

Abstract
A previous genome-wide association study (GWAS) of more than 100,000 individuals identified molecular-genetic predictors of educational attainment. We undertook in-depth life-course investigation of the polygenic score derived from this GWAS using the four-decade Dunedin Study (N = 918). There were five main findings. First, polygenic scores predicted adult economic outcomes even after accounting for educational attainments. Second, genes and environments were correlated: Children with higher polygenic scores were born into better-off homes. Third, children’s polygenic scores predicted their adult outcomes even when analyses accounted for their social-class origins; social-mobility analysis showed that children with higher polygenic scores were more upwardly mobile than children with lower scores. Fourth, polygenic scores predicted behavior across the life course, from early acquisition of speech and reading skills through geographic mobility and mate choice and on to financial planning for retirement. Fifth, polygenic-score associations were mediated by psychological characteristics, including intelligence, self-control, and interpersonal skill. Effect sizes were small. Factors connecting DNA sequence with life outcomes may provide targets for interventions to promote population-wide positive development.
Keywords: genetics, behavior genetics, intelligence, personality, adult development
In 2013, scientists reported the first successful genome-wide association study (GWAS) of a social-science outcome, educational attainment (Rietveld et al., 2013). Their analysis of millions of genetic variants in more than 100,000 individuals hinted at the existence of a molecular map to success in schooling written in the alphabet of DNA. As anticipated, rather than finding a so-called gene for education, this study revealed a genetic continuum: Some individuals carry very few alleles associated with educational attainment, the bulk of the population carries some such alleles, and a few people carry many. This continuum, measured as a polygenic score (Chabris, Lee, Cesarini, Benjamin, & Laibson, 2015), has since been shown to predict educational attainment in cohorts on three continents and even differences in educational attainment between siblings in the same family (Conley et al., 2015; de Zeeuw et al., 2014; Domingue, Belsky, Conley, Harris, & Boardman, 2015; Rietveld, Esko, et al., 2014; Ward et al., 2014). Although the magnitudes of associations are small, these findings have provoked controversy and concern about misuse and misinterpretation (Henig, 2015). In an effort to provide an empirical foundation for productive public discussion of the new science of sociogenomics, we ask three questions in the current article: (a) Do genetic discoveries for educational attainment predict outcomes beyond schooling? (b) If so, what are the developmental and behavioral pathways that connect differences in DNA sequences with divergent life outcomes? (c) Do psychological characteristics act as mediators of genetic associations? Although these questions may seem premature, it is important to ask them now, before technologies using genetics to predict social outcomes become possible.
These questions were addressed by examination of data prospectively collected from a population-representative birth cohort followed through midlife, the Dunedin Study (Poulton, Moffitt, & Silva, 2015). Across 13 repeated in-person assessments, Dunedin Study members were evaluated for developmental milestones in childhood; for traits, behaviors, and aspirations through adolescence; and ultimately for attainments and outcomes in adulthood (Table 1). Because attrition has been minimal (5% at the latest wave in 2012), the findings illustrate genetic associations with life courses and life outcomes without bias from selective attrition as a result of illness or challenging life circumstances. We tested a series of hypotheses about the scope, pathways, and psychological mechanisms of genetic influence on socioeconomic attainments across the first half of the life course. We tracked a deeply phenotyped cohort from early childhood through midlife, examining preselected developmentally appropriate manifestations of achievement-related behaviors. We report a large number of outcome variables in order to provide a complete account of these data. In the interest of reproducibility the analysis plan was posted in advance.
Table 1.
