

Abstract
Abnormalities of membranous and endochondral ossification in patients with adolescent idiopathic scoliosis (AIS) remain incompletely understood. To investigate abnormalities in the melatonin signaling pathway and cellular response to melatonin in AIS, a case-control study of osteogenic and chondrogenic differentiation was performed using human mesenchymal stem cells (hMSCs). AIS was diagnosed by physical and radiographic examination. hMSCs were isolated from the bone marrow of patients with AIS and control subjects (n=12 each), and purified by density gradient centrifugation. The expression levels of melatonin receptors (MTs) 1 and 2 were detected by western blotting. Osteogenic and chondrogenic differentiation was induced by culturing hMSCs in osteogenic and chondrogenic media containing vehicle or 50 nM melatonin. Alkaline phosphatase (ALP) activity assays, quantitative glycosaminoglycan (GAG) analysis, and reverse transcription-quantitative polymerase chain reaction analysis were performed. Compared with controls, MT2 demonstrated low expression in the AIS group. Melatonin increased ALP activity, GAG synthesis and upregulated the expression of genes involved in osteogenic and chondrogenic differentiation including, ALP, osteopontin, osteocalcin, runt-related transcription factor 2, collagen type II, collagen type X, aggrecan and sex-determining region Y-box 9 in the normal control hMSCs, but did not affect the AIS groups. Thus, AIS hMSCs exhibit abnormal cellular responses to melatonin during osteogenic and chondrogenic differentiation, which may be associated with abnormal membranous and endochondral ossification, and skeletal growth. These results indicate a potential modulating role of melatonin via the MT2 receptor on abnormal osteogenic and chondrogenic differentiaation in patients with AIS.
Keywords: melatonin, adolescent idiopathic scoliosis, osteogenic differentiation, chondrogenic differentiation, human mesenchymal stem cells
Introduction
Adolescent idiopathic scoliosis (AIS) is a complex three-dimensional structural deformity with lateral curvature of the spine, which presents in late childhood and occurs in up to 3% of school-aged children worldwide (1,2). Genetic factors, growth and sex hormone secretion, connective tissue structure, muscle structure, vestibular dysfunction, melatonin deficiency, and platelet abnormalities are associated with the etiology of AIS (3–6). Studies have previously demonstrated that patients with AIS have abnormal skeletal growth (7) and persistent lower bone mineral density (8) compared with sex- and age-matched controls. Additionally, previous studies have suggested that relative anterior overgrowth with disproportionate endochondral-membranous bone growth contributes to the development of AIS (9). A melatonin receptor 1B (MTNR1B) gene polymorphism (10) and asymmetric expression of melatonin receptor mRNA in the bilateral paravertebral muscles (11) have been previously reported to be associated with AIS. Furthermore, neuroendocrine research findings implicating melatonin deficiency and melatonin signaling pathway dysfunction as the source of AIS have been a focus of great interest (12).
Melatonin signaling pathway dysfunction has previously been reported in the osteoblasts of patients with severe AIS (13,14). The abnormal response of osteoblasts to melatonin and abnormal expression of MTNR1B in osteoblasts of patients with AIS has been previously reported (15,16). However, to the best of our knowledge, no results of functional research on abnormalities in the melatonin signaling pathway affecting the regulation of osteogenic and chondrogenic differentiation in AIS have previously been reported.
Mesenchymal stem cells (MSCs) are multipotent stromal cells that can differentiate into a variety of unique mesenchymal cell types, including osteoblasts, chondrocytes, and adipocytes (17). Intramembranous and endochondral ossification, two important components of bone formation, begin with MSC proliferation and condensation (18). Previous studies have demonstrated that melatonin is important for the regulation of osteogenic and chondrogenic differentiation of MSCs (19–21).
The present study was conducted to provide further understanding and evidence of the expression of melatonin membrane receptor 2 (MT2) in human MSCs (hMSCs), and to examine the effects of melatonin on osteogenic and chondrogenic differentiation of hMSCs isolated from patients with AIS and normal control subjects. It was hypothesized that an abnormality of the melatonin signaling pathway may be involved in abnormal skeletal growth caused by osteogenic and chondrogenic differentiation in subjects with AIS.
Materials and methods
Patient data
The Ethics Committee of Sun Yat-sen University (Guangzhou, China) approved this study, and all participants provided written informed consent. Patients with severe AIS and sex-matched control subjects were recruited. The AIS group comprised patients who met current clinical criteria for severe AIS and were candidates for surgery. These patients provided detailed histories and underwent physical examinations, standard posteroanterior radiography of the whole spine while standing, and other tests, including magnetic resonance imaging and/or computed tomography. Patients with congenital scoliosis and scoliosis secondary to neuromuscular disorders, endocrine disorders, skeletal dysplasia, connective tissue abnormalities and syndromic disorders were excluded. Control subjects were surgery candidates recruited from the traumatology department. Two experienced orthopedic surgeons preoperatively confirmed the absence of other deformities of the skeletal system, hereditary diseases, and disorders affecting bone growth and metabolism in the control subjects.
Isolation and culture of bone marrow-derived hMSCs
hMSCs were isolated from bone marrow obtained from patients with AIS and control donors as described previously (19). The bone marrow samples were diluted with phosphate-buffered saline (PBS). Cells were then fractionated using a Lymphoprep (MP Biomedicals, LLC., Santa Ana, CA, USA) density gradient by resuspension in low-glucose Dulbecco's modified Eagle medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.), seeded, and incubated at 37°C/5% CO2. After 48 h, non-adherent cells were removed by changing the medium. Subsequently, the medium was changed every 3 days. When the cells reached 80–90% confluence, they were trypsinized, counted, and replated. Cells from passages 3–6 were used for the experiments. Phenotype analysis of cultured MSCs was conducted using a flow cytometer (data not shown), as described previously (18).
