

Abstract
The skin is a barrier organ that interacts with the external environment. Being continuously exposed to potential microbial invasion, the dermis and epidermis home a variety of immune cells in both homeostatic and inflammatory conditions. Tools to obtain skin cell release for cytofluorimetric analyses are, therefore, very useful in order to study the complex network of immune cells residing in the skin and their response to microbial stimuli. Here, we describe an efficient methodology for the digestion of mouse skin to rapidly and efficiently obtain single-cell suspensions. This protocol allows maintenance of maximum cell viability without compromising surface antigen expression. We also describe how to take and digest skin samples from different anatomical locations, such as the ear, trunk, tail, and footpad. The obtained suspensions are then stained and analyzed by flow cytometry to discriminate between different leukocyte populations.
Keywords: Medicine, Issue 110, Leukocytes, immune system, skin, dermis, flow cytometry, single-cell preparation, collagenase
Introduction
The skin is one of the largest organs of the body and its large surface is continuously exposed to the external environment. Therefore the skin has to protect the organism from potential threats in order to maintain homeostasis, both by physical means, and by providing active protection toward potential pathogen entry. In a similar way to the gut and lung mucosa, the skin homes a variety of immune cells that continuously interact with the epithelium in order to maintain immune surveillance. This complex system that involves both immune and non-immune cells of the skin has been acknowledged since the early days of immunology, when in 1978 the term "Skin-associated lymphoid tissue" (SALT) 1 was first coined to describe such complexity.
The large number of immune cells residing in the skin helps the organism not only to fight potential invasions, but also to orchestrate wound healing and to maintain tolerance toward self-antigens and the skin microbiota 2,3.
Amongst skin resident immune cells, Dendritic Cells (DCs) have a crucial role in shaping immune responses and maintaining homeostasis 4. Dermal DCs and Langerhans cells readily respond to pathogen invasions and, upon migration to lymph nodes activate T cells and induce the expression of skin homing receptors on the newly generated effector T cells 5. DCs also have a fundamental role in regulating skin homeostasis. DCs migrating from the dermis to the lymph nodes in homeostatic conditions transport skin sequestered antigens with the purpose of inducing T cell tolerance mostly through the differentiation of regulatory T cells (Tregs) specific for skin antigens 6-8.
T cells represent the most abundant population of immune cells in the skin. Not only do they infiltrate the skin during an infection but they also represent a stable population among skin resident lymphocytes 9,10. Both CD8+ and CD4+ resident memory (rm)T cells reside in the skin and, following an infection, can respond long before effector T cells are recruited from the blood. In the skin, also resides a population of tissue resident Tregs capable of maintaining tolerance toward skin antigens and tissue homeostasis. These cells are rapidly and potently activated after exposure to their cognate antigen and have therefore been defined as memory Tregs 11,12.
Along with DCs and T cells, many innate immune cells, such as NK cells, gamma delta T cells, group 2 ILCs 13, Mast Cells and Macrophages, populate the skin and contribute to host protection. To analyze such a complex environment, it is mandatory to obtain single-cell suspensions from skin specimens with high efficiency while at the same time preserving the expression of surface markers for flow cytofluorimetric analysis or sorting.
Murine skin presents an outer epidermal layer, constituted mainly of keratinocytes, Langerhans cells and dendritic epidermal T cells; and the dermal layer beneath. The dermis homes most of the immune cells and is made by an extracellular matrix in which collagen fibers are the most abundant, especially collagen-I and collagen-IV. Unlike human skin, mouse skin is covered in fur, and thus more populated with hair follicles. It is, also, much thinner than human skin and contains a muscular layer, the panniculus carnosus, which helps wound healing 14. These characteristics render it more difficult to efficiently disrupt the collagen net of the dermis in order to get immune cells out.
In this paper we provide a suitable method for digesting the extracellular matrix that relies on a cocktail of high purity collagenase 1 and 2 and thermolysin. Moreover, since the analysis of skin from different regions requires different experimental procedures, we will show different methods to efficiently obtain skin samples from the footpad, tail, ear and trunk. We will then label the samples in order to evaluate the presence of DCs (MHCII+CD11c+CD45+) Macrophages (CD11b+F4/80+CD45+), CD8+, and CD4+ T cells.