Tracking the Development of Socioeconomic Success
| Phenotype | Measure or data source | Age |
|---|---|---|
| Success in schooling | ||
| Highest degree | Structured interview | 15–38 |
| Success beyond schooling | ||
| Adult-attainment factor | Occupation (prestige score based on NZ Census data), income, assets, credit-problems scale, difficulty-paying-expenses scale, days of social-welfare-benefit use (NZ Social Welfare Administration), credit score (Veda credit bureau) | 38 |
| Social mobility | Childhood social class based on parental occupation; adult attainment measured using education, occupation, and the adult-attainment factor | Birth–15, 38 |
| Pathways to success | ||
| Developmental milestones | Interviews with mothers | 3 |
| Reading ability | Burt Word Reading Test (Scottish Council for Research in Education, 1976) | 7–18 |
| Aspirations | Questionnaire | 15 |
| Standardized testing | NZ Ministry of Education test record | 18 |
| Geographic mobility | Life-history calendar interview | 21–38 |
| Financial planfulness | Structured interview and informant reports | 32–38 |
| Mate selection | Structured interview in which Dunedin Study members reported their relationship status and, for those in a serious relationship, their partners’ highest educational degree and income | 38 |
| Skills and abilities | ||
| Cognitive ability | Peabody Picture Vocabulary Test (Dunn, 1965), Stanford-Binet Intelligence Scale (Terman & Merrill, 1960), Wechsler Intelligence Scales for Children– Revised (Wechsler, 1974) | 3–13 |
| Self-control skills | Staff observations, parent and teacher reports, and interviews with Dunedin Study members | 3–11 |
| Interpersonal skill | Staff observations | 3–9 |
| Physical health | Medical exams, anthropometry, lung function testing, clinical interviews with parents | 3–11 |
Note: NZ = New Zealand.
Method
Sample
Participants were members of the Dunedin Study, a longitudinal investigation of health and behavior in a complete birth cohort. Dunedin Study members (N = 1,037; 91% of eligible births; 52% male) were all individuals born between April 1972 and March 1973 in Dunedin, New Zealand, who were eligible on the basis of residence in the province and who participated in the first assessment at age 3. The cohort represented the full range of socioeconomic status (SES) in the general population of New Zealand’s South Island. On adult health, the cohort matched the New Zealand National Health and Nutrition Survey (e.g., body mass index, smoking, visits to the doctor; Poulton et al., 2015). The cohort was primarily White; fewer than 7% self-identified as having non-European ancestry, matching the population of the South Island (Poulton et al., 2015). Assessments were carried out at birth and at ages 3, 5, 7, 9, 11, 13, 15, 18, 21, 26, 32, and, most recently, 38 years, when 95% of the 1,007 Dunedin Study members still alive took part. At each assessment, each Dunedin Study member was brought to the research unit for a full day of interviews and examinations.
Genotyping and imputation
We used Illumina HumanOmni Express 12 BeadChip arrays (Version 1.1; Illumina, Hayward, CA) to assay common single-nucleotide polymorphism (SNP) variation in the genomes of our cohort members. We imputed additional SNPs using the impute2 software (Version 2.3.1; https://mathgen.stats.ox.ac.uk/impute/impute_v2.html; Howie, Donnelly, & Marchini, 2009) and the 1000 Genomes Phase 3 reference panel (1000 Genomes Project, 2016). Imputation was conducted on autosomal SNPs appearing in dbSNP (Version 140; http://www.ncbi.nlm.nih.gov/SNP/; Sherry et al., 2001) that were “called” in more than 98% of the Dunedin Study samples. Invariant SNPs were excluded. Prephasing and imputation were conducted using a 50-million-base-pair sliding window. The resulting genotype database included genotyped SNPs and SNPs imputed with 90% probability of a specific genotype among the non-Maori members of the Dunedin cohort (n = 918). We analyzed SNPs in Hardy-Weinberg equilibrium (p > .01).
Polygenic scoring
We calculated polygenic scores according to the method described by Dudbridge (2013) using the PRsice software (Version 1.22; http://prsice.info/; Euesden, Lewis, & O’Reilly, 2015). To calculate the polygenic score for educational attainment, we matched genotypes from our data with GWAS results for educational attainment reported by the Social Science Genetic Association Consortium (Rietveld et al., 2013) and used the approximately 2.3 million matched genotypes to score Dunedin Study members’ genetic predisposition to educational attainment.
