

Abstract
Cold-induced mortality has historically been a key aspect of mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae), population control, but little is known about the molecular basis for cold tolerance in this insect. We used RNA-seq analysis to monitor gene expression patterns of mountain pine beetle larvae at four time points during their overwintering period—early-autumn, late-autumn, early-spring, and late-spring. Changing transcript profiles over the winter indicates a multipronged physiological response from larvae that is broadly characterized by gene transcripts involved in insect immune responses and detoxification during the autumn. In the spring, although transcripts associated with developmental process are present, there was no particular biological process dominating the transcriptome.
Keywords: Cold tolerance, Mountain pine beetle, Insect overwintering, Glycerol, Physiological stress, Lodgepole pine, Heat shock proteins, Anti-freeze, Cytochromes P450
Introduction
Changing climate and large contiguous stands of susceptible lodgepole pine (Pinus contorta Douglas) have resulted in a large mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae) outbreak in British Columbia and other parts of western North America (Raffa et al., 2008). The beetle and its fungal associates are projected to kill an estimated 750 million cubic meters of merchantable pine in British Columbia by 2017 (Natural Resources Canada, 2015). The large scale of this outbreak in British Columbia, in combination with the expansion of mountain pine beetle into previously unaffected areas of jack pine, Pinus bankisana Lamb, forests in Alberta (Cullingham et al., 2011), have caused substantial economic losses in British Columbia (Schneider et al., 2010) and continue to have massive ecological effects on the landscape.
Cold-induced mortality has historically been a key aspect of mountain pine beetle population control. The mountain pine beetle spends much of its typically one-year life cycle as larvae in the phloem tissue of susceptible host trees. While they overwinter under the bark, larvae have had, under typical conditions, to survive winter temperatures below –30°C (Cole, 1981). Mountain pine beetle larvae possess cold tolerance mechanisms, such as the production of glycerol (Bentz & Mullins, 1999) that enable supercooling of insect bodily fluids and tolerance of temperatures far below freezing. However, if a cold snap occurs early in winter, before the larvae have become cold acclimated, massive mortality can occur (Bale, 2002). Cold-induced mortality has historically controlled bark beetle populations in British Columbia and prevented the insects from moving further north or east than their historically known range. Because of the recent move of this insect across the Rocky Mountains and into the jack pine forests of Alberta (Cullingham et al., 2011; Janes et al., 2014), understanding the cold tolerance mechanisms of mountain pine beetle is becoming increasingly important for forest management and the development of predictive models.
We used RNA-seq analysis to monitor transcript profiles of mountain pine beetle larvae at four time points during their overwintering period—early-autumn, late-autumn, early-spring, and late-spring. Changing transcript profiles over the winter indicates a multipronged approach to cold readiness, overwintering, and transition into spring development in mountain pine beetle larvae. We uncovered shifts in transcript levels for several groups of genes that are likely to be important in the overwintering success of mountain pine beetle larvae.
Methods and Materials
Collection of larval specimens
Overwintering larvae were collected as described in Bonnett et al., 2012. In brief, larvae were sampled from eleven naturally infested lodgepole pine trees located at two sampling sites near Tête Jaune Cache, British Columbia, Canada (N53°3′36.00″, W119°36′54.00″and N52°55′4.00″, W119°21′23.00″). Each tree was fitted with three iButton temperature data loggers (Maxim, Sunnyvale, CA) that recorded ambient temperature every thirty minutes. Overwintering mountain pine beetle larvae were live-collected from under the bark, immediately flash frozen with liquid nitrogen in individual vials in the field, transported on dry ice, and stored at –80°C until RNA extractions were conducted. Collection dates were 26 September 2008, 7 November 2008, 25 March 2009, and 27 May 2009.
RNA extractions
Immediately prior to RNA extractions, larval beetles were transitioned to –20°C in RNAlater-ICE Frozen Tissue Transition Solution (Ambion, Life Technologies) according to the manufacturer’s protocol, and placed in individual wells of a 96-well, 1 mL round bottom polypropylene block (Corning Life Sciences) with one stainless steel grinding ball per well. RNA extractions were performed using the MagMAX-96 Total RNA Isolation Kit (Ambion, Life Technologies). Prior to extraction, RNAlater-ICE solution was removed by pipette, and 100 µL of lysis/binding solution from the MagMax-96 Total RNA Isolation Kit was immediately added to each sample well. Samples were ground for eight cycles (1500 strokes/min for 30 s followed by 30 s on ice) using a Geno/Grinder 2000 (SPEX CertiPrep). Instructions for the MagMax-96 Total RNA Isolation Kit were followed with a few minor modifications: when initially transferring samples from the grinding plate to the processing plate, the 60 µL of isopropanol was added directly to the wells in the grinding plate to aid with sample transfer; during the DNA digestion step, 60 µL of diluted TURBO DNase was added to each sample to ensure complete DNA degradation; before the final elution step, the elution buffer was heated to 70°C to ensure maximum bead dispersion, as suggested in the troubleshooting guide. RNA isolates from individual larvae were assessed for concentration and purity on a Nanodrop-1000 Spectrophotometer (Thermo Fisher Scientific).
RNA sample pooling and replicates
For the RNA-seq analysis, we used four biological replicates for each of the four sampling dates. Due to the small size of the larvae, each ‘biological’ replicate was comprised of pooled individual larvae. For the small September larvae, up to eight RNA extractions from individual larvae were pooled to get a single biological replicate. For the November, March, and May larvae, four individuals were pooled for each biological replicate. Pooled RNA samples were concentrated using the RNeasy Mini Kit (Qiagen), following the instructions for the RNA Cleanup Protocol. An aliquot of purified RNA for each biological replicate, containing from 5–11 µg of RNA in a maximum volume of 100 µL, was submitted to the Canada’s Michael Smith Genome Sciences Centre (Vancouver, BC, Canada) for whole transcriptome shotgun sequencing.
RNA-seq method
Paired-end Illumina HiSeq 2000 platform sequencing was performed as described in Robert et al. (2013). 50 bp sequence reads were requested, although 75 bp reads were generated for some of the sequencing lanes because of advancing sequencing technologies. The 16 samples were multiplexed into four sequencing lanes, with one replicate of each treatment randomly assigned per lane so that every lane contained all four sampling dates. The paired-end fastq files for each library were mapped to the 13,088 gene models identified in the male mountain pine beetle genome sequence (Keeling et al., 2013) via CLC Genomics Workbench (CLC bio: http://www.clcbio.com/) using the parameters listed in Robert et al. (2013). The average fragment length for paired-end reads was approximately 220bp. Read counts are used as a measure of transcript expression for a given gene model in each sample.
