI (echo planar imaging) method with BOLD (blood oxygenation level-dependent) was used with the following scan parameters: TE = 20 ms, flip angle = 80 °, matrix size = 120 × 128, field of view = 220 × 206.3 mm, slice thickness = 3 mm (0.48 mm gap), number of slices = 32, TR = 2,000 ms. These parameters resulted in a 1.7 × 1.7 × 3 mm voxel size. 168 whole-brain volumes were acquired each run using an interleaved bottom to top sequence, with one complete volume collected every 2 seconds (TR = 2000 ms). A high resolution, T1 weighted 3D image was also acquired with the following parameters: TR = 2300 ms, TE = 3.36 ms, flip angle = 9 ° matrix size = 256 × 256, field of view = 256 mm, slice thickness = 1 mm, number of slices = 160. The acquisition of the anatomical scan took approximately 9 minutes.
Image analysis
Data analysis was performed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm). The following steps were used for data preprocessing. (1) Slice timing correction for interleaved acquisition using sinc interpolation. (2) 4th degree b-splice interpolation for realignment to the first volume. (3) Trilinear coregistration with the anatomical image. (4) 16 nonlinear iterations for normalization and voxels were resliced to 2 × 2 × 2 mm. (5) 4 × 4 × 8 mm full width half maximum Gaussian kernal smoothing. Up to two volumes where movement exceeded 4 mm in any of the x, y or z dimensions were replaced with the mean of the images immediately before and after the outlying volume. Participants with >2 volumes with >4 mm of movement were excluded from the study. We normalized all brains to the standard T1 Montreal Neurological Institute (MNI) adult template (12 linear affine parameters for brain size and position, 8 non-linear iterations and 2×2×2 non-linear basis functions). We normalized the children’s brain to an adults template because the examination of developmental changes required a comparison using the same template, and also because one previous study has found that normalization accuracy of typical children’s brain is comparable using the MNI template or the CCHMC template (Cincinnati Children’s Hospital Medical Center, (Wilke, Schmithorst & Holland, 2002) which was based on MRI data of 200 children from 5–19 years of age (Machilsen, d’Agostino, Maes, Vandermeulen, Hahn, Lagae & Stiers, 2007).
Statistical analyses at the first level were calculated using an event-related design with four lexical trial types, the perceptual control trials, and the fixation baseline trials as six conditions of interest. A high pass filter with a cutoff period of 128 s was applied. Trials were modeled using a canonical hemodynamic response function (HRF). Data from each subject were entered into a general linear model using an event-related analysis procedure. Group results were obtained using random-effects analyses by combining subject-specific summary statistics across the group as implemented in SPM8. The main effect of all lexical conditions versus the baseline fixation condition was tested using a one sample t-test separately for each language (Chinese, English). We formed a union of these two maps that served as a mask for all subsequent analyses to ensure that developmental effects were due to activation and not deactivation differences, because deactivation may be related with the function of default mode network that is not the main interest in the current study. To determine whether the developmental effects depended on language, an ANCOVA of age by language with accuracy on the visual rhyming task as a covariate was calculated for each lexical condition (O+P+,O+P−,O−P+,O−P−) minus baseline and for the all lexical conditions (lexical) minus baseline. The main effects of age and language were also determined based on the contrast of lexical minus baseline. Accuracy served as a covariate so that the differences of brain activation were less likely due to performance confounds. All reported brain activations survive at P<.001 uncorrected, voxels>15 which equals FWE corrected P <.05 in our data (i.e. using our voxel size, and spatial smoothing FWHM etc.) according to AlphaSim (Cox, 1996). In addition to the voxel-wise analyses, we adopted nine regions of interest (ROI) from the main effect of language. These ROIs were defined as 6 mm radius spheres centered at the peak voxels in the language effect and eigenvalues for each participant were extracted for the ROIs. We then calculated age by language ANCOVAs on these ROIs with accuracy on the visual rhyming task as a covariate.
In order to examine the relationship between task performance and brain activation, we examined all regions that showed a significant interaction either in the voxel-wise or the ROI analysis. In the brain-behavior correlation analyses, we focused on children because of the ceiling effect for accuracy in adults. Eigenvalues for each child participant were extracted for a sphere with a 6 mm radius and we correlated this with accuracy on the visual rhyming task within each child group with the phonological ability (i.e. accuracy on the auditory rhyming task) partialled out. There were 22 Chinese children and 19 English children in the partial correlation analyses, because some children did not participate in the auditory rhyming task. Phonological ability (i.e. auditory rhyming) was partialled out because it is generally higher for English speakers than Chinese speakers due to the nature of the language. There was also evidence that it is more correlated with reading in English than in Chinese (McBride-Chang, Cho, Liu, Wagner, Shu, Zhou, Cheuk & Muse, 2005a). In order to make references to reading and reading development using the visual rhyming judgment task in the current study, we wanted to rule out the influence of phonological ability.
Results
Behavioral results
ANCOVAs of language (Chinese, English) with age as a covariate were calculated for performance on the visual rhyming task in the scanner separately for accuracy and reaction time (see Table 1). There was a significant main effect of language for accuracy (F(1,99) = 8.242, p < 0.01) with English participants achieving higher accuracies than Chinese participants. The main effect of language was not significant for reaction time (F(1,99) = 22.290, p = 0.133). There was a significant effect of age for accuracy (F(1,99) = 77.039, p < 0.001) and reaction time (F(1,99) = 52.704, p < 0.001) with higher accuracies and faster reaction times in older participants than younger ones. The interaction of language by age was significant for accuracy (F(1,99) = 4.119, p < 0.05) but not reaction time (F(1,99) = 0.022, p =.881) (see Table 1). The interaction was driven by a greater developmental increase in accuracy in Chinese (F(1,48) = 55.861, p <.001) than in English (F(1,51) = 23.815, p <.001).
Most of the participants were also given an auditory rhyming task outside of the scanner as a measure of their phonological ability. An ANCOVA of language revealed a significant effect of age (F(1,79)=15.193, p<.001) driven by higher accuracy in older participants than in younger ones. The main effect of language was significant (F(1,79)=6.033, p<.05) with higher accuracies in English participants than in Chinese participants. There was also a significant interaction of age by language (F(1,79)=5.887, p<.05) driven by greater age-related increase in Chinese (F(1,38)=13.822, p<.005) than in English (F(1,41)=1.718, p=.197). Phonological ability was also correlated with the visual rhyming task in the scanner (r=.578, p<.001).
fMRI results
We submitted the lexical minus fixation baseline contrast to an ANCOVA of language with age and accuracy on the visual rhyming task as covariates. Developmental differences within each language are presented in Table 2 and Table 3, and Figure 1 shows the brain activation maps for adults minus children and children minus adults separately for English and Chinese. There were developmental increases in English in inferior parietal lobule (IPL), middle occipital gyrus (MOG) and inferior frontal gyrus (IFG) in the left hemisphere and developmental decreases in cingulate gyrus. There were developmental increases in Chinese in right MOG, left superior parietal lobule and left middle frontal gyrus and developmental decreases in right superior temporal gyrus (STG). Because we used fixation baseline in the contrasts, the results may have reflected all levels of cognitive processing in reading such as visuo-orthographic recognition, mapping from orthography to phonology, phonological manipulation and activation of meaning.