For each genotype, we counted the number of education-associated alleles (0, 1, or 2) and multiplied this count by the effect size estimated in the original GWAS. (Most genotypes had effect sizes very near 0.) We then summed weighted counts across all genotypes to calculate each Dunedin Study member’s score. We used all matched SNPs to compute polygenic scores, irrespective of nominal significance for their association with educational attainment. Scores ranged from −30.51 to 73.77 (M = 17.73, SD = 17.94) and were normally distributed in the Dunedin birth cohort. We standardized scores so that the mean was zero and the standard deviation was 1 (see Fig. S1 in the Supplemental Material available online). Given the original GWAS results, Dunedin Study members with polygenic scores greater than 0 would be expected to complete more years of schooling, and Dunedin Study members with polygenic scores below 0 would be expected to complete fewer years of schooling. We used the same method to calculate polygenic scores for height (based on results from the Genetic Investigation of Anthropometric Traits Consortium’s most recent GWAS of height; Wood et al., 2014). To account for potential population stratification, we adjusted polygenic score analyses for the first 10 principal components computed from the genome-wide SNP data using the EIGENSOFT smartPCA tool (Version 5.0.2; http://www.hsph.harvard.edu/alkes-price/software/; Price et al., 2006; Price, Zaitlen, Reich, & Patterson, 2010).
Measurement of life-course-development phenotypes
More detailed descriptions of study measures described later in this section and relevant citations are provided in the Supplemental Material.
Social-class origins
We measured social class origins as the average SES across repeated assessments throughout Dunedin Study members’ childhoods. SES was determined from the higher of either parent’s occupational status throughout the Dunedin Study members’ childhoods.
Attainment
We measured educational attainment as the highest degree completed by a Dunedin Study member through the time of the age-38 assessment. We measured attainment beyond education from Dunedin Study members’ reports of their occupation, income, assets, credit problems, and difficulties paying expenses when they were 38 years old and from electronic record searches of social-welfare and credit-score databases.
Pathways to success
We measured the age at which Dunedin Study members achieved early developmental milestones on the basis of data gathered from interviews with their mothers when the members were 3 years old. We measured reading ability from scores on the Burt Word Reading Test (Scottish Council for Research in Education, 1976), taken when Dunedin Study members were 7, 9, 11, 13, 15, and 18 years old. We measured educational and socioeconomic aspirations from surveys completed by the Dunedin Study members at the age of 15. We measured academic performance from scores on standardized tests taken at the ages of 15 to 18. We measured geographic mobility from member life-history calendar reports about place of work and residence from the ages of 21 to 38. We measured financial planfulness on the basis of data gathered from surveys of Dunedin Study members’ friends and relatives and from structured interviews with the members themselves when they were 32 and 38 years old. We measured the SES of members’ romantic partners from members’ reports of their partners’ income and education in structured interviews conducted when the members were 38 years old.
Life satisfaction
When they were 38 years old, Dunedin Study members completed a five-item Satisfaction With Life scale (e.g., “In most ways my life is close to ideal,” “So far I have gotten the important things I want in life”; Pavot & Diener, 1993).
Traits and abilities
We measured cognitive ability and cognitive development using the Peabody Picture Vocabulary Test (Dunn, 1965), administered when Dunedin Study members were 3 years old; the Stanford-Binet Intelligence Scale (Terman & Merrill, 1960), administered when members were 5 years old; and the Wechsler Intelligence Scales for Children–Revised (WISC-R; Wechsler, 1974), administered when members were 7 to 13 years old. We measured Dunedin Study members’ childhood self-control skills from observational ratings of their lack of control (when they were 3 and 5 years old) and parent, teacher, and self-reports of impulsive aggression, hyperactivity, lack of persistence, inattention, and impulsivity (when they were 5–11 years old). We measured Dunedin Study members’ childhood interpersonal skills from reports made by trained research workers after standardized testing sessions when they were 3 to 9 years old. We measured childhood health from medical exams, anthropometry, lung function testing, and interviews with parents at assessments made between Dunedin Study members’ birth and the age of 11.