Western blotting analysis
For protein extraction, hMSCs were seeded at a density of 1×106 cells in a 75-cm2 flask and cultured in DMEM containing 10% FBS. The cells were cultured for an additional 3 days after reaching confluence. The cells were then trypsinized and lysed in radioimmunoprecipitation assay buffer containing 1 mm protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA) for 30 min, as described in a previous study (16). The mixture was centrifuged at 15,000 × g for 20 min at 4°C. The supernatant was collected for protein detection. Protein was extracted using a BCA Protein assay kit (CWbiotech, Beijing, China) and stored at −80°C.
For western blotting, ~16 µl of the prepared supernatant was mixed with an equal volume of 5X sodium dodecyl sulfate (SDS) gel loading buffer. The samples were placed in a boiling water bath for 5 min. Samples were centrifuged at 15,000 × g for 5 min. The supernatant (30 µg), in a 25 µl volume, was subjected to SDS-polyacrylamide gel electrophoresis (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using 10% separating gel and 4% stacking gel for 50 min at 180 V. The gel was removed from the tank and electroblotted onto a methanol-activated nitrocellulose membrane (EMD Millipore, Billerica, MA, USA) using the semi-dry method for 60 min at 200 mA. The membrane was washed three times with Tris-buffered saline Tween-20 (TBST) buffer and blocked with 5% skim milk for 1 h at 4°C. Subsequently, it was probed with mouse anti-human melatonin primary antibody for MT1 and MT2 (1:1,000; Santa Cruz Biotechnology Inc., Dallas, TX, USA; cat. no. sc-398788) and mouse anti-human β-actin primary antibody (1:2,000; Santa Cruz Biotechnology, Inc.; cat. no. sc-130300) overnight at 4°C. The membrane was washed three times with TBST buffer and incubated with horseradish peroxidase-conjugated goat anti-mouse secondary antibody (1:4,000; Santa Cruz Biotechnology, Inc.; cat. no. 395763) for 1 h at room temperature. After washing three times, the immunocomplex was visualized using electrochemiluminescence (ECL) western blotting detection reagents (Thermo Fisher Scientific, Inc.) and an ECL camera (ImageQuant LAS 4000 mini; GE Healthcare Life Sciences, Chalfont, UK).
Following reaction with the first primary antibody, the same membrane was rinsed with TBST buffer and the antibodies were stripped with a stripping buffer for 30 min. Then, it was incubated with anti-β-actin antibody overnight at 4°C as the reference protein. Following conjugation with the secondary antibody, the immunocomplex was visualized as described. An affinity-isolated antigen-specific antibody directed against the β-actin protein (Santa Cruz Biotechnology, Inc.) was used as a reference protein for western blots. All results were confirmed in duplicate.
Osteogenic differentiation
Osteogenic differentiation assays were conducted on four groups: Control osteogenesis (Control-Os), control osteogenesis + melatonin (Control-Os/Mel), AIS osteogenesis (AIS-Os), and AIS osteogenesis + melatonin (AIS-Os/Mel) groups. hMSCs were plated at a density of 15,000 cells/cm2 in 6-well plates for the reverse transcription-quantitative polymerase chain reaction (RT-qPCR) assay and 12-well plates for the alkaline phosphatase (ALP) activity assay in DMEM with 10% FBS, as described previously (19). When the cells reached 70–80% confluence, the growth medium was changed to osteogenic differentiation medium, which consisted of DMEM, 10% FBS, 0.1 µM dexamethasone, 10 mM β-glycerol phosphate (Sigma-Aldrich) and 50 µg/ml ascorbic acid (Sigma-Aldrich). For the ALP activity and RT-qPCR assays, cells were treated for 12 days with osteogenic differentiation medium in the presence of melatonin at a final concentration of 50 nM.
ALP activity assay
ALP activity was examined using a p-nitrophenyl phosphate (pNPP) assay (Sigma-Aldrich). Following treatment with osteogenic differentiation medium, the cells were washed and lysed in 0.2 ml PBS containing 0.1 M glycine, 1 mM MgCl2 and 0.05% Triton X-100 for 10 min at 4°C. The lysate was incubated with pNPP solution at 37°C for 30 min, and absorbance at 405 nm was measured using a microplate reader. The protein content was determined and ALP activity was expressed as p-NP units/mg protein/30 min.
Chondrogenic differentiation
Chondrogenic differentiation assays were also performed on four groups: Control chondrogenesis (Control-CHO), control chondrogenesis + melatonin (Control-CHO/Mel), AIS chondrogenesis (AIS-CHO), and AIS chondrogenesis + melatonin (AIS-CHO/Mel) groups. A high-density micromass culture system was used as described previously (22). Briefly, culture-expanded MSCs were trypsinized, washed, and then resuspended at a density of 2×107 cells/ml in a chemically defined chondrogenic medium consisting of high-glucose DMEM supplemented with 10 ng/ml recombinant human transforming growth factor-β3 (Peprotech, Inc., Rocky Hill, NJ, USA), 100 nM dexamethasone (Sigma-Aldrich), 50 µg/ml ascorbic acid 2-phosphate (Sigma-Aldrich), 1 mM sodium pyruvate (Sigma-Aldrich), 40 µg/ml proline (Sigma-Aldrich), and insulin-transferrin-selenium + Universal Culture Supplement Premix (BD Biosciences, Franklin Lakes, NJ, USA; final concentrations, 6.25 µg/ml bovine insulin, 6.25 µg/ml transferrin, 6.25 µg/ml selenous acid, 5.33 µg/ml linoleic acid, and 1.25 mg/ml bovine serum albumin). Droplets (15 µl) were placed carefully in each interior well of a 24-well plate. Cells were allowed to adhere at 37°C for 1.5 h, followed by addition of 500 µl chondrogenic medium containing vehicle or 50 nM melatonin (Sigma-Aldrich). The medium was changed every 3 days, and induced cartilage tissues were harvested on day 14 for RT-qPCR assay and on day 21 for quantitative analysis of glycosaminoglycan (GAG).