Table 2.
Brain activations for developmental changes in English on the contrast of lexical minus fixation.
| Anatomical Region | H | BA | Voxels | Z-score | x | y | z | |
|---|---|---|---|---|---|---|---|---|
| developmental increase | Inferior parietal lobule | Left | 40 | 42 | 3.99 | −30 | −50 | 44 |
| Superior parietal lobule, inferior parietal lobule | Left | 7, 40 | 40 | 4.53 | −28 | −64 | 44 | |
| Inferior parietal lobule | Left | 40 | 28 | 3.84 | −48 | −44 | 42 | |
| Inferior frontal gyrus | Left | 44,6 | 20 | 3.75 | −40 | 2 | 20 | |
| Middle occipital gyrus, inferior temporal gyrus | Left | 19 | 18 | 3.58 | −30 | −78 | 28 | |
| developmental decrease | Cingulate gyrus | Left | – | 90 | 4.56 | −6 | 10 | 20 |
Note: H, hemisphere; BA, Brodmann Area
Table 3.
Brain activations for developmental changes in Chinese on the contrast of lexical minus fixation.
| Anatomical Region | H | BA | Voxels | Z-score | x | y | z | |
|---|---|---|---|---|---|---|---|---|
| developmental increase | Middle occipital gyrus | Right | 18 | 96 | 5.17 | 24 | −92 | 2 |
| Middle frontal gyrus | Left | 9 | 25 | 3.97 | −48 | 8 | 36 | |
| Cuneus | Left | 18 | 69 | 4.36 | −26 | −86 | 6 | |
| Superior parietal lobule | Left | 7,39 | 90 | 4.12 | −30 | −78 | 30 | |
| 67 | 3.89 | 36 | −80 | −6 | ||||
| developmental decrease | Superior temporal gyrus | Right | 22 | 84 | 4.73 | 50 | −52 | 20 |
Note: H, hemisphere; BA, Brodmann Area
Figure 1.
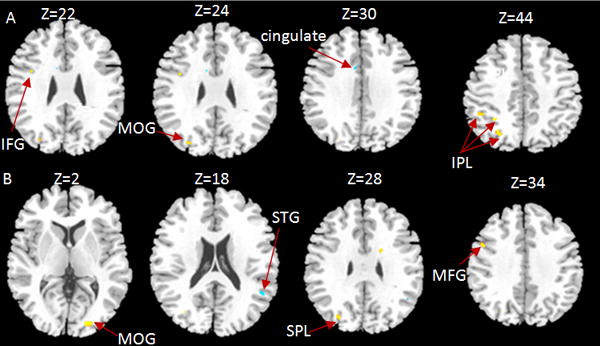
Developmental changes in the rhyming task. Age-related increases are indicated in warm colors and age-related decreases in cold colors. (A) Developmental increases in English in left inferior frontal gyrus (IFG), left middle occipital gyrus (MOG) and left inferior parietal lobule (IPL). Developmental decreases in English in cingulate gyrus. (B) Developmental increases in Chinese in right middle occipital gyrus (MOG), left superior parietal lobule (SPL), and left middle frontal gyrus (MFG). Developmental decrease in Chinese in right superior temporal gyrus (STG).
We also found significant interactions between age and language at two clusters within left IPL (IPL1: x= −28, y=−64, z= 44; voxel=28; IPL2: x=−30, y=−50, z=44; voxel=25; see Figure 2A). This interaction overlapped with the developmental increases at left IPL in English (see Figure 2B). Therefore, the interaction at these two IPL regions was due to greater developmental increases in English compared to Chinese (see Table 2 & Figure 2C). For the model with site as a covariate, we found almost identical clusters within the left IPL for the interaction between age and language (one cluster at x=−28,y=−64, z=44, and one cluster at x=−32, y=−50, z=48), suggesting that site variation is minor in the current study. For the calculation on each lexical condition, we found interaction at left IPL (x= −26, y=−34, z= 30; voxel=62) and left IFG (x= −40, y=2, z= 20; voxel=16) in the O+P− minus baseline and at the left IPL (IPL1: x= −28, y=−64, z= 44; voxel=27; IPL2: x=−48, y=−40, z=50; voxel=17) in the O−P+ condition; we found no significant interaction in the two non-conflicting conditions (O+P+ minus baseline and O−P− minus baseline). Further data analysis revealed that the interactions in O+P− and O−P+ were both driven by greater developmental increase in English than in Chinese.
Figure 2.
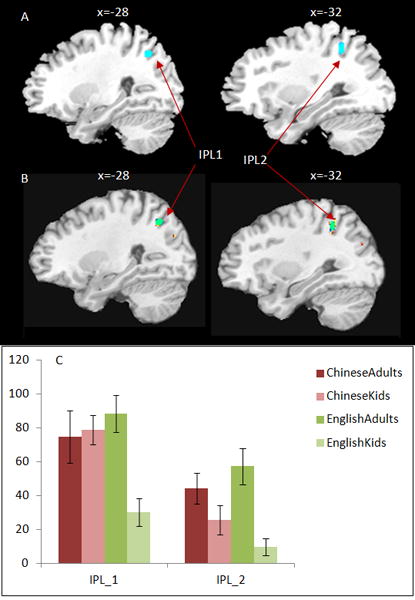
Whole brain analysis showing interactions between language and age. (A) This interaction occurred at two locations in the inferior parietal lobule (IPL1: x= −28, y=−64, z= 44; voxel=28; IPL2: x=−30, y=−50, z=44; voxel=25). (B) This interaction (Green) overlapped with the developmental increases in English (yellow). (C) Bar charts show that the interaction in both clusters in IPL was driven by developmental increases in English. The error bars are standard errors.