Height
Study members’ height at age 38 was measured to the nearest millimeter using a stadiometer (Harpenden; Holtain, Ltd., Crosswell, Wales).
Ethical approvals
The study protocol was approved by the institutional ethical review boards of the participating universities. Dunedin Study members gave informed consent before participating. The Otago University ethics committee provided ethical approval for the Dunedin Study. Participants gave written consent before data were collected. When participants were children, their parents gave informed consent.
Data sharing
Dunedin Study data are available to researchers on application. A managed-access process ensures that approval is granted to research that comes under the terms of participant consent and privacy (see the Supplemental Material for data-sharing details).
Statistical analysis
We analyzed continuous dependent variables using linear regression models to estimate standardized regression coefficients (reported as Pearson’s r). We analyzed dichotomous dependent variables using Poisson regression models to estimate relative risks (RRs). We analyzed time-to-event data for developmental milestones using Cox models to estimate hazard ratios. We analyzed ordered categorical outcomes using ordered logit models to estimate odds ratios. We analyzed repeated measures longitudinal data on reading ability and cognitive development using multilevel longitudinal growth models (Singer & Willett, 2003). Finally, we conducted mediation analyses using the system of equations described by Baron and Kenny (1986) and the methods described by Preacher and his colleagues (Preacher & Hayes, 2008; Preacher & Kelley, 2011) to calculate total, direct, and indirect effects and to estimate the proportion of effects mediated by each of the mediators. Growth model and mediation analyses are described further in the Supplemental Material. All models were adjusted for sex.
Results
Analyses included the 918 non-Maori Dunedin Study members who provided DNA samples. Cohort members’ genomes were scored according to published GWAS results for educational attainment (Rietveld et al., 2013; polygenic scores were standardized so that they had a mean of 0 and a standard deviation of 1; see Fig. S1 in the Supplemental Material). The analyses proceeded in three parts. Part 1 analyses examined divergent outcomes of high- and low-scoring children, first in education and then in the acquisition of social and economic capital through midlife and the social mobility those attainments reflected. Part 2 analyses examined how higher-scoring children came to grow apart from their lower-scoring peers. Analysis tested genetic differences in the timing of early-life milestones; in the age at which children learned to read; in the decision to test for secondary-education credentials and university enrollment, and performance on those tests; in geographic mobility in search of training and employment; and in selection of mates, formation of households, and forging of careers. Part 3 analyses examined candidate psychological characteristics through which genetic influences on development and life outcomes might come about.
Part 1: What did discovered genetics of educational attainment mean for life outcomes beyond schooling?
Part 1 analyses tested the hypothesis that Dunedin Study members’ polygenic scores would predict their life attainments at the age of 38, roughly the midpoint in the human life span. All analyses were adjusted for the first 10 principal components computed from genome-wide SNP data (see Table S1 in the Supplemental Material) to adjust for potential population stratification (i.e., genome-wide patterning of differences in allele frequency that might induce spurious correlations between the polygenic score and study outcomes). Unadjusted estimates are reported in Table S1 in the Supplemental Material.
Did individuals with higher polygenic scores achieve higher degrees?
In replication of the original discovery about the genetics of educational attainment, our results showed that Dunedin cohort members with higher polygenic scores tended to go on to achieve higher degrees compared with peers who had lower scores (r = .15, p < .001; Fig. 1a). This correlation between polygenic score and educational attainment was nearly identical to the estimate from the original report (Rietveld et al., 2013). As in previous studies, the genetic effect was small in magnitude; for example, having a polygenic score 1 standard deviation above the mean was associated with a 19% increase in likelihood of completing a university degree (RR = 1.19, 95% confidence interval (CI) = [1.07, 1.32]).
Fig. 1.