The raw RNA-seq data is available at the National Center for Biotechnology Information Sequence Read Archive (NCBI SRA) database (accession numbers: SRX1043704–SRX1043719) under the TRIA umbrella BioProject (PRJNA169907) that identifies aggregated research project data generated from the TRIA research collaborations on mountain pine beetle systems genomics.
DESeq statistical analysis based on Anders & Huber (2010) was also performed as described in Robert et al. (2013) to calculate fold-changes for each gene as well as p-values adjusted (padj) for multiple comparisons using a Benjamini–Hochberg correction (Benjamini & Hochberg, 1995). We used a 0.1% false discovery rate (padj < 0.001) to identify transcript expression levels that were significantly different between timepoints. The DESeq output is publicly available via a DOI-based repository and can be found in ‘Data Availability’.
Results
Sequencing results
Sixteen libraries were sequenced with four biological replicates at each time point (Table 1). Overall, 581 transcripts (representing 4.43% of the 13,088 gene models, or predicted genes, identified in the draft mountain pine beetle genome) with significant shifts in expression occurred uniquely between the time points in September and November (Autumn) and 534 (representing 4.08% of the gene models identified in the draft genome) occurred uniquely between the time points in March to May (Spring); 145 transcripts (representing 1.12% of the gene models in the draft genome) showed significantly shifts in both the fall and winter (Fig. 1). Of the shared 145 transcripts, 107 increased in autumn and decreased in the spring whereas 29 transcripts showed the opposite pattern (i.e., decreased in autumn and increased in spring). The remaining nine significantly changing transcripts either both increased in autumn and spring, or both decreased in autumn and spring.
Table 1. Summary information for sequence data mapped to the male mountain pine beetle genome (Keeling et al., 2013).
| Sampling date | Replicate | NCBI accession number | Read length | Total pairs mapped | Uniquely mapped pairs |
|---|---|---|---|---|---|
| Sept. 26, 2008 | 1 | SRX1043704 | 50 | 51,228,868 | 20,383,049 |
| 2 | SRX1043705 | 50 | 75,540,024 | 29,715,613 | |
| 3 | SRX1043706 | 75 | 66,662,746 | 26,832,203 | |
| 4 | SRX1043707 | 75 | 41,552,862 | 16,802,040 | |
| Nov. 7, 2008 | 1 | SRX1043708 | 50 | 64,102,946 | 25,410,259 |
| 2 | SRX1043709 | 50 | 52,214,600 | 20,525,750 | |
| 3 | SRX1043710 | 75 | 36,280,228 | 14,402,268 | |
| 4 | SRX1043711 | 75 | 71,571,952 | 28,268,130 | |
| Mar. 25, 2009 | 1 | SRX1043712 | 50 | 59,279,874 | 23,425,001 |
| 2 | SRX1043713 | 50 | 54,870,752 | 21,496,616 | |
| 3 | SRX1043714 | 75 | 58,248,050 | 23,687,547 | |
| 4 | SRX1043715 | 75 | 67,215,526 | 26,666,241 | |
| May 27, 2009 | 1 | SRX1043716 | 50 | 55,731,196 | 21,959,708 |
| 2 | SRX1043717 | 50 | 43,999,520 | 17,182,971 | |
| 3 | SRX1043718 | 75 | 73,662,736 | 29,422,596 | |
| 4 | SRX1043719 | 75 | 51,860,004 | 20,696,257 |
Figure 1. Venn diagram of total transcriptome.
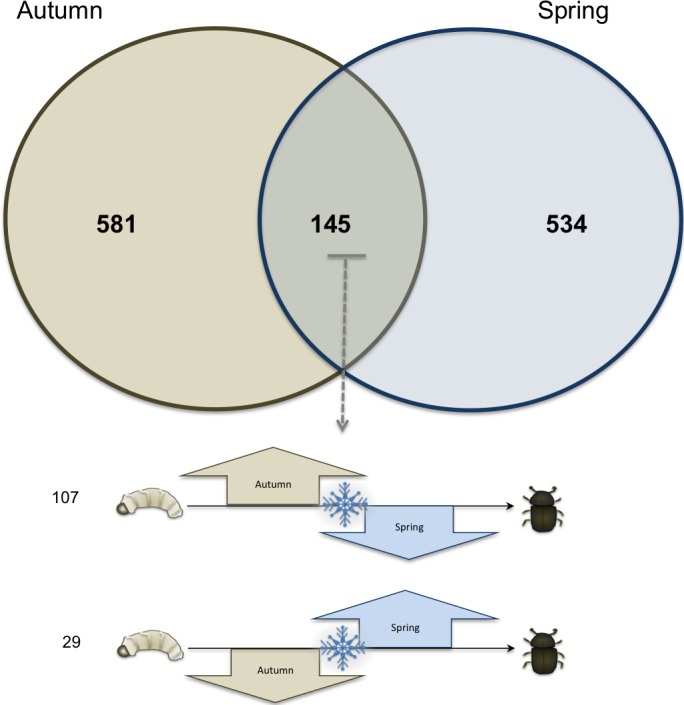
Venn diagram showing the number of significantly changing transcripts unique to autumn (brown circle), and unique to spring (blue circle), and those that showed significantly shifts in both autumn and spring (middle). Of the 145 gene transcripts that showed a significant difference in both autumn and spring, 107 increased in autumn and decreased in spring, and 29 are decreased in autumn and then increased in spring.
Transcriptome changes during autumn (September to November)
Between September and November, 726 transcripts showed significantly different expression levels (Bonferroni adjusted p-value < 0.001). Of these 726, 458 were significantly up-regulated (ranging from 1.6-fold increase to >100-fold increase) and 268 were significantly down-regulated (ranging from a 1.7-fold decrease to >300-fold decrease).
The largest fold changes that occurred between sampling dates in September and in November were gene transcripts annotated in the broad group of stress response physiology. The top expression fold-change was annotated as a putative galactose-specific C-type lectin (102.75-fold increase), an enzyme associated with the immune response in insects (e.g., Yu et al., 2005). Of the 11 transcripts that were significantly up-regulated over 10-fold between September and November (Table 2): four were of unknown function and were associated with physiological stress arising from insect contact with pathogens and host plant defenses as well as general cellular stabilization responses in response to stress. These six transcripts were annotated as a putative alpha-esterase, glycoside hydrolase family protein 48, cyclin-dependent kinase 5 activator, two heat shock proteins, and a glutathione S-transferase.
Table 2. Positive fold changes greater than 10.
Differentially expressed transcripts (identified by the gene model ID in the mountain pine beetle draft genome (Keeling et al., 2013) with positive fold changes >10 between September and November, and their annotations.
| NCBI accession | Fold change | Adjusted p-value | Annotation |
|---|---|---|---|
| KB740939.1 | 102.7 | 1.19 × 10−6 | galactose-specific C-type lectin (Aedes aegypti) |
| KB740085.1 | 85.9 | 1.18 × 10−48 | hypothetical protein (Tribolium castaneum) |
| KB740839.1 | 19.1 | 6.43 × 10−13 | alpha-esterase (Tribolium castaneum) |
| KB740648.1 | 15.8 | 4.46 × 10−12 | hypothetical protein (Tribolium castaneum) |
| KB740914.1 | 14.2 | 0.00052 | glycoside hydrolase family protein 48 (Dendroctonus ponderosae) |
| KB740085.1 | 14.1 | 3.2 × 10−08 | GF10288 (Drosophila ananassae) |
| KB740939.1 | 12.7 | 1.03 × 10−5 | cyclin-dependent kinase 5 activator (Tribolium castaneum) |
| KB741213.1 | 11.9 | 2.25 × 10−8 | No hits |
| KB740969.1 | 11.9 | 1.19 × 10−21 | heat shock protein 68 (Tribolium castaneum) |
| KB740969.1 | 11.1 | 5.65 × 10−50 | heat shock protein 68 (Tribolium castaneum) |
| KB741231.1 | 10.8 | 2.69 × 10−7 | putative glutathione s-transferase (Tribolium castaneum) |
Conversely, of the 41 transcripts that showed the largest (>10-fold) down-regulation between September and November, almost half (19) have no known function, a further nine were annotated as insect cuticle proteins, and the remaining 13 transcripts were variously annotated, but contain a number of catabolic enzymes such as proteases and lyases (Table 3).
Table 3. Negative fold changes greater than 10.
Differentially expressed transcripts (identified by the gene model ID in the mountain pine beetle draft genome (Keeling et al., 2013) with negative fold changes >10 between September and November, and their annotations.
| NCBI accession | Fold change | Adjusted p-value | Annotation |
|---|---|---|---|
| KB741103.1 | –10.18 | 0.00022 | Pupal cuticle protein |
| KB741156.1 | –10.20 | 8.08 × 10−05 | CLIP-domain serine protease (Tribolium castaneum) |
| KB740994.1 | –11.16 | 6.50 × 10−8 | hypothetical protein (Tribolium castaneum) |
| KB741213.1 | –11.30 | 7.35 × 10−20 | No hits |
| KB741217.1 | –11.71 | 0.00048 | cytochrome P450 (Tribolium castaneum) |
| KB740848.1 | –12.02 | 4.45 × 10−5 | cuticle protein (Tribolium castaneum) |
| KB741159.1 | –12.41 | 0.00073 | No hits |
| KB741077.1 | –12.48 | 5.14 × 10−9 | No hits |
| KB740085.1 | –12.71 | 4.51 × 10−5 | hypothetical protein (Tribolium castaneum) |
| KB740993.1 | –13.09 | 4.45 × 10−5 | cuticle protein (Tribolium castaneum) |
| KB741103.1 | –13.52 | 3.47 × 10−15 | No hits |
| KB740939.1 | –13.67 | 7.13 × 10−5 | PREDICTED: similar to pol (Hydra magnipapillata) |
| KB740540.1 | –13.73 | 3.88 × 10−15 | hypothetical protein (Tribolium castaneum) |
| KB740970.1 | –13.95 | 1.61 × 10−18 | polysaccharide lyase (Dendroctonus ponderosae) |
| KB740193.1 | –14.26 | 1.83 × 10−12 | nicotinic acetylcholine receptor (Tribolium castaneum) |
| KB740085.1 | –14.29 | 3.34 × 10−8 | hypothetical protein (Tribolium castaneum) |
| KB739553.1 | –14.98 | 8.58 × 10−5 | acid phosphatase (Tribolium castaneum) |
| KB740193.1 | –15.16 | 9.54 × 10−7 | nicotinic acetylcholine receptor (Musca domestica) |
| KB741019.1 | –16.07 | 2.20 × 10−16 | No hits |
| KB741019.1 | –16.85 | 1.07 × 10−12 | No hits |
| KB741269.1 | –17.96 | 1.57 × 10−5 | hypothetical protein (Tribolium castaneum) |
| KB740727.1 | –18.73 | 0.00022 | glucose dehydrogenase (Tribolium castaneum) |
| KB741062.1 | –20.00 | 1.78 × 10−73 | endopolygalacturonase ( Dendroctonus ponderosae) |
| KB740972.1 | –22.23 | 0.00064 | ectodermal protein (Tribolium castaneum) |
| KB741177.1 | –23.82 | 1.02 × 10−9 | hypothetical protein (Tribolium castaneum) |
| KB741019.1 | –26.06 | 4.58 × 10−6 | No hits |
| KB741018.1 | –26.81 | 0.00057 | cuticle protein (Culex quinquefasciatus) |
| KB741067.1 | –31.69 | 1.25 × 10−6 | larval cuticle protein (Apriona germari) |
| KB741207.1 | –32.00 | 0.00012 | Hypothetical protein (Tribolium castaneum) |
| KB740798.1 | –33.75 | 0.00026 | hypothetical protein (Tribolium castaneum) |
| KB740972.1 | –34.89 | 2.75 × 10−11 | No hits |
| KB740425.1 | –36.77 | 0.00037 | Hypothetical protein (Tribolium castaneum) |
| KB740686.1 | –55.12 | 0.00056 | Hypothetical protein (Drosophila pseudoobscura pseudoobscura) |
| KB740085.1 | –70.29 | 0.00014 | cuticular protein (Tribolium castaneum) |
| KB740880.1 | –124.19 | 9.00 × 10−5 | hypothetical protein (Tribolium castaneum) |
| KB738495.1 | –157.19 | 1.89 × 10−5 | cuticular protein (Tribolium castaneum) |
| KB740540.1 | –181.29 | 0.00093 | hypothetical protein (Tribolium castaneum) |
| KB740085.1 | –206.30 | 8.84 × 10−5 | cuticular protein (Tribolium castaneum) |
| KB740966.1 | –298.81 | 1.06 × 10−7 | Hypothetical protein (Drosophila erecta) |
| KB736863.1 | –301.59 | 0.00044 | No hits |
| KB740984.1 | –313.04 | 9.44 × 10−7 | cuticle protein (Tribolium castaneum) |
Transcript changes during spring (March to May)
Between the sampling points in March and May, 679 gene transcripts showed significant differences (Bonferroni adjusted p-value < 0.001). The 456 transcripts significantly up-regulated between March and May, as a group, had very different annotations than the transcript changes associated with the autumn period between September and November. The top six transcripts, in terms of fold-changes, between March and May all exhibited well over a 100 fold increase. Of these six transcripts, the greatest up-regulation was >1000-fold increase in a transcript annotated as a cathepsin L-like proteinase, a digestive enzyme in beetles (Murdock et al., 1987). The second highest fold change (202-fold up-regulation) is annotated as a pol (polyp-specific) gene similar to a sequence expressed in the polyp stage of the freshwater cnidarian, Hydra magnipapillata. The remaining transcripts that exhibited >100-fold up-regulation were all without functional annotations in current databases.