Nine ROIs were identified according to the main effect of language (see Table 4) including 5 regions that were more activated in English than in Chinese: left inferior temporal gyrus (ITG: −44, −60, −10), left superior temporal gyrus (−56, −40, 8), left inferior frontal gyrus (−46, 10, 24), left inferior parietal lobule (−44, −38, 46), and left medial frontal gyrus at (−4, 12, 52) and 4 regions that are more activated in Chinese than in English: left superior parietal lobule (−36, −68, 44), left precentral gyrus (−54, −2, 40), right middle occipital gyrus (40, −78, −4), and posterior cingulate (2, −70,10). An ANCOVA of language (English, Chinese) by age (children, adults) with accuracy on the visual rhyming task as a covariate was calculated for each ROI. Here we only report regions where a significant interaction of language and age was revealed. A significant interaction was found at the left STG (F(1,91) = 9.766, p = .002) and left ITG (F(1,92) = 8.312, p = .005). Post-hoc tests revealed that the interaction was driven by greater developmental increases in English (F(1,46) = 5.500, p = .023 for STG; F(1,45) = 10.258, p =.003 for ITG) than in Chinese (F(1,44) = 1.887, p =.176 for STG; F(1,46) = 0.01, p =.922 for ITG) (see Figure 3). A significant interaction was also found at the right MOG (F(1,97) = 5.168, p = .025), which was driven by greater developmental increases in Chinese (F(1,46) = 4.991, p =.03) than in English (F(1,50) = 0.043, p =.837) (see Figure 3).
Table 4.
Brain regions that showed a significant main effect of language.
| Anatomical Region | H | BA | Voxels | Z-score | x | y | z | |
|---|---|---|---|---|---|---|---|---|
| English> Chinese | Inferior temporal gyrus, inferior occipital gyrus, middle occipital gyrus | Left | 19,37 | 411 | inf | −44 | −60 | −10 |
| Inferior frontal gyrus | Left | 45,46 | 691 | inf | −46 | 10 | 24 | |
| Medial frontal gyrus, superior frontal gyrus | Left/right | 6 | 447 | 7.25 | −4 | 12 | 52 | |
| Superior temporal gyrus | Left | 22 | 35 | 5.82 | −56 | −40 | 8 | |
| Inferior parietal lobule | Left | 40 | 250 | 5.83 | −44 | −38 | 46 | |
| Chinese> English | Middle occipital gyrus | Right | 18,19 | 137 | 4.47 | 40 | −78 | −4 |
| Superior parietal lobule | Left | 7 | 278 | 6.18 | −36 | −68 | 44 | |
| Precentral gyrus | Left | 6 | 36 | 5.84 | −54 | −2 | 40 | |
| Posterior cingulate | Right/left | – | 1601 | 7.08 | 2 | −70 | 10 |
Note: H, hemisphere; BA, Brodmann Area
Figure 3.
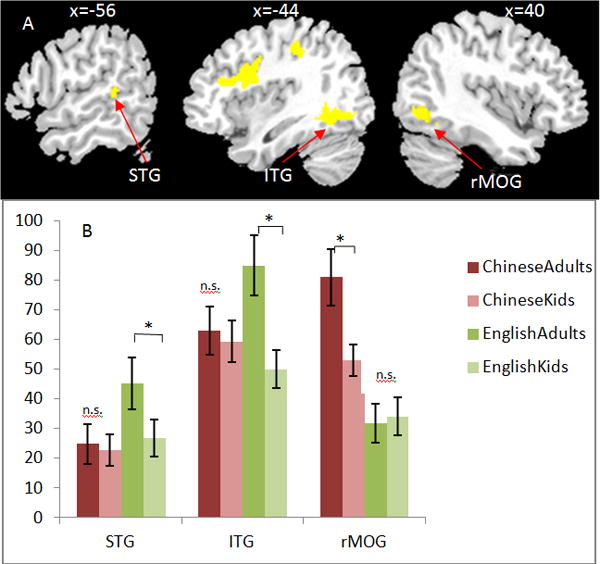
Regions of interest analysis showing interactions between language and age. (A) Regions were taken from the main effect of language that showed greater activation for English in left superior temporal gyrus (STG) and left inferior temporal gyrus (ITG) and greater activation for Chinese in right middle occipital gyrus (MOG). (B) There is a greater developmental increase in English than in Chinese at left STG and left ITG, whereas there is a greater developmental increase in Chinese than in English at right MOG. The error bars are standard errors.
In the brain-behavior correlation analysis, we extracted eigenvalues from five regions: the two clusters at the left IPL which showed a significant interaction in the voxel-wise analysis, and the left STG, the left ITG, and the right MOG which showed a significant interaction in the ROI analysis. We then correlated the eigenvalues with the accuracy in the rhyming task for the children in each language with phonological ability partialled out. For the two clusters at the left IPL, we averaged the eigenvalues and did one correlation due to their proximal location and similar function reflected by their similar patterns in the developmental effect, interaction and correlation analyses. We found that higher accuracy was correlated with greater activation in the left IPL for English children (r = .470, p < .05), but not for Chinese children, and the correlation was significantly higher for English children than for Chinese children (z = 1.805, p <.05 for right-tailed test) (see Figure 4). No significant correlations were found at the left STG, the left ITG, or the right MOG.
Figure 4.
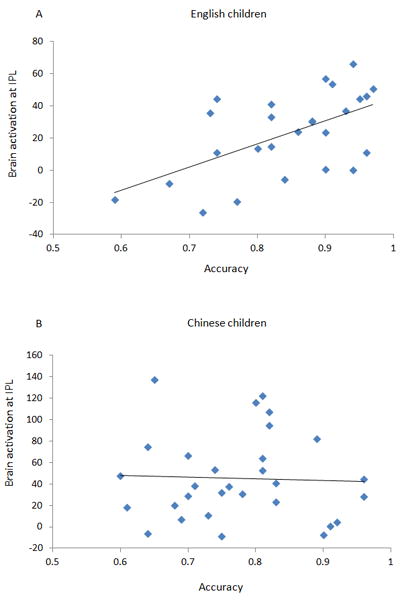
Brain-behavior correlations. (A) Accuracy on the visual rhyming task was positively correlated with activation in left IPL in English children (r=0.470, p=0.024), not Chinese children (r=−0.169, p=0.499) (B).
Discussion
In this study, we found a significant developmental increase in the involvement of the left IPL, STG and ITG only for English. We also found that brain activation at left IPL was positively correlated with accuracy on the visual rhyming judgment only within English children. In contrast, we found a developmental increase in the activation of the right MOG only for Chinese. Although orthographic and phonological processes are involved in both English and Chinese reading acquisition, our study provides evidence for the argument that even in a phonological task, phonological processing is more involved in English, whereas holistic visuo-orthographic processing is more involved in Chinese. Our study is novel and important because it is the first to show increasing divergence over development in the brain network of reading between two different writing systems, suggesting that the brain adapts to the writing system with experience.
For children learning to read English and other alphabets, the most popular and effective approach emphasizes children’s awareness of the phonological structure of speech, because this awareness helps establish the relationship between graphemes and phonemes that in turn facilitates reading development due to the semi-regular relationship between letters and sounds. There thus exists a very close association between reading and phonological processing across all alphabetic languages (Bradley & Bryant, 1983; Goswami, 1993; Perfetti, 1987). In our view, this extensive practice leads to a biological adaptation; that neural systems for phonological processing are heavily involved in reading in alphabetic languages. Therefore, the left STG that is associated with phonological representations (Turkeltaub et al., 2003; Dehaene, 2010) and left IPL that is associated with conversion between orthography and phonology (Booth, Burman, Meyer, Gitelman, Parrish & Mesulam, 2002) show developmental increases in our study.
A recent study by our group found that proximal locations in left STG ((−54, −38, 8) vs. (−56, −40, 8) in the current study) and left IPL ((−48, −44, 48) vs. (−32, −50, 48) in the current study) showed greater developmental increases in English than Chinese during an auditory rhyming task (Brennan, Cao, Pedroarena-Leal, McNorgan & Booth, 2012). There are two other studies that have also found greater involvement of left IPL in alphabetic compared to logographic reading and the location of IPL in these studies is close to the current study ((−39, −44, 54) and (−30, −62, 44) respectively) (Chen, Fu, Iversen, Smith & Matthews, 2002; Tham, Rickard, Rajapakse, Choong, Ng, Lim & Ho, 2005). The left IPL has also been found to be more involved in reading words with conflicting orthographic and phonological information (O+P−, O−P+) than words with non-conflicting information (O+P+, O−P−) (Bitan, Burman, Chou, Dong, Cone, Cao, Bigio & Booth, 2007), suggesting the fine-grain mapping between representational systems is more important when orthography and phonology is conflicting. This is consistent with the current finding that the left IPL is more involved in English reading than Chinese reading over development especially for conflicting conditions. Together, these results suggest that left STG and left IPL are important regions for phonological processing in English reading acquisition. Consistent with the argument that the nature of the mapping between orthography and phonology should make a difference in phonological processing during reading, a previous study found that more regular (transparent) alphabetic writing systems show greater involvement of the STG and IPL compared to less regular (opaque) alphabetic writing systems (Paulesu et al., 2000), suggesting the role of these regions in grapheme-phoneme conversion. Chinese is even more arbitrary in its mapping between orthography and phonology, which may explain why previous studies have found that the IPL and STG are less involved in Chinese character reading than in alphabetic reading (Sakurai et al., 2000; Chen et al., 2002; Tan et al., 2003; Tan et al., 2005; Tham et al., 2005; Liu et al., 2007), and is consistent with the lack of developmental differences in these regions for Chinese in our study.
A number of behavioral studies in English have shown that phonological skill is strongly correlated with reading performance (Ehri, Nunes, Willows, Schuster, Yaghoub-Zadeh & Shanahan, 2001; Snowling, Goulandris, Bowlby & Howell, 1986), and that phonological skill is the best predictor of reading achievement (Goswami, 1993), presumably because phonological skill is essential in the conversion from orthography to phonology at the grapheme-phoneme level. Left IPL, the region that has been proposed to be involved in the mapping between orthography and phonology during reading, has been found sensitive to reading skill in alphabetic languages (Booth et al., 2003a). Neuroimaging studies in English have also found reduced activation in the left IPL in children and adults with reading disability compared to normal controls (Rumsey, Nace, Donohue, Wise, Maisog & Andreason, 1997; Shaywitz et al., 1998). Consistent with this, we found that higher reading skill in English children is correlated with greater involvement of the left IPL even when phonological ability is partialled out, suggesting that grapheme-phoneme conversion is important in English reading independent of phonological ability. However, this region does not show a reading skill effect in Chinese children, suggesting that grapheme-phoneme conversion ability is a less important determinant of reading skill in non-alphabetic writing systems.
Computational models are an ideal tool for studying how learning to read is influenced by the nature of the mapping between orthographic and phonological representations. Harm and Seidenberg (1999) implemented a connectionist model that they initially trained on phonology, and later on the mapping from orthographic to phonological representations, simulating the natural process of early language and later reading instruction. Comparisons between the literate and illiterate network found that rhyming words were more similar in their phonological representations in the literate network, suggesting that the acquisition of an alphabetic orthography restructures the phonological network. It is possible that the orthographic training induced changes in the model correspond to the developmental and reading skill related increases in activation we see in phonological processing regions for our English speakers. Yang et al (2009) recently applied this connectionist model to learning to read in Chinese (Yang, McCandliss, Shu & Zevin, 2009). As with the Harm and Seidenberg (1999) model, mapping between orthographic and phonological representations was achieved by a layer of hidden units between these networks. Compared to models trained on English, Chinese models required more hidden units to accommodate the greater number of arbitrary relations between orthography and phonology. Because hidden unit representations in Chinese encode sub-lexical regularities at the phonetic radical level which is a larger grain size than graphemes and introduce larger errors into phonological representations, activation in the hidden units is much sparser in these models. These connectionist models thus provide an explanation for the greater developmental and reading skill increases in grapheme-phoneme level phonological processing regions observed for the English speakers in our study as compared to the Chinese speakers. As indicated in the model by Yang, et al. 2009, sub-lexical phonological processing also emerges during Chinese reading acquisition, suggesting that the mapping between Chinese characters and syllables is not completely arbitrary. Indeed, the majority (85%) of Chinese characters have a phonetic radical that provides information about the character’s pronunciation (Zhu, 1988), however only 23% of them are reliable (Xing, 2004). Therefore, sub-lexical processing in Chinese is at a larger grain size and less reliable than English, which may explain the reduced activation in the STG and IPL regions. In contrast, in a number of previous studies (Bolger et al., 2005; Tan et al., 2005; Nelson, Liu, Fiez & Perfetti, 2009), researchers have observed that left middle frontal gyrus is more involved in Chinese reading than in English reading, which may be related to the larger grain size and greater arbitrariness in Chinese reading. The current study also found a significant developmental increase in the left middle frontal gyrus within Chinese, even though there was a lack of significant interaction with language, suggesting the importance of this region in Chinese phonological processing during reading. However, this developmental increase in left middle frontal gyrus may be confounded by the fact that older Chinese participants also have greater experience of being bilingual than younger Chinese. Studies have suggested that bilingual participants tend to involve the left middle frontal gyrus to a greater degree in language tasks than monolingual participants, presumably due to language selection (Archile-Suerte, Zevin, Ramos & Hernandez, 2013; Khateb, Abutalebi, Michel, Pegna, Lee-Jahnke & Annoni, 2007).