Association between polygenic score and educational and adult achievement. In (a), mean polygenic score is graphed as a function of educational attainment. Error bars represent 95% confidence intervals. For the 1972–73 birth cohort we studied, compulsory education ended at age 15 years, at which point students could elect to take a School Leaving Certificate exam. Fifteen percent of our sample obtained no educational credential; 15% obtained the School Leaving Certificate but did not progress further; 42% completed sixth-form or Bursary Certificates (roughly equivalent to a full high school diploma in the United States); and 29% completed a university degree. In (b), the scatterplot (with best-fitting regression line) shows the relationship between Dunedin Study members’ polygenic scores (x-axis) and their adult-attainment-factor z scores (y-axis). The adult attainment factor was composed of occupational prestige, income, assets, credit problems, difficulties paying expenses, social-welfare-benefit use, and credit score. Each plotted point represents mean x and y coordinates for a bin of 10 Dunedin Study members.
Did individuals with higher polygenic scores go on to achieve socioeconomic success beyond schooling?
Adult socioeconomic attainments of Dunedin Study members were measured using data from structured interviews about jobs, income, wealth, and financial difficulties and by conducting administrative-record searches of governmental and credit-bureau databases. Factor analysis of these multiple measures was used to compute an adult-attainment-factor score (see Table S2 and Fig. S2 in the Supplemental Material). By midlife, individuals with higher polygenic scores tended to be more socioeconomically successful: They held more prestigious occupations, earned higher incomes, had accumulated more assets, reported fewer difficulties paying their expenses, relied less on social-welfare benefits, and had higher credit scores (r = .13, p < .001 for the adult-attainment factor; Fig. 1b). It may seem unsurprising that a polygenic score that predicted educational attainment also continued to predict success after education was complete. However, less than half of the genetic association with adult attainment was accounted for by educational attainment; when we repeated our genetic analysis of the adult-attainment factor and included education as a covariate, the adjusted effect size was .07 (p = .035). Genetic effect sizes for the individual attainment measures and effect sizes after adjustment for educational attainment are shown in Fig. S3 in the Supplemental Material.
In sum, in the Dunedin cohort, individuals with higher polygenic scores tended to grow up to become more successful, not only in schooling, but also in their economic and professional lives. This success depended only partly on their educational attainment.
Were children with higher polygenic scores more often born into socially advantaged families?
In previous research, the correlation between parent and offspring polygenic scores was estimated to be approximately .6 (Conley et al., 2015). Moreover, if a generation of individuals who achieve more occupational and economic success carry a certain genotype or set of genotypes, it stands to reason that their own children will inherit not only their genetics, but also their social success. This hypothesis of social stratification of genotypes was tested by comparing polygenic scores of children whose parents occupied different social positions. Parents’ SES was measured from repeated assessments conducted when the cohort members were growing up (i.e., during their first 15 years of life; see the Supplemental Material). Our findings point to a gene-environment correlation: The polygenic score for educational attainment was stratified by childhood SES such that children with higher polygenic scores tended to have grown up in families with higher SES, whereas children with lower polygenic scores tended to have grown up in families with lower SES (r = .13, p < .001).
Were children with higher polygenic scores more likely to achieve upward social mobility?
Analyses of social mobility tested whether the higher life attainments of children with higher polygenic scores were independent of their social origins. The analysis of adult socioeconomic outcomes was repeated, but with the addition of a statistical control for the SES of a child’s family during his or her first 15 years of life (see the Supplemental Material). Three interrelated outcomes were considered: the Dunedin Study member’s educational attainment; their attained adult SES, measured as occupational prestige (in parallel to the status of their parents); and their adult-attainment-factor score. Children with higher polygenic scores tended to attain more regardless of whether they began life in a family that was well-off or one that was socially disadvantaged (more education: r = .10, p = .002; more prestigious occupations: r = 0.11, p < .001; higher adult-attainment-factor scores: r = .11, p = .002). Figure 2 shows associations between the polygenic score and adult attainment in groups of Dunedin Study members with low, middle, and high childhood SES.
Fig. 2.

