

Abstract
Clostridium difficile is a global health burden and the leading cause of antibiotic-associated diarrhoea worldwide, causing severe gastrointestinal disease and death. Three well characterised toxins are encoded by this bacterium in two genetic loci, specifically, TcdB (toxin B) and TcdA (toxin A) in the Pathogenicity Locus (PaLoc) and binary toxin (CDT) in the genomically distinct CDT locus (CdtLoc). Toxin production is controlled by regulators specific to each locus. The orphan response regulator, CdtR, encoded within the CdtLoc, up-regulates CDT production. Until now there has been no suggestion that CdtR influences TcdA and TcdB production since it is not carried by all PaLoc-containing strains and CdtLoc is not linked genetically to PaLoc. Here we show that, in addition to CDT, CdtR regulates TcdA and TcdB production but that this effect is strain dependent. Of clinical relevance, CdtR increased the production of TcdA, TcdB and CDT in two epidemic ribotype 027 human strains, modulating their virulence in a mouse infection model. Strains traditionally from animal lineages, notably ribotype 078 strains, are increasingly being isolated from humans and their genetic and phenotypic analysis is critical for future studies on this important pathogen. Here we show that CdtR-mediated toxin regulation did not occur in other strain backgrounds, including a ribotype 078 animal strain. The finding that toxin gene regulation is strain dependent highlights the regulatory diversity between C. difficile isolates and the importance of studying virulence regulation in diverse lineages and clinically relevant strains. Our work provides the first evidence that TcdA, TcdB and CDT production is linked by a common regulatory mechanism and that CdtR may act as a global regulator of virulence in epidemic 027 strains.
Author Summary
Clostridium difficile is the leading cause of antibiotic-associated diarrhoea. The TcdB, TcdA and binary toxins produced by C. difficile are encoded in two genomically distinct loci: TcdB and TcdA in the Pathogenicity Locus (PaLoc) and binary toxin (CDT) in the CDT locus (CdtLoc). Toxin production is primarily controlled by regulators specific to each locus. Because the presence of these loci varies amongst different strains of C. difficile, no rational link for their co-regulation has ever been proposed. Here we have shown that the regulator of CDT production, CdtR, also regulates production of TcdA and TcdB in a strain dependent manner. These results represent the first evidence that TcdA and TcdB production is linked to the production of CDT by a common regulatory mechanism. Collectively, our results establish CdtR as an important virulence regulator in two clinically important, epidemic strains of C. difficile, and further highlights the need to investigate regulatory mechanisms of important virulence factors in diverse strain backgrounds.
Introduction
C. difficile antibiotic-associated diarrhoea is a toxin mediated disease [1,2]. During infection, TcdA, TcdB and CDT are secreted into the colonic epithelium by this bacterium, leading to diarrhoea that can progress to serious, life threatening inflammatory diseases, including pseudomembranous colitis and toxic megacolon [3]. The production of these toxins varies between strains. TcdB is the most commonly encoded toxin and is most often co-located with the TcdA gene in the PaLoc region [4], both toxins act as monoglucosyltransferases that irreversibly modify Rho family members [3]. PaLoc variants that produce TcdB and not TcdA are, however, becoming increasingly common, for example, they represented 23% of strains in one recent study of human strains in China [5]. CDT is encoded in a specific locus, CdtLoc (Fig 1) [6] the carriage of which has also increased significantly over the last decade; in 2004 6% of clinical isolates encoded CDT whereas 33.5% now encode this toxin [7,8]. CDT is an ADP-ribosyltransferase that is not essential for disease, but may be important for colonisation during an infection [9]. CdtLoc is not carried by all PaLoc-containing strains and it is not linked genetically to PaLoc.
Fig 1. Schematic representation of the CDT loci from representative strains.

a, The full length CdtLoc from the ribotype 027 strains (M7404 and R20291). b, The CdtLoc from the ribotype 078 strain, JGS6133. The cdtR pseudogene is shown in black and is grey after the premature stop codon. c, The CdtLoc from the ribotype 012 strain 630 carrying cdtAB pseudogenes (shown in black). d, The CdtLoc in the CDT negative strain CD37 is replaced with a 68 bp sequence. The boundaries of the CdtLoc are indicated with vertical lines and the flanking genes are blue.
Regulation of toxin production in C. difficile is somewhat strain dependent, suggesting that toxin regulatory mechanisms have evolved independently to modulate pathogenesis [10–13]. The TcdR alternative sigma factor and TcdC anti-sigma factor, which are encoded with tcdA and tcdB in the PaLoc, act as the primary mechanism controlling the production of these toxins [14,15]. TcdA and TcdB regulation has also been linked to many important cellular processes in the C. difficile life cycle, including sporulation, by Spo0A, the master sporulation regulator, motility, via the flagella regulator SigD, and nutrient acquisition, by the regulators of carbon and amino acid metabolism, CcpA and CodY [10,11,16–20]. The ribotype 027 strains associated with epidemics of severe CDI appear to be more virulent than strains previously isolated, a phenotype that has been partly attributed to increased TcdA and TcdB production [21–23]. By comparison, little is known about the regulation of CDT production beyond the involvement of CdtR, and until now, no link had been identified between the control of CDT, TcdA and TcdB production. The difficulty in genetically manipulating strains from different C. difficile clonal lineages has also prevented a broader analysis of the role of this regulator across different strain types.
In this study, we investigated the role of CdtR in different strains of C. difficile including two epidemic ribotype 027 strains. As expected, CdtR was found to regulate the production of CDT. Surprisingly, however, CdtR also regulated the production of the PaLoc encoded toxins, TcdA and TcdB, in the two ribotype 027 strains. Further analysis showed that regulation occurred at the transcriptional level and probably resulted from indirect regulation of the positive regulator of PaLoc gene expression, TcdR. Importantly, further analysis showed the importance of CdtR for C. difficile pathogenesis, with cdtR mutants causing less severe disease than the wild type strain in a mouse infection model. To determine whether CdtR function is conserved across evolutionarily diverse isolates, ribotype 078 (JGS6133) and 012 (630) strains were investigated. CdtR regulated CDT production in the ribotype 078 strain; the ribotype 012 strain does not encode CDT. Notably, and in contrast to the ribotype 027 strains, CdtR did not regulated TcdA or TcdB production in either strain background, highlighting the regulatory variation of key virulence factors between C. difficile strains. These results highlight the importance of investigating regulatory mechanisms in clinically important strains of C. difficile and suggest that CdtR-mediated toxin regulation is an important virulence mechanism in the epidemic ribotype 027 strains.
Results
CdtR regulates production of CDT, TcdA and TcdB in two epidemic, ribotype 027 strains of C. difficile
To investigate the role of CdtR in the regulation of CDT production in the epidemic ribotype 027 strains, we constructed two independent cdtR mutants in the Canadian isolate M7404 and a cdtR mutant in the UK isolate R20291 and confirmed their genotype by Southern hybridisation (S1 Fig). Western blot analysis showed that the cdtR mutants produced less CDTa and CDTb compared to the wild type and that complementation with cdtR in trans resulted in over-expression of both toxin subunits (Fig 2A and 2B). Consistent with these results, ADP-ribosyltransferase assays demonstrated that the cdtR mutants had significantly reduced levels of CDT activity compared to the wild type, while the complemented cdtR mutants showed CDT activity greater than that of the wild type strain (Fig 2D and 2E). Overall, these data show, for the first time, that CdtR is important for regulating CDT production in epidemic ribotype 027 C. difficile strains.
Fig 2. Analysis of CDT production.

a–c, Western immunoblot using CDTa-specific and cross-reactive Ib-specific antibodies and precipitated supernatants from the strains indicated. CD37 (non-toxigenic), V = vector control, R+ = cdtR complemented. The arrows indicate the 48 kDa CDTa and 99 kDa CDTb proteins. d–f, CDT activity assessed by ADP-ribosyltransferase assay. Samples were separated by SDS-PAGE and biotinylated (ADP-ribosylated) actin detected by HRP-streptavidin. Relative CDT activity was assessed by densitometry compared to the non-toxigenic control strain CD37. A = actin, A-A = ADP-ribosylated actin. Data represent the mean ± SEM (n = 3). *, p ≤ 0.05.
Unexpectedly, Western blots performed using TcdA- and TcdB-specific antibodies showed that the cdtR mutants produced less TcdA and TcdB than the wild type, while the complemented cdtR derivatives expressed high levels of both toxins (Fig 3A and 3C). Cytotoxicity assays were performed using HT29 and Vero cells to measure the activity of TcdA and TcdB, respectively, in the culture supernatants from the isogenic panel of M7404 and R20291 strains. The TcdA and TcdB activities of all of strains increased over time and, consistent with the Western blot results, showed lower activity in supernatants from the cdtR mutants compared to the wild type, confirming that the cdtR mutants were less cytotoxic in vitro (Fig 3B and 3D). TcdA and TcdB activity of the complemented cdtR mutants was consistently higher than the cdtR mutants and the wild type across all time points (Fig 3B and 3D). These data show that CdtR regulates the production of TcdA and TcdB in both M7404 and R20291, which is the first demonstration of a common regulator modulating the expression of all three toxins in C. difficile.
Fig 3. Analysis of TcdA and TcdB production.

a, c, e, Western immunoblot using TcdA-specific and TcdB-specific antibodies with precipitated supernatant from the strains indicated. CD37 (non-toxigenic), V = vector control, R+ = cdtR complemented. Arrows indicate the 308 kDa TcdA and 270 kDa TcdB proteins. Supernatants were collected at 12, 24, 48 and 72 hours post inoculation and assayed by doubling dilution cytotoxicity assays. b, The panel of M7404 strains were assayed using HT29 cells and Vero cells. d, R20291 panel of strains assayed using HT29 cells and Vero cells. Data represent the mean ± SEM (n = 3–5). *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001.
CdtR positively regulates expression of genes from the pathogenicity locus (PaLoc) of C. difficile
To investigate the molecular mechanism of regulation, we determined if CdtR controlled toxin production at the transcriptional level. Using the isogenic panel of M7404 strains, reverse-transcription droplet digital PCR (RT-ddPCR) analysis was employed to quantitatively compare the level of expression of each of the toxin encoding genes (tcdA, tcdB, cdtA) and the PaLoc-encoded toxin regulators (tcdR, tcdC). The relative transcription of all three toxin genes and tcdR was significantly decreased in both cdtR mutants compared to the wild type (Fig 4A–4D). Over-expression of cdtR in the complemented strains resulted in a dramatic over-expression of tcdA, tcdB, cdtA and tcdR (Fig 4A–4D). Although TcdC is predicted to be non-functional in the 027 strains [12], we analysed the expression of tcdC and found it to be similar in the isogenic panel of strains (Fig 4E). Previous work in strain 630Δerm, a derivative of the non-027 historical strain 630 which belongs to ribotype 012, showed that the flagella synthesis regulator, SigD, is an important regulator of TcdA and TcdB production via the regulation of tcdR expression [24]. Analysis of our isogenic panel of M7404 strains showed no change in sigD transcription, suggesting that CdtR does not influence the expression of sigD in this strain background and that therefore the modulation of tcdA and tcdB expression does not occur via SigD (Fig 4F). Collectively, these results indicate that regulation of all three toxins by CdtR in M7404 occurs at a transcriptional level and that the regulation of tcdA and tcdB occurs via the upregulation of tcdR transcription.
Fig 4. Transcriptional analysis of M7404 cdtR mutant and complemented strains compared to wild-type.
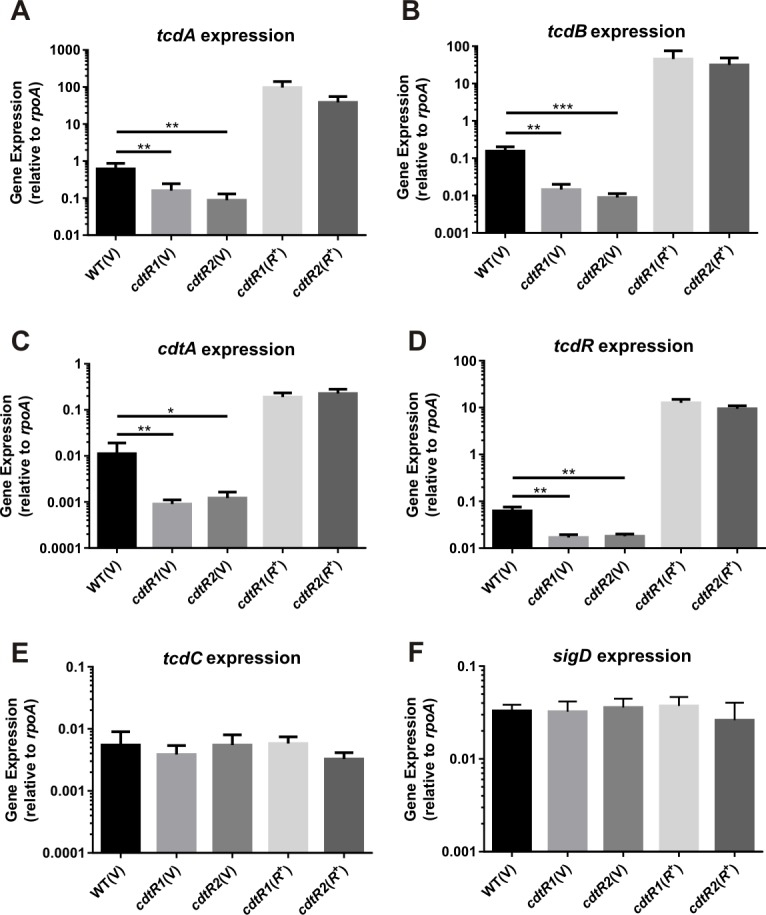
RNA was isolated from strains for analysis of (a) tcdA, (b) tcdB, (c) cdtA, (d) tcdR, (e) tcdC and (f) sigD expression. Levels of gene expression were normalised to rpoA. Data represent the mean normalised gene expression ± SEM from five independent biological replicates. *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001.
CdtR is important for modulating C. difficile virulence
Our observation that CdtR increases toxin production in ribotype 027 strains has important implications for the virulence capacity of these clinically important strains. To determine if the modulatory effect of CdtR on toxin production influences C. difficile disease, we examined whether cdtR inactivation altered virulence in our mouse infection model [2]. It was previously shown that infection with a cdtA mutant of M7404 resulted in disease that was indistinguishable from the parent strain [2]. This mutant no longer produced CDT, but had an intact cdtR gene and continued to produce TcdA and TcdB at wild type levels. A reduction in CDT levels is therefore not likely to have a major effect on disease in our animal model. All of the mice infected with the isogenic cdtR-series of M7404 derivatives were colonised with C. difficile at similar levels (S2 Fig). Wild type-infected mice rapidly lost weight and the majority were euthanized 40 to 48 hours post infection in accordance with animal ethics guidelines, with a mean time to death of 48 ± 5.1 hours and a survival rate of 13% (Fig 5). Mice infected with either of the two independent cdtR mutants had significantly higher survival rates (Mantel-Cox log rank test, P < 0.0001) of 100% and 96% for cdtR1(V) and cdtR2(V), respectively, and showed no overt signs of disease nor significant weight loss (Fig 5). By comparison, mice infected with either complemented mutant, cdtR1(R +) or cdtR2(R +), had a wild-type virulence phenotype, with marked weight loss, and a mean time to death of 31.5 ± 2.5 hours and 57.6 ± 7.5 hours, respectively, reflected in survival rates of 0% and 33% (Fig 5).
Fig 5. Virulence of M7404 wild-type, cdtR mutant and complemented strains in mice.

a, Kaplan-Meier survival curve showing time from infection to euthanasia of mice infected with different strains of C. difficile in hours. (n = 15). b, Time from inoculation of mice to death in hours. Data represents mean ± S.E.M. (n = 15). Data represent the mean ± SEM ****, p ≤ 0.0001.
Damage to the colon and caecum of C. difficile-infected mice results from the production of TcdA and TcdB [2]. We therefore performed histopathological analysis to assess the damage to colonic and caecal tissues collected from the groups of infected and uninfected mice under study here. All tissues were de-identified and independently scored using a previously defined set of scoring parameters that included overall tissue damage, polymorphonucleocyte (PMN) influx, crypt damage and oedema [2]. Tissues of uninfected mice only had minimal surface damage to the intestinal epithelia resulting from the disruption of microbiota by antibiotic pre-treatment and tissue processing (Fig 6A) with low colon and caecum damage scores of 4.7 and 3.8, respectively (Fig 6B and 6C). By comparison, wild type-infected mice had severely inflamed tissues, with extensive damage to the epithelial surface, crypt branching and hyperplasia, goblet cell loss, significant PMN influx and mucosal and sub-mucosal oedema (Fig 6A). These histopathologies were reflected in the high damage scores of 12.9 and 13.6 for their colonic and caecal tissues, respectively (Fig 6B and 6C).
Fig 6. Histopathology of C. difficile infected tissues.
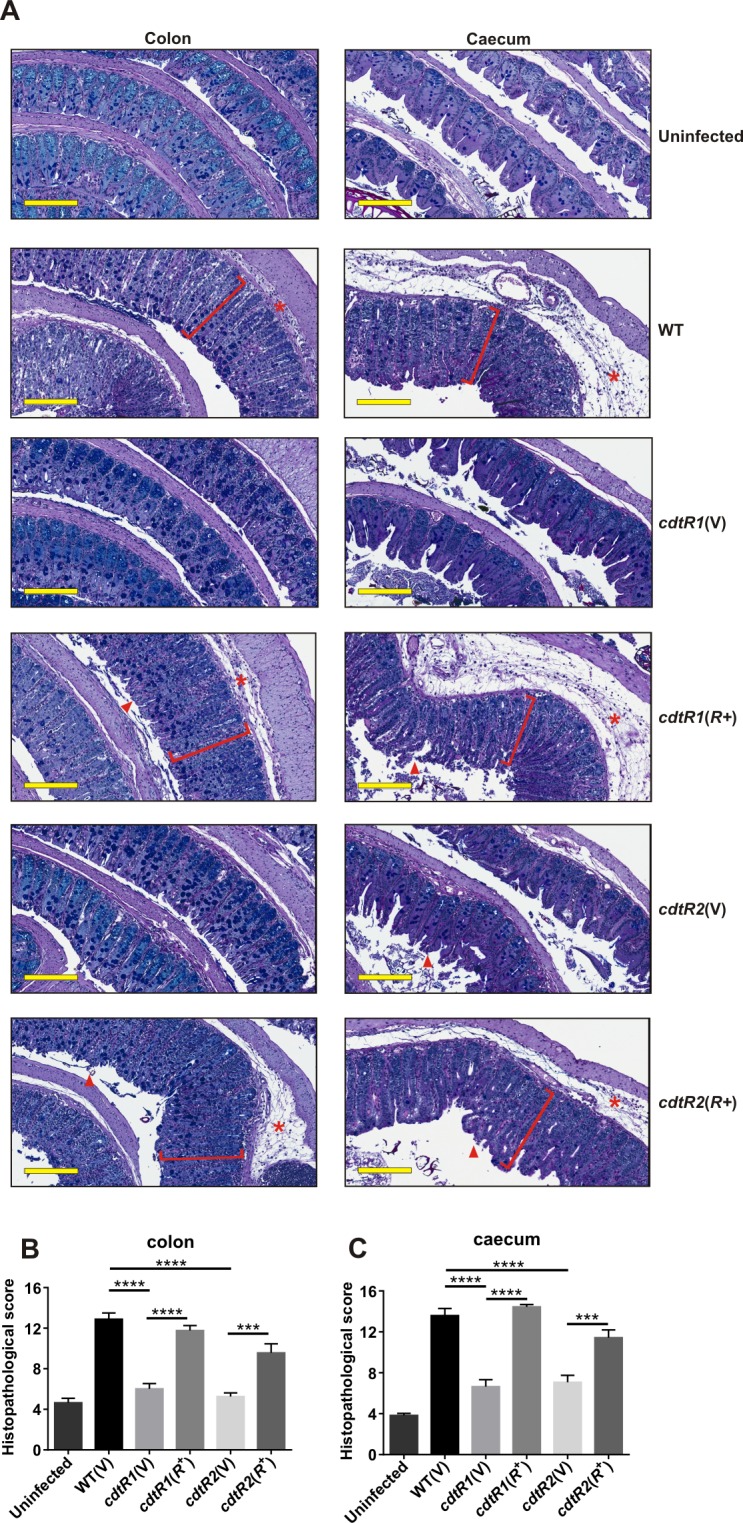
Representative images of sections of colon and caeca collected from uninfected mice or mice infected with different strains of C. difficile, fixed and strained with PAS-Alcian blue. Red brackets ([) indicate crypt hyperplasia, arrow heads (▲) represent surface epithelial damage and asterisks (*) represent oedema and inflammation. Scale bars (200 μm) are shown in yellow. Histopathology damage scores from uninfected or infected colons (b) and caeca (c). Data represent the mean ± SEM ****, p ≤ 0.0001.
Mice infected with either cdtR mutant had tissue architecture similar to that seen with the uninfected mice. Very little colonic and caecal damage was observed, with some surface epithelial damage to crypts occurring and no apparent crypt hyperplasia, and little PMN influx into the mucosa (Fig 6A), with correspondingly low colonic (6.0 and 5.3) and caecal (6.6 and 7.1) damage scores (Fig 6B and 6C). However, mice infected with either cdtR-complemented mutant had similar levels of tissue damage to mice infected with the wild-type strain. Severe crypt damage was observed in the majority of these tissues, particularly the caeca, with crypt hyperplasia, loss of goblet cells, PMN influx and severe oedema in the mucosa and sub-mucosa (Fig 6A) and high histopathological damage scores were determined for both the colon (11.8 and 9.6) and the caecum (14.5 and 11.4) (Fig 6B and 6C).
To confirm that the reduced virulence of the cdtR mutants can be attributed to reduced levels of TcdA and TcdB production in vivo, the cytotoxicity of the intestinal contents collected from mice infected with the panel of C. difficile strains was assessed. No cytotoxicity was observed against HT29 or Vero cells by faecal samples collected from uninfected mice whereas mice infected with the wild type strain showed high levels of cytotoxicity against HT29 and Vero cells, indicating the production of TcdA and TcdB in vivo (S4 Fig). By comparison, samples collected from mice infected with the two cdtR mutants (cdtR1(V) and cdtR2(V)) showed reduced cytotoxicity against HT29 or Vero cells, indicating decreased levels of TcdA and TcdB production in vivo compared to the wild type strain (S4 Fig). Intestinal contents collected from mice infected with the two complemented cdtR mutants ((cdtR1(R +) and cdtR2(R +)) showed increased levels of cytotoxicity against HT29 and Vero cells, indicating restored in vivo TcdA and TcdB production. Collectively, the survival data, histopathological and in vivo cytotoxicity analysis support the hypothesis that CdtR modulates the virulence of the C. difficile ribotype 027 strain M7404 due to the role that it plays in regulating TcdA and TcdB production.
CdtR differentially regulates toxin production in a ribotype 078 strain of C. difficile
To investigate whether TcdA and TcdB regulation by CdtR occurs in other strains, we assessed this phenotype in a ribotype 078 animal isolate, JGS6133. Ribotype 078 strains are commonly isolated from animals, but have also been implicated in severe human infections [25,26]. Our sequencing analysis revealed that the cdtR gene in JGS6133 contains a naturally occurring stop codon mutation at codon 322, which has been described previously in other 078 strains [27], and results in a 142 amino acid truncation that is likely to result in a non-functional protein (Fig 1B). Western blots showed that JGS6133 complemented with the full length cdtR gene in trans, and hence producing functional CdtR, had significantly more CDTa and CDTb than the wild-type and vector control strains, which produced similar amounts of both proteins (Fig 2C). Increased CDT production by the JGS6133 cdtR + derivative was reflected in increased toxin activity as assessed by ADP-ribosyltransferase assays (Fig 2F). By contrast, production of TcdA and TcdB was not altered in the JGS6133 cdtR + strain compared to the wild type (Fig 3E). This result suggests that CdtR-mediated regulation of TcdA and TcdB production is not conserved within all strains of C. difficile.
CdtR does not regulate TcdA and TcdB production in strain 630
Derivatives of a ribotype 012 strain, designated 630, have been routinely used for the analysis of gene regulation and other phenotypes in C. difficile due to the relative ease of their genetic manipulation. Many strains of C. difficile, including strain 630, contain cdtAB pseudogenes within the CdtLoc (Fig 1C). These genes contain multiple frameshift mutations and stop codons and do not encode a functional CDT, but still encode a full length CdtR protein [28]. The possibility that CdtR functions to regulate other important processes, such as TcdA and TcdB production, may provide a rationale for the retention of functional CdtR in strains that carry the cdtAB pseudogenes. We were therefore interested in investigating CdtR-mediated toxin regulation in a strain with these pseudogenes.
To determine whether CdtR regulates TcdA and TcdB production in strains of C. difficile with cdtAB pseudogenes we transferred the cdtR complementation vector, pJIR4218, into strain 630. Western blot analysis showed that derivatives of 630 over-expressing cdtR had similar levels of TcdA and TcdB production as the isogenic vector control strains (S3A and S3B Fig). RT-ddPCR analysis confirmed that cdtR was over-expressed in these strains relative to the wild type and the vector control (S3C Fig). Taken together these results suggest that CdtR is not important for the regulation of TcdA and TcdB production in strain 630. Since CdtR is also not important for TcdA and TcdB regulation in a ribotype 078 strain, regulation of these toxins by CdtR may be specific to the ribotype 027 strains.
Discussion
The emergence of epidemic ribotype 027 strains over a decade ago prompted several investigations of the genetic and phenotypic characteristics that may have led to the global dominance of these strains. These features may include higher sporulation rates, resistance to key antibiotics and unique aspects of toxin regulation [11,12,29–31]. The presence of a full length CdtLoc was also initially considered to be important in this regard because it encodes CDT [21,23], however, despite numerous studies, the importance of this toxin in virulence remains undefined [2,32,33]. The results of the work presented here suggest that CdtLoc, and specifically CdtR, may play an indirect but significant role in disease pathogenesis of the ribotype 027 strains by regulating TcdA and TcdB production.
Our work confirms that CdtR enhances CDT production. Strikingly, ribotype 078 strains produce CDT even though they contain a conserved mutation in cdtR and they have CDT activity that is not significantly different from that of strains without this mutation [27,34]. Our data supports these observations since CDT production was detected from the ribotype 078 strain JGS6133 which contains this naturally occurring cdtR mutation. However, CDT production from JGS6133 was enhanced when functional CdtR was expressed in this strain. Even though CdtR is not essential for CDT production, our work suggests that the presence of this regulator increases the expression of this toxin.
Although CdtR was previously shown to be an important regulator of CDT production [6] there was no evidence to suggest that it played a role in TcdA or TcdB production, particularly since the pathogenicity loci encoding these toxins are not genomically linked. Our data clearly show that CdtR is an important regulator of TcdA and TcdB production, as well as CDT, in two ribotype 027 strains, and that this regulatory capacity plays a role in virulence since inactivation of cdtR attenuated the virulence of strain M7404 in a mouse infection model. The results obtained with the mouse infection experiments are directly relevant to the disease-causing capacity of these strains. This is a significant finding as it is the first report of a regulatory link between the two pathogenicity loci, PaLoc and CdtLoc.
The ability to regulate toxin production through a variety of mechanisms may provide a selective advantage to C. difficile since it may allow virulence factors to be produced in response to different and specific environmental cues. Ribotype 027 strains appear to have evolved to differ in their regulatory responses in comparison to other C. difficile lineages. C. difficile regulates toxin production in response to many environmental stimuli, including metabolisable carbon sources and quorum signalling molecules, through several different regulatory proteins [20,35,36]. TcdR is the primary positive regulator of TcdA and TcdB production, while TcdC represses toxin production [12,15]. TcdR is highly conserved between strains and many regulators directly influence its expression [20,24,27,35]. Our data suggest that CdtR may regulate TcdA and TcdB production by controlling tcdR expression. CdtR belongs to the LytTR family of DNA binding response regulators and may function by binding to the tcdR promoter, thereby regulating TcdR expression and, consequently, tcdA and tcdB expression [6]. However, we could not identify canonical LytTR DNA binding sites upstream of the tcdR, tcdA or tcdB genes. Similarly, LytTR DNA binding sequences with the conserved sequence and spacing could not be identified within the promoters of other genes identified to regulate toxin production in C. difficile in other studies, including sigD, codY or ccpA. It may be that the CdtR binding sites are too dissimilar to typical LytTR sites to be identified or that CdtR does not directly bind to these regions; instead, an unidentified, CdtR-controlled intermediate regulator may be modulating PaLoc toxin gene expression.
CdtR-mediated TcdA and TcdB regulation may have specifically evolved in ribotype 027 C. difficile strains since the co-regulation of these toxins with CDT appears to be ribotype specific. CdtR does not play a similar role in other ribotypes tested here to that seen in ribotype 027 strains and this phenotype does not appear to be conserved between divergent strain backgrounds. The strains included in our assessment belong to three of the five defined evolutionary clades of C. difficile, specifically, clade 2 for ribotype 027 (M7404 and R20291), clade 5 for ribotype 078 (JGS6133) and clade 1 for ribotype 012 (630) [37]. Our observations in strain 630 are particularly relevant; many studies are performed using this isolate because it is relatively easy to genetically manipulate. It is clear from our research and other studies [10,11] that strain 630 characteristics may not always reflect those of other strains. Similar observations have been made for clade 5 ribotype 078 strains, which are genetically and phenotypically divergent from strains belonging to other clades [11,30,38,39].
Although the global regulators CodY and CcpA are conserved in strains of C. difficile, it has been shown that these regulators control toxin production experimentally only in a strain 630 background and their role in other strains, including the 027 and 078 strains, is not known [19,20]. Similarly, several flagella structural and regulatory genes, including SigD, have only been linked to toxin production in C. difficile in strain 630 [24]. While the genetic organisation of the flagella genes within the F1 and F3 flagella regions are similar in strain 630 and the 027 strains, the sequence variation in these regions is thought to contribute to their different motility phenotypes [30]. By comparison, the F3 region, which contains several genes involved in toxin regulation in strain 630, is absent in the 078 strains and is thought to explain the lack of motility in these strains [30]. It has been shown that several of the conserved flagella structural proteins encoded in the F1 and F3 flagella regions, including FliC, FliD and FlgE, are important for toxin production in strain 630 but do not contribute to toxin production in the 027 strain, R20291 [10]. Further research is required to determine if other conserved flagella genes, known to regulate toxin production in strain 630, play a similar role in 027 and 078 strain backgrounds.
To date, only one study has investigated the regulation of toxin production in an 078 strain and showed that the master sporulation regulator, Spo0A, differentially regulates toxin production in an 078 strain, two epidemic 027 strains and a strain 630 derivative [11]. Dingle et al. [40] found that strains from clade 5, including the 078 strains, carry a PaLoc similar to that found in other ribotypes but that genes outside of this region are highly divergent. It was suggested that the 078 strains may have originated from a divergent, non-toxigenic strain that obtained the PaLoc in a separate event in comparison to other toxigenic lineages. We present data supporting the concept that strains from this background have evolved different toxin regulatory mechanisms from the more commonly studied 027 strains and strain 630 derivatives.
The results presented here clearly show that modulation of tcdA and tcdB expression by CdtR may be specific to the ribotype 027 strains and is likely to be an important factor contributing to their increased virulence. Furthermore, the fluidity of the regulatory systems that control gene expression in C. difficile, exemplified by the toxin gene expression studies presented here, reflect the plasticity and dynamic nature of the C. difficile genome [37].
In conclusion, we have provided the first evidence that TcdA and TcdB production is linked to the production of CDT by a common regulatory mechanism and that CdtR acts as a global regulator of toxin production and virulence in two ribotype 027 strains. The observed differences in virulence between the ribotype 027 strains and other historical isolates have been attributed, in part, to elevated toxin production, mainly as a result of mutations in the tcdC gene [21–23]. Another key genetic difference identified between these strains is the possession of a full length CdtLoc [30]. Our results suggest that possession of the CdtLoc in the 027 strains enhances virulence by the CdtR-mediated up-regulation of TcdA and TcdB production. Therefore, we postulate that the ability of the epidemic ribotype 027 strains to coordinate production of all known C. difficile toxins, CDT, TcdA and TcdB, by CdtR is a key factor in the increased virulence of these strains.
Methods
Bacterial strains and growth conditions
All bacterial strains are defined in Table 1. Culture media were from Oxoid or Becton Dickinson (BD) and all antibiotics and supplements used are from Sigma-Aldrich, Amresco or Merck unless otherwise stated. E. coli and B. subtilis strains were cultured at 37°C in 2xYT media [41] supplemented with either chloramphenicol (25 µg/ml for E. coli; 5 μg/ml for B. subtilis) or tetracycline (10 µg/ml). C. difficile strains were cultured in HIS broth[42] or on HIS agar supplemented with 0.1% (w/v) L-cysteine and 0.375% (w/v) glucose or TY broth[1] with D-cycloserine (250 µg/ml), thiamphenicol (10 µg/ml), lincomycin (50 µg/ml) or anhydrous tetracycline (50 ng/ml), as required. C. difficile cultures were grown in a Don Whitley A35 Anaerobic Workstation in an atmosphere of 10% (v/v) H2, 10% (v/v) CO2 and 80% (v/v) N2 at 37°C.
Table 1. Bacterial strains and plasmids.
| Strain or plasmid | Characteristics | Source or reference |
|---|---|---|
| Strain | ||
| E. coli | ||
| DH5α | F– Φ80dlacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17(rK –, mK +) deoR thi-1 supE44 gyrA96 relA1 | Life Technologies |
| Top10 | F–mcrA Δ(mrr - hsdRMS - mcrBC) Φ80lacZΔM15 ΔlacX74 nupG recA1 araD139 Δ(ara-leu)7697 galU galK rpsL (StrR) endA1 λ– | Life Technologies |
| MM294 | F–, endA1, hsdR17(rK –mK +), supE44, thi-1, recA + | [50] |
| B. subtilis | ||
| BS34A | B. subtilis donor that carries a single copy of Tn916 on its chromosome, TcR | [51] |
| JIR6342 | BS34A carrying cdtR TargeTron plasmid, pJIR4153 | This study |
| JIR6345 | BS34A carrying cdtR complementation plasmid, pJIR4218 | This study |
| JIR6346 | BS34A carrying shuttle plasmid, pDLL24 | This study |
| C. difficile | ||
| M7404 | Canadian BI/NAP1/027 isolate | [6] |
| R20291 | UK BI/NAP1/027 isolate | [39] |
| CD37 | Non-toxigenic C. difficile isolate | [52] |
| JGS6133 | Porcine ribotype 078 isolate | [12] |
| DLL3094 | M7404 (pDLL24) | This study |
| cdtR1 (or JIR8707) | M7404 cdtR::TargeTron (mutant 1), LnR | This study |
| cdtR2 (or JIR8708) | M7404 cdtR::TargeTron (mutant 2), LnR | This study |
| JIR8739 | JIR8707 (pDLL24), LnR, TmR | This study |
| JIR8740 | JIR8707 (pJIR4218), LnR, TmR | This study |
| JIR8741 | JIR8708 (pDLL24), LnR, TmR | This study |
| JIR8742 | JIR8708 (pJIR4218), LnR, TmR | This study |
| JIR8729 | R20291 cdtR::TargeTron, LnR | This study |
| JIR8745 | JIR8729 (pDLL24), LnR, TmR | This study |
| JIR8746 | JIR8729 (pJIR4218), LnR, TmR | This study |
| JIR8747 | R20291 (pDLL24), TmR | This study |
| JIR8733 | JGS6133 (pDLL24), TmR | This study |
| JIR8735 | JGS6133 (pJIR4218) | This study |
| Plasmid | ||
| pDLL4 | C. difficile shuttle vector allows plasmid to be mobilised by Tn916, TmR | [12] |
| pDLL24 | pDLL4 carrying lacZα, CmR | This study |
| pJIR4218 | pDLL24 carrying cdtR and its promoter, CmR | This study |
| pDLL45 | pMTL9361 derivative with HindIII and BsrGI sites removed from rep; TmR | This study |
| pDLL55 | Derivative of pDLL45 carrying lacZα; TmR | This study |
| pJIR4135 | Group II intron of pDLL45 retargeted to insert between codons 288/289 of the cdtR gene, CmR | This study |
| pJIR4153 | pDLL55 (StuI/HindIII)Ω pJIR4135 group II intron (StuI/HindIII) | This study |
Polymerase Chain Reaction (PCR)
All oligonucleotide primers are listed in Table 2. PCR cycling conditions (unless otherwise stated) were as follows: initial denaturation step at 94°C for 4 min, followed by 30 cycles of denaturation at 94°C for 30 sec, an annealing step at 50°C for 30 sec and an extension step at 72°C for 1 min per 1 kb. A final extension step was performed at 72°C for 10 minutes. PCRs were performed with Phusion DNA polymerase (New England Biolabs) and 2x Failsafe PCR buffer E (Epicentre Biotechnology). Splice-overlap extension (SOE)-PCR to re-target the Targetron was performed as described in the TargeTron Gene Knockout System users guide (Sigma-Aldrich) with modifications as previously described [43].
Table 2. Oligonucleotide primers.
| Primer | Sequence (5’- 3’) | Use |
|---|---|---|
| JRP5448 | AAAAAAGCTTATAATTATCCTTATAAAACCATTTCGTGCGCCCAGATAGGGTG | cdtR-288a-IBS (+) |
| JRP5449 | CAGATTGTACAAATGTGGTGATAACAGATAAGTCCATTTCTATAACTTACCTTTCTTTGT | cdtR-288a-EBS1d (-) |
| JRP5450 | TGAACGCAAGTTTCTAATTTCGGTTTTTTATCGATAGAGGAAAGTGTCT | cdtR-288a-EBS2 (+) |
| JRP3867 | CGAAATTAGAAACTTGCGTTCAGTAAAC | EBS universal (-) |
| JRP5632 | AAAAGGATCCCTTCTATAATTAGAAGTTAAATAATTCTTC | Amplify cdtR gene and upstream region, introduces BamHI site (+) |
| JRP5633 | AAAACTGCAGGAGACATCTCTTTTTTCTATTTATTATG | Amplify cdtR gene and upstream introduces PstI site (-) |
| DLP458 | TAATAAAAATACTGCCCTCGACAAA | tcdA-specific for RT-ddPCR (+) |
| DLP459 | ATAAATTGCATGTTGCTTCATAACT | tcdA-specific for RT-ddPCR (-) |
| JRP6107 | GCTATTAGCGAGGATAACGATTTC | tcdB-specific for RT-ddPCR (+) |
| JRP6108 | CTTTCCTAGTTCCATCATAAATCTACCA | tcdB-specific for RT-ddPCR (-) |
| JRP2443 | CAAGAAATAACTCAGTAGATGATTTGCAA | tcdR-specific for RT-ddPCR (+) |
| JRP2444 | TCTCCCTCTTCATAATGTAAAACTCTACTAAG | tcdR-specific for RT-ddPCR (-) |
| JRP6104 | AGCACAAAGGATATTGCTCTACT | tcdC-specific for RT-ddPCR (+) |
| JRP6105 | AAATGACCTCCTCATGGTCTTC | tcdC-specific for RT-ddPCR (-) |
| JRP3845 | TGCAATACTACTTACAAGGCTCCTATAGA | cdtA-specific for RT-ddPCR (+) |
| JRP3846 | TCTTTCCCATTCTTTAGCCTTTTC | cdtA-specific for RT-ddPCR (-) |
| JRP6238 | GATGCATGCTTTATTCGTGTACATA | cdtR-specific for RT-ddPCR (+) |
| JRP6239 | CGACATATATGGCCATTACTCATT | cdtR-specific for RT-ddPCR (-) |
| JRP2285 | GGATGATATGATGAAGGTTAGAAACCT | rpoA-specific for RT-ddPCR (+) |
| JRP2286 | CCCAATCCAAGTTCTTCTAGTTTTTG | rpoA-specific for RT-ddPCR (-) |
(+) = forward primer, (-) = reverse primer
Isolation and manipulation of nucleic acids
Plasmid DNA was isolated from E. coli, B. subtilis and C. difficile using QIAprep spin miniprep columns (Qiagen) following the manufacturer’s instructions. Genomic DNA was isolated from C. difficile as previously described [44]. Standard methods of DNA digestion, modification and ligation were used. DNA sequencing was carried out using BigDye Terminator v3.1 Ready Reaction Mix (Applied Biosystems) following the manufacturer’s instructions. Sequencing reactions were resolved on an Applied Biosystems 3730 DNA Analyzer. Sequences were analysed using ContigExpress (Invitrogen).
Construction of recombinant plasmids
All plasmids are outlined in Table 1. Construction of the cdtR TargeTron plasmid was performed as previously described, with some modifications [43]. Briefly, the group II intron from pDLL45 was retargeted by SOE-PCR to insert between nucleotides 288 and 289 of the cdtR gene using the primer pairs JRP5448 and JRP3867 and JRP5449 and JRP5450 (Table 2) to generate a 350 bp product, which was digested with BsrGI and HindIII and cloned into the corresponding sites of pDLL45, resulting in pJIR4135. A StuI-HindIII fragment was then sub-cloned from pJIR4135 into the corresponding sites of pDLL55, resulting in pJIR4153.
The cdtR complementation plasmid was constructed by PCR amplifying the cdtR gene and approximately 300 bp of its promoter region from C. difficile M7404 using the primers JRP5632 and JIR5633 (Table 2). The resulting 1.1 kb fragment was purified using a PCR purification kit (Qiagen) following the manufacturer’s instructions, digested with BamHI and PstI and cloned into the corresponding sites of pDLL24, resulting in pJIR4218.
Transfer of plasmid DNA into C. difficile by conjugation
Plasmid DNA was introduced into the B. subtilis conjugative donor strain BS34A as previously described [45]. The resulting strain was used as the donor for the conjugative transfer of plasmid DNA into C. difficile strains as before [11].
Isolation of cdtR mutants
C. difficile cdtR mutants were isolated using the method previously described [11] and confirmed by PCR and Southern hybridisation analysis. Complementation of the mutation was achieved using the cdtR complementation plasmid, pJIR4218. The cloning vector, pDLL24, was transferred into the cdtR mutant and the wild-type strain to construct vector (v) controls.
Detection of TcdA, TcdB, CDTa and CDTb by western blotting
Toxins were partially purified and concentrated eight-fold from 72 hour C. difficile TY culture supernatants by methanol-chloroform precipitation [11]. Protein concentrations were determined using the BCA protein assay kit (Pierce) as per the manufacturer’s instructions. Concentrated supernatant proteins (10 µg) were separated by 10% (v/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) [46] and transferred onto a nitrocellulose membrane (Whatman). Membranes were analysed as previously described [43]. TcdA and TcdB were detected using TcdA-specific monoclonal and TcdB-specific polyclonal antibodies (tgcBIOMICS), respectively. CDTa and CDTb were detected, respectively, using a CDTa-specific antibody and C. perfringens Ib-specific antibody that is cross reactive with CDTb [47]. CDTa, CDTb and TcdB-bound antibodies were detected using horseradish peroxidase conjugated anti-rabbit goat antibodies (Millipore) and TcdA-bound antibodies were detected using anti-mouse goat antibodies (Millipore). The Western Lightning Chemiluminescence reagent kit (Perkin-Elmer) was used to detect the bands, which were visualised by exposure to X-ray film or on a BioRad ChemiDoc XRS+ system.
ADP-ribosyltransferase assays
Toxins were partially purified from culture supernatants by precipitation with 70% ammonium sulphate as described previously [6]. ADP ribosyltransferase assays were performed as previously described [48]. Briefly, precipitated supernatant protein (50 µg) was incubated for 60 minutes at 37°C with 10 µg of actin in assay buffer (20 mM Tris-HCl pH 7.5, 1 µM dithiothreitol (DTT), 40 µM ATP, 40 µM CaCl2, 5 µM MgCl2) and 10 µM of biotinylated NAD+ (Trevigen). The reaction was heat inactivated at 95°C for 5 minutes in 4x SDS sample buffer (240 mM Tris-Cl (pH 6.8), 40% glycerol (v/v), 8% SDS (w/v), 5% (v/v) 2-mercaptoethanol, 0.05% (v/v) bromophenol blue and separated by 10% SDS-PAGE. Proteins were transferred onto a nitrocellulose membrane and biotinylated proteins were detected with horseradish peroxidase-conjugated streptavidin (GE Healthcare Life Sciences) and the Western Lightning Chemiluminescence reagent kit (Perkin-Elmer), following the manufacturer’s instructions. Relative band intensities were determined by densitometry using Image Lab Software (Bio-Rad). Data were analysed using GraphPad Prism 6 and statistical significance assessed using an unpaired t-test with a 95% confidence interval.
Vero and HT29 cell cytotoxicity assays
C. difficile strains were grown overnight in 20 ml of HIS broth with thiamphenicol and lincomycin, as required. The cultures then were used to inoculate 50 ml of TY broth with selection, such that each culture had a starting OD600 of approximately 0.05. Aliquots (5 ml) were taken at 12, 24, 48 and 72 hours, pelleted by centrifugation (10,000 g, 10 min, room temperature) and the supernatants filter sterilised through 0.45 µM and 0.2 µM filters (Sartorius). Supernatants were stored on ice until use. Vero cell and HT29 cell cytotoxicity assays were performed using the filtered C. difficile supernatants as previously described [43]. The levels of TcdA and TcdB produced by the C. difficile strains in vivo was assessed by determining the cytotoxicity of the intestinal contents collected 24 hours post infection against HT29 and Vero cells. Intestinal samples were resuspended in 100 mg/ml in PBS, diluted one in eight, filter sterilised and applied to Vero and HT29 cells, as described previously [2]. The endpoint (toxin titre) was scored as the last dilution with 100% cytopathic effect (CPE). Data were analysed using GraphPad Prism 6 and statistical significance assessed using an unpaired t-test with a 95% confidence interval.
RT-ddPCR analysis of C. difficile gene expression
Total RNA was extracted using TRIzol® (Life Technologies) following the manufacturer’s instructions. Forty ml of C. difficile TY broth cultures with an OD600 of approximately 0.3 for tcdC and cdtA expression analysis, and 10 ml of TY broth culture grown to OD600 of approximately 1.8 for tcdA, tcdB and tcdR expression analysis, were used. A total of 200 ng of RNA was converted to cDNA using SuperScript III Reverse Transcriptase (Life Technologies), following the manufacturer’s instructions. Transcript levels were quantified using the QX200 Droplet Digital PCR System (BioRad) using QX200 ddPCR EvaGreen Supermix and 0.1–5 µg of total cDNA and specific primers (Table 2) at a concentration of 200 nM. Transcription levels of each gene was normalised to transcription levels of the housekeeping gene rpoA. Data were analysed using GraphPad Prism 6 and statistical significance assessed using a Mann Whitney U test.
C. difficile virulence trials
Groups of five male six to eight week old C57BL/6 mice were used in C. difficile virulence trials as previously described [2], except mice were switched back to plain drinking water on the day of infection. Mice were administered 106 C. difficile spores by oral gavage and were humanely euthanised at the onset of severe disease or at the end of the experiment, as previously defined [2]. Animal handling and experimentation were performed in accordance with institutional guidelines (Monash University animal ethics committee numbers MARP/2014/135 and SOBSB/M/2010/25). Faecal samples were taken daily to monitor C. difficile shedding using HIS agar supplemented with 0.1% (w/v) cysteine, 0.1% (w/v) taurocholate, 0.375% (w/v) glucose, 250 µg/ml D-cycloserine, 8 µg/ml cefoxitin, 10 µg/ml erythromycin, 12 µg/ml norfloxacin, 32 µg/ml moxalactam. Data were analysed using GraphPad Prism 6 and statistical significance assessed using a log-rank (Mantel-Cox) test.
Histology
The entire colon and caecum were collected from each mouse and Swiss-rolled [49] prior to fixation to allow for cross-sectional examination of the entire length of the colon. Tissues were stained with PAS-Alcian blue and histopathological assessment of damage and scoring of tissues was performed blind by independent observers using a previously defined set of parameters [2].
Supporting Information
Schematic diagram of the cdtR genomic location and surrounding genes in (a) wild-type M7404 or R20291 and (b) TargeTron-derived cdtR insertion mutants. Southern hybridisation using a cdtR specific probe (red) showed a size increase from a 3.2 kb AvaII fragment in the wild type (black arrow) to a 5.0 kb AvaII fragment in the independent cdtR mutants (white arrow) in both (c) M7404 and (e) R20291 strain backgrounds. Hybridization of an ermB probe (purple) to a 5.0 kb AvaII fragment (black arrow) in the (d) M7404 cdtR mutants and (f) R20291 cdtR mutant confirmed the TargeTron insertion.
(TIF)
Colonisation efficiencies are shown as total colony forming unit (CFU) of C. difficile isolated per gram of faeces collected from mice at 24 and 48 hours. Mice surviving beyond 48 hours had similar levels of colonisation. Data represent the mean ± SEM (n = 6–15).
(TIF)
Western immunoblots were performed using precipitated supernatant proteins from the CD37 non-toxigenic strain, two 630 vector control strains and two 630 strains carrying the cdtR + complementation vector, pJIR4218, and detected using antibodies specific for (a) TcdA and (b) TcdB. c, Expression of cdtR in 630, 630 carrying the vector control and 630 carrying the cdtR + complementation vector normalised to rpoA expression.
(TIF)
Faecal samples collected from uninfected and C. difficile infected mice 24 hours post infection were assayed for cytotoxicity by doubling dilution cytotoxicity assays using (a) HT29 cells and (b) Vero cells. Data represent the mean ± SEM (n = 5).
(TIF)
Acknowledgments
We thank M. Popoff for providing CDTa-specific and Ib-specific antibodies, P. Howarth for technical assistance, S. Larcombe for the construction of pDLL24, S. Mileto for helpful discussions and B. Cunningham for construction of DLL3094 and technical assistance.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by grants from the Australian National Health and Medical Research Council (GNT1065985, awarded to JIR, GNT1051584, awarded to DL, and GNT1051586, awarded to DL). DL was supported by a research fellowship from the Australian Research Council (FT120100779). SAL was the recipient of an Australian Postgraduate Award. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Lyras D, O'Connor JR, Howarth PM, Sambol SP, Carter GP, et al. (2009) Toxin B is essential for virulence of Clostridium difficile . Nature 458: 1176–1179. 10.1038/nature07822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Carter GP, Chakravorty A, Pham Nguyen TA, Mileto S, Schreiber F, et al. (2015) Defining the Roles of TcdA and TcdB in Localized Gastrointestinal Disease, Systemic Organ Damage, and the Host Response during Clostridium difficile Infections. MBio 6: e00551 10.1128/mBio.00551-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Awad MM, Johanesen PA, Carter GP, Rose E, Lyras D (2014) Clostridium difficile virulence factors: Insights into an anaerobic spore-forming pathogen. Gut Microbes 5: 579–593. 10.4161/19490976.2014.969632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Rupnik M, Janezic S (2016) An Update on Clostridium difficile Toxinotyping. J Clin Microbiol 54: 13–18. 10.1128/JCM.02083-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Yan Q, Zhang J, Chen C, Zhou H, Du P, et al. (2013) Multilocus sequence typing (MLST) analysis of 104 Clostridium difficile strains isolated from China. Epidemiol Infect 141: 195–199. 10.1017/S0950268812000453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Carter GP, Lyras D, Allen DL, Mackin KE, Howarth PM, et al. (2007) Binary toxin production in Clostridium difficile is regulated by CdtR, a LytTR family response regulator. J Bacteriol 189: 7290–7301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Goncalves C, Decre D, Barbut F, Burghoffer B, Petit JC (2004) Prevalence and characterization of a binary toxin (actin-specific ADP-ribosyltransferase) from Clostridium difficile . J Clin Microbiol 42: 1933–1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Snydman DR, McDermott LA, Jacobus NV, Thorpe C, Stone S, et al. (2015) U.S.-Based National Sentinel Surveillance Study for the Epidemiology of Clostridium difficile-Associated Diarrheal Isolates and Their Susceptibility to Fidaxomicin. Antimicrob Agents Chemother 59: 6437–6443. 10.1128/AAC.00845-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Schwan C, Stecher B, Tzivelekidis T, Van Ham M, Rohde M, et al. (2009) Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Pathog 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Baban ST, Kuehne SA, Barketi-Klai A, Cartman ST, Kelly ML, et al. (2013) The role of flagella in Clostridium difficile pathogenesis: comparison between a non-epidemic and an epidemic strain. PLoS One 8: e73026 10.1371/journal.pone.0073026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Mackin KE, Carter GP, Howarth P, Rood JI, Lyras D (2013) Spo0A differentially regulates toxin production in evolutionarily diverse strains of Clostridium difficile . PLoS One 8: e79666 10.1371/journal.pone.0079666 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Carter GP, Douce GR, Govind R, Howarth PM, Mackin KE, et al. (2011) The anti-sigma factor TcdC modulates hypervirulence in an epidemic BI/NAP1/027 clinical isolate of Clostridium difficile . PLoS Pathog 7: e1002317 10.1371/journal.ppat.1002317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Cartman ST, Kelly ML, Heeg D, Heap JT, Minton NP (2012) Precise manipulation of the Clostridium difficile chromosome reveals a lack of association between the tcdC genotype and toxin production. Appl Environ Microbiol 78: 4683–4690. 10.1128/AEM.00249-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Matamouros S, England P, Dupuy B (2007) Clostridium difficile toxin expression is inhibited by the novel regulator TcdC. Mol Microbiol 64: 1274–1288. [DOI] [PubMed] [Google Scholar]
- 15. Mani N, Lyras D, Barroso L, Howarth P, Wilkins T, et al. (2002) Environmental response and autoregulation of Clostridium difficile TxeR, a sigma factor for toxin gene expression. J Bacteriol 184: 5971–5978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Saujet L, Monot M, Dupuy B, Soutourina O, Martin-Verstraete I (2011) The key sigma factor of transition phase, SigH, controls sporulation, metabolism, and virulence factor expression in Clostridium difficile . J Bacteriol 193: 3186–3196. 10.1128/JB.00272-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. McKee RW, Mangalea MR, Purcell EB, Borchardt EK, Tamayo R (2013) The second messenger cyclic Di-GMP regulates Clostridium difficile toxin production by controlling expression of sigD . J Bacteriol 195: 5174–5185. 10.1128/JB.00501-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Martin MJ, Clare S, Goulding D, Faulds-Pain A, Barquist L, et al. (2013) The agr locus regulates virulence and colonization genes in Clostridium difficile 027. J Bacteriol 195: 3672–3681. 10.1128/JB.00473-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Antunes A, Martin-Verstraete I, Dupuy B (2011) CcpA-mediated repression of Clostridium difficile toxin gene expression. Mol Microbiol 79: 882–899. 10.1111/j.1365-2958.2010.07495.x [DOI] [PubMed] [Google Scholar]
- 20. Dineen SS, McBride SM, Sonenshein AL (2010) Integration of metabolism and virulence by Clostridium difficile CodY. J Bacteriol 192: 5350–5362. 10.1128/JB.00341-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Loo VG, Poirier L, Miller MA, Oughton M, Libman MD, et al. (2005) A predominantly clonal multi-institutional outbreak of Clostridium difficile—Associated diarrhea with high morbidity and mortality. N Engl J Med 353: 2442–2449. [DOI] [PubMed] [Google Scholar]
- 22. Warny M, Pepin J, Fang A, Killgore G, Thompson A, et al. (2005) Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 366: 1079–1084. [DOI] [PubMed] [Google Scholar]
- 23. McDonald LC, Killgore GE, Thompson A, Owens RC Jr, Kazakova SV, et al. (2005) An epidemic, toxin gene-variant strain of Clostridium difficile . N Engl J Med 353: 2433–2441. [DOI] [PubMed] [Google Scholar]
- 24. El Meouche I, Peltier J, Monot M, Soutourina O, Pestel-Caron M, et al. (2013) Characterization of the SigD regulon of C. difficile and its positive control of toxin production through the regulation of tcdR . PLoS One 8: e83748 10.1371/journal.pone.0083748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Keel K, Brazier JS, Post KW, Weese S, Songer JG (2007) Prevalence of PCR ribotypes among Clostridium difficile isolates from pigs, calves, and other species. Journal of Clinical Microbiology 45: 1963–1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Goorhuis A, Bakker D, Corver J, Debast SB, Harmanus C, et al. (2008) Emergence of Clostridium difficile infection due to a new hypervirulent strain, polymerase chain reaction ribotype 078. Clinical Infectious Diseases 47: 1162–1170. 10.1086/592257 [DOI] [PubMed] [Google Scholar]
- 27. Bouvet PJ, Popoff MR (2008) Genetic relatedness of Clostridium difficile isolates from various origins determined by triple-locus sequence analysis based on toxin regulatory genes tcdC, tcdR, and cdtR . J Clin Microbiol 46: 3703–3713. 10.1128/JCM.00866-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Sebaihia M, Wren B. W., Mullany P., Fairweather N. F., Minton N., Stabler NRT R., Roberts A. P., Cerdeno-Tarraga A. M., Wang H., Holden AW M. T., Churcher C., Quail M. A., Baker S., Bason N., Brooks TC K., Cronin A., Davis P., Dowd L., Fraser A., Feltwell ZH T., Holroyd S., Jagels K., Moule S., Mungall K., Price, E. C., et al. (2006) The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat Gen 38: 779–786. [DOI] [PubMed] [Google Scholar]
- 29. Merrigan M, Venugopal A, Mallozzi M, Roxas B, Viswanathan VK, et al. (2010) Human hypervirulent Clostridium difficile strains exhibit increased sporulation as well as robust toxin production. J Bacteriol 192: 4904–4911. 10.1128/JB.00445-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Stabler RA, He M, Dawson L, Martin M, Valiente E, et al. (2009) Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium. Genome Biol 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Vohra P, Poxton IR (2011) Comparison of toxin and spore production in clinically relevant strains of Clostridium difficile . Microbiology 157: 1343–1353. 10.1099/mic.0.046243-0 [DOI] [PubMed] [Google Scholar]
- 32. Geric B, Carman RJ, Rupnik M, Genheimer CW, Sambol SP, et al. (2006) Binary toxin-producing, large clostridial toxin-negative Clostridium difficile strains are enterotoxic but do not cause disease in hamsters. J Infect Dis 193: 1143–1150. [DOI] [PubMed] [Google Scholar]
- 33. Schwan C, Kruppke AS, Nolke T, Schumacher L, Koch-Nolte F, et al. (2014) Clostridium difficile toxin CDT hijacks microtubule organization and reroutes vesicle traffic to increase pathogen adherence. Proc Natl Acad Sci U S A 111: 2313–2318. 10.1073/pnas.1311589111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Metcalf DS, Weese JS (2011) Binary toxin locus analysis in Clostridium difficile . J Med Microbiol 60: 1137–1145. 10.1099/jmm.0.028498-0 [DOI] [PubMed] [Google Scholar]
- 35. Antunes A, Camiade E, Monot M, Courtois E, Barbut F, et al. (2012) Global transcriptional control by glucose and carbon regulator CcpA in Clostridium difficile . Nucleic Acids Res 40: 10701–10718. 10.1093/nar/gks864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Darkoh C, DuPont HL, Norris SJ, Kaplan HB (2015) Toxin synthesis by Clostridium difficile is regulated through quorum signaling. MBio 6: e02569 10.1128/mBio.02569-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Knight DR, Elliott B, Chang BJ, Perkins TT, Riley TV (2015) Diversity and Evolution in the Genome of Clostridium difficile . Clin Microbiol Rev 28: 721–741. 10.1128/CMR.00127-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Elliott B, Dingle KE, Didelot X, Crook DW, Riley TV (2014) The complexity and diversity of the Pathogenicity Locus in Clostridium difficile clade 5. Genome Biol Evol 6: 3159–3170. 10.1093/gbe/evu248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Stabler RA, Gerding DN, Songer JG, Drudy D, Brazier JS, et al. (2006) Comparative phylogenomics of Clostridium difficile reveals clade specificity and microevolution of hypervirulent strains. J Bacteriol 188: 7297–7305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Dingle KE, Griffiths D, Didelot X, Evans J, Vaughan A, et al. (2011) Clinical Clostridium difficile: clonality and pathogenicity locus diversity. PLoS One 6: e19993 10.1371/journal.pone.0019993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Sambrook J, Russell David W (1989) Molecular cloning: a laboratory manual Vol. 3: Cold spring harbor laboratory press. [Google Scholar]
- 42. Smith CJ, Markowitz SM, Macrina FL (1981) Transferable tetracycline resistance in Clostridium difficile . Antimicrobial Agents and Chemotherapy 19: 997–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Carter GP, Larcombe S, Li L, Jayawardena D, Awad MM, et al. (2014) Expression of the large clostridial toxins is controlled by conserved regulatory mechanisms. International Journal of Medical Microbiology. [DOI] [PubMed] [Google Scholar]
- 44. O'Connor JR, Lyras D, Farrow KA, Adams V, Powell DR, et al. (2006) Construction and analysis of chromosomal Clostridium difficile mutants. Molecular Microbiology 61: 1335–1351. [DOI] [PubMed] [Google Scholar]
- 45. Anagnostopoulos C, Spizizen J (1961) Requirements for Transformation in Bacillus subtilis . J Bacteriol 81: 741–746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680–685. [DOI] [PubMed] [Google Scholar]
- 47. Perelle S, Gibert M, Bourlioux P, Corthier G, Popoff MR (1997) Production of a complete binary toxin (actin-specific ADP- ribosyltransferase) by Clostridium difficile CD196. Infect Immun 65: 1402–1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Yonogi S, Matsuda S, Kawai T, Yoda T, Harada T, et al. (2014) BEC, a novel enterotoxin of Clostridium perfringens found in human clinical isolates from acute gastroenteritis outbreaks. Infect Immun 82: 2390–2399. 10.1128/IAI.01759-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Moolenbeek C, Ruitenberg EJ (1981) The "Swiss roll": a simple technique for histological studies of the rodent intestine. Lab Anim 15: 57–59. [DOI] [PubMed] [Google Scholar]
- 50. Backman K, Ptashne M, Gilbert W (1976) Construction of plasmids carrying the cI gene of bacteriophage lambda. Proc Natl Acad Sci U S A 73: 4174–4178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Roberts AP, Hennequin C, Elmore M, Collignon A, Karjalainen T, et al. (2003) Development of an integrative vector for the expression of antisense RNA in Clostridium difficile . J Microbiol Meth 55: 617–624. [DOI] [PubMed] [Google Scholar]
- 52. Smith CJ, Markowitz SM, Macrina FL (1981) Transferable tetracycline resistance in Clostridium difficile . Antimicrob Agents Chemother 19: 997–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Schematic diagram of the cdtR genomic location and surrounding genes in (a) wild-type M7404 or R20291 and (b) TargeTron-derived cdtR insertion mutants. Southern hybridisation using a cdtR specific probe (red) showed a size increase from a 3.2 kb AvaII fragment in the wild type (black arrow) to a 5.0 kb AvaII fragment in the independent cdtR mutants (white arrow) in both (c) M7404 and (e) R20291 strain backgrounds. Hybridization of an ermB probe (purple) to a 5.0 kb AvaII fragment (black arrow) in the (d) M7404 cdtR mutants and (f) R20291 cdtR mutant confirmed the TargeTron insertion.
(TIF)
Colonisation efficiencies are shown as total colony forming unit (CFU) of C. difficile isolated per gram of faeces collected from mice at 24 and 48 hours. Mice surviving beyond 48 hours had similar levels of colonisation. Data represent the mean ± SEM (n = 6–15).
(TIF)
Western immunoblots were performed using precipitated supernatant proteins from the CD37 non-toxigenic strain, two 630 vector control strains and two 630 strains carrying the cdtR + complementation vector, pJIR4218, and detected using antibodies specific for (a) TcdA and (b) TcdB. c, Expression of cdtR in 630, 630 carrying the vector control and 630 carrying the cdtR + complementation vector normalised to rpoA expression.
(TIF)
Faecal samples collected from uninfected and C. difficile infected mice 24 hours post infection were assayed for cytotoxicity by doubling dilution cytotoxicity assays using (a) HT29 cells and (b) Vero cells. Data represent the mean ± SEM (n = 5).
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.