While the importance of phonological skill in English reading acquisition is well established (McBride-Chang & Kail, 2002; Perfetti, 1984; Snowling et al., 1986), a number of studies have pointed out that phonological skill alone is insufficient for the development of reading speed and fluency; reading acquisition in English also requires development of orthographic skills (Frith, 1986; O’Brien et al., 2011). Our current results revealed a greater developmental increase for English compared to Chinese in left ITG, suggesting special importance of this orthographic processing region in English reading development. The left posterior ventral occipito-temporal cortex (−42, −70, −10) appears to be specialized for processing high spatial-frequency stimuli, whereas the homologous right cortex (42, −70, −10) seems to be specialized for processing low spatial-frequency stimuli (Seghier & Price, 2011). Their left posterior ventral occipito-temporal cortex is close to our left ITG (−44, −60, −10) and their right homologous region is also close to our rMOG where we found a greater developmental increase for Chinese than English (40, −78, −4). Similarly, other studies found that the left ITG is involved in local feature detection (i.e., letter identification) in comparison to the right MOG, which is more involved in holistic visual analysis (i.e., font style identification) (Barton, Sekunova, Sheldon, Johnston, Iaria & Scheel, 2010; Shulman, Sullivan, Gish & Sakoda, 1986; Fink, 1999; for a review, see Van Kleeck, 1989). Some have suggested that high-spatial frequency information processing is more important for English word reading, whereas low-spatial frequency information is more important for Chinese word reading (Hsiao & Cottrell, 2009). English has a large lexicon size (i.e. about 20,000 base words) but a small number of orthographic units (only 26 letters), whereas Chinese has a much smaller lexicon (i.e. about 4,500 characters) with a much larger number of orthographic units (more than 1,000 stroke patterns, a functional unit smaller than radicals and larger than strokes). Thus, compared with Chinese characters, words in the English lexicon are visually more similar to each other, requiring more in depth processing of high spatial frequency information to recognize words (Hsiao & Cottrell, 2009). The left ITG has been found to be involved in both English and Chinese reading (Fu, Chen, Smith, Iversen & Matthews, 2002; Liu, Zhang, Tang, Mai, Chen, Tardif & Luo, 2008; Xue, Jiang, Chen & Dong, 2008), but one meta-analysis study (Tan et al., 2005) found that it is more involved in English reading than in Chinese reading at (−44, −56, −12) which is close to our effect (−44, −60, −10). Our study is important because it shows that the importance of the left ITG to reading increases with experience in English but not in Chinese.
Behavioral studies suggest that Chinese reading acquisition is marked by increased reliance on orthography and decreased reliance on phonology (Peng, 1985; Song, 1995). Because Chinese characters map to spoken syllables rather than individual phonemes, Chinese readers do not rely on grapheme-phoneme conversion (Hsiao & Cottrell, 2009). In addition, because spoken Chinese is highly homophonic, a Chinese child is confronted with the fact that many written characters correspond to the same syllable. Relying on phonological units to access semantics of a printed character thus would produce an indeterminate meaning. The nature of rampant homophony of written Chinese, together with its visual-orthographic demands, has led to an emphasis on orthographic instruction in learning to read Chinese. One prevalent strategy in primary school is that children are required to spend a great amount of time repeatedly copying single characters. The extensive experience of writing facilitates the acquisition of holistic visual configurations, as shown by learning related increases in the right MOG for Chinese second language learners (Cao, Vu, Chan, Lawrence, Harris, Guan, Xu & Perfetti, 2012). Although visuo-orthographic processing has been found to be left-lateralized in the inferior temporal/fusiform gyri (the so-called visual word form area) in alphabetic languages, the right hemisphere homologous region is also involved to a lesser degree (Cohen, Lehericy, Chochon, Lemer, Rivaud & Dehaene, 2002). However, several studies have found that unlike English, visuo-orthographic processing is bilateral in Chinese reading, probably due to the greater demand on holistic processing of Chinese characters. Studies have found activation in the right MOG as well as in the left visual word form area for Chinese reading when visual perceptual processes were controlled (Luke, Liu, Wai, Wan & Tan, 2002; Bolger et al., 2005; Tan et al., 2005). It has also been documented that there is increased involvement of the right MOG over development in Chinese (Cao et al., 2010; Cao et al., 2009), such that Chinese adults have greater involvement of this region as compared to alphabetic readers (Bolger et al., 2005; Liu & Perfetti, 2003; Tan et al., 2005; Nakamura, Dehaene, Jobert, Le Bihan & Kouider, 2005). The right MOG has been found to be essential to reading in studies that reported a lesion in the right occipital cortex produces alexia in right-handed patients in alphabetic languages (Estañol, 1999; Davous & Boller 1994) and in Japanese (Hirose, 1977) . However, lesion studies have not been able to demonstrate that the right occipital cortex is more involved in Chinese reading than in English reading. Our finding of developmental increases in right MOG in Chinese but not English suggests that this region is especially important in Chinese reading acquisition presumably because of its involvement of holistic visuo-orthographic processes required by the nature of logographic writing systems.
Although the present study represents the first direct investigation of cross-script developmental changes in the neural basis of reading, there are several limitations in the current study. First, monosyllabic words were used in English, whereas two-character words were used in Chinese as monosyllabic words would not allow for a parallel phonological awareness task because of the many homophones in Chinese. This may have resulted in the rhyming judgment task in Chinese being more difficult than that in English. Even though task performance was taken account by including accuracy as a covariate in our analyses, it is possible that task differences influenced some of the cross-script developmental changes. Second, the present study used only a single task (i.e. rhyming judgment to visually presented words), and therefore, a more complex design with multiple tasks would be helpful to reveal the functions of each area in reading different scripts. Third, we had a limited behavioral battery (i.e. rhyming judgments to auditorily presented words), and therefore, other behavioral tests such as visuo-orthographic assessment would be interesting to be include in brain-behavior correlation analyses in order to provide more direct evidence of specific effect of reading experience in different scripts.
Our study provides neurological evidence that in English, as compared to Chinese, a reading task requiring access to phonological representations relies more heavily on temporo-parietal regions involved in mapping from orthography to phonological representations. In addition, our study suggests that detailed orthographic processing in left visual association areas is more characteristic of English reading, while holistic orthographic analysis in right visual association areas is more important in Chinese reading. Only by using a cross linguistic developmental design were we able to show that there is increasing divergence between alphabetic and logographic writing systems due to the unique nature of their orthographic representations and their mappings of these representations to phonology.
References
- Anthony JL, Lonigan CJ, Driscoll K, Phillips BM, Burgess SR. Phonological sensitivity: A quasi-parallel progression of word structure units and cognitive operations. Reading Research Quarterly. 2003;38:470–487. [Google Scholar]
- Archila-Suerte P, Zevin J, Ramos AI, Hernandez AE. The neural basis of non-native speech perception in bilingual children. Neuroimage. 2013;67:51–63. doi: 10.1016/j.neuroimage.2012.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barton JJ, Sekunova A, Sheldon C, Johnston S, Iaria G, Scheel M. Reading words, seeing style: the neuropsychology of word, font and handwriting perception. Neuropsychologia. 2010;48:3868–3877. doi: 10.1016/j.neuropsychologia.2010.09.012. [DOI] [PubMed] [Google Scholar]
- Bitan T, Burman DD, Chou TL, Lu D, Cone NE, Cao F, Bigio JD, Booth JR. The interaction between orthographic and phonological information in children: an fMRI study. Human Brain Mapping. 2007;28:880–891. doi: 10.1002/hbm.20313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger DJ, Perfetti CA, Schneider W. Cross-cultural effect on the brain revisited: universal structures plus writing system variation. Human Brain Mapping. 2005;25:92–104. doi: 10.1002/hbm.20124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TR, Mesulam MM. Functional anatomy of intra- and cross-modal lexical tasks. Neuroimage. 2002;16:7–22. doi: 10.1006/nimg.2002.1081. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TR, Mesulam MM. Relation between brain activation and lexical performance. Human Brain Mapping. 2003a;19:155–169. doi: 10.1002/hbm.10111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Zhang L, Choy J, Gitelman DR, Parrish TR, Mesulam MM. Modality-specific and -independent developmental differences in the neural substrate for lexical processing. Journal of Neurolinguistics. 2003b;16:383–405. [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Zhang L, Gitelman DR, Parrish TR, Mesulam MM. Development of brain mechanisms for processing orthographic and phonological representations. Journal of Cognitive Neuroscience. 2004;16:1234–1249. doi: 10.1162/0898929041920496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley L, Bryant PE. Categorizing sounds and learning to read-a causal connection. Nature. 1983;301:419–421. [Google Scholar]
- Brennan C, Cao F, Pedroarena-Leal N, McNorgan C, Booth JR. Learning to read reorganizes the oral language network only in alphabetic writing systems. Human Brain Mapping. 2012 doi: 10.1002/hbm.22147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Lee R, Shu H, Yang Y, Xu G, Li K, Booth JR. Cultural constraints on brain development: Evidence from a developmental study of visual word processing in Mandarin Chinese. Cerebral Cortex. 2010;20:1223–1233. doi: 10.1093/cercor/bhp186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Peng D, Liu L, Jin Z, Fan N, Deng Y, Booth JR. Developmental differences of neurocognitive networks for phonological and semantic processing in Chinese word reading. Human Brain Mapping. 2009;30:797–809. doi: 10.1002/hbm.20546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Vu M, Chan H, Lawrence J, Harris L, Guan Q, Xu Y, Perfetti CA. Writing affects the brain network of reading in Chinese: an fMRI study. Human Brain Mapping. 2012 doi: 10.1002/hbm.22017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen L, Lehericy S, Chochon F, Lemer C, Rivaud S, Dehaene S. Language-specific tuning of visual cortex? Functional properties of the Visual Word Form Area. Brain. 2002;125:1054–1069. doi: 10.1093/brain/awf094. [DOI] [PubMed] [Google Scholar]
- Cox RW. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research. 1996;29:162–173. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- Chen Y, Fu S, Iversen SD, Smith SM, Matthews PM. Testing for dual brain processing routes in reading: a direct contrast of Chinese character and pinyin reading using FMRI. Journal of Cognitive Neuroscience. 2002;14:1088–1098. doi: 10.1162/089892902320474535. [DOI] [PubMed] [Google Scholar]
- Davous P, Boller F. Transcortical alexia with agraphia following a right temporo-occipital hematoma in a right-handed patient. Neuropsychologia. 1994;32:1263–1272. doi: 10.1016/0028-3932(94)90108-2. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Pegado F, Braga LW, Ventura P, Nunes Filho G, Jobert A, Dehaene-Lambertz G, Kolinsky R, Morais J, Cohen L. How learning to read changes the cortical networks for vision and language. Science. 2010;330:1359–1364. doi: 10.1126/science.1194140. [DOI] [PubMed] [Google Scholar]
- Ehri LC, Nunes SR, Willows DA, Schuster BV, Yaghoub-Zadeh Z, Shanahan T. Phonemic awareness instruction helps children learn to read: Evidence from the National Reading Panel’s metanalaysis. Reading Research Quarterly. 2001;36:250–287. [Google Scholar]
- Estañol B, Vega-Boada F, Corte-Franco G, Juárez S, Hernández R, García-Ramos G. Alexia without agraphia due to the lesion in the right occipital lobe in a right-handed man. Detection of hemispheric lateralization of handedness and language in a right-handed patient. Revue Neurologique. 1999;28:243–245. [PubMed] [Google Scholar]
- Fink GR, Marshall JC, Halligan PW, Dolan RJ. Hemispheric asymmetries in global/local processing are modulated by perceptual salience. Neuropsychologia. 1999;37:31–40. doi: 10.1016/s0028-3932(98)00047-5. [DOI] [PubMed] [Google Scholar]
- Frith U. A developmental framework for developmental dyslexia. Annals Dyslexia. 1986;36:69–81. doi: 10.1007/BF02648022. [DOI] [PubMed] [Google Scholar]
- Fu S, Chen Y, Smith S, Iversen S, Matthews PM. Effects of word form on brain processing of written Chinese. Neuroimage. 2002;17:1538–1548. doi: 10.1006/nimg.2002.1155. [DOI] [PubMed] [Google Scholar]
- Goswami U. Phonological skills and learning to read. Annals of the New York Academy Sciences. 1993;682:296–311. doi: 10.1111/j.1749-6632.1993.tb22977.x. [DOI] [PubMed] [Google Scholar]
- Gountouna VE, Job DE, McIntosh AM, Moorhead TW, Lymer GK, Whalley HC, Hall J, Waiter GD, Brennan D, McGonigle DJ, Ahearn TS, Cavanagh J, Condon B, Hadley DM, Marshall I, Murray AD, Steele JD, Wardlaw JM, Lawrie SM. Functional Magnetic Resonance Imaging (fMRI) reproducibility and variance components across visits and scanning sites with a finger tapping task. Neuroimage. 2010;49:552–60. doi: 10.1016/j.neuroimage.2009.07.026. [DOI] [PubMed] [Google Scholar]
- Harm MW, Seidenberg MS. Phonology, reading acquisition, and dyslexia: insights from connectionist models. Psychological Review. 1999;106:491–528. doi: 10.1037/0033-295x.106.3.491. [DOI] [PubMed] [Google Scholar]
- Hirose G, Kin T, Murakami E. Alexia without agraphia associated with right occipital lesion. Journal of Neurology, Neurosurgery, and Psychiatry. 1977;40:225–227. doi: 10.1136/jnnp.40.3.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho CS, Chan DW, Chung KK, Lee SH, Tsang SM. In search of subtypes of Chinese developmental dyslexia. Journal of Experimental Child Psychology. 2007;97:61–83. doi: 10.1016/j.jecp.2007.01.002. [DOI] [PubMed] [Google Scholar]
- Hsiao JH, Cottrell GW. Not all visual expertise is holistic, but it may be leftist: the case of Chinese character recognition. Psychological Science. 2009;20:455–463. doi: 10.1111/j.1467-9280.2009.02315.x. [DOI] [PubMed] [Google Scholar]
- Khateb A, Abutalebi J, Michel CM, Pegna AJ, Lee-Jahnke H, Annoni JM. Language selection in bilinguals: a spatio-temporal analysis of electric brain activity. International Journal of Psychophysiology. 2007;65:201–213. doi: 10.1016/j.ijpsycho.2007.04.008. [DOI] [PubMed] [Google Scholar]
- Kim KK, Byun E, Lee SK, Gaillard WD, Xu B, Theodor WH. Verbal working memory of Korean–English bilinguals: An fMRI study. Journal of Neurolinguistics. 2011;24:1–13. [Google Scholar]
- Liu C, Zhang W, Tang Y, Mai X, Chen H, Tardif T, Luo YJ. The visual word form area: evidence from an fMRI study of implicit processing of Chinese characters. Neuroimage. 2008;40:1350–1361. doi: 10.1016/j.neuroimage.2007.10.014. [DOI] [PubMed] [Google Scholar]
- Liu Y, Dunlap S, Fiez J, Perfetti CA. Evidence for neural accommodation to a writing system following learning. Human Brain Mapping. 2007;28:1223–1234. doi: 10.1002/hbm.20356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Perfetti CA. The time course of brain activity in reading English and Chinese: an ERP study of Chinese bilinguals. Human Brain Mapping. 2003;18:167–175. doi: 10.1002/hbm.10090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luke KK, Liu HL, Wai YY, Wan YL, Tan LH. Functional anatomy of syntactic and semantic processing in language comprehension. Human Brain Mapping. 2002;16:133–145. doi: 10.1002/hbm.10029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo B, Shan C, Zhu R, Weng X, He S. Functional foveal splitting: evidence from neuropsychological and multimodal MRI investigations in a Chinese patient with a splenium lesion. PLoS One. 2011;6:e23997. doi: 10.1371/journal.pone.0023997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machilsen B, d’Agostino E, Maes F, Vandermeulen D, Hahn HK, Lagae L, Stiers P. Linear normalization of MR brain images in pediatric patients with periventricular leukomalacia. Neuroimage. 2007;35:686–97. doi: 10.1016/j.neuroimage.2006.12.037. [DOI] [PubMed] [Google Scholar]
- McBride-Chang C, Cho J, Liu H, Wagner R, Shu H, Zhou A, Cheuk C, Muse A. Changing models across cultures: associations of phonological awareness and morphological structure awareness with vocabulary and word recognition in second graders from Beijing, Hong Kong, Korea, and the United States. Journal of Experimental Child Psychology. 2005a;92:140–160. doi: 10.1016/j.jecp.2005.03.009. [DOI] [PubMed] [Google Scholar]
- McBride-Chang C, Chow BW-Y, Zhong YP, Burgess S, Hayward W. Chinese character acquisition and visual skills in two Chinese scripts. Reading and Writing. 2005b;18:99–128. [Google Scholar]
- McBride-Chang C, Kail RV. Cross-cultural similarities in the predictors of reading acquisition. Child Development. 2002;73:1392–1407. doi: 10.1111/1467-8624.00479. [DOI] [PubMed] [Google Scholar]
- Muter V, Hulme C, Snowling M, Taylor S. Segmentation, not rhyming, predicts early progress in learning to read. Journal of Experimental Child Psychology. 1997;65:370–396. doi: 10.1006/jecp.1996.2365. [DOI] [PubMed] [Google Scholar]
- Nakamura K, Dehaene S, Jobert A, Le Bihan D, Kouider S. Subliminal convergence of Kanji and Kana words: further evidence for functional parcellation of the posterior temporal cortex in visual word perception. Journal of Cognitive Neuroscience. 2005;17:954–968. doi: 10.1162/0898929054021166. [DOI] [PubMed] [Google Scholar]
- Nation K, Hulme C. Phonemic segmentation, not onset-rime segmentation, predicts early reading and spelling skills. Reading Research Quarterly. 1997;32:154–167. [Google Scholar]
- Nelson JR, Liu Y, Fiez J, Perfetti CA. Assimilation and accommodation patterns in ventral occipitotemporal cortex in learning a second writing system. Human Brain Mapping. 2009;30:810–820. doi: 10.1002/hbm.20551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Brien BA, Wolf M, Miller LT, Lovett MW, Morris R. Orthographic processing efficiency in developmental dyslexia: an investigation of age and treatment factors at the sublexical level. Annals of Dyslexia. 2011;61:111–135. doi: 10.1007/s11881-010-0050-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olulade OA, Flowers DL, Napoliello EM, Eden GF. Developmental differences for word processing in the ventral stream. Brain and Language. 2013;125:134–145. doi: 10.1016/j.bandl.2012.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paulesu E, McCrory E, Fazio F, Menoncello L, Brunswick N, Cappa SF, Cotelli M, Cossu G, Corte F, Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith CD, Frith U. A cultural effect on brain function. Nature Neuroscience. 2000;3:91–96. doi: 10.1038/71163. [DOI] [PubMed] [Google Scholar]
- Peng D, Guo DJ, Zhang SL. Access to the lexical information of Chinese characters during semantic judgment. Acta Psychologica Sinica. 1985;3:227–233. [Google Scholar]
- Perfetti CA. Reading acquisition and beyond: Decoding includes cognition. American Journal of Education. 1984;93:40–60. [Google Scholar]
- Perfetti CA. Language, Speech, and Print: Some Asymmetries in the Acquisition of Literacy. In: Horowitz R, Samuels SJ, editors. Comprehending Oral and Written Language. New York: Academic Press; 1987. pp. 355–369. [Google Scholar]
- Price CJ, Devlin JT. The interactive account of ventral occipitotemporal contributions to reading. Trends in Cognitive Sciences. 2011;15:246–253. doi: 10.1016/j.tics.2011.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rumsey JM, Nace K, Donohue B, Wise D, Maisog JM, Andreason P. A positron emission tomographic study of impaired word recognition and phonological processing in dyslexic men. Archives of Neurology. 1997;54:562–573. doi: 10.1001/archneur.1997.00550170042013. [DOI] [PubMed] [Google Scholar]
- Sakurai Y, Momose T, Iwata M, Sudo Y, Ohtomo K, Kanazawa I. Different cortical activity in reading of Kanji words, Kana words and Kana nonwords. Cognitive Brain Research. 2000;9:111–115. doi: 10.1016/s0926-6410(99)00052-x. [DOI] [PubMed] [Google Scholar]
- Seghier ML, Price CJ. Explaining left lateralization for words in the ventral occipitotemporal cortex. The Journal of Neuroscience. 2011;31:14745–14753. doi: 10.1523/JNEUROSCI.2238-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaywitz BA, Skudlarski P, Holahan JM, Marchione KE, Constable RT, Fulbright RK, Zelterman D, Lacadie C, Shaywitz SE. Age-related changes in reading systems of dyslexic children. Annals of Neurology. 2007;61:363–370. doi: 10.1002/ana.21093. [DOI] [PubMed] [Google Scholar]
- Shaywitz SE, Shaywitz BA, Pugh KR, Fulbright RK, Constable RT, Mencl WE, Shankweiler DP, Liberman AM, Skudlarski P, Fletcher JM, Katz L, Marchione KE, Lacadie C, Gatenby C, Gore JC. Functional disruption in the organization of the brain for reading in dyslexia. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:2636–2641. doi: 10.1073/pnas.95.5.2636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shu H, Peng H, McBride-Chang C. Phonological awareness in young chinese children. Developmental Science. 2008;11:171–181. doi: 10.1111/j.1467-7687.2007.00654.x. [DOI] [PubMed] [Google Scholar]
- Shu H, Zhou X, Wu N. Utilizing phonological cues in Chinese characters: A developmental study. Acta Psychologica Sinica. 2000;32:164–169. [Google Scholar]
- Shulman GL, Sullivan MA, Gish K, Sakoda WJ. The role of spatial-frequency channels in the perception of local and global structure. Perception. 1986;15:259–273. doi: 10.1068/p150259. [DOI] [PubMed] [Google Scholar]
- Snowling M, Goulandris N, Bowlby M, Howell P. Segmentation and speech perception in relation to reading skill: a developmental analysis. Journal of Experimental Child Psychology. 1986;41:489–507. doi: 10.1016/0022-0965(86)90006-8. [DOI] [PubMed] [Google Scholar]
- Song H, Zhang HC, Shu H. Developmental changes in functions of orthography and phonology in Chinese reading. Acta Psychologica Sinica. 1995;2:139–144. [Google Scholar]
- Sutton BP, Goh J, Hebrank A, Welsh RC, Chee MW, Park DC. Investigation and validation of intersite fMRI studies using the same imaging hardware. Journal of Magnetic Resonance Imaging. 2008;28:21–28. doi: 10.1002/jmri.21419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Laird AR, Karl L, Fox PT. Neuroanatomical correlates of phonological processing of Chinese characters and alphabetic words: A meta-analysis. Human Brain Mapping. 2005;25:83–91. doi: 10.1002/hbm.20134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Spinks JA, Feng CM, Siok WT, Perfetti CA, Xiong J, Fox PT, Gao JH, Kalogirou E. Neural systems of second language reading are shaped by native language. Human Brain Mapping. 2003;18:158–166. doi: 10.1002/hbm.10089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tham WW, Rickard Liow SJ, Rajapakse JC, Choong Leong T, Ng SE, Lim WE, Ho LG. Phonological processing in Chinese-English bilingual biscriptals: an fMRI study. Neuroimage. 2005;28:579–587. doi: 10.1016/j.neuroimage.2005.06.057. [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Garaeu L, Flowers DL, Zefirro TA, Eden G. Development of the neural mechanisms for reading. Nature Neuroscience. 2003;6:767–773. doi: 10.1038/nn1065. [DOI] [PubMed] [Google Scholar]
- Van Kleeck MH. Hemispheric differences in global versus local processing of hierarchical visual stimuli by normal subjects: new data and a meta-analysis of previous studies. Neuropsychologia. 1989;27:1165–1178. doi: 10.1016/0028-3932(89)90099-7. [DOI] [PubMed] [Google Scholar]
- Wagner RK, Torgeson JK. The nature of phonological processing and its causal role in the acquisition of reading skills. Psychological Bulletin. 1987;101:192–212. [Google Scholar]
- Wilke M, Schmithorst VJ, Holland SK. Assessment of spatial normalization of whole-brain magnetic resonance images in children. Human Brain Mapping. 2002;17:48–60. doi: 10.1002/hbm.10053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xing H, Shu H, Li P. The acquisition of Chinese characters: Corpus analyses and connectionist simulations. Journal of Cognitive Science. 2004;5:1–49. [Google Scholar]
- Xue G, Jiang T, Chen C, Dong Q. Language experience shapes early electrophysiological responses to visual stimuli: the effects of writing system, stimulus length, and presentation duration. Neuroimage. 2008;39:2025–2037. doi: 10.1016/j.neuroimage.2007.10.021. [DOI] [PubMed] [Google Scholar]
- Yang J, McCandliss BD, Shu H, Zevin JD. Simulating Language-specific and Language-general Effects in a Statistical Learning Model of Chinese Reading. Journal of Memory Language. 2009;61:238–257. doi: 10.1016/j.jml.2009.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeno S. The Educator’s Word Frequency Guide. Brewster, NY: Touchstone Applied Science Associates; 1996. [Google Scholar]
- Zhou XL. Phonology plays a limited role in semantic activation. In: Peng DL, Shu H, Chen XC, editors. Cognitive research on the Chinese language. Shandong: Shandong Educational Publishing; 1997. [Google Scholar]
- Zhu X. Analysis of cueing function of phonetic components in modern Chinese. In: Yuan X, editor. Proceedings of the symposium on the Chinese language and characters. Beijing, Chinese: Guang Ming Daily Press; 1988. pp. 85–99. [Google Scholar]