Scatterplots showing the association between Dunedin Study members’ polygenic scores and their adult-attainment-factor z scores, separately for children born in low-, middle-, and high-socioeconomic status (SES) families. Each plotted point represents the mean x and y coordinates for a bin of about 10 Dunedin Study members. The solid red line is the best-fitting regression line for the raw data. The dashed lines show the mean level of attainment for each SES subgroup. The distribution of polygenic scores within each subgroup is shown in the box-and-whiskers plots at the bottom of the figure. The vertical line in the center of each box marks the median, and the left and right edges of each box correspond to the 25th and 75th percentiles, respectively. The whiskers indicate 95% of the range. The black vertical lines behind the box plots show the cohort mean. The plots align with the scales on the x-axes of the graphs.
Dunedin Study data confirmed that children with higher polygenic scores had grown up in families with more socioeconomic resources (Krapohl & Plomin, 2016). But the data also showed that even for children born into socially disadvantaged circumstances, higher polygenic scores predicted upward social mobility.
Part 2: How did children with higher polygenic scores grow apart from their peers?
If children with higher polygenic scores do achieve higher levels of attainment in schooling and beyond, it is important to know how this comes about. The intermediate phenotypes that link DNA sequence with life outcomes can provide clues about genetic mechanisms and can also suggest targets for interventions designed to improve children’s outcomes (Belsky, Moffitt, & Caspi, 2013). The next analysis examined how children with higher polygenic scores grew apart from their peers beginning during the early school years and continuing through midlife.
Children with higher polygenic scores were more likely to say their first words at younger ages
When Dunedin Study members were 3 years old, their mothers were interviewed about the ages at which the members achieved each of a series of developmental milestones. The milestones, ordered by the normative age at which they were reached, were smiling, walking, talking, feeding oneself, daytime potty training, communicating using sentences, and nighttime potty training (see Fig. S5 in the Supplemental Material). Dunedin Study members with higher polygenic scores began talking earlier, on average, than peers with lower scores (hazard ratio = 1.12, 95% CI = [1.05, 1.19], p < .001), and were also somewhat quicker to begin communicating using sentences (hazard ratio = 1.06, 95% CI = [1.00, 1.13], p = 0.052), although this difference was not statistically significant at the α = 0.05 threshold. This accelerated development was restricted to verbal ability; Dunedin Study members with higher polygenic scores did not reach other developmental milestones ahead of peers.
Children with higher polygenic scores acquired reading skills at younger ages
Study members’ reading skill was assessed with the Burt Word Reading Test at each measurement session from ages 7 to 18 years. We used longitudinal multilevel growth models to test genetic associations with the model intercept and linear and quadratic slopes of change in reading over time (see Fig. S6 in the Supplemental Material). The model intercept captured the cohort mean reading score at age 7 (b = 30.50). The linear-slope term captured average annual change in reading score from age 7 to age 18 (b = 12.50). The quadratic-slope term captured deceleration of change; that is, it captured the convexity of the trajectory across childhood (b = −0.60). All model terms were statistically significant (p < .001). We tested genetic influence on growth by modeling intercept and slope terms of the growth curve as functions of the polygenic score and covariates. Polygenic score coefficients measured the effect of a 1-standard-deviation difference in polygenic score on reading at age 7 (intercept), on the linear change per year in reading score between the ages of 7 and 18 (linear slope), and on the deceleration of that change with increasing age (quadratic slope).
Growth-curve modeling found that by age 7, children with higher polygenic scores were already stronger readers (intercept: b = 2.79, SE = 0.57, p < .001). Thereafter, these children improved their performance at a faster rate (linear slope: b = 0.25, SE = 0.09, p = 0.005) and reached their peak performance at an earlier age (quadratic slope: b = −0.03, SE = 0.01, p < .001; Fig. 3). These results show that, on this educational fundamental, Dunedin Study members with higher polygenic scores were often already ahead of their peers by the second grade, and this gap in ability tended to expand through the middle-school years, although genetic differences were small.
Fig. 3.