Quantitative analysis of GAG
Micromasses were washed and digested in PBS containing 0.03% papain (Merck Millipore, Darmstadt, Germany), 5 mM cysteine hydrochloride (Sigma-Aldrich), and 10 mM ethylenediaminetetraacetic acid (Sigma-Aldrich) for 16 h at 65°C. GAG concentrations were measured using the 1,9-dimethylmethylene blue (DMMB) dye binding assay (Sigma-Aldrich). Briefly, an aliquot of the lysate was reacted with DMMB solution for 10 min, and absorbance at 525 nm was measured using Varioskan Flash (Thermo Scientific, Inc.). DNA concentrations were calculated using the fluorescent dye Hoechst 33258 binding assay (Sigma-Aldrich) and a SpectraMax M5 microplate reader (Molecular Devices, LLC, Sunnyvale, CA, USA). For comparison, GAG content was normalized to DNA content.
RT-PCR analysis
Total RNA was extracted from cells or micromasses using RNAiso Plus reagent (Takara Biotechnology Co., Ltd., Dalian, China) and then converted to cDNA using PrimeScript RT master mix (Takara Biotechnology Co., Ltd.), according to the manufacturer's protocols. qPCR was performed with an iQ5 system (Bio-Rad Laboratories, Inc.) using SYBR Green I master mix (Toyobo Co., Ltd., Osaka, Japan), with the following thermocycling conditions: Initial denaturation, 95°C for 10 min; 40 cycles of 95°C for 15 sec and 60°C for 60 sec. The expressions of osteogenic marker genes, including those for ALP, osteopontin, osteocalcin, and runt-related transcription factor 2 (RUNX2); and chondrogenic marker genes, including those for collagen type II (COL2A1), collagen type X (COL10A1), aggrecan, and sex-determining region Y-box 9 (SOX9), were examined. The expression level of the glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) served as a reference. The primer sequences used are listed in Table I. Each PCR was processed in triplicate. The Cq value of GAPDH was subtracted from that of the target gene (ΔCq), and the average of triplicate ΔCq values was recorded. The relative expression level of each gene was determined using the 2−ΔΔCq method (23).
Table I.
Primers used for reverse transcription-quantitative polymerase chain reaction assays.
| Gene (accession no.) | Primer sequence | Product length (bp) |
|---|---|---|
| GAPDH (NM_002046) | 5′-AGAAAAACCTGCCAAATATGATGAC-3′ | 126 |
| 5′-TGGGTGTCGCTGTTGAAGTC-3′ | ||
| ALP (NM_000478.3) | 5′-AGCACTCCCACTTCATCTGGAA-3′ | 77 |
| 5′-GAGACCCAATAGGTAGTCCACATTG-3′ | ||
| Osteopontin (NM_013227) | 5′-GCGAGGAGTTGAATGGTG-3′ | 140 |
| 5′-CTTGTGGCTGTGGGTTTC-3′ | ||
| Osteocalcin (NM_199173) | 5′-CAGCGAGGTAGTGAAGAGA-3′ | 143 |
| 5′-GACTGGTGTAGCCGAAAG-3′ | ||
| RUNX2 (NM_001024630) | 5′-AGAAGGCACAGACAGAAGCTTGA-3′ | 78 |
| 5′-AGGAATGCGCCCTAAATCACT-3′ | ||
| COL2A1 (NM_001844) | 5′-GGCAATAGCAGGTTCACGTACA-3′ | 79 |
| 5′-CGATAACAGTCTTGCCCCACTT-3′ | ||
| COL10A1 (NM_000493) | 5′-CAAGGCACCATCTCCAGGAA-3′ | 70 |
| 5′-AAAGGGTATTTGTGGCAGCATATT-3′ | ||
| Aggrecan (NM_001135) | 5′-TGCATTCCACGAAGCTAACCTT-3′ | 84 |
| 5′-GACGCCTCGCCTTCTTGAA-3′ | ||
| SOX9 (NM_000346) | 5′-AGCGAACGCACATCAAGAC-3′ | 110 |
| 5′-GCTGTAGTGTGGGAGGTTGAA-3′ |
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; ALP, alkaline phosphatase; RUNX2, runt-related transcription factor 2; COL2A1, collagen type II; COL10A1, collagen type X; SOX9, sex-determining region-Y box 9.
Statistical analysis
All quantitative data are presented as the mean ± standard deviation. Differences in responses between hMSCs from AIS and control subjects were examined by one-way analysis of variance followed by a Newman-Keuls post-hoc t-test using SPSS statistical software (version 16.0; SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
Expression of MTs in hMSCs
Study participants were 12 patients with severe AIS (8 females, 4 males; age 14–19 years) and 12 control subjects (8 females, 4 males; age 15–24 years; Tables II and III). hMSCs isolated from bone marrow were defined morphologically by a fibroblast-like appearance. Overall, expression levels of MT1 and MT2 were irregular in hMSCs from patients with AIS. MT1 expression was low in MSCs from 4 subjects with AIS (P2, P3, P6 and P10), and MT2 expression was low in MSCs from 6 subjects with AIS (P2-P5, P10 and P11; extremely low in P2 and P10; Fig. 1A). By contrast, all MSC samples isolated from control subjects exhibited uniform bands for MT1 and MT2 (Fig. 1B). When the levels from all samples were combined, MT2 demonstrated significantly reduced expression in the AIS group compared with the control group (P=0.00261; Fig. 1C).
Table II.
Clinical data from patients with AIS.
| Case | Diagnosis | Major curve range | Sex | Age at surgery (years) | Cobb angle (°) | Risser sign |
|---|---|---|---|---|---|---|
| P1 | AIS | T5-T11 | F | 14 | 57 | Grade III |
| P2 | AIS | T5-L1 | F | 14 | 51 | Grade II |
| P3 | AIS | T3-T11 | F | 15 | 66 | Grade III |
| P4 | AIS | T7-L3 | F | 19 | 68 | Grade V |
| P5 | AIS | T6-L2 | F | 14 | 75 | Grade II |
| P6 | AIS | T5-T11 | F | 15 | 72 | Grade IV |
| P7 | AIS | T10-L4 | F | 14 | 74 | Grade IV |
| P8 | AIS | T3-T10 | F | 16 | 70 | Grade IV |
| P9 | AIS | T7-L3 | M | 16 | 59 | Grade IV |
| P10 | AIS | T5-L2 | M | 16 | 55 | Grade IV |
| P11 | AIS | T4-T11 | M | 14 | 67 | Grade III |
| P12 | AIS | T6-L3 | M | 15 | 58 | Grade III |
AIS, adolescent idiopathic scoliosis; T, thoracic vertebra; L, lumbar vertebra; F, female; M, male.
Table III.
Clinical data for control subjects.
| Case | Diagnosis | Sex | Age at surgery (years) |
|---|---|---|---|
| N1 | Fracture | F | 16 |
| N2 | Fracture | F | 17 |
| N3 | Musculoskeletal tumor | F | 15 |
| N4 | Fracture | F | 18 |
| N5 | Fracture | F | 19 |
| N6 | Fracture | F | 24 |
| N7 | Fracture | F | 18 |
| N8 | Fracture | F | 15 |
| N9 | Fracture | M | 16 |
| N10 | Musculoskeletal tumor | M | 17 |
| N11 | Fracture | M | 19 |
| N12 | Fracture | M | 19 |
F, female; M, male.
Figure 1.

MTs expression in hMSCs. (A) hMSCs isolated from 12 patients with AIS (P1-P12). Protein expression of MT1 and MT2 was irregular in hMSCs from patients with AIS. hMSCs from four subjects (P2, P3, P6, and P10) demonstrated low MT1 expression, and those from six subjects (P2-P5, P10, and P11) exhibited low MT2 expression. Expression was extremely low in two subjects (P2 and P10). (B) hMSCs isolated from 12 control subjects (N1-N12). All hMSCs isolated from control subjects exhibited uniform bands for MT1 and MT2. (C) Overall, MT2 demonstrated reduced expression in the AIS group compared with control group. n=12. *P<0.05 vs. control group. hMSCs, human mesenchymal stem cells; AIS, adolescent idiopathic scoliosis; MT, melatonin receptor.
Lack of response of hMSCs to melatonin treatment in osteo- genic differentiation in AIS patients
Following osteogenic differentiation for 12 days, treatment with 50 nM melatonin significantly enhanced ALP activity in normal control MSCs compared with untreated normal controls (P=0.01685), indicating that these MSCs were sensitive to melatonin during osteogenesis. By contrast, melatonin did not promote ALP activity in the AIS groups (Fig. 2). Similarly, melatonin treatment enhanced the mRNA expression levels of osteogenic marker genes, including ALP, osteopontin, osteocalcin and RUNX2, in the normal control group compared with the untreated normal control (P= 0.00472, P= 0.0177, P= 0.0285 and P=0.00726, respectively), however melatonin exhibited no effect on the AIS groups (Fig. 3). Thus, MSCs from patients with AIS appeared to be unresponsive to melatonin treatment during osteogenesis.
Figure 2.
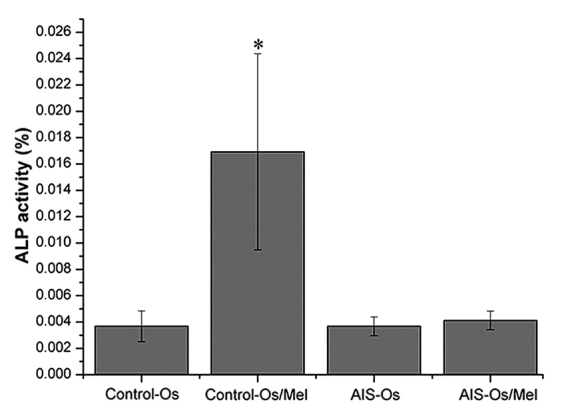
Effect of melatonin on ALP activity in human mesenchymal stem cells. Osteogenic differentiation for 12 days, treatment with 50 nM melatonin significantly enhanced ALP activity in normal control samples. By contrast, melatonin did not promote ALP activity in the AIS groups. The result of ALP activity was representative of three independent experiments. *P<0.05 vs. control-Os group. ALP, alkaline phosphatase; Os, osteogenesis; Mel, melatonin; AIS, adolescent idiopathic scoliosis.
Figure 3.

Effect of melatonin on osteogenic differentiation and the expression of marker genes in human mesenchymal stem cells. mRNA expression was measured at day 12, and relative expression levels were calculated using the 2−ΔΔCt method. Melatonin treatment enhanced the expression of osteogenic marker genes, including (A) ALP, (B) osteopontin, (C) osteocalcin, and (D) RUNX2, in the control groups, but not in the AIS groups. *P<0.05 vs. control-Os group. ALP, alkaline phosphatase; Os, osteogenesis; AIS, adolescent idiopathic scoliosis; RUNX2, runt-related transcription factor 2.
Lack of response of hMSCs to melatonin treatment in chon-drogenic differentiation in AIS patients
During chondrogenic differentiation, melatonin treatment increased GAG synthesis in normal control MSCs, which was elevated significantly on day 21 compared with untreated normal control cells (P=0.0318), however, melatonin exhibited no effect in the AIS groups (Fig. 4). Consistent with the results of quantitative GAG analysis, during chondrogenic differentiation, melatonin significantly upregulated the mRNA expression levels of COL2A1, COL10A1, aggrecan and SOX9 in the normal control group compared with untreated cells (P=0.00415, P=0.01942, P=0.02773 and P=0.0069, respectively), which are critical for chondrogenic differentiation, however, no effect was observed in the AIS groups (Fig. 5). The SOX9 level was marginally downregulated in the AIS-CHO/Mel group compared with the AIS-CHO group, although this difference was not significant.
Figure 4.

Effect of melatonin on quantitative analysis of GAG in human mesenchymal stem cells. GAG content was analyzed quantitatively and normalized by DNA content after 21 days of differentiation. Melatonin treatment increased GAG synthesis significantly in the normal control groups, but not in the AIS groups. *P<0.05 vs. control-CHO group. GAG, glycoaminoglycan; CHO, chondrogenesis; AIS, adolescent idiopathic scoliosis; Mel, melatonin.
Figure 5.

Effect of melatonin on chondrogenic differentiation and the expression of marker genes in human mesenchymal stem cells. mRNA expression levels were measured using reverse transcription-quantitative polymerase chain reaction and normalized to GAPDH expression after 14 days of differentiation. Relative expression levels were calculated using the 2−ΔΔCt method. Melatonin significantly upregulated the expression of (A) COL2A1, (B) COL10A1, (C) the aggrecan gene and (D) SOX9 in the normal control groups, but not in the AIS groups. *P<0.05 vs. control-CHO group. COL2A1, collagen type II; CHO, chondrogenesis; AIS, adolescent idiopathic scoliosis; Mel, melatonin; COL10A1, collagen type X; SOX9, sex-determining region-Y box 9.
Discussion
This study investigated the effects of melatonin on osteogenic and chondrogenic differentiation of hMSCs in patients with AIS and control subjects. The lack of hMSC response to melatonin during osteogenic and chondrogenic differentiation in AIS samples may be attributed to alterations in the melatonin signaling pathway.
MT1 and MT2, the two MT subtypes in mammals, are members of the seven-transmembrane domain G protein-coupled receptor family (24). Multiple studies have demonstrated that an MT2 polymorphism is associated with the occurrence of AIS and the severity of spinal curvature (10,16,25,26). Expression of MT2 mRNA is asymmetric in the bilateral paravertebral muscles, with greater expression levels on the concave compared with the convex side in patients with AIS (11). Wang et al (27) also demonstrated that mRNA expression of MT2 was significantly reduced in growth plate chondrocytes from patients with AIS compared with control subjects. In the present study, MT2 expression was reduced in the AIS group compared with control group. MT1 expression was lower in 4 subjects with AIS and MT2 expression was lower in 6 subjects with AIS (extremely low in 2 subjects). Subjects P2 and P10 demonstrated low MT1 expression and extremely low MT2 expression, however, these factors were not associated with AIS severity; these patients demonstrated Cobb angles of 51° and 55°, respectively, which were not the most severe in this sample population. However, in a previous study MT2 expression was not detectable in the osteoblasts of 4 out of 11 patients with AIS (15). Additionally, a previous study demonstrated that longer arm span, as part of abnormal systemic skeletal growth, was correlated with low MT2 expression in subjects with AIS (28). Comprehensive investigation of MT2, including RNA and protein expression, and also the gene promoter polymorphism and signaling pathway analysis, should be performed using hMSCs from subjects with AIS and controls to further understand potential MT2 abnormalities.
Melatonin enhances ALP activity in the differentiation processes of hMSCs cultured in osteogenic medium via MT2 and the mitogen-activated protein kinase kinase (MEK)/extracellular signal-regulated kinase 1/2 (ERK 1/2) signaling cascade (29). Previously, melatonin was demonstrated to stimulate proliferation and type I collagen synthesis in human bone cells in vitro at maximal stimulatory doses of 50–100 nM (30), and to promote osteoblast differentiation and mineralization of matrix (31) through the bone morphogenetic protein/ERK/Wnt signaling pathways (32), suggesting that this hormone is involved in regulating bone growth. Furthermore, a previous study reported that melatonin stimulated proliferation and ALP activity during human osteoblastic differentiation in vitro, in a dose-dependent manner at pharmacological concentrations (33). Melatonin suppresses the proliferation and promotes the differentiation of rat dental papilla cells, and enhances mineralized matrix formation (34). In the present study, melatonin treatment led to increased expression of osteoblast differentiation markers in control subjects, as demonstrated by ALP activity and RT-qPCR assays. These results are in accordance with our previous research (19,20). However, melatonin exhibited no observable effect on hMSCs from subjects with AIS during osteogenic differentiation. Man et al (15) also reported that melatonin was unable to promote proliferation and differentiation of osteoblasts in subjects with AIS, which might be associated with low bone mineral density in these patients (35–38).
Proliferative and hypertrophic chondrocytes in the anterior spinal columns of patients with AIS may affect spinal curve development (39). Increased numbers of these chondrocytes were observed in pinealectomized (PNX) chickens, and rapid bone elongation was more pronounced in chickens following PNX-induced osteoporosis, which may also contribute to the development of scoliosis (40). The results of the present study demonstrated that melatonin enhanced chondrogenic differentiation of hMSCs in control subjects, in accordance with previous findings (21), however, melatonin did not affect chondrogenic differentiation in subjects with AIS. Abnormal responses to melatonin during the proliferation and differentiation of growth-plate chondrocytes in subjects with AIS have also been reported previously (27).
Previous studies have suggested that AIS is associated with the loss of coupling between endochondral and membranous ossification during the developmental period (9,41–43). Longitudinal growth, achieved mainly by endochondral ossification, is more rapid than circumferential growth in vertebral bodies. By contrast, circumferential growth in the vertebral bodies and pedicles is slower in patients with AIS and achieved by membranous ossification. Dissociation between longitudinal and circumferential growth, resulting in overgrowth of the anterior column in the scoliotic spine, may contribute to the development of AIS (9). The results of the present study suggest that the melatonin signaling pathway is dysfunctional in MSCs of patients with AIS, which may be important for abnormal membranous and endochondral ossification during growth in these subjects.
In the present study, a physiological concentration of melatonin demonstrated functional abnormalities of the melatonin signaling pathway in regulating hMSCs from patients with AIS, and responses to pharmacological and high concentrations of melatonin should be examined further. Furthermore, the mechanism underlying the effects of MT2 on bone development in patients with AIS requires further elucidation. IN the present study, there were concerns regarding the in vitro culture of hMSCs from primary cells and differentiation by osteogenesis or chondrogenesis. Thus, future studies should explore differences in treatment length and compare cellular responses to melatonin following osteogenic and chondrogenic induction, and in osteoblasts and chondrocytes from patients with AIS and controls.
To the best of our knowledge, the current report is the first to describe the abnormal response of hMSCs to melatonin during osteogenic and chondrogenic differentiation in patients with AIS. These findings provide further evidence supporting the presence of an abnormal systemic melatonin signaling pathway in these patients, which may be associated with abnormal membranous and endochondral skeletal growth. Notably, the present study suggests a possible explanation for the modulating role of melatonin via the MT2 receptor on abnormal osteogenic and chondrogenic differentiation in patients with AIS, which may be a mechanism underlying the etiopathogenesis of AIS.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (grant nos. 81371908 and 81472039), the Program for New Century Excellent Talents in University (grant no. NCET-12-0564), and Fundamental Research Funds for the Central Universities, Young Teachers Fund of Sun Yat-sen University (grant no. 13YKPY23).
References
- 1.Hresko MT. Clinical practice. Idiopathic scoliosis in adolescents. N Engl J Med. 2013;368:834–841. doi: 10.1056/NEJMcp1209063. [DOI] [PubMed] [Google Scholar]
- 2.Wise CA, Gao X, Shoemaker S, Gordon D, Herring JA. Understanding genetic factors in idiopathic scoliosis, a complex disease of childhood. Curr Genomics. 2008;9:51–59. doi: 10.2174/138920208783884874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Machida M. Cause of idiopathic scoliosis. Spine (Phila Pa 1976) 1999;24:2576–2583. doi: 10.1097/00007632-199912150-00004. [DOI] [PubMed] [Google Scholar]
- 4.Cheung KM, Wang T, Qiu GX, Luk KD. Recent advances in the aetiology of adolescent idiopathic scoliosis. Int Orthop. 2008;32:729–734. doi: 10.1007/s00264-007-0393-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Peng Y, Liang G, Pei Y, Ye W, Liang A, Su P. Genomic polymorphisms of G-protein estrogen receptor 1 are associated with severity of adolescent idiopathic scoliosis. Int Orthop. 2012;36:671–677. doi: 10.1007/s00264-011-1374-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kulis A, Goździalska A, Drag J, Jaśkiewicz J, Knapik-Czajka M, Lipik E, Zarzycki D. Participation of sex hormones in multifactorial pathogenesis of adolescent idiopathic scoliosis. Int Orthop. 2015;39:1227–1236. doi: 10.1007/s00264-015-2742-6. [DOI] [PubMed] [Google Scholar]
- 7.Chu WC, Lam WW, Chan YL, Ng BK, Lam TP, Lee KM, Guo X, Cheng JC. Relative shortening and functional tethering of spinal cord in adolescent idiopathic scoliosis? Study with multiplanar reformat magnetic resonance imaging and somatosensory evoked potential. Spine (Phila Pa 1976) 2006;31:E19–E25. doi: 10.1097/01.brs.0000193892.20764.51. [DOI] [PubMed] [Google Scholar]
- 8.Cheng JC, Hung VW, Lee WT, Yeung HY, Lam TP, Ng BK, Guo X, Qin L. Persistent osteopenia in adolescent idiopathic scoliosis-longitudinal monitoring of bone mineral density until skeletal maturity. Stud Health Technol Inform. 2006;123:47–51. [PubMed] [Google Scholar]
- 9.Guo X, Chau WW, Chan YL, Cheng JC. Relative anterior spinal overgrowth in adolescent idiopathic scoliosis. Results of disproportionate endochondral-membranous bone growth. J Bone Joint Surg Br. 2003;85:1026–1031. doi: 10.1302/0301-620X.85B7.14046. [DOI] [PubMed] [Google Scholar]
- 10.Qiu XS, Tang NL, Yeung HY, Lee KM, Hung VW, Ng BK, Ma SL, Kwok RH, Qin L, Qiu Y, Cheng JC. Melatonin receptor 1B (MTNR1B) gene polymorphism is associated with the occurrence of adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 2007;32:1748–1753. doi: 10.1097/BRS.0b013e3180b9f0ff. [DOI] [PubMed] [Google Scholar]
- 11.Qiu Y, Wu L, Wang B, Yu Y, Zhu Z. Asymmetric expression of melatonin receptor mRNA in bilateral paravertebral muscles in adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 2007;32:667–672. doi: 10.1097/01.brs.0000257536.34431.96. [DOI] [PubMed] [Google Scholar]
- 12.Lombardi G, Akoume MY, Colombini A, Moreau A, Banfi G. Biochemistry of adolescent idiopathic scoliosis. Adv Clin Chem. 2011;54:165–182. doi: 10.1016/B978-0-12-387025-4.00007-8. [DOI] [PubMed] [Google Scholar]
- 13.Azeddine B, Letellier K, Wang da S, Moldovan F, Moreau A. Molecular determinants of melatonin signaling dysfunction in adolescent idiopathic scoliosis. Clin Orthop Relat Res. 2007;462:45–52. doi: 10.1097/BLO.0b013e31811f39fa. [DOI] [PubMed] [Google Scholar]
- 14.Moreau A, Wang DS, Forget S, Azeddine B, Angeloni D, Fraschini F, Labelle H, Poitras B, Rivard CH, Grimard G. Melatonin signaling dysfunction in adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 2004;29:1772–1781. doi: 10.1097/01.BRS.0000134567.52303.1A. [DOI] [PubMed] [Google Scholar]
- 15.Man GC, Wang WW, Yeung BH, Lee SK, Ng BK, Hung WY, Wong JH, Ng TB, Qiu Y, Cheng JC. Abnormal proliferation and differentiation of osteoblasts from girls with adolescent idiopathic scoliosis to melatonin. J Pineal Res. 2010;49:69–77. doi: 10.1111/j.1600-079X.2010.00768.x. [DOI] [PubMed] [Google Scholar]
- 16.Man GC, Wong JH, Wang WW, Sun GQ, Yeung BH, Ng TB, Lee SK, Ng BK, Qiu Y, Cheng JC. Abnormal melatonin receptor 1B expression in osteoblasts from girls with adolescent idiopathic scoliosis. J Pineal Res. 2011;50:395–402. doi: 10.1111/j.1600-079X.2011.00857.x. [DOI] [PubMed] [Google Scholar]
- 17.Bruder SP, Fink DJ, Caplan AI. Mesenchymal stem cells in bone development, bone repair, and skeletal regeneration therapy. J Cell Biochem. 1994;56:283–294. doi: 10.1002/jcb.240560303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kelly DJ, Jacobs CR. The role of mechanical signals in regulating chondrogenesis and osteogenesis of mesenchymal stem cells. Birth Defects Res C Embryo Today. 2010;90:75–85. doi: 10.1002/bdrc.20173. [DOI] [PubMed] [Google Scholar]
- 19.Zhang L, Su P, Xu C, Chen C, Liang A, Du K, Peng Y, Huang D. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARγ expression and enhancing Runx2 expression. J Pineal Res. 2010;49:364–372. doi: 10.1111/j.1600-079X.2010.00803.x. [DOI] [PubMed] [Google Scholar]
- 20.Zhang L, Zhang J, Ling Y, Chen C, Liang A, Peng Y, Chang H, Su P, Huang D. Sustained release of melatonin from poly (lactic-co-glycolic acid) (PLGA) microspheres to induce osteogenesis of human mesenchymal stem cells in vitro. J Pineal Res. 2013;54:24–32. doi: 10.1111/j.1600-079X.2012.01016.x. [DOI] [PubMed] [Google Scholar]
- 21.Gao W, Lin M, Liang A, Zhang L, Chen C, Liang G, Xu C, Peng Y, Chen C, Huang D, Su P. Melatonin enhances chondrogenic differentiation of human mesenchymal stem cells. J Pineal Res. 2014;56:62–70. doi: 10.1111/jpi.12098. [DOI] [PubMed] [Google Scholar]
- 22.Zhang L, Su P, Xu C, Yang J, Yu W, Huang D. Chondrogenic differentiation of human mesenchymal stem cells: A comparison between micromass and pellet culture systems. Biotechnol Lett. 2010;32:1339–1346. doi: 10.1007/s10529-010-0293-x. [DOI] [PubMed] [Google Scholar]
- 23.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 24.von Gall C, Stehle JH, Weaver DR. Mammalian melatonin receptors: Molecular biology and signal transduction. Cell Tissue Res. 2002;309:151–162. doi: 10.1007/s00441-002-0581-4. [DOI] [PubMed] [Google Scholar]
- 25.Qiu XS, Tang NL, Yeung HY, Cheng JC, Qiu Y. Lack of association between the promoter polymorphism of the MTNR1A gene and adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 2008;33:2204–2207. doi: 10.1097/BRS.0b013e31817e0424. [DOI] [PubMed] [Google Scholar]
- 26.Nelson LM, Ward K, Ogilvie JW. Genetic variants in melatonin synthesis and signaling pathway are not associated with adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 2011;36:37–40. doi: 10.1097/BRS.0b013e3181e8755b. [DOI] [PubMed] [Google Scholar]
- 27.Wang WW, Man GC, Wong JH, Ng TB, Lee KM, Ng BK, Yeung HY, Qiu Y, Cheng JC. Abnormal response of the proliferation and differentiation of growth plate chondrocytes to melatonin in adolescent idiopathic scoliosis. Int J Mol Sci. 2014;15:17100–17114. doi: 10.3390/ijms150917100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yim AP, Yeung HY, Sun G, Lee KM, Ng TB, Lam TP, Ng BK, Qiu Y, Moreau A, Cheng JC. Abnormal skeletal growth in adolescent idiopathic scoliosis is associated with abnormal quantitative expression of melatonin receptor, MT2. Int J Mol Sci. 2013;14:6345–6358. doi: 10.3390/ijms14036345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Radio NM, Doctor JS, Witt-Enderby PA. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J Pineal Res. 2006;40:332–342. doi: 10.1111/j.1600-079X.2006.00318.x. [DOI] [PubMed] [Google Scholar]
- 30.Nakade O, Koyama H, Ariji H, Yajima A, Kaku T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J Pineal Res. 1999;27:106–110. doi: 10.1111/j.1600-079X.1999.tb00603.x. [DOI] [PubMed] [Google Scholar]
- 31.Roth JA, Kim BG, Lin WL, Cho MI. Melatonin promotes osteoblast differentiation and bone formation. J Biol Chem. 1999;274:22041–22047. doi: 10.1074/jbc.274.31.22041. [DOI] [PubMed] [Google Scholar]
- 32.Park KH, Kang JW, Lee EM, Kim JS, Rhee YH, Kim M, Jeong SJ, Park YG, Kim SH. Melatonin promotes osteoblastic differentiation through the BMP/ERK/Wnt signaling pathways. J Pineal Res. 2011;51:187–194. doi: 10.1111/j.1600-079X.2011.00875.x. [DOI] [PubMed] [Google Scholar]
- 33.Satomura K, Tobiume S, Tokuyama R, Yamasaki Y, Kudoh K, Maeda E, Nagayama M. Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J Pineal Res. 2007;42:231–239. doi: 10.1111/j.1600-079X.2006.00410.x. [DOI] [PubMed] [Google Scholar]
- 34.Liu J, Zhou H, Fan W, Dong W, Fu S, He H, Huang F. Melatonin influences proliferation and differentiation of rat dental papilla cells in vitro and dentine formation in vivo by altering mitochondrial activity. J Pineal Res. 2013;54:170–178. doi: 10.1111/jpi.12002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Burner WL, III, Badger VM, Sherman FC. Osteoporosis and acquired back deformities. J Pediatr Orthop. 1982;2:383–385. doi: 10.1097/01241398-198210000-00006. [DOI] [PubMed] [Google Scholar]
- 36.Cheng JC, Guo X. Osteopenia in adolescent idiopathic scoliosis. A primary problem or secondary to the spinal deformity? Spine (Phila Pa 1976) 1997;22:1716–1721. doi: 10.1097/00007632-199708010-00006. [DOI] [PubMed] [Google Scholar]
- 37.Cheng JC, Guo X, Sher AH. Persistent osteopenia in adolescent idiopathic scoliosis. A longitudinal follow up study. Spine (Phila Pa 1976) 1999;24:1218–1222. doi: 10.1097/00007632-199906150-00008. [DOI] [PubMed] [Google Scholar]
- 38.Yu WS, Chan KY, Yu FW, Yeung HY, Ng BK, Lee KM, Lam TP, Cheng JC. Abnormal bone quality versus low bone mineral density in adolescent idiopathic scoliosis: A case-control study with in vivo high-resolution peripheral quantitative computed tomography. Spine J. 2013;13:1493–1499. doi: 10.1016/j.spinee.2013.05.018. [DOI] [PubMed] [Google Scholar]
- 39.Zhu F, Qiu Y, Yeung HY, Lee KM, Cheng JC. Histomorphometric study of the spinal growth plates in idiopathic scoliosis and congenital scoliosis. Pediatr Int. 2006;48:591–598. doi: 10.1111/j.1442-200X.2006.02277.x. [DOI] [PubMed] [Google Scholar]
- 40.Aota Y, Terayama H, Saito T, Itoh M. Pinealectomy in a broiler chicken model impairs endochondral ossification and induces rapid cancellous bone loss. Spine J. 2013;13:1607–1616. doi: 10.1016/j.spinee.2013.05.017. [DOI] [PubMed] [Google Scholar]
- 41.Dickson RA, Lawton JO, Archer IA, Butt WP. The pathogenesis of idiopathic scoliosis. Biplanar spinal asymmetry. J Bone Joint Surg Br. 1984;66:8–15. doi: 10.1302/0301-620X.66B1.6693483. [DOI] [PubMed] [Google Scholar]
- 42.Parent S, Labelle H, Skalli W, Latimer B, de Guise J. Morphometric analysis of anatomic scoliotic specimens. Spine (Phila Pa 1976) 2002;27:2305–2311. doi: 10.1097/00007632-200211010-00002. [DOI] [PubMed] [Google Scholar]
- 43.Dayer R, Haumont T, Belaieff W, Lascombes P. Idiopathic scoliosis: Etiological concepts and hypotheses. J Child Orthop. 2013;7:11–16. doi: 10.1007/s11832-012-0458-3. [DOI] [PMC free article] [PubMed] [Google Scholar]