

Abstract
Background
Pseudomonas aeruginosa is the predominant pathogen associated with the decline of pulmonary function in cystic fibrosis (CF) patients. Both environment-to-host acquisition and patient-to-patient transmission have been described for P. aeruginosa infection. Epidemic clones and bacterial phenotypic adaptation to the CF lung have been recognised as independent risk factors for disease progression. So far, there is no established link between genotypic prevalence and phenotypic traits. Here, we look at the major CF patient cohort in Italy to identify shared P. aeruginosa clones and associated common phenotypic traits.
Results
A comprehensive analysis of P. aeruginosa genotypes to determine the presence of high-risk shared clones and their association to specific phenotypic traits has been performed in a major Italian CF centre. Pulsed-field gel electrophoresis (PFGE) of P. aeruginosa isolates from 338 CF subjects identified 43 profiles shared by two or more patients and 214 profiles exclusive to individual patients. There was no evidence of a P. aeruginosa outbreak, but four most prevalent pulsotypes were detected. Common phenotypic traits were recorded intra-pulsotypes, but we detected heterogeneity inter-pulsotypes. Two of the four major pulsotypes included P. aeruginosa isolates with hallmarks of adaptation to the CF airways, including loss of motility, low production of siderophore, pyocyanin and proteases, and antibiotic resistance. One of these pulsotypes grouped a high percentage of hypermutable isolates. No clear correlation between epidemiological and clinical data was found.
Conclusions
We conclude that CF patients of this cohort shared common pulsotypes, but their phenotypic heterogeneity indicates an absence of specific traits associated to P. aeruginosa genotypic prevalence.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-016-0760-1) contains supplementary material, which is available to authorized users.
Keywords: Cystic fibrosis, Pseudomonas aeruginosa, Bacterial genotype, Bacterial phenotype, Bacterial adaptation
Background
Pseudomonas aeruginosa is the most common respiratory pathogen in patients with cystic fibrosis (CF) infecting approximately 80 % of subjects, starting from adolescence [1]. The predominant mechanism by which P. aeruginosa is acquired is controversial. Few dominant clones, including P. aeruginosa PA14 and clone C strain, are distributed worldwide and highly prone to infect CF patients, suggesting environment-to-host acquisition [2, 3]. However patient-to-patient transmission of P. aeruginosa has been increasingly reported in a few CF centres [4]. So far, few strains, such as clone C and the Liverpool epidemic strain (LES), have been indicated as highly pathogenic and transmissible causing epidemics within and between several CF clinics [5–9]. LES and the Melbourne strains have also been associated with a worse prognosis and higher rates of mortality, respectively [10, 11]. Thus, person-to-person P. aeruginosa transmission may represent a serious threat for CF patients, and this has opened a debate on infection control issues and the management of CF patients.
The pathogenicity of P. aeruginosa in CF is promoted by the diversification of the bacterial population and the presence of multiple phenotypes [12]. Common phenotypic traits, such as mucoidy, immotility, type-III secretion system deficiency, lasR mutation, hypermutability and lipopolysaccharide (LPS) modifications are consistently acquired by most P. aeruginosa strains to promote long-term persistence in CF patients. Few of these P. aeruginosa phenotypes (e.g. mucoidy, lasR mutant phenotype and hypermutability) have been associated with the more severe lung function [13–15]. While it is well-established that the bacterial intensive genetic adaptation has a key role in the progression of chronic lung infection, the link between specific P. aeruginosa phenotypic traits and genotypic prevalence remains to be established.
In this study we addressed a comprehensive analysis of P. aeruginosa genotypes at the CF centre in Verona, Italy, to establish the presence of a prevalent clone due to possible patient-to-patient transmission and its association to specific phenotypic traits. Results did not point to the presence of a P. aeruginosa outbreak, though sporadic events of possible transmission may have occurred. However, we detected prevalent pulsotypes which are characterised by phenotypic heterogeneity. These data indicate the absence of specific traits in P. aeruginosa isolates among prevalent pulsotypes.
Methods
Patients and bacterial strains
Between July 2008 and April 2009, 1,352 clinical isolates of P. aeruginosa were sampled from 338 patients with CF attending the CF centre in Verona. Patients were followed prospectively and only those intermittently or chronically colonised were selected for the study. Isolation and identification of P. aeruginosa from sputum were carried out by plating onto MacConkey agar and incubating for 48–72 h, and by API system 20NE (bioMerieux SA, Lyon, France). Provisional isolate differentiation was made on the basis of colony size, morphology, pigmentation (visual assessment), and mucoidy. P. aeruginosa isolates were stored at −80 °C in the MAST CRYOBANK™ (Mast Diagnostics, Bootle, United Kingdom). In the CF centre, patients undergo routine sputum culture four times a year. One positive culture for each CF patient was selected for this study. Four P. aeruginosa isolates were collected from this positive culture, and one was blindly chosen for detailed analysis.