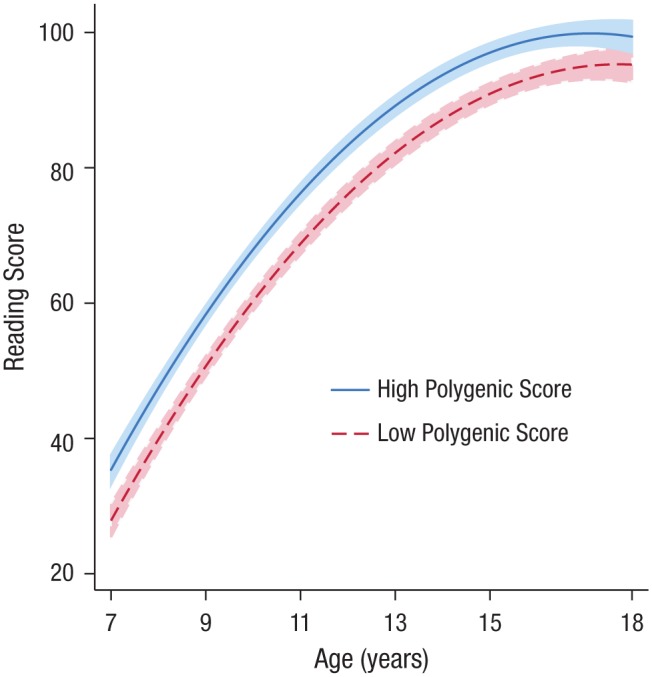
Children with higher polygenic scores acquired reading skills more rapidly. Association between age and reading skill (as measured by the Burt Word Reading Test; Scottish Council for Research in Education, 1976), separately for children with high polygenic scores (≥ 1 SD above the mean; n = 159) and those with low polygenic scores (≥ 1 SD below the mean; n = 147). The shaded areas show 95% confidence intervals.
Adolescents with higher polygenic scores had higher aspirations as high school students
When Dunedin Study members were 15 years old, they were asked about the highest level of education they planned to complete and also about the kind of job they hoped to have some day. At this critical developmental juncture, when adolescents of this New Zealand birth cohort (1972–1973) were choosing whether to remain in school or to begin working, adolescents in the Dunedin cohort who had higher polygenic scores aspired to higher educational attainments (r = 0.15, p < .001; for aspiration to a university degree, RR = 1.24, 95% CI = [1.11, 1.37]) and more prestigious occupations (r = 0.12, p = 0.001; for aspiration to a high-status “professional” occupation, such as a medical doctor or engineer, RR = 1.16, 95% CI = [1.06, 1.27]).
Adolescents with higher polygenic scores tested at higher levels in high school
Students distinguish themselves academically by selecting into more competitive tracks and by their performance within those tracks. At the time the Dunedin Study members were in high school, New Zealand pupils sat for standardized exams in the fifth, sixth, and seventh forms (ages 15–17 years). For the 1972–1973 birth cohort, the age-15 certificate exam was required to earn a School-Leaving Certificate (the minimum secondary education credential at the time); the age-16 Sixth-Form Certificate was used for entry to various tertiary institutions; and the age-17 bursary exam was the method through which the government allocated funds (bursaries) to support living costs during university. Dunedin Study members brought their official exam records to the research unit, and their scores were recorded. Adolescents with higher polygenic scores were less likely to have left school without testing for a credential (RR = 0.78, 95% CI = [0.66, 0.93], p = 0.006), and were more likely to advance to the next testing level at each age (ordered logit odds ratio = 1.32, 95% CI = [1.12, 1.55], p = .001. They also performed better on the tests (r = .24 for the age-15 certificate exam, p < .001; r = .19 for the age-16 sixth-form exam, p < .001; and r = .19 for the bursary exam, p = .032). These findings show that adolescents with higher polygenic scores distinguished themselves from peers by more often competing at advanced academic levels and by outperforming peers on standardized tests.
Dunedin Study members with higher polygenic scores were more likely to pursue occupational opportunities outside of New Zealand
Success in competitive professional environments sometimes depends on “going the extra mile.” The next analysis tested whether Dunedin Study members with higher polygenic scores did so literally, using data on where members lived and worked from the time they were 21 years old through the end of follow-up (obtained from life-history calendars completed by the Dunedin Study members at each adult assessment; see the Supplemental Material). Overseas work experience is common for New Zealanders, including Dunedin-cohort members. By age 38, more than a third of the Dunedin cohort (42%) had worked in a foreign country for a spell of at least 12 months. The most common destination for overseas work experience was Australia (about 41% of those who worked abroad did so in Australia but not elsewhere). Work experience in a foreign country beyond Australia has special significance in New Zealand and is known as “the Big OE” (for overseas experience; Wikipedia, 2014). Dunedin Study members with higher polygenic scores were more likely to have an OE (RR = 1.17, 95% CI = [1.05, 1.32], p = .007). Most New Zealanders who work abroad ultimately return home to raise their families. At the time of the age-38 interviews, 18% of Dunedin Study members lived and worked in Australia, and an additional 7% lived and worked in another foreign country. Dunedin Study members with higher polygenic scores were more likely to be among these migrants (RR = 1.18, 95% CI = [1.05, 1.32], p = .005); compared with the polygenic scores of those living in New Zealand, scores for migrants to Australia were higher by 0.19 SD, 95% CI = [0.02, 0.36], p = .026, and scores for migrants to other countries were higher by 0.27 SD, 95% CI = [0.02, 0.51], p = .032 (Fig. 4). These findings suggest that Dunedin Study members with higher polygenic scores distinguished themselves in the labor force by more often pursuing job opportunities beyond New Zealand.
Fig. 4.

Association between polygenic score and likelihood of migrating out of New Zealand. The value at the end of each arrow is the average standard-deviation difference in polygenic scores (with the standard error of the estimate in parentheses) between Dunedin Study members who moved to that area (North America, n = 14; Europe, n = 41; Asia and Africa, n = 13; Australia, n = 162) and members who remained in or returned to New Zealand. Migrants were defined as Dunedin Study members who had lived and worked abroad for a minimum of 12 months since the age of 21 and who were still living abroad at the age-38 assessment.
Dunedin Study members with higher polygenic scores were more financially planful
At ages 32 and 38, friends and relatives who knew each Dunedin Study member well reported about the member’s ability to manage money (96% response rate). In addition, Dunedin Study members were interviewed about financial building blocks (investments and retirement savings) and saving behaviors; scores on financial building blocks and savings behavior scales were averaged to calculate a financial planfulness score (see the Supplemental Material). Dunedin Study members with higher polygenic scores were rated by their informants as having fewer difficulties managing their money (r = −.08, p = .013) and were more financially planful on average (r = .09, p = .008). These findings show that in addition to acquiring academic credentials and professional experience to command higher earnings, Dunedin Study members with higher polygenic scores tended to be better managers of their financial resources.
Dunedin Study members with higher polygenic scores selected partners with higher socioeconomic attainments
In addition to education, wages, and investments, so-called marriage markets contribute to a person’s accumulation of social and financial resources (Breen & Salazar, 2011). According to prior research, men and women who are better-off tend to pair with one another, and this pattern of homophilous mating also occurs for people who are less well off (Schwartz, 2013). By midlife, most Dunedin Study members were in a serious relationship. Dunedin Study members with higher polygenic scores were no more likely to be in a serious relationship than members with lower scores (RR = 1.00, 95% CI = [0.98, 1.03], p = .776). Dunedin Study members in serious relationships were interviewed about their partners’ education and income. This partner information was available for 83% of the 918 Dunedin Study members for whom we had genetic data (n = 759). Information was used to classify partners’ SES as low (31%), middle (49%), or high (20%; see the Supplemental Material). Dunedin Study members with higher polygenic scores tended to have partners with higher SES (r = .09, p = .011; Fig. 5). These findings suggest that Dunedin Study members with higher polygenic scores bolstered the socioeconomