

Abstract
How do flying birds respond to changing environments? The behaviour of budgerigars, Melopsittacus undulatus, was filmed as they flew through a tapered tunnel. Unlike flying insects—which vary their speed progressively and continuously by holding constant the optic flow induced by the walls—the birds showed a tendency to fly at only two distinct, fixed speeds. They switched between a high speed in the wider section of the tunnel, and a low speed in the narrower section. The transition between the two speeds was abrupt, and anticipatory. The high speed was close to the energy-efficient, outdoor cruising speed for these birds, while the low speed was approximately half this value. This is the first observation of the existence of two distinct, preferred flight speeds in birds. A dual-speed flight strategy may be beneficial for birds that fly in varying environments, with the high speed set at an energy-efficient value for flight through open spaces, and the low speed suited to safe manoeuvring in a cluttered environment. The constancy of flight speed within each regime enables the distances of obstacles and landmarks to be directly calibrated in terms of optic flow, thus facilitating simple and efficient guidance of flight through changing environments.
Keywords: birds, flight, vision, visual guidance, speed
1. Introduction
Although considerable attention has been devoted to investigating how birds migrate over long distances (for review, see [1]), we know relatively little about how birds cope with the challenges of short-range navigation. For example, do birds tailor their flight to adapt to changes in the environment? Short-range guidance has been investigated extensively in flying insects, where it has been shown that cues based on optic flow—the motion of the image of the environment in the eye—play a significant role in controlling a number of different aspects of flight (for reviews, see [2–4]). Specifically, there is good evidence, from experiments in either tapered tunnels or tunnels displaying moving patterns on the walls, to suggest that flying insects, for example, vinegar flies (Drosophila melanogaster), honeybees (Apis mellifera) and bumblebees (Bombus terrestris) regulate their flight speed by holding constant the magnitude of the optic flow that is experienced by their visual system as they fly through the environment [5–8]. This method of controlling flight speed ensures that the speed is high during flight in an open environment and is progressively reduced to lower, safer levels as the environment becomes more cluttered.
2. Material and methods
Ten birds were used in the experiments. Each bird was released four times at each end of a tapered tunnel (figure 1a, inset). Consequently, each bird experienced four flights in the widening direction of the tunnel, and four flights in the narrowing direction. This resulted in a total of 80 flights: 40 in the narrowing direction and another 40 in the widening direction.
Figure 1.
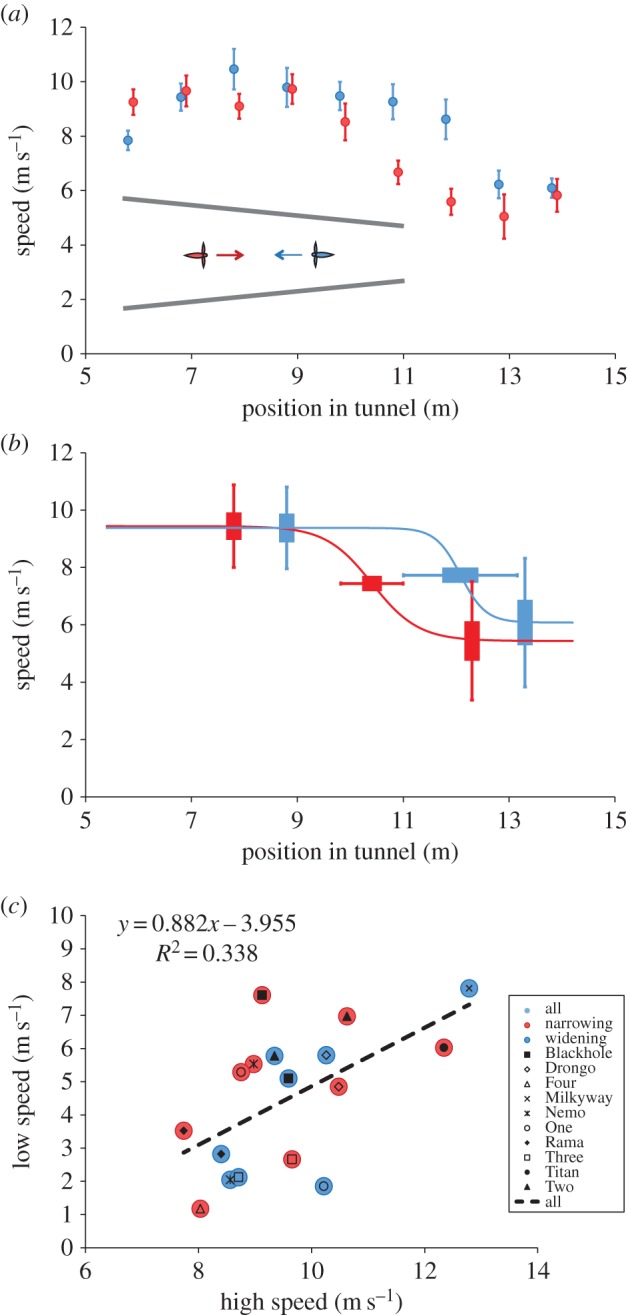
(a (inset)) Schematic overhead view of tapered tunnel, illustrating flights in the narrowing direction (red) and the widening direction (blue). (a (graph)) Comparison of mean three-dimensional speed profiles for flights in the narrowing direction (red circles) and the widening direction (blue circles), obtained by averaging data from 10 birds (four flights per bird in each direction). For clarity, the red symbols are shifted slightly to the right of the blue symbols, with the latter representing the actual positions along the tunnel. The error bars show s.e.m. (b) Variation of the high speed, the low speed and the transition points for flights in the narrowing direction (red) and the widening direction (blue) across individual birds. The error bars show s.d. (outer segment) and s.e.m. (inner segment). (c) Correlation between the high speed and the low speed for individual birds (individual symbols), for flights in the narrowing (red) and widening (blue) directions. Note that in four cases no optimal fit could be found, explaining why there are data points missing in the figure.
The birds' flights were tracked and reconstructed in three dimensions using two high-speed video cameras. The median speed of each bird in each metre segment along the tunnel's longitudinal axis was then evaluated for each flight trajectory. We then fitted the data to a logistic model, which is a general model that can assume a linear, a sigmoidal or a stepwise profile, depending upon its parameters to investigate the effects of a narrowing or a widening tunnel on flight behaviour. Details of the experimental protocol, video recording, three-dimensional reconstruction and data analysis are given in the electronic supplementary material.
3. Results
Figure 1a shows the average speed profiles of the birds for flights in the narrowing direction (red) and the widening direction (blue). The circles show mean flight speeds averaged across all birds and flights. The curves in figure 1b represent logistic functions fitted to the average data (see the electronic supplementary material). For each flight direction the speed profile, in its entirety, is approximated well by a sigmoidal fit, but poorly by a linear fit. In the wider section of the tunnel, the birds flew at a constant, high speed (9.44 m s−1). When they reached a position near the midpoint of the tunnel, they dropped their speed abruptly to a lower level (5.44 m s−1), which was maintained for the rest of the flight. When traversing the tunnel in the opposite direction (i.e. when flying through a progressively widening tunnel), the birds commenced flight at the low speed and maintained this speed until a position approximately 2 m ahead of the midpoint, beyond which they switched to the high speed. However, the steepness of the transition is the same in both directions (paired t-test, p = 0.94; see the electronic supplementary material). It must be noted that figure 1b shows the flight profiles averaged over all of the birds, and since the location of the speed transition varies slightly from bird to bird, this makes the speed transition in the average speed profile appear smoother and less abrupt than in the profiles for the individual birds (data not shown).
To examine variations in the behaviour among the 10 birds that we studied, we fitted separate logistic functions to the data of each bird, and measured the variability in the high speed, the low speed and the points of transition (switching) between the high speed and the low speed for flights in the narrowing direction (red curve, figure 1b), and between the low speed and high speed for flights in the widening direction (blue curve, figure 1b). The standard deviations and standard errors of these variables, shown in figure 1b, indicate that all of the four quantities are quite consistent across all birds.
To examine the variability across individual birds, we examined the relationship between the high speed and the low speed for each bird, for flights in the narrowing as well as the widening direction. This relationship is shown in figure 1c, which plots the high speed versus the low speed for each bird, with the circles representing data for individual birds. Inspection of these data, and a linear regression on it, reveals that (i) the low speed is correlated with the high speed (R2 = 0.34, p < 0.05), that is, a bird with a higher than average ‘high’ flight speed also tends to have a higher than average ‘low’ flight speed; and (ii) for each bird, there is a more or less constant difference, of about 6 m s−1 on average, between the high speed and the low speed. (iii) The small, correlated variation of the high speed and low speed from bird to bird (figure 1c) may reflect small variations in the physiological or behavioural characteristics of individual birds.
4. Discussion
The above observations suggest that budgerigars fly primarily at two distinct speeds. The speed in the wide section is significantly higher than that in the narrow section, regardless of flight direction (p < 0.001 in either case; pairwise two-tailed t-test, details in the electronic supplementary material). Thus, the transition between the two speeds is rather abrupt, and not gradual. Evidently, the birds tailor the speed of their flight to the passage that they are negotiating: a narrowing passage elicits a reduction in flight speed—possibly for reasons of safety—whereas a widening passage elicits an increase in flight speed, presumably to expedite the journey through the passage.
In broad terms, this behaviour is similar to that displayed by flying insects [3–8]. Bees flying through a tapered tunnel reduce their speed approximately linearly from a maximum value recorded at the widest part of the tunnel to a minimum at the narrowest part, indicating that they control their flight speed by holding constant the optic flow experienced by the visual system during flight [6,8]. However, budgerigars do not vary their speed in a linear manner. They display only two distinct flight speeds, and the transition between the two speeds is abrupt. Thus, unlike insects, the birds are not holding the induced optic flow constant; rather, they switch to the lower speed when the optic flow exceeds a certain value, and to the higher speed when the optic flow drops below this value.
Why are budgerigars different from bees? For budgerigars, oxygen consumption and respiratory rate are U-shaped functions of speed, with a minimum occurring at about 10 m s−1 [9]. Such U-shaped functions have also been measured or theoretically predicted for other bird species [10,11]. The higher flight speed of approximately 9.5 m s−1 observed in our tapered tunnel closely matches this energy-efficient speed. Thus, the birds may prefer to fly at the energy-efficient speed whenever the tunnel is wide enough to permit safe flight, and switch to the lower speed of approximately 5.5 m s−1 when the tunnel becomes narrower than a critical value. Interestingly, the lower flight speed—presumably a ‘manoeuvring’ speed—is also rather constant and prevails across a number of other experiments with budgerigars [12–14].
The data in figure 1b,c also reveal that the speed profile for flights in the widening direction can be replicated by shifting the profile for flights in the narrowing direction to the right by a distance of about 1.7 m. This suggests that birds switch their speed by looking at a section of the tunnel that is 0.85 m ahead of their current location. 0.85 m is half the distance between the switching points, as illustrated in figure 2. From the geometry of the tunnel (figure 2), we infer that the birds are looking forward at an angle of approximately 24° relative to the flight direction. (This does not imply that the birds are looking exclusively along this direction, but suggests that this is the direction of highest sensitivity of the visual subsystem that is used to assess flight speed in relation to tunnel width.) This means that the viewpoints for birds flying in the two directions will converge at a position where the width of the tunnel is 0.76 m. Thus, the birds reduce or increase their flight speed in a predictive fashion according to whether the perceived oncoming width of the tunnel becomes smaller or larger than 0.76 m. Predictive control of flight speed has also been observed in bumblebees [15]. However, while flying insects tailor their speed to the width of the passage in a smooth, continuously varying manner [3–8], budgerigars, as we show here, seem to adopt only two discrete speeds. The advantage of a dual-speed flight system that incorporates only two ‘known’, pre-set speeds is that, for each speed, the distances to obstacles can be directly calibrated in terms of the optic flow that they elicit. Additionally, the decision to switch from one speed to the other can be performed in a simple, straightforward manner by monitoring the optic flow that is generated by the oncoming environment.
Figure 2.
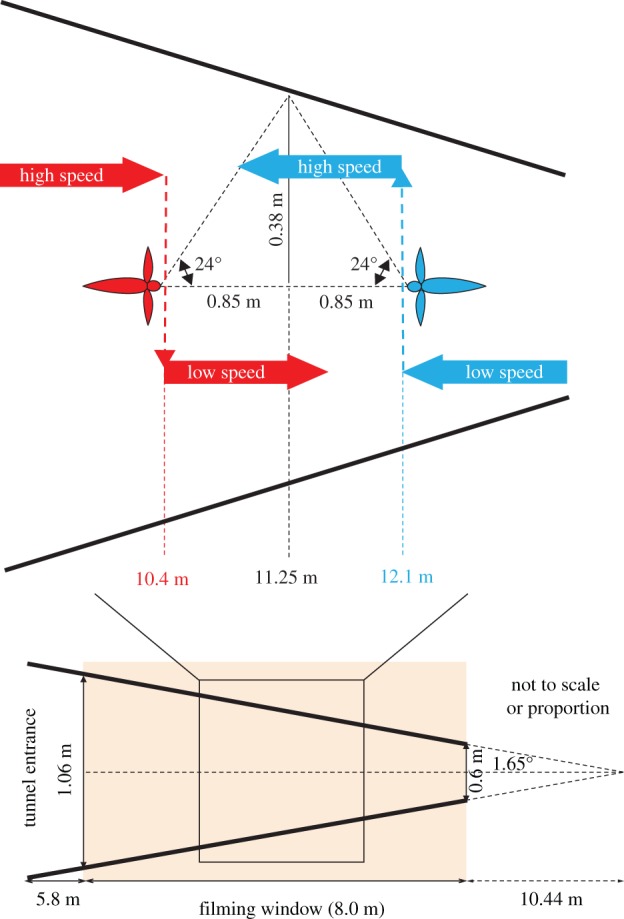
Schematic of bird flight through the tapered tunnel in the narrowing direction (red) and the widening direction (blue), showing the locations at which the flight speed drops from ‘high’ to ‘low’ (dashed red arrow) and increases from ‘low’ to ‘high’ (dashed blue arrow), based on the data shown in figure 1. Note: This figure is not to scale or proportion.
In principle, switching between the two cardinal speeds can be accomplished by gauging the width of the oncoming environment either by monitoring optic flow as we have suggested above, or, alternatively, by using stereo cues. In birds, the visual overlap ranges from 10 to 40°, and is used primarily for close-range vision mostly in the lower visual field [16]. It is therefore unlikely to provide stereo cues over our inferred frontal field, which is at least 48° wide (2 × 24°). However, our present experiments do not allow us to distinguish between the use of the two cues, which would be an interesting topic for future research.
In the case of the budgerigar, the use of two cardinal flight speeds requires that these speeds be predetermined and regulated by some means other than visual input. This could be achieved, for example, by (a) incorporating two standard flight modes that generate different thrusts, each mode representing the use of a flight speed that is locally optimum in terms of energy consumption; (b) regulating flight speed by sensing the airspeed (e.g. [17]) or (c) using a combination of (a) and (b). These possibilities could be examined by investigating flights in a tapered tunnel that provides headwinds or tailwinds at various controlled speeds.
We have seen that budgerigars switch their speed from ‘high’ to ‘low’ when the oncoming passage becomes narrower than 0.76 m. This width is roughly 2.5 times the average wingspan of these birds (around 30 cm). A recent study revealed that, during flight through very narrow apertures, budgerigars close their wings only when the aperture is narrower than their wingspan [13]. Thus, budgerigars are ‘body aware’, displaying an ability to gauge the widths of passages precisely in relation to their wingspan. Given this, one may speculate, at the risk of some oversimplification, that budgerigar flight is governed by three simple operating rules: (i) fly at the high (energy-efficient) speed of approximately 9.5 m s−1 when the passage through the environment is wider than 2.5× the wingspan; (ii) switch to the low (manoeuvring) speed (approx. 5.5 m s−1) when the oncoming passage is narrower than 2.5× the wingspan and (iii) close wings when the oncoming passage is narrower than the wingspan.
Supplementary Material
Acknowledgements
We thank Hong Diem Vo for her assistance in conducting the experiments, and the anonymous reviewers for their suggestions for improving the manuscript.
Ethics
All animal care and data collection protocols have been approved by the Animal Ethics Committee of the University of Queensland (Permit QBI/274/15/ARC).
Data accessibility
Data are available as the electronic supplementary material.
Authors' contributions
The study was designed by I.S. and M.V.S. The data were collected by I.S. Both authors wrote the manuscript, provided intellectual input, edited/approved the final version and agree to be held accountable for the work performed.
Competing interests
We declare we have no competing interests.
Funding
This work was funded by ARC Discovery Grant DP 110103277, Human Frontiers in Science Grant RGP0003/2013, a Queensland Premier's Fellowship, and an ARC Distinguished Outstanding Researcher Award (DP 140100914).
References
- 1.Wiltschko R, Wiltschko W. 2009. Avian navigation. Auk 126, 717–743. ( 10.1525/auk.2009.11009) [DOI] [Google Scholar]
- 2.Borst A. 2009. Drosophila's view on insect vision. Curr. Biol. 19, R36–R47. ( 10.1016/j.cub.2008.11.001) [DOI] [PubMed] [Google Scholar]
- 3.Srinivasan MV. 2011. Visual control of navigation in insects and its relevance for robotics. Curr. Opin. Neurobiol. 21, 535–543. ( 10.1016/j.conb.2011.05.020) [DOI] [PubMed] [Google Scholar]
- 4.Srinivasan MV. 2011. Honeybees as a model for the study of visually guided flight, navigation, and biologically inspired robotics. Physiol. Rev. 91, 413–460. ( 10.1152/physrev.00005.2010) [DOI] [PubMed] [Google Scholar]
- 5.David CT. 1982. Compensation for height in the control of groundspeed by Drosophila in a new, ‘barber's pole’ wind tunnel. J. Comp. Physiol. 147, 485–493. ( 10.1007/BF00612014) [DOI] [Google Scholar]
- 6.Srinivasan M, Zhang S, Lehrer M, Collett T. 1996. Honeybee navigation en route to the goal: visual flight control and odometry. J. Exp. Biol. 199, 237–244. [DOI] [PubMed] [Google Scholar]
- 7.Baird E, Srinivasan MV, Zhang S, Cowling A. 2005. Visual control of flight speed in honeybees. J. Exp. Biol. 208, 3895–3905. ( 10.1242/jeb.01818) [DOI] [PubMed] [Google Scholar]
- 8.Portelli G, Ruffier F, Roubieu FL, Franceschini N. 2011. Honeybees’ speed depends on dorsal as well as lateral, ventral and frontal optic flows. PLoS ONE 6, e19486 ( 10.1371/journal.pone.0019486) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tucker VA. 1968. Respiratory exchange and evaporative water loss in the flying budgerigar. J. Exp. Biol. 48, 67–87. [Google Scholar]
- 10.Bundle MW, Hansen KS, Dial KP. 2007. Does the metabolic rate–flight speed relationship vary among geometrically similar birds of different mass? J. Exp. Biol. 210, 1075–1083. ( 10.1242/jeb.02727) [DOI] [PubMed] [Google Scholar]
- 11.Tobalske BW. 2007. Biomechanics of bird flight. J. Exp. Biol. 210, 3135–3146. ( 10.1242/jeb.000273) [DOI] [PubMed] [Google Scholar]
- 12.Bhagavatula PS, Claudianos C, Ibbotson MR, Srinivasan MV. 2011. Optic flow cues guide flight in birds. Curr. Biol. 21, 1794–1799. ( 10.1016/j.cub.2011.09.009) [DOI] [PubMed] [Google Scholar]
- 13.Schiffner I, Vo HD, Bhagavatula PS, Srinivasan MV. 2014. Minding the gap: in-flight body awareness in birds. Front. Zool. 11, 64 ( 10.1186/s12983-014-0064-y) [DOI] [Google Scholar]
- 14.Schiffner I, Srinivasan MV. 2015. Direct evidence for vision-based control of flight speed in Budgerigars. Sci. Rep. 5, 10992 ( 10.1038/srep10992) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Baird E, Kornfeldt T, Dacke M. 2010. Minimum viewing angle for visually guided ground speed control in bumblebees. J. Exp. Biol. 213, 1625–1632. ( 10.1242/jeb.038802) [DOI] [PubMed] [Google Scholar]
- 16.Martin GR. 2009. What is binocular vision for? A birds’ eye view. J. Visi. 9, 14 ( 10.1167/9.11.14) [DOI] [PubMed] [Google Scholar]
- 17.Pennycuick CJ. 2008. Modelling the flying bird. Amsterdam, The Netherlands: Elsevier. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are available as the electronic supplementary material.