Patterns in transcript changes during overwintering (September to May)
Of the transcripts that exhibited increased levels between September and November, 107 also decreased significantly between March and May. Many of these were annotated as genes responsible for cold tolerance and as indicators of physiological response to stress. Transcripts associated with cold tolerance included those for two enzymes—glycerol-3-phosphate dehydrogenase and glycerol kinase—that immediately precede the production of glycerol. Those two were also significantly up-regulated in the autumn, stable over the winter, and significantly down-regulated in the spring (Fig. 2).
Figure 2. Glycerol biosynthesis.

Fold changes between each sampling date through the winter for transcripts annotated as key enzymes in the biosynthetic pathway for glycerol: glycerol-3-phosphate dehydrogenase (G3PDH, KB741112.1) and glycerol kinase (GK, KB740969.1). An asterisk indicates a significant fold change within the two time points measured within each sampling period (Bonferroni adjusted p-value < 0.001). The diagram on the right illustrates a subset of the glycerol biosynthetic pathway (modified with permission from Fraser, 2011) showing the location G3PDH and GK as the enzymes that catalyze the steps immediately preceding the production of glycerol. Metabolites are shown in ovals: dihydroxyacetone phosphate (DHAP), glycerol-3-phosphate (G3P), glyceraldehyde (GA); enzymes are shown in rectangles: glycerol-3-phosphate dehydrogenase (G3PDH), glycerol kinase (GK), alcohol dehydrogenase (ADH).
Other transcripts annotated as proteins that play a role in cold tolerance included those for a trehalose receptor that increased 14-fold, and for several heat shock proteins. For example, two heat shock protein transcripts and a heat shock transcription factor transcript that increased between September and November, significantly decreased during the spring (Fig. 3). A significant change in glutathione-S-transferase (Fig. 4) and cytochromes P450 (Table 4) also occur for larvae in the autumn and spring.
Figure 3. Heat shock proteins.
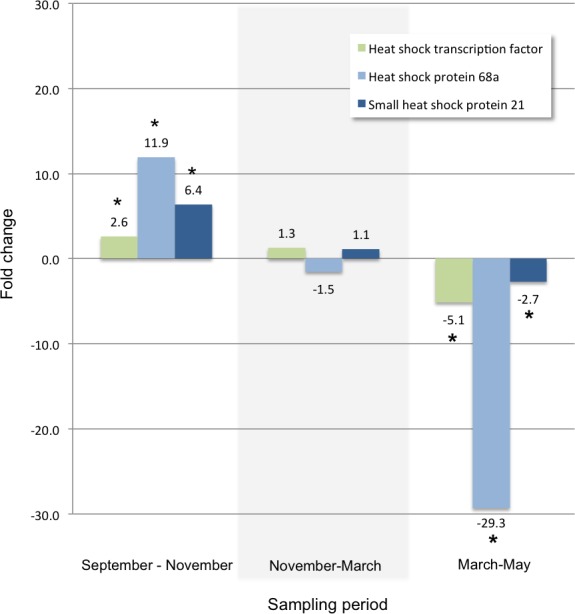
Fold changes between each sampling date through the winter for transcripts annotated with: a heat shock transcription factor (KB740969.1), heat shock protein 68a (KB740941.1), and small heat shock protein 21 (KB740193.1). An asterisk indicates a significant fold change between the time points within each sampling period (Bonferroni adjusted p-value < 0.001).
Figure 4. Glutathione-S-transferase.

Fold changes between each sampling date through the winter for transcripts annotated as glutathione-S-transferase 1 (KB741231.1) and glutathione-S-transferase 2 (KB740293.1). An asterisk indicates a significant fold change between the time points within each sampling period (Bonferroni adjusted p-value < 0.001).
Table 4. P450 table.
Fold changes and adjusted p-values for cytochromes P450 that increased in the autumn sampling period (transcript expression levels between September and November) and then either decreased or did not change through the winter (November and March) and spring (March and May) sampling periods. An asterisk indicates a significant difference (Bonferroni adjusted p-value < 0.001) between time points within the sampling period. Shaded rows indicate transcripts that increased significantly in the autumn and decreased significantly the in the spring.