Protocol
Study approval: The experimental protocols were approved by the Italian Ministry of Health (Rome, Italy) according to the Decreto legislativo 27 gennaio 1992, n. 116 "Attuazione della Direttiva n. 86/609/CEE in materia di protezione degli animali utilizzati a fini sperimentali o ad altri fini scientifici."
1. Obtaining Skin Samples
Euthanize the mouse by cervical dislocation.
- Ear:
- Cut out the hairless part of the ears of the mouse.
- Separate dorsal and ventral side by pulling them apart with forceps. Remove any remaining cartilages by gently scraping the inner part of the ears with forceps.
- Trunk skin:
- Shave the whole skin of the animal both with an electric razor and with delicate depilatory lotion.
- Cut off the desired skin sample (from 2 cm2 up to the whole animal skin).
- If cutting off the entire dorsal and ventral mouse skin, make a vertical cut in the middle of the mouse back and then cut along the borders of the shaved areas around the mouse body.
- Gently separate the skin from the peritoneum and back muscles, keeping the skin sample still with the forceps while separating the skin sample with the closed round edged tips of a surgical scissors or a scalpel.
- Prepare a 100 mm cell culture plate containing 10 ml of ice cold PBS.
- Eliminate subcutaneous fat by submerging the skin sample in PBS in the plate prepared in step 1.3.3 and scraping the skin sample with forceps.
- Tail:
- Cut off the tail of the mouse.
- Make a vertical cut on the tail with a surgical scissors or surgical scalpel, starting from the tail base and reaching to the tip of the tail.
- Use forceps to detach the skin from the tail. Grab the edge of the cut at the base of the tail and gently pull toward the tip while holding the body of the tail with forceps.
- Remove any excess fat by gently scraping the skin sample.
- Footpad
- Cut out the entire mouse foot with surgical scissors. Avoiding cutting any hairy skin.
- Cut the skin of the foot longitudinally from the ankle to the beginning of the digits.
- Hold the ankle bones with forceps or by hand while grabbing the skin with forceps and pull the skin toward the digits as if taking off a glove.
2. Digestion
Prepare the digestion cocktail from the stock solution (prepared as in Materials List) with the following recipe: Enzyme mix (see Materials List for specifications) 300 μg/ml, DNAse1 50 U/ml. Dilute in RPMI 5% FBS.
Chop skin samples with scissors into small pieces into a 35 mm Petri dish with 2 ml of Digestion Cocktail (For trunk skin: 2 ml of Digestion Cocktail every 2 cm2 of skin, up to 3 ml total).
Incubate at 37 °C for 90 min. Gently shake the Petri dish 1-2 times during incubation.
Pour digested skin samples on a 70 μm cell strainer in 66 mm Petri dish containing 1 ml of RPMI1640 5% FBS.
Wash the strainer with 5 ml of RPMI 1640 5% FBS.
Squeeze the sample through the cell strainer with a syringe plunger.
Flush plunger, Petri dish and strainer with 20 ml of RPMI1640 5% FBS, transfer into 50 ml tube.
3. Antibodies and Labeling
Wash the obtained suspension twice with 5 ml of PBS 1% FBS (or BSA).
- Label the obtained cell suspension with the desired antibodies.
- Resuspend the samples in 100 μl/sample of a mix containing 2 μg/ml of aCD16/CD32 (clone 2.4G2) diluted in PBS 1% FBS.
- Incubate for 15 min at 4 °C in order to block non-specific binding of the desired antibodies via Fc receptors.
- Add the antibodies indicated in Table 1.
- Prepare a mix diluting the desired antibodies in 100 μl/sample of PBS 1% FBS at a concentration of 4 μg/ml.
- Add 100 μl of the mix prepared in step 3.2.3.1 to each sample so that in the tube, there will be a volume of 200 μl of antibody mix at a final concentration of 2 μg/ml.
- Incubate for 30 min at 4 °C and cover from light.
Wash with 1 ml of PBS 1% FBS, centrifuge for 5 min at 400 x g, discard the supernatant and resuspend in 300 μl of PBS 1% FBS.
Add DAPI at a final concentration of 2 μg/ml and incubate at 4 °C, protected from light for 10 min.
- Analyze the samples obtained by FACS.
- In order to isolate leukocytes, use the following gating strategy:
- First eliminate any dead cells by gating on the DAPI- population.
- Select singlets in the FSC-A vs. FSC W plot as indicated in Figure 1.
- Gate the cells based on size and granularity typical of the leukocytes of interest.
- Select CD45+ cells in the FSC-A CD45 plot as indicated in Figure 1.
- Distinguish the different populations of leukocytes by the expression of surface markers as follows:
- Identify DCs as CD11c+ MHC II+ cells. These can be later analyzed for the expression of CD207 to distinguish Langerhans cells and CD207+ Dermal DCs from CD207- Dermal DCs.
- Identify Macrophages as CD64+ F4/80+ cells.
- Identify B cells as CD19+ MHCII+ cells.
- Identify T cells as CD8+or CD4+ cells.
Representative Results
Skin from different region of the body was digested according to the protocol described and stained with the indicated antibodies. The gating strategy is depicted in Figure 1. First select live cells (DAPI- cells), then gate on singlets (FSC-A vs. FSC-W) and on cells with lymphocyte morphology (FSC-A vs. SSC-A). When indicated, CD45+ cells from hematopoietic origin are selected.
DCs are labeled with anti-CD11c, -MHCII, and -CD45 antibodies, Macrophages are identified as F4/80+ CD64+ CD45+ cells, B cells as CD19+CD45+ MHCII+ cells, CD4+T lymphocytes as CD4+CD45+ cells, and CD8+ T lymphocytes as CD8+CD45+ cells (Figure 2) .
This method guarantees high viability of the digested population with more than 50% of total viable cells (DAPI-) in all regions of the skin that are analyzed (Figure 3A). An indication of the number of CD45+ cells yield per cm2 is depicted in Figure 3B.
An estimate of the total number of DCs, Macrophages, B cells and CD4+ or CD8+ T cells obtainable from the digestion of 1 cm2 of mouse skin is indicated in Figure 3C.
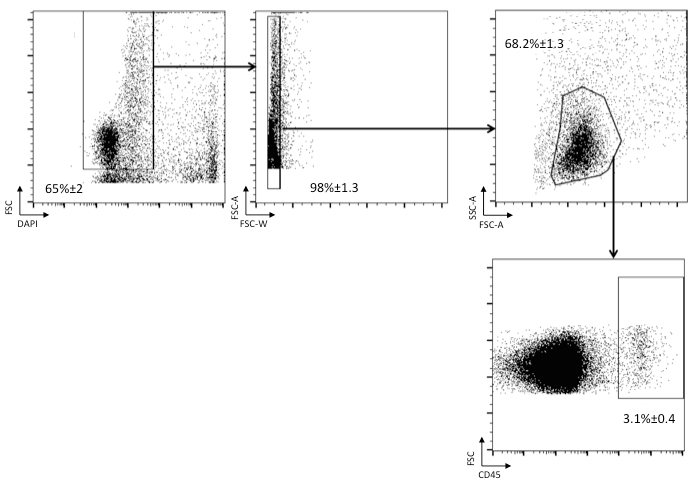 Figure 1. Identification of cells of hematopoietic origin in skin single-cell suspensions. Single-cell suspensions have been stained with DAPI and the anti-CD45 antibody. Among live cells (DAPI-negative), singlets (FSC-A vs FSC-H) are analyzed to select cells with leukocyte morphology (FSC-A vs. SSC-A). Cells of hematopoietic origin are identified as CD45+.
Figure 1. Identification of cells of hematopoietic origin in skin single-cell suspensions. Single-cell suspensions have been stained with DAPI and the anti-CD45 antibody. Among live cells (DAPI-negative), singlets (FSC-A vs FSC-H) are analyzed to select cells with leukocyte morphology (FSC-A vs. SSC-A). Cells of hematopoietic origin are identified as CD45+.
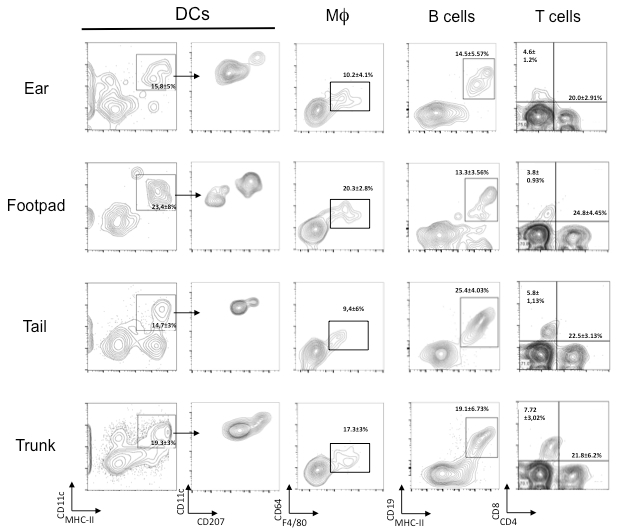 Figure 2. Analysis of immune cells present in the mouse skin derived from different regions. Cells are gated as indicated in Figure 1. DCs are identified as CD11c+MHC II+ CD45+ cells and later stained with CD207 to distinguish Langerhans cells and CD207+ dermal DCs from CD207- ; Macrophages are identified as CD64+F4/80+CD45+ cells; T lymphocytes are divided in CD8+CD45+ cells and CD4+CD45+ cells; B cells are identified as MHCII+CD19+CD45+ cells. Percentages are referred to the total CD45+ cells.
Figure 2. Analysis of immune cells present in the mouse skin derived from different regions. Cells are gated as indicated in Figure 1. DCs are identified as CD11c+MHC II+ CD45+ cells and later stained with CD207 to distinguish Langerhans cells and CD207+ dermal DCs from CD207- ; Macrophages are identified as CD64+F4/80+CD45+ cells; T lymphocytes are divided in CD8+CD45+ cells and CD4+CD45+ cells; B cells are identified as MHCII+CD19+CD45+ cells. Percentages are referred to the total CD45+ cells.
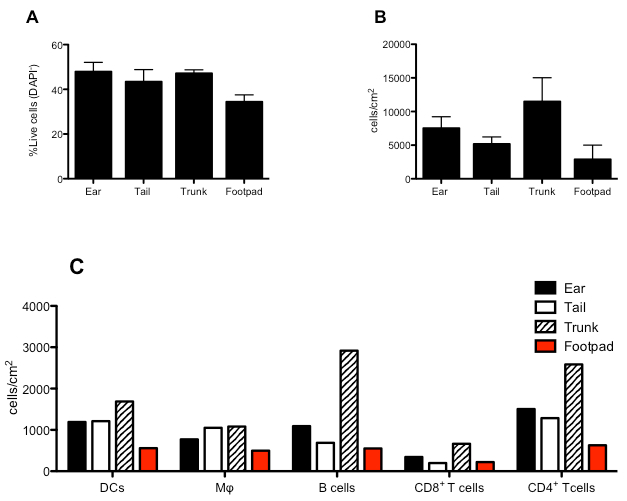 Figure 3. Quantitative analysis of the immune cells present in the mouse skin from different regions. (A) Percentage of viable (DAPI-) cells derived from the indicated regions of mouse skin. (B) Absolute numbers CD45+ cells in the different regions of mouse skin expressed in number of cells per cm2 of skin. (C) Estimate number of DCs, MΦ, B cells and CD4+ or CD8+ T cells derived from 1 cm2 of skin from the indicated regions of mouse skin.
Figure 3. Quantitative analysis of the immune cells present in the mouse skin from different regions. (A) Percentage of viable (DAPI-) cells derived from the indicated regions of mouse skin. (B) Absolute numbers CD45+ cells in the different regions of mouse skin expressed in number of cells per cm2 of skin. (C) Estimate number of DCs, MΦ, B cells and CD4+ or CD8+ T cells derived from 1 cm2 of skin from the indicated regions of mouse skin.
| Antibody | Staining | Concentration | Time | Temperature |
| DCs Mix | ||||
| CD11c | PE-Cy7 | 2 µg/ml | 30 min | 4 °C |
| MHC II | PE | " | " | " |
| CD45 | PE-Cy5.5 | " | " | " |
| CD207 | APC | " | " | " |
| Macrophages Mix | ||||
| F4/80 | APC-Cy7 | 2 µg/ml | 30 min | 4 °C |
| MHC II | PE | " | " | " |
| CD45 | PE-Cy5.5 | " | " | " |
| CD64 | PE-Cy7 | " | " | " |
| T and B cells Mix | ||||
| CD8a | PE | 2 µg/ml | 30 min | 4 °C |
| CD4 | APC-Cy7 | " | " | " |
| CD19 | APC | " | " | " |
| CD45 | PE-Cy5.5 | " | " | " |
Table 1.Indications for the staining of the different populations of immune cells populating the skin. Three mixes are indicated for staining of DCs, Macrophages and lymphocytes.
Discussion
We have described a method for the preparation of single-cell suspensions from different mouse skin regions. The method of digestion we have adopted, not only gives high yields but also preserves the expression of surface markers, which is fundamental for the subsequent FACS analysis.
The use of a cocktail of collagenase I and II and thermolysin, guarantees minimum batch to batch differences in enzyme activity making this method highly reproducible. Other published methods rely on different enzyme cocktails but in our hands they give lower yields and, more importantly, are less reproducible. Our method, however, does not discriminate between cells populating the epidermis and cells populating the dermis. Since in many cases it could be useful to distinguish between these two different compartments, we direct the reader to these previous works that use such methods.15
In our experience skin samples from the ear and tail are easier to digest than the trunk skin where the subcutaneous grease layer has to be thoroughly scraped in order to make the dermis accessible to the enzyme cocktail. The de-greasing step, though as well as the cutting should be performed quickly and gently in order to maintain cell viability. A prolonged or too thorough degreasing step can result in damage to the immune cells resident in the skin and therefore result in much lower yield and decreased viability of the recovered cells.
It should also be noted that to preserve the integrity of surface markers, the digestion should be performed in a medium devoid of reducing agents such as β-mercaptoethanol. To avoid unwanted damage to surface markers, the incubation time should be kept as shorter as possible and, in any event, not prolonged for more than 90 min.
Particular care should be, also, taken when pouring the digested skin samples into the cell strainer. It is important to thoroughly wash the digested skin in order to wash away the liberated cells before smashing the digested sample with the syringe plunger. With this step the skin sample is washed from the liberated immune cells that then do not undergo any more physical stress. The smashing should also be performed gently in order not to kill any cells and both the cell strainer and the syringe plunger should be thoroughly washed in the subsequent washing step.
Skin digestion can be also performed in a 50 ml falcon in a water bath at 37 °C. In this case the skin can be chopped beforehand and then put into the falcon tube. Note that the skin samples should always be submerged in digestion buffer during the incubation and that ear-pieces stick to the sides of falcon tubes. When using falcon tubes, therefore, make sure the pieces of chopped skin stay submerged in the digestion medium.
This method can be useful in many different experimental settings, particularly when studying non-inflamed skin. During inflammation, vasodilation and tissue swelling loosen the structure of the tissue and immune cells from blood are massively recruited to the skin. The efficiency of cell recovery is, thus, much higher. In homeostatic conditions, an effective way to digest the tight collagen matrix of the dermis is, in fact, needed in order to recover the immune cell constituting the complex network populating the skin.
The reader might also be interested in a comprehensive review that covers the origin and phenotype of different dendritic cells and macrophages populating the skin in both homeostatic and inflammatory conditions.16
In different experimental settings, different portions of mouse skin can be treated. For instance, in skin transplantation models, a portion of the tail could be transplanted on the trunk of a receiving animal; when studying edema formation or immunization procedures, stimuli are often injected on the footpad of mice, while the ear is frequently used to assess T cell memory with DTH assays. With our method we could efficiently extract skin resident immune cells from different locations making our method useful for the experimental setting of choice.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
This work was supported by grants from the Associazione Italiana per la Ricerca sul Cancro (AIRC, IG14593, MFAG13235), the Fondazione Cariplo (Grant 2010-0678 and NANOVAC) and the Fondazione Regionale per la Ricerca Biomedica.
References
- Streilein WJ. Circuits and signals of the skin-associated lymphoid tissues (SALT) Journal of Investigative Dermatology. 1985;85(1 Suppl):10s–13s. doi: 10.1111/1523-1747.ep12275413. [DOI] [PubMed] [Google Scholar]
- Tay SS, Roediger B, Tong PL, Tikoo S, Weninger W. The Skin-Resident Immune Network. Current dermatology reports. 2014;3:13–22. doi: 10.1007/s13671-013-0063-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen W, et al. Adaptive immunity to murine skin commensals. Proceedings of the National Academy of Sciences of the United States of America. 2014;111(29):E2977–E2986. doi: 10.1073/pnas.1401820111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broggi A, Zanoni I, Granucci F. Migratory conventional dendritic cells in the induction of peripheral T cell tolerance. Journal of leukocyte biology. 2013;94(5):903–911. doi: 10.1189/jlb.0413222. [DOI] [PubMed] [Google Scholar]
- Pan J, et al. DCs metabolize sunlight-induced vitamin D3 to'program'T cell attraction to the epidermal chemokine CCL27. Nature Immunology. 2007;8(3):285–293. doi: 10.1038/ni1433. [DOI] [PubMed] [Google Scholar]
- Vitali C, et al. Migratory, and not lymphoid-resident, dendritic cells maintain peripheral self-tolerance and prevent autoimmunity via induction of iTreg cells. Blood. 2012;120(6):1237–1245. doi: 10.1182/blood-2011-09-379776. [DOI] [PubMed] [Google Scholar]
- Azukizawa H, et al. Steady state migratory RelB+ langerin+ dermal dendritic cells mediate peripheral induction of antigen-specific CD4+ CD25+ Foxp3+ regulatory T cells. European journal of immunology. 2011;41(5):1420–1434. doi: 10.1002/eji.201040930. [DOI] [PubMed] [Google Scholar]
- Idoyaga J, et al. Specialized role of migratory dendritic cells in peripheral tolerance induction. The Journal of clinical investigation. 2013;123(2):844–854. doi: 10.1172/JCI65260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyman O, Conrad C, Tonel G, Gilliet M, Nestle FO. The pathogenic role of tissue-resident immune cells in psoriasis. Trends in immunology. 2007;28(2):51–57. doi: 10.1016/j.it.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Di Meglio P, Perera GK, Nestle FO. The multitasking organ: recent insights into skin immune function. Immunity. 2011;35(6):857–869. doi: 10.1016/j.immuni.2011.12.003. [DOI] [PubMed] [Google Scholar]
- Rosenblum MD, et al. Response to self antigen imprints regulatory memory in tissues. Nature. 2011;480(7378):538–542. doi: 10.1038/nature10664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenblum MD, Truong HA, Abbas AK, Gratz IK. 177: Maintenance of memory regulatory T cells in peripheral tissues. Cytokine. 2013;63(3):284–285. [Google Scholar]
- Kim BS, et al. TSLP elicits IL-33-independent innate lymphoid cell responses to promote skin inflammation. Science translational medicine. 2013;5(170) doi: 10.1126/scitranslmed.3005374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gopinathan KM. New insights into skin structure: scratching the surface. Advanced drug delivery reviews. 2002;54(Suppl 1):S3–S17. doi: 10.1016/s0169-409x(02)00121-7. [DOI] [PubMed] [Google Scholar]
- Sandrine H, et al. CD207+ CD103+ dermal dendritic cells cross-present keratinocyte-derived antigens irrespective of the presence of Langerhans cells. The Journal of Experimental Medicine. 2010;207(1):189–206. doi: 10.1084/jem.20091964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard M, Samira T, Sandrine H. The origins and functions of dendritic cells and macrophages in the skin. Nature Reviews Immunology. 2014;14(6):417–428. doi: 10.1038/nri3683. [DOI] [PubMed] [Google Scholar]