Isolates genotyping
Pulsed Field Gel Electrophoresis (PFGE) of P. aeruginosa isolates was performed using the Genepath System apparatus and the CHEF Bacterial Genomic DNA Plug kit (Bio-Rad Laboratories, Hercules, USA), following the protocol by Grundmann et al. [16]. DNA band patterns were analysed with InfoQuest FP software version 5.1 (Bio-Rad Laboratories, Hercules, USA), using the Dice correlation coefficient and Unweighted Pair Group Method with Arithmetic Mean (position tolerance and optimisation of 1.5 %) [17]. By applying the criteria of Tenover and colleagues [17], based on the differences in the numbers of bands, we identified a cut-off value of 90 % similarity to correctly cluster the PFGE profiles. A cluster was consequently defined as a group of isolates showing ≥90 % similarity of the PFGE profile.
Isolates phenotyping
Swimming and twitching capacities were evaluated as described [18]. Proteases, siderophore and pyocyanin secretion, mutation frequency and lasR mutant phenotype were assayed as described in the Additional file 1.
Minimum Inhibitory Concentrations (MIC) measurement
MICs were determined according to Clinical and Laboratory Standards Institute (CLSI) guidelines [19]. MIC90 is the concentration active against 90 % of microorganisms tested. Isolates were defined as multidrug-resistant (MDR) and extensively drug-resistant (XDR) according to the terminology approved by the European Centre for Disease Prevention and Control (ECDC) and the Centres for Disease Control and Prevention (CDC) [20].
Results
Patients with CF
The clinical data of CF patients selected for the study, including FEV1 (Forced Expiratory Volume in 1 s) and their F508del mutation status, are shown in Additional file 1: Table S1. Of the 338 patients, 161 were males and 177 females. The mean age at enrollment was 26.7 years (range 1–50 years). Nineteen pairs of siblings, including four pairs of twins, were included in the study. Seventy-eight patients were homozygous and 147 heterozygous for F508del. The mean FEV1 at the time of patient enrollment was 61.5 (range 17–126). The percentage of patients chronically colonised (at least three positive cultures recorded with at least one month intervals between them) by P. aeruginosa was 50 %, while the rest were intermittently colonised (with only one or two positive cultures), according to the European Consensus [21].
Genomic profile of P. aeruginosa isolates
PFGE was carried out for all 338 P. aeruginosa isolates. By setting the similarity cut-off at 90 %, 214 PFGE profiles were classified as unique pulsotypes of single patients (Fig. 1). Of the 43 clusters of strains detected, 25 clusters included at least two or more P. aeruginosa isolates showing identical PFGE profile (100 % similarity) or belonging to highly related subtypes (≥90 % similarity), and these were classified as “PFGE patterns” (P). The remaining 18 clusters grouped two or more P. aeruginosa isolates which were highly related to each other (≥90 % similarity), but none of them identical, and these were classified as “PFGE correlated profiles” (CP). Four major PFGE clusters, including more than three isolates, were detected: P1, P6, P14 and CP2 (Fig. 2). The contacts investigation showed no clear link among patients with the same PFGE profile. Cluster P1 included P. aeruginosa isolates from eight CF patients, distributed in four highly related subtypes. Two P1 subtypes (P1.0 and P1.1) included three indistinguishable P. aeruginosa isolates each. Cluster P6 included isolates from six patients, five of them identical (100 % similarity) (P6.0) and one highly related (97 % similarity) to subtype P6.0. Cluster P14 included isolates from 12 patients, distributed in six highly related subtypes, one of them grouping four indistinguishable isolates (P14.0) and three grouping two indistinguishable isolates each (P14.1, P14.2 and P14.3). Cluster CP2 included isolates highly related to each other derived from eight CF patients’ clustering with a similarity ≥90 % (CP2.0 – CP2.7). The remaining 304 isolates presented unique PFGE profiles (63.3 % of the total strains) or were included in other minor PFGE clusters, grouping two or three P. aeruginosa isolates (26.6 %) with a similarity of at least 90 %. One pair of siblings harboured an indistinguishable genotype (P13), while two pairs were infected by highly related P. aeruginosa isolates (CP7 and CP18). The genotype and the characteristics of patients with P. aeruginosa isolates belonging to the prevalent PFGE clusters (P1, P6, P14 and CP2) are reported in Table 1 and summarized in Table 2. Overall, these findings indicate that a P. aeruginosa outbreak was not found, despite the presence of prevalent pulsotypes, potentially due to sporadic cross-transmission.
Fig. 1.
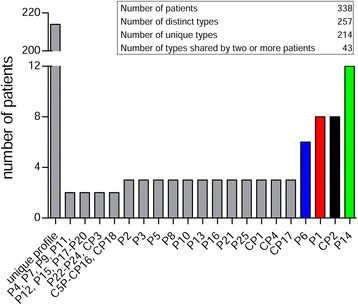
Distribution of isolates among profiles of different sizes. P. aeruginosa isolates were subjected to PFGE and DNA band patterns were analysed. The distribution of isolates among the profiles is shown
Fig. 2.
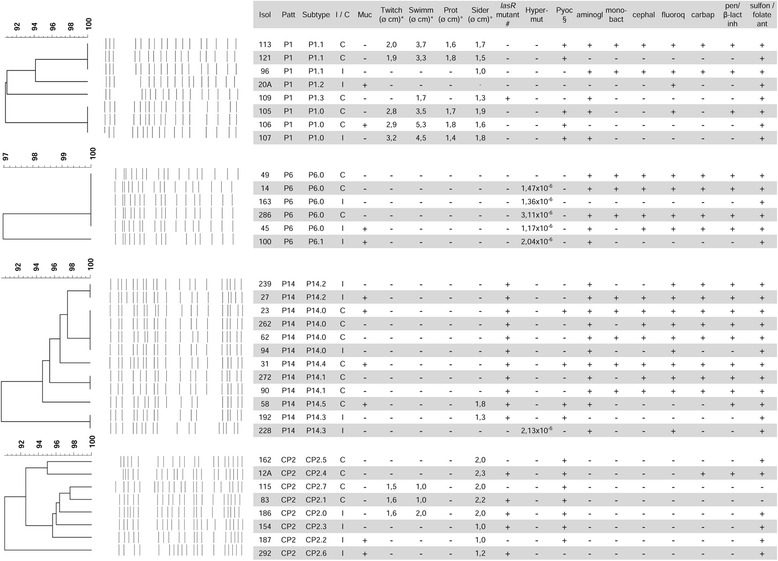
Genotypic and phenotypic characterisation of P. aeruginosa isolates belonging to the four more frequent PFGE profiles. PFGE dendrograms of the four major clusters (P1, P6, P14, CP2) are reported. Phenotypic traits of adaptation to CF airways and intermittent or chronic status of isolates are indicated. Resistance to at least one member of a specific antibiotic class is indicated as +. * swimming and twitching motility zone diameter, as measured by subsurface stab assay; ° halo diameter; # isolates with iridescent and metallic sheen of the colony surface, that is typical for a lasR mutant, are indicated as +; § visible pyocyanin presence +; no visible pyocyanin -; Abbreviations: Isol isolate, Patt PFGE pattern, I/C intermittent or chronic, Muc mucoidy, Twitch twitching motility, Swimm swimming motility, Prot proteases, Sider siderophores, Hyper-mut hyper-mutability (mutation frequency), Pyoc pyocyanin, aminogl aminoglycosides, mono-bact mono-bactams, cephal cephalosporins, fluoroq fluoroquinolones, carbap carbapenems, pen/β-lact inh penicillins/β-lactamase inhibitors, sulfon/folate ant sulfonamides/folate antagonists
Table 1.
Demographics of CF patients harbouring P. aeruginosa isolates belonging to the most representative patterns
| Isolate | Pattern | Subtype | Sex | Age (yrs range) | FEV1 (% predicted) | P. aeruginosa status | Age at 1st P. aeruginosa isolation (range) |
|---|---|---|---|---|---|---|---|
| 113 | P1 | P1.1 | F | 46–50 | 31 | C | 26–30 |
| 121a | P1 | P1.1 | M | 6–10 | 54 | C | 0–5 |
| 96b | P1 | P1.1 | F | 21–25 | 82 | I | 16–20 |
| 20A | P1 | P1.2 | F | 26–30 | 68 | I | 0–5 |
| 109b | P1 | P1.3 | M | 36–40 | 52 | C | 31–35 |
| 105 | P1 | P1.0 | M | 31–35 | 66 | C | 0–5 |
| 106 | P1 | P1.0 | M | 36–40 | 70 | C | 21–25 |
| 107b | P1 | P1.0 | M | 41–45 | 84 | I | 26–30 |
| 49b | P6 | P6.0 | M | 34 | 39 | C | 6–10 |
| 14 | P6 | P6.0 | M | 36–40 | 41 | C | 16–20 |
| 163b | P6 | P6.0 | F | 16–20 | 112 | I | 6–10 |
| 286 | P6 | P6.0 | F | 21–25 | 27 | C | 0–5 |
| 45 | P6 | P6.0 | F | 26–30 | 43 | I | 16–20 |
| 100a | P6 | P6.1 | F | 31–35 | 48 | I | 11–15 |
| 239a | P14 | P14.2 | M | 21–25 | 55 | I | 0–5 |
| 27b | P14 | P14.2 | M | 21–25 | 96 | I | 11–15 |
| 23b | P14 | P14.0 | M | 26–30 | 55 | C | 0–5 |
| 262 | P14 | P14.0 | F | 31–35 | 82 | C | 16–20 |
| 62b | P14 | P14.0 | M | 31–35 | 55 | C | 16–20 |
| 94b | P14 | P14.0 | F | 31–35 | 50 | I | 16–20 |
| 31 | P14 | P14.4 | F | 31–35 | 26 | C | 16–20 |
| 272a | P14 | P14.1 | M | 11–15 | 105 | C | 6–10 |
| 90 | P14 | P14.1 | F | 21–25 | 64 | C | 0–5 |
| 58 | P14 | P14.5 | F | 16–20 | 39 | C | 11–15 |
| 192b | P14 | P14.3 | M | 26–30 | 100 | I | 0–5 |
| 228b | P14 | P14.3 | M | 31–35 | 29 | I | 11–15 |
| 162 | CP2 | CP2.5 | M | 21–25 | 61 | C | 0–5 |
| 12A | CP2 | CP2.4 | F | 11–15 | 51,8 | C | 0–5 |
| 115b | CP2 | CP2.7 | M | 36–40 | 79 | C | 21–25 |
| 83b | CP2 | CP2.1 | M | 11–15 | 100 | C | 0–5 |
| 186b | CP2 | CP2.0 | F | 6–10 | 115 | I | 6–10 |
| 154 | CP2 | CP2.3 | F | 31–35 | 76 | I | 6–10 |
| 187 | CP2 | CP2.2 | F | 41–45 | 50 | I | 21–25 |
| 292 | CP2 | CP2.6 | M | 36–40 | 51 | I | 21–25 |
Abbreviations and symbols: M male, F female, FEV 1 forced expiratory volume in 1 s, C chronic, I intermittent; a F508del +/+; b F508del +/−
Table 2.
CF patient demographics
| Pattern (N° of patients) | Male % | Mean age (yrs) | FEV1 % | F508del homozygous % | Chronically colonised % | Mean age at the first colonisation (yrs) | Annual median rate of decline in FEV1 percent predicted over three years (%) |
|---|---|---|---|---|---|---|---|
| P1 (8) | 62.5 | 32.6 (10–46) | 63.4 (31–84) | 12.5 | 75 | 16.4 (1–32) | 4.4 (0.5 to 11.3) |
| P6 (6) | 33.3 | 27.4 (20–36) | 51.7 (27–112) | 16.7 | 67 | 11.3 (2–20) | 2.7 (−9.5 to 12) |
| P14 (12) | 50 | 26.7 (13–34) | 63 (26–100) | 14.3 | 83 | 10.3 (0–18) | 6.1 (−1.5 to 15) |
| CP2 (8) | 50 | 25.9 (10–41) | 73 (50–115) | 0 | 50 | 11.6 (0–25) | 9.2 (−2 to 21.1) |
| Others (304) | 47 | 26.5 (1–50) | 61.3 (17–113) | 24.3 | 67 | 11.6 (0–47) | 5.8 (−37.5 to 39) |
Phenotypic characterisation of the major P. aeruginosa clusters
Phenotypic characterisation was carried out on 34 P. aeruginosa isolates belonging to the four prevalent clusters (P1, P6, CP2, P14) identified by PFGE analysis (Fig. 2, Additional file 1: Table S2). Most of the isolates in cluster P1 were non-mucoid (75 %), motile (62.5 % with twitching and 75 % with swimming motility), protease, siderophore and pyocyanin producers (respectively 62.5, 87.5 and 62.5 %), non-hypermutable (100 %), lasR wt (87.5 %); two isolates were XDR and one isolate was MDR (at least MDR: 37.5 %) [20]. Isolates belonging to cluster P6 were prevalently non-mucoid (66.7 %), non-motile (100 % lacking both twitching and swimming motility), protease and siderophore low-producers (100 % lacking both protease and siderophore secretion), lasR wt (100 %), hypermutable (83.3 %) and pyocyanin low producers (100 %). Three isolates were XDR and one was MDR (at least MDR: 66.7 %). Isolates of cluster P14 were mainly non-mucoid (66.7 %), non-motile (100 % lacking both twitching and swimming motility), protease and siderophore low-producers (100 % lacking protease and 83.3 % lacking siderophore secretion), lasR mutated (91.7 %), non-hypermutable (91.7 %) and pyocyanin low-producers. Five isolates were XDR and three were MDR (at least MDR: 66.7 %). Cluster CP2 included isolates mainly non-mucoid (75 %), non-motile (62.5 % lacking both twitching and swimming motility), protease non-secreting (100 %), siderophore secreting (100 %), non-hypermutable (100 %), lasR mutated (62.5 %); none of the isolates was MDR or XDR. Notably, no clear difference in antimicrobial resistance between isolates from intermittently and chronically colonised/infected patients within the same PFGE profile was observed. Overall, these results demonstrate the phenotypic heterogeneity among major pulsotypes.
Considering all the isolates of the four clusters, it should be pointed out that only a few were mucoid even though several patients were chronically infected. Decrease in the twitching and swimming motility, reduced protease, siderophore and pyocyanin secretion, lasR mutant phenotype, hypermutability and MDR phenotype, which are hallmarks of bacterial adaptation, were detected prevalently in cluster P6 (Fig. 3).
Fig. 3.
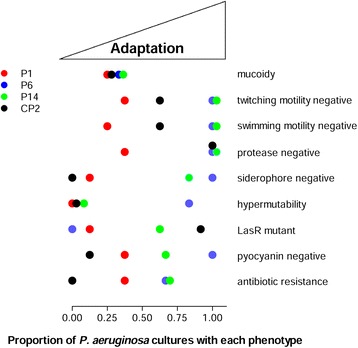
Phenotypic characterisation of P. aeruginosa isolates belonging to the four more frequent PFGE profiles. Characterisation of phenotypic traits of adaptation to CF airways were assayed on 34 P. aeruginosa isolates belonging to the four major clusters (P1, P6, P14, CP2) identified by PFGE analysis. The proportion of isolates with a specific phenotypic trait was calculated and represented for each profile
Discussion
This work provides the results of the genotypic characterisation of P. aeruginosa isolates collected from 338 patients at the CF centre in Verona between 2008 and 2009. The objective was to establish the presence of a high-risk outbreak due to possible patient-to-patient transmission. The PFGE analysis revealed the presence of 43 clusters that included isolates from two to 12 patients, indicating possible transmission in this clinic. However, a P. aeruginosa outbreak was not evident.
In this study the prevalent cluster included 12 subjects representing the 3.6 % of all the CF patients recruited. In other surveys which identified PFGE profiles shared by 7–20 % of patients, the authors concluded that the risk of cross-colonisation was extremely low [22–24]. According to these criteria, at the CF centre in Verona the risk of patient-to-patient transmission is low, although it cannot be excluded in some cases. It has been reported that different management of the P. aeruginosa infections in terms of antimicrobial chemotherapy and infection control is probably responsible for the discordant epidemiology [25]. At the CF centre in Verona, strong antibiotic treatment was administered only in the presence of pulmonary exacerbations and for eradication protocols. Overall, the pre-existing rules for infection control in this CF centre and the strategy of eradicating P. aeruginosa when isolated for the first time may have counteracted outbreaks of epidemic strains, as shown by our results. Moreover, the structure and organisation of this centre changed in 2012, in accordance with the infection control policies suggested in the European Consensus for the standards of care for CF patients [26]. The implementation of infection control strategies may have further reduced the risk of cross-transmission compared to the study period analysed (2008–2009).
The long-term colonisation of CF patients’ airways by P. aeruginosa selects clonal patho-adaptive variants with different features compared to the initially acquired strain [27]. Of these features, the most frequently described are the loss of invasive functions such as twitching and swimming motility or virulence factors as pyocyanin, several proteases, acquisition of mucoid and hypermutable phenotype. These phenotypic changes favour the persistence of P. aeruginosa variants in the CF lung. Despite the well-known relevance of P. aeruginosa phenotypes, associated to adaptation to the CF airways, to the progression of lung disease, as far as we are aware this is the first work which attempts to examine the link between predominant P. aeruginosa pulsotypes and phenotypic traits. Our phenotypic analysis showed that, despite a phenotypic homogeneity within the pulsotype, the clusters were heterogeneous when compared with each other, indicating an absence of specific traits characterising the predominant pulsotypes. More importantly, these data indicate that the potential patient-to-patient transmission cannot be due to a specific P. aeruginosa phenotypic trait analysed in this study. However, we cannot exclude that other adaptive traits (e.g., metabolic changes, lipopolysaccharide modifications) may be involved in cross-transmission.
Among the limitations of this study, we acknowledge that environmental microbiologic sampling was not performed. As a consequence we cannot exclude that the most common pulsotypes were acquired from an environmental source. In addition, we only tested a single P. aeruginosa isolate per sputum sample. A previous study reported that the great majority of patients only harbour a unique genotype [28]. However, co-infection with multiple P. aeruginosa isolates is also well-described in CF patients [29, 30]. Secondly, this study relied on the analysis of isolates recovered from the sputum. And although sputum culture appears to be a reliable means for detecting P. aeruginosa in the lower airways [31], it should be considered that different regions of the CF lung may be colonised by different isolates. We cannot exclude that other strains may be dominant in other regions of the CF lung, e.g. the more distal areas which are difficult to access, and probably affected by more stringent environmental conditions (e.g. oxygen depletion). Overall, these two limitations may lead to an underestimation of the prevalent pulsotypes, and of the genetic variability among P. aeruginosa isolates. The choice to use PFGE as a typing tool may also be questionable, as it is expensive, labour-intensive and does not allow comparisons with data from other groups, different to the multilocus sequence typing (MSLT) and random amplified polymorphic DNA (RAPD). However, PFGE was considered to be the gold-standard method for microbial typing when the survey was initiated. Recently it was shown that the discriminatory power of PFGE, MSLT and RAPD are comparable [32]. Furthermore, while MSLT was shown to be highly predictive in labelling strains as unique, PFGE was the best technique for predicting clonality. Despite all the criticisms, there is no evidence to support the claim of an outbreak at the CF centre in Verona, although the risk of cross-transmission among patients cannot be excluded.
Conclusions
This survey suggested an absence of a P. aeruginosa outbreak at the CF centre in Verona, despite the presence of prevalent clusters which may suggest cross-transmission in a few of the cases. We suggest that strict segregation procedures for infection control and the strategy of first eradication of P. aeruginosa may have reduced the incidence of nosocomial transmission. These surveillance studies, although complex and expensive, have the potential to monitor the emergence of epidemic high-risk clones and implement infection control strategies within the CF centres. In addition, the inter-pulsotypes phenotypic heterogeneity described in this work indicated the absence of specific traits associated to major pulsotypes. Further studies are needed to rule out any contribution of other determinants to transmission.
Abbreviations
CF, cystic fibrosis; CP, PFGE correlated profiles; FEV1, forced expiratory volume in 1 second; MDR, multidrug-resistant; MIC, minimum inhibitory concentrations; MSLT, multilocus sequence typing; P, PFGE patterns; PFGE, pulsed-field gel electrophoresis; RAPD, random amplified polymorphic DNA; XDR, extensively drug-resistant
Acknowledgements
Not applicable.
Funding
This study was supported by funds from the European Union Seventh Framework Programme (FP7) collaborative Action (grant NABATIVI, contract number 223670). The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Availability of data and materials
All the data supporting the findings are present in the manuscript and in the Additional file 1.
Authors’ contributions
CC, PM, DMC, BMA and AB conceived and designed the study. CC, RB, EPe and IDF designed and performed the genotypic and phenotypic analysis. PM, FF, GB and BMA were responsible for P. aeruginosa isolates collection. EPi and PI collected and analysed clinical data. GT performed statistical analysis. PM was responsible for the authorisation request to the ethical committee. CC, RB and AB integrated and analysed the data. AB coordinated the study. CC and AB wrote the manuscript. All authors have been involved in revising the manuscript critically. All authors have read and approved the final manuscript.
Competing interests
The authors declare no competing financial interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
The study was approved by the Ethical Committee of Azienda Ospedaliera Universitaria Integrata di Verona (project # 1612), in accordance with the laws of the Italian Ministero della Salute. Written informed consent to participate to the study was obtained from the patients enrolled or their parents, in accordance with the Ethical Committee rules.
Additional file
Additional Methods and References; Tables S1. Demographics of CF patients enrolled in the study; Table S2. Evaluation of pyocyanin production by P. aeruginosa isolates from CF patients. (DOCX 102 kb)
Contributor Information
Cristina Cigana, Phone: +39 02 2643 5476, Email: cigana.cristina@hsr.it.
Paola Melotti, Email: paola.melotti@ospedaleuniverona.it.
Rossella Baldan, Email: baldan.rossella@hsr.it.
Elisa Pedretti, Email: pedretti_elisa@libero.it.
Emily Pintani, Email: emily.pintani@ospedaleuniverona.it.
Patrizia Iansa, Email: patrizia.iansa@ospedaleuniverona.it.
Ida De Fino, Email: defino.ida@hsr.it.
Flavio Favari, Email: flavio.favari@ospedaleuniverona.it.
Gabriella Bergamini, Email: gabriella.bergamini@univr.it.
Gloria Tridello, Email: gloria.tridello@ospedaleuniverona.it.
Daniela M. Cirillo, Email: cirillo.daniela@hsr.it
Baroukh M. Assael, Email: baroukh.assael@ospedaleuniverona.it
Alessandra Bragonzi, Phone: +39 02 2643 4189, Email: bragonzi.alessandra@hsr.it.
References
- 1.Saiman L, Siegel J. Infection control in cystic fibrosis. Clin Microbiol Rev. 2004;17:57–71. doi: 10.1128/CMR.17.1.57-71.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wiehlmann L, Wagner G, Cramer N, Siebert B, Gudowius P, Morales G, Kohler T, van Delden C, Weinel C, Slickers P, et al. Population structure of Pseudomonas aeruginosa. Proc Natl Acad Sci U S A. 2007;104(19):8101–8106. doi: 10.1073/pnas.0609213104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Romling U, Wingender J, Muller H, Tummler B. A major Pseudomonas aeruginosa clone common to patients and aquatic habitats. Appl Environ Microbiol. 1994;60(6):1734–1738. doi: 10.1128/aem.60.6.1734-1738.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cramer N, Wiehlmann L, Tummler B. Clonal epidemiology of Pseudomonas aeruginosa in cystic fibrosis. Int J Med Microbiol. 2010;300(8):526–533. doi: 10.1016/j.ijmm.2010.08.004. [DOI] [PubMed] [Google Scholar]
- 5.Armstrong D, Nixon GM, Carzino R, Bigham A, Carlin JB, Robins-Browne RM, Grimwood K. Detection of a widespread clone of Pseudomonas aeruginosa in a pediatric cystic fibrosis clinic. Am J Respir Crit Care Med. 2002;166:983–987. doi: 10.1164/rccm.200204-269OC. [DOI] [PubMed] [Google Scholar]
- 6.Cheng K, Smyth RL, Govan JR, Doherty C, Winstanley C, Denning N, Heaf DP, van Saene H, Hart CA. Spread of beta-lactam-resistant Pseudomonas aeruginosa in a cystic fibrosis clinic. Lancet. 1996;348:639–642. doi: 10.1016/S0140-6736(96)05169-0. [DOI] [PubMed] [Google Scholar]
- 7.Fluge G, Ojeniyi B, Hoiby N, Digranes A, Ciofu O, Hunstad E, Haanaes OC, Storrosten OT. Typing of Pseudomonas aeruginosa strains in Norwegian cystic fibrosis patients. Clin Microbiol Infect. 2001;7:238–243. doi: 10.1046/j.1469-0691.2001.00247.x. [DOI] [PubMed] [Google Scholar]
- 8.Scott FW, Pitt TL. Identification and characterization of transmissible Pseudomonas aeruginosa strains in cystic fibrosis patients in England and Wales. J Med Microbiol. 2004;53:609–615. doi: 10.1099/jmm.0.45620-0. [DOI] [PubMed] [Google Scholar]
- 9.Römling U, Kader A, Sriramulu DD, Simm R, Kronvall G. Worldwide distribution of Pseudomonas aeruginosa clone C strains in the aquatic environment and cystic fibrosis patients. Environ Microbiol. 2005;7:1029–1038. doi: 10.1111/j.1462-2920.2005.00780.x. [DOI] [PubMed] [Google Scholar]
- 10.Al-Aloul M, Crawley J, Winstanley C, Hart CA, Ledson MJ, Walshaw MJ. Increased morbidity associated with chronic infection by an epidemic Pseudomonas aeruginosa strain in CF patients. Thorax. 2004;59:334–336. doi: 10.1136/thx.2003.014258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Griffiths AL, Jamsen K, Carlin JB, Grimwood K, Carzino R, Robinson PJ, Massie J, Armstrong DS. Effects of segregation on an epidemic Pseudomonas aeruginosa strain in a cystic fibrosis clinic. Am J Respir Crit Care Med. 2005;171:1020–1025. doi: 10.1164/rccm.200409-1194OC. [DOI] [PubMed] [Google Scholar]
- 12.Smith EE, Buckley DG, Wu Z, Saenphimmachak C, Hoffman LR, D'Argenio DA, Miller SI, Ramsey BW, Speert DP, Moskowitz SM, et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc Natl Acad Sci U S A. 2006;103(22):8487–8492. doi: 10.1073/pnas.0602138103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hoffman LR, Kulasekara HD, Emerson J, Houston LS, Burns JL, Ramsey BW, Miller SI. Pseudomonas aeruginosa lasR mutants are associated with cystic fibrosis lung disease progression. J Cyst Fibros. 2009;8(1):66–70. doi: 10.1016/j.jcf.2008.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li Z, Kosorok MR, Farrell PM, Laxova A, West SE, Green CG, Collins J, Rock MJ, Splaingard ML. Longitudinal development of mucoid Pseudomonas aeruginosa infection and lung disease progression in children with cystic fibrosis. JAMA. 2005;293(5):581–588. doi: 10.1001/jama.293.5.581. [DOI] [PubMed] [Google Scholar]
- 15.Ferroni A, Guillemot D, Moumile K, Bernede C, Le Bourgeois M, Waernessyckle S, Descamps P, Sermet-Gaudelus I, Lenoir G, Berche P, et al. Effect of mutator P. aeruginosa on antibiotic resistance acquisition and respiratory function in cystic fibrosis. Pediatr Pulmonol. 2009;44(8):820–825. doi: 10.1002/ppul.21076. [DOI] [PubMed] [Google Scholar]
- 16.Grundmann H, Schneider C, Hartung D, Daschner FD, Pitt TL. Discriminatory power of three DNA-based typing techniques for Pseudomonas aeruginosa. J Clin Microbiol. 1995;33(3):528–534. doi: 10.1128/jcm.33.3.528-534.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Swaminathan B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239. doi: 10.1128/jcm.33.9.2233-2239.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.O'Toole GA, Kolter R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol Microbiol. 1998;30:295–304. doi: 10.1046/j.1365-2958.1998.01062.x. [DOI] [PubMed] [Google Scholar]
- 19.CLSI M100–S24 . Performance standards for antimicrobial susceptibility testing; 24th informational supplement. Wayne: Clinical and Laboratory Standards Institute; 2014. [Google Scholar]
- 20.Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi: 10.1111/j.1469-0691.2011.03570.x. [DOI] [PubMed] [Google Scholar]
- 21.Doring G, Conway SP, Heijerman HG, Hodson ME, Hoiby N, Smyth A, Touw DJ. Antibiotic therapy against Pseudomonas aeruginosa in cystic fibrosis: a European consensus. Eur Respir J. 2000;16(4):749–767. doi: 10.1034/j.1399-3003.2000.16d30.x. [DOI] [PubMed] [Google Scholar]
- 22.Spencker F, Haupt S, Claros MC, Walter S, Lietz T, Schille R, Rodloff AC. Epidemiologic characterization of Pseudomonas aeruginosa in patients with cystic fibrosis. Clin Microbiol Infect. 2000;6:600–607. doi: 10.1046/j.1469-0691.2000.00171.x. [DOI] [PubMed] [Google Scholar]
- 23.Vosahlikova S, Drevinek P, Cinek O, Pohunek P, Maixnerova M, Urbaskova P, van den Reijden TJ, Dijkshoorn L, Nemec A. High genotypic diversity of Pseudomonas aeruginosa strains isolated from patients with cystic fibrosis in the Czech Republic. Res Microbiol. 2007;158(4):324–329. doi: 10.1016/j.resmic.2007.02.003. [DOI] [PubMed] [Google Scholar]
- 24.Speert D, Campbell ME, Henry DA, Milner R, Taha F, Gravelle A, Davidson AG, Wong LT, Mahenthiralingam E. Epidemiology of Pseudomonas aeruginosa in cystic fibrosis in British Columbia, Canada. Am J Respir Crit Care Med. 2002;166:988–993. doi: 10.1164/rccm.2203011. [DOI] [PubMed] [Google Scholar]
- 25.Cramer N, Wiehlmann L, Ciofu O, Tamm S, Hoiby N, Tummler B. Molecular epidemiology of chronic Pseudomonas aeruginosa airway infections in cystic fibrosis. PLoS One. 2012;7(11):e50731. doi: 10.1371/journal.pone.0050731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kerem E, Conway S, Elborn S, Heijerman H. Standards of care for patients with cystic fibrosis: a European consensus. J Cyst Fibros. 2005;4(1):7–26. doi: 10.1016/j.jcf.2004.12.002. [DOI] [PubMed] [Google Scholar]
- 27.Bragonzi A, Paroni M, Nonis A, Cramer N, Montanari S, Rejman J, Di Serio C, Doring G, Tummler B. Pseudomonas aeruginosa microevolution during cystic fibrosis lung infection establishes clones with adapted virulence. Am J Respir Crit Care Med. 2009;180(2):138–145. doi: 10.1164/rccm.200812-1943OC. [DOI] [PubMed] [Google Scholar]
- 28.Logan C, Habington A, Lennon G, Grogan J, Byrne M, O'Leary J, O'Sullivan N. Genetic relatedness of Pseudomonas aeruginosa isolates among a paediatric cystic fibrosis patient cohort in Ireland. J Med Microbiol. 2012;61(Pt 1):64–70. doi: 10.1099/jmm.0.035642-0. [DOI] [PubMed] [Google Scholar]
- 29.Mowat E, Paterson S, Fothergill JL, Wright EA, Ledson MJ, Walshaw MJ, Brockhurst MA, Winstanley C. Pseudomonas aeruginosa population diversity and turnover in cystic fibrosis chronic infections. Am J Respir Crit Care Med. 2011;183(12):1674–1679. doi: 10.1164/rccm.201009-1430OC. [DOI] [PubMed] [Google Scholar]
- 30.Workentine ML, Sibley CD, Glezerson B, Purighalla S, Norgaard-Gron JC, Parkins MD, Rabin HR, Surette MG. Phenotypic heterogeneity of Pseudomonas aeruginosa populations in a cystic fibrosis patient. PLoS One. 2013;8(4):e60225. doi: 10.1371/journal.pone.0060225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Blau H, Linnane B, Carzino R, Tannenbaum EL, Skoric B, Robinson PJ, Robertson C, Ranganathan SC. Induced sputum compared to bronchoalveolar lavage in young, non-expectorating cystic fibrosis children. J Cyst Fibros. 2014;13(1):106–110. doi: 10.1016/j.jcf.2013.05.013. [DOI] [PubMed] [Google Scholar]
- 32.Waters V, Zlosnik JE, Yau YC, Speert DP, Aaron SD, Guttman DS. Comparison of three typing methods for Pseudomonas aeruginosa isolates from patients with cystic fibrosis. Eur J Clin Microbiol Infect Dis. 2012;31(12):3341–3350. doi: 10.1007/s10096-012-1701-z. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All the data supporting the findings are present in the manuscript and in the Additional file 1.