|
Autumn |

|
Winter |

|
Spring | |||
|---|---|---|---|---|---|---|---|---|
| NCBI accession | CYP ID | Fold change | Adjusted p-value | Fold change | Adjusted p-value | Fold change | Adjusted p-value | |
| KB741217.1 | CYP6CR2 | 8.01* | <0.0001 | –2.79 | 0.0055 | 1.25 | 0.2165 | |
| KB741284.1 | CYP345E3 | 4.86* | <0.0001 | –1.85 | 0.1206 | –4.15* | <0.0001 | |
| KB740988.1 | CYP4G56 | 3.89* | <0.0001 | 1.00 | 1.0000 | –0.60 | 0.3440 | |
| KB739995.1 | CYP4EX1 | 3.29* | 0.0001 | –1.76 | 0.1901 | 0.04 | 0.9668 | |
| KB737912.1 | CYP6BW2 | 3.16* | <0.0001 | –3.98* | <0.0001 | 1.27 | 0.0678 | |
| KB740970.1 | CYP9AP1 | 3.03* | <0.0001 | –2.37 | 0.0016 | –0.48 | 0.4740 | |
| KB740940.1 | CYP4CV1 | 2.43* | 0.0003 | –1.13 | 0.9780 | –0.72 | 0.5954 | |
| KB740092.1 | CYP349B1 | 2.37* | <0.0001 | –1.48 | 0.1789 | –5.34* | <0.0001 | |
| KB741266.1 | CYP6DF1 | 2.22* | <0.0001 | –1.42 | 0.2963 | –0.42 | 0.5217 | |
| KB740988.1 | CYP9Z36 | 2.11* | 0.0010 | –3.66* | <0.0001 | –2.00* | 0.0004 | |
| KB741292.1 | CYP6BS2 | 1.99* | 0.0004 | –1.81 | 0.0170 | 1.00 | 0.0543 | |
Discussion
As mountain pine beetle larvae grow and develop within the infested host tree, they are exposed during the autumn and winter to decreasing temperatures, conifer host chemical defenses (Bohlmann, 2012; Trapp & Croteau, 2001; Keeling & Bohlmann, 2006a.; Keeling & Bohlmann, 2006b), and pathogens (Winder, Macey & Cortese, 2010). In addition, overwintering mountain pine beetle larvae may be exposed to other possible changes in their environment such as nutrient and moisture content of host tissue or changing microbiomes of the host tissue and the feeding galleries. Many of the major fold changes in the larvae transcriptome observed between September and November occur in a group of transcripts that can be broadly grouped as “stress response genes.” Because this is a period in which mountain pine beetle larvae have shown the highest mortality (Bale, 2002), the general working hypothesis is that larvae die if they are not fully acclimated, including the production anti-freeze compounds. Our results support this, but additionally suggest that early winter larvae are under enormous physiological stress from other factors as well, such as, for example, host chemical defenses and pathogens. The autumn transcriptome signature is consistent with proteomic evidence of an increase in oxidative stress in overwintering larvae between September and November (Bonnett et al., 2012). Within this response however, it is difficult to separate the response to a seasonal drop in temperature from the other environmental pressures occurring at this time. The diversity of enzyme groups and pathways that are up-regulated between September and November indicates a multi-pronged approach to cold hardiness and overwintering. We identified several groups of genes likely to be associated with one or more “stress” factors: cold hardiness (heat shock proteins, glycerol biosynthesis, sugars and animo acids), immune-responsive proteins, and detoxification mechanisms.
Between September and November, two heat shock protein (Hsp) transcripts are strongly up-regulated (annotated as similar to Hsp68 in Tribolium cataneum), an additional two transcripts are significantly up-regulated to a lesser extent (one 90kDa protein and one small heat shock protein 21), and a heat shock protein transcription factor transcript is significantly up-regulated as well. Heat shock proteins are associated with abiotic stresses such as temperature change, drought, dehydration, and chemical exposure (King & MacRae, 2015; Zhao & Jones, 2012)—all of which occur during the early months of larval growth and development under the bark of the host tree. These transcripts likely play a role in protein stabilization or sequestration in preparation for below-freezing temperatures during the winter months. Similar expression of heat shock proteins has been shown in Drosophila melanogaster, specifically expression of Hsp68 was induced by acute cold stress. (Colinet & Hoffmann, 2012; Colinet, Lee & Hoffmann, 2010) and in leafminer species (Huang & Kang, 2007; Huang, Wang & Kang, 2009). The expression pattern of a putative HSP transcription factor, an Hsp 68a, and a small Hsp 21 (a large increase in the early winter, stable transcription throughout the winter months, then a large decrease during the onset of spring) (Fig. 3), identifies these gene candidates for further study.
Glycerol is a known cryoprotectant in mountain pine beetle and is one of the few metabolites that have been shown to accumulate in overwintering larvae (Bentz & Mullins, 1999). Our results support the importance of glycerol production in early winter and support similar quantitative RT-PCR results found in overwintering larvae in Fraser (2011) and J Fraser (2010, unpublished data); studies that were conducted using overwintering larvae collected concurrently with the larvae sampled for this study. Key enzymes in the two catalytic steps preceding the final synthesis of glycerol were significantly up-regulated between September and November (Fig. 2). This suggests that production of glycerol is increased between September and November, in agreement with previous work (Fraser, 2011). Also in agreement with Fraser (2011), our results further suggest that glycerol is biosynthesized via dihydroxyacetone phosphate (DHAP) and glycerol-3-phosphate (G3P) rather than via glyceraldehyde (Fig. 2). Although the glyceraldehyde route could conceivably also be responsible for increased glycerol synthesis in some circumstances, we did not find evidence for this pathway in our data set, which supports the findings in Fraser (2011) as well.
Although a protein identified as trehalose-phosphate synthase was up-regulated in the autumn and down-regulated in the spring in the proteomic data reported by Bonnett et al. (2012), only a single gene transcript in our data set was annotated as a trehalose 6-phosphate synthase isoform. The expression of this transcript did not change significantly in the autumn, through the winter, or in the spring. Rozsypal et al. (2013) also found that levels of trehalose and proline remain at high, constant levels for overwintering codling moth larvae and suggest that these metabolites are important for protein and membrane stabilization during bouts of freezing temperatures as described in Jain & Roy (2009). Our data suggests a similar pattern for mountain pine beetle overwintering larvae.
In addition to preparation for sub-freezing temperatures, mountain pine beetle larvae also showed a stress response in the form of the expression of immune response transcripts. The largest significant transcript increase between September and November was annotated as a galactose-specific C-type lectin. In Drosophila, these enzymes are associated with immune response, but the precise role of specific proteins in this large group remains unclear (Tanji, Ohashi-Kobayashi & Natori, 2006). We identified 11 gene models in the mountain pine beetle genome annotated as ‘lectins’; two of these were highly up-regulated in larvae between September and November. This could represent either a local response to a particular pathogen, or a general stress-induced response as a result of toxic conditions of a colonized host tree.
In autumn, several genes associated with detoxification of host metabolites are also strongly up-regulated. Glutathione S-transferases and an alpha-esterases are two enzyme groups that have been implicated with detoxification of xenobiotics, such as host defense compounds, in insects (Pitt et al., 2014; Robert et al., 2013; Campbell et al., 2003; Kostaropoulos et al., 2001). As insect larvae are still living and growing in a dying tree, they would be exposed to not only decreasing temperatures, but also to the full suite of conifer host chemical defenses (Clark, Huber & Carroll, 2012; Bohlmann, 2012; Keeling & Bohlmann, 2006a.; Keeling & Bohlmann, 2006b). A large group of enzymes commonly associated with detoxification of plant defense compounds, cytochromes P450 (Feyereisen, 1999), are also significantly up-regulated in our data set during autumn. Eleven gene transcripts annotated as cytochromes P450 increase significantly in the autumn. Of these 11, three also significantly decrease in the spring (CYP345E3, CYP349B1, and CYP9Z36). These enzymes would be strong candidates for a role in larval detoxification of host tissue. CYP345E3 is similar in sequence to CYP345E2, which modifies monoterpene substrates (Keeling et al., 2013). Expressed sequence tag data suggests CYP345E3 is not localized to the antenna like CYP345E2, but found in the midgut and fat body tissues of adults (Keeling et al., 2016), also suggesting a role in detoxification. Interestingly, several of the cytochromes P450 that increased significantly in larvae during autumn (CYP6CR2, CYP4EX1, CYP9AP1, and CYP6BW2) comprise almost the entire list of significantly decreasing cytochrome P450s in starved versus fed adult mountain pine beetle (Robert et al., 2013). Because these transcripts do not decrease in spring, are elevated in antennae-rich mountain pine beetle expressed sequence tag libraries (Keeling et al., 2012), and implicated in odourant degradation processes in adult cotton leafworms (Pottier et al., 2012), it may be possible that the development of odour processing begins as early as the formation of eye-antennal imaginal discs in early instar larvae (Haynie & Bryant, 2005). In addition, CYP6CR2 has high sequence similarity to CYP6CR1, which is an epoxidase in exo-brevicomin biosynthesis, a pheromone produced by male mountain pine beetle after leaving the brood tree, but that decreases during host tree selection and mating (Song et al., 2014). Theses cytochromes P450 could also play a role as the developing larvae navigate and feed on host tissue containing a diversity of associated fungi that can act as a food source, or threat, under the bark (Lee, Kim & Breuil, 2006). The opposite process occurs in the adults, as after host colonization, odorant detection and processing becomes irrelevant as the focus shifts to mating and reproduction (Robert et al., 2013).
In spring, larvae must begin the physiological process of preparing for emergence, dispersal flight, and a chance of reproduction during the subsequent summer months as well as a number of unidentified processes that require further study. For larvae that survived the winter, the late-instar preparation for pupation shows decreased levels of stress response gene transcripts and increases in developmental gene transcripts generally. The various transcripts showing greater than 100-fold increases March and May are largely without known functional annotation in current databases. Developmental processes of overwintering beetles in the spring thus seem to represent a largely unknown set of processes that need further investigation. The function of the pol in Hydra spp. polyps is unknown, but possible functions include control of osmotic pressure, secretion of this protein to form an outer protective layer for the polyp, secretion as a component of the extracellular matrix, secretion as an anti-bacterial defense, or secretion as a signaling molecule (Aerne et al., 1996). The distant phylogenetic relationship of cnidarians to beetles make it difficult to predict potential function. Because this transcript is strongly and significantly up-regulated (greater than 200-fold increase in expression) between March and May, the protein product could play a significant role in bark beetle late-stage developmental processes.
Conclusions
We suggest a multipronged physiological response from larvae in a toxic and cold environment during autumn. This response can be broadly characterized by gene transcripts involved in insect immune responses and detoxification, which are strongly up-regulated in the autumn and occurs within a background of preparation for winter via the production of glycerol, heat shock proteins, trehalose and other known strategies to tolerant below-freezing temperatures. In the spring, although gene transcripts associated with developmental process are present, the bulk of the transcripts are not yet associated with known gene functions.
Funding Statement
This work was supported by the following: Natural Science and Engineering Research Council of Canada (NSERC) Postdoctoral Fellowship (http://www. nserc-crsng.gc.ca/index_eng.asp); Genome Canada (http://www.genomecanada.ca); Genome British Columbia (http://www.genomebc.ca); Genome Alberta (http://genomealberta.ca); Canada Research Chairs (CRC) Program (http://www.chairs-chaires.gc.ca); Canada Foundation for Innovation (CFI) (http://www.innovation.ca); and British Columbia Knowledge Development Fund (BCKDF) (http://www.aved.gov.bc.ca/research andinnovation/Funding/BCKDF/). We thank collaborators within the Tria 1 and Tria 2 projects (http://www.thetriaproject.ca), a study of Mountain Pine Beetle System Genomics. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
Dezene P.W. Huber is an Academic Editor for PeerJ.
Author Contributions
Jeanne A. Robert conceived and designed the experiments, performed the experiments, analyzed the data, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Tiffany Bonnett conceived and designed the experiments, performed the experiments, analyzed the data, reviewed drafts of the paper.
Caitlin Pitt conceived and designed the experiments, performed the experiments, reviewed drafts of the paper.
Luke J. Spooner, Macaire M.S. Yuen and Christopher I. Keeling analyzed the data, reviewed drafts of the paper.
Jordie Fraser conceived and designed the experiments, reviewed drafts of the paper.
Jörg Bohlmann contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Dezene P.W. Huber conceived and designed the experiments, contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
DNA Deposition
The following information was supplied regarding the deposition of DNA sequences:
The raw RNA-seq data is available at the National Center for Biotechnology Information Sequence Read Archive (NCBI SRA) database (accession numbers: SRX1043704–SRX1043719) under the TRIA umbrella BioProject (PRJNA169907) that identifies aggregated research project data generated from the TRIA research collaborations on mountain pine beetle systems genomics.
Data Availability
The following information was supplied regarding data availability:
The DESeq output is publicly available via Figshare accessible from the following links:
Sept–Nov
DOI: 10.6084/m9.figshare.815969
URL: https://figshare.com/s/1031731941b18df7121a
Sept–May
DOI: 10.6084/m9.figshare.815968
URL: https://figshare.com/s/5c3490cb9f906604b0cd
Sept–March
DOI: 10.6084/m9.figshare.815967
URL: https://figshare.com/s/4d0d34c9445a6bc52422
March–May
References
- Aerne et al. (1996).Aerne B, Gröger H, Schuchert P, Spring J, Volker S. The polyp and its medusa: a molecular approach. Scientia Marina. 1996;60(1):7–16. [Google Scholar]
- Anders & Huber (2010).Anders S, Huber W. Differential expression analysis for sequence count data. Genome Biology. 2010;11(10):R106. doi: 10.1186/gb-2010-11-10-r106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bale (2002).Bale JS. Insects and low temperatures: from molecular biology to distributions and abundance. Philosophical Transactions of the Royal Society B: Biological Sciences. 2002;357(1423):849–862. doi: 10.1098/rstb.2002.1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamini & Hochberg (1995).Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. Series B (Methodological) 1995;57(1):289–300. [Google Scholar]
- Bentz & Mullins (1999).Bentz BJ, Mullins DE. Ecology of Mountain pine beetle (coleoptera: scolytidae) cold hardening in the intermountain west. Environmental Entomology. 1999:577–587. doi: 10.1093/ee/28.4.577. [DOI] [Google Scholar]
- Bohlmann (2012).Bohlmann J. Pine terpenoid defences in the mountain pine beetle epidemic and in other conifer pest interactions: specialized enemies are eating holes into a diverse, dynamic and durable defence system. Tree Physiology. 2012;32(8):943–945. doi: 10.1093/treephys/tps065. [DOI] [PubMed] [Google Scholar]
- Bonnett et al. (2012).Bonnett TR, Robert JA, Pitt C, Fraser J, Keeling CI, Bohlmann J, Huber D. Global and comparative proteomic profiling of overwintering and developing mountain pine beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae), larvae. Insect Biochemistry and Molecular Biology. 2012;42(12):890–901. doi: 10.1016/j.ibmb.2012.08.003. [DOI] [PubMed] [Google Scholar]
- Campbell et al. (2003).Campbell PM, Robin GCDeQ, Court LN, Dorrian SJ, Russell RJ, Oakeshott JG. Developmental expression and gene/enzyme identifications in the alpha esterase gene cluster of Drosophila melanogaster. Insect Molecular Biology. 2003;12(5):459–471. doi: 10.1046/j.1365-2583.2003.00430.x. [DOI] [PubMed] [Google Scholar]
- Clark, Huber & Carroll (2012).Clark EL, Huber DPW, Carroll AL. The legacy of attack: implications of high phloem resin monoterpene levels in lodgepole pines following mass attack by mountain pine beetle, Dendroctonus ponderosae Hopkins. Environmental Entomology. 2012;41:392–398. doi: 10.1603/EN11295. [DOI] [PubMed] [Google Scholar]
- Cole (1981).Cole W. Some risks and causes of mortality in mountain pine beetle populations: a long-term analysis. Researches on Population Ecology. 1981;23:116–144. doi: 10.1007/BF02514096. [DOI] [Google Scholar]
- Colinet & Hoffmann (2012).Colinet H, Hoffmann AA. Comparing phenotypic effects and molecular correlates of developmental, gradual and rapid cold acclimation responses in Drosophila melanogaster. Functional Ecology. 2012;26:84–93. doi: 10.1111/j.1365-2435.2011.01898.x. [DOI] [Google Scholar]
- Colinet, Lee & Hoffmann (2010).Colinet H, Lee SF, Hoffmann A. Temporal expression of heat shock genes during cold stress and recovery from chill coma in adult Drosophila melanogaster. FEBSJ. 2010;277:174-185. doi: 10.1111/j.1742-4658.2009.07470.x. [DOI] [PubMed] [Google Scholar]
- Cullingham et al. (2011).Cullingham CI, Cooke JE, Dang S, Davis CS, Cooke BJ, Coltman DW. Mountain pine beetle host-range expansion threatens the boreal forest. Molecular Ecology. 2011;20(10):2157–2171. doi: 10.1111/j.1365-294X.2011.05086.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feyereisen (1999).Feyereisen R. Insect p450 enzymes. Annual Review of Entomology. 1999;44:507–533. doi: 10.1146/annurev.ento.44.1.507. [DOI] [PubMed] [Google Scholar]
- Fraser (2011).Fraser J. MSc thesis. 2011. Cold tolerance and seasonal gene expression in Dendroctonus ponderosae. [Google Scholar]
- Haynie & Bryant (2005).Haynie JL, Bryant PJ. Development of the eye-antenna imaginal disc and morphofenesis of the adult head in drosophila melanogaster. Journal of Experimental Zoology. 2005;237(3):293–308. doi: 10.1002/jez.1402370302. [DOI] [PubMed] [Google Scholar]
- Huang & Kang (2007).Huang L-H, Kang L. Cloning and interspecific altered expression of heat shock protein genes in two leafminer species in response to thermal stress. Insect Molecular Biology. 2007;16:491–500. doi: 10.1111/j.1365-2583.2007.00744.x. [DOI] [PubMed] [Google Scholar]
- Huang, Wang & Kang (2009).Huang L-H, Wang CZ, Kang L. Cloning and expression of five heat shock protein genes in relation to cold hardening and development in the leafminer, Liriomyza sativa. Journal of Insect Physiology. 2009;55:279–285. doi: 10.1016/j.jinsphys.2008.12.004. [DOI] [PubMed] [Google Scholar]
- Jain & Roy (2009).Jain NK, Roy I. Effect of trehalose on protein structure. Protein Science. 2009;18(1):24–36. doi: 10.1002/pro.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janes et al. (2014).Janes JK, Yisu L, Keeling CI, Yuen M, Boone CK, Cooke JEK, Bohlmann J, Huber DPW, Murray BW, Coltman DW, Sperling FAH. How the mountain pine beetle (Dendroctonus ponderosae) breached the Canadian Rocky Mountains. Molecular Biology and Evolution. 2014;31(7):1803–1815. doi: 10.1093/molbev/msu135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keeling & Bohlmann (2006a).Keeling CI, Bohlmann J. Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defence of conifers against insects and pathogens. New Phytologist. 2006a;170:657–675. doi: 10.1111/j.1469-8137.2006.01716.x. [DOI] [PubMed] [Google Scholar]
- Keeling & Bohlmann (2006b).Keeling CI, Bohlmann J. Diterpene resin acids in conifers. Phytochemistry. 2006b;67:2415–2423. doi: 10.1016/j.phytochem.2006.08.019. [DOI] [PubMed] [Google Scholar]
- Keeling et al. (2012).Keeling CI, Henderson H, Li M, Yuen M, Clark EL, Fraser JD, Huber D, Liao NY, Docking TR, Birol I, Chan SK, Taylor GA, Palmquist D, Jones S, Bohlmann J. Transcriptome and full-length cDNA resources for the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major insect pest of pine forests. Insect Biochemistry and Molecular Biology. 2012;42(8):525–536. doi: 10.1016/j.ibmb.2012.03.010. [DOI] [PubMed] [Google Scholar]
- Keeling et al. (2016).Keeling CI, Li M, Sandhu HK, Henderson H, Yuen MMS, Bohlmann J. Quantitative metabolome, proteome, and transcriptome analysis of midgut and fat body tissues in the mountain pine beetle, Dendroctonus ponderosa Hopkins, and insights into pheromone biosynthesis. Insect Biochemistry and Molecular Biology. 2016;70:170–183. doi: 10.1016/j.ibmb.2016.01.002. [DOI] [PubMed] [Google Scholar]
- Keeling et al. (2013).Keeling CI, Yuen M, Liao NY, Docking TR, Chan SK, Taylor GA, Palmquist DL, Jackman SD, Nguyen A, Li M, Henderson H, Janes JK, Zhao Y, Pandoh P, Moore R, Sperling FA, Huber DP, Birol I, Jones SJ, Bohlmann J. Draft genome of the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major forest pest. Genome Biology. 2013;14(3) doi: 10.1042/BJ20051921. R27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King & MacRae (2015).King A, MacRae TH. Insect heat shock proteins during stress and diapause. Annual Review of Entomology. 2015;60:59–75. doi: 10.1146/annurev-ento-011613-162107. [DOI] [PubMed] [Google Scholar]
- Kostaropoulos et al. (2001).Kostaropoulos I, Papadopoulos AI, Metaxakis A, Boukouvala E, Papadopoulou-Mourkidou E. Glutathione S-transferase in the defence against pyrethroids in insects. Insect Biochemistry and Molecular Biology. 2001;31:313–319. doi: 10.1016/S0965-1748(00)00123-5. [DOI] [PubMed] [Google Scholar]
- Lee, Kim & Breuil (2006).Lee S, Kim JJ, Breuil C. Mountain pine beetle initiative working paper. Canadian Forest Service; Victoria: 2006. Diversity of fungi associated with mountain pine beetle, Dendroctonus ponderosae, and infested lodgepole pines in British Columbia. [Google Scholar]
- Murdock et al. (1987).Murdock LL, Brookhart G, Dunn PE, Foard DE, Kelley S, Kitch L, Shade RE, Shukle RH, Wolfson JL. Cysteine digestive proteinases in Coleoptera. Comparative Biochemistry and Physiology. Part B: Comparative Biochemistry. 1987;87(4):783–787. doi: 10.1016/0305-0491(87)90388-9. [DOI] [Google Scholar]
- Natural Resources Canada (2015).Natural Resources Canada . Mountain pine beetle. Ottawa: Natural Resources Canada; 2015. [October 2015]. [Google Scholar]
- Pitt et al. (2014).Pitt C, Robert JA, Bonnett TR, Keeling CI, Bohlmann J, Huber DPW. Proteomics indicators of rapidly shifting physiology from whole moutain pine beetle, dendroctonus ponderoase (Coleoptera: Curculionidae), adults during early host colonization. PLoS ONE. 2014;9(10):e2109. doi: 10.1371/journal.pone.0110673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pottier et al. (2012).Pottier M-A, Bozzolan F, Chertemps T, Jacquin-Joly E, Lalouette L, Siaussat D, Maïbèche-Coisne M. Cytochrome p450s and cytochrome p450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Molecular Biology. 2012;21:568-580. doi: 10.1111/j.1365-2583.2012.01160.x. [DOI] [PubMed] [Google Scholar]
- Raffa et al. (2008).Raffa KF, Aukema BH, Bentz BJ, Carroll AL, Hicke JA, Turner MG, Romme WH. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: dynamics of biome-wide bark beetle eruptions. BioScience. 2008;58(6):501–518. [Google Scholar]
- Robert et al. (2013).Robert JA, Pitt C, Bonnett TR, Yuen MMS, Keeling CI, Bohlmann J, Huber DPW. Disentangling detoxification: gene expression analysis of feeding mountain pine beetle illuminates molecular-level host chemical defense detoxification mechanisms. PLoS ONE. 2013;8(11):e2109. doi: 10.1371/journal.pone.0077777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozsypal et al. (2013).Rozsypal J, Kostal V, Sahradnickova H, Simek P. Overwintering strategy and mechanisms of cold tolerantce in the codling moth (cydia pomonella) PLoS ONE. 2013;8(4):e2109. doi: 10.1371/journal.pone.0061745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider et al. (2010).Schneider RR, Latham MC, Stelfox B, Farr D, Boutin S. Effects of a severe mountain pine beetle epidemic in western Alberta, Canada under two forest management scenarios. International Journal of Environmental Research. 2010 doi: 10.1155/2010/417595. Article 417595. [DOI] [Google Scholar]
- Song et al. (2014).Song M, Gorzalski A, Nguyen TT, Liu X, Jeffrey C, Blomquist GJ, Tittiger C. Exo-Brevicomin biosynthesis in the fat body of the mountain pine beetle, Dendroctonus ponderosae. Journal of Chemical Ecology. 2014;40:181-189. doi: 10.1007/s10886-014-0381-9. [DOI] [PubMed] [Google Scholar]
- Tanji, Ohashi-Kobayashi & Natori (2006).Tanji T, Ohashi-Kobayashi A, Natori S. Participation of a galactose-specific c-type lectin in drosophila immunity. Biochemistry Journal. 2006;396(pt 1):127–138. doi: 10.1042/BJ20051921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapp & Croteau (2001).Trapp SC, Croteau RB. Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics. 2001;158(2):811–832. doi: 10.1093/genetics/158.2.811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winder, Macey & Cortese (2010).Winder RS, Macey DE, Cortese J. Dominant bacteria associated with broods of mountain pine beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae, scolytinae) Journal of the Entomological Society of British Columbia. 2010;107:43–56. [Google Scholar]
- Yu et al. (2005).Yu X-Q, Tracy ME, Ling E, Scholz FR, Trenczek T. A novel c-type immulection-3 from Manduca sexta is translocated from hemolymph into the cytoplasm of hemocytes. Insect Biochemistry and Molecular Biology. 2005;35(4):285–295. doi: 10.1016/j.ibmb.2005.01.004. [DOI] [PubMed] [Google Scholar]
- Zhao & Jones (2012).Zhao L, Jones WA. Expression of heat shock protein genes in insect stress response. Invertebrate Survival Journal. 2012;9:93–101. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The following information was supplied regarding data availability:
The DESeq output is publicly available via Figshare accessible from the following links:
Sept–Nov
DOI: 10.6084/m9.figshare.815969
URL: https://figshare.com/s/1031731941b18df7121a
Sept–May
DOI: 10.6084/m9.figshare.815968
URL: https://figshare.com/s/5c3490cb9f906604b0cd
Sept–March
DOI: 10.6084/m9.figshare.815967
URL: https://figshare.com/s/4d0d34c9445a6bc52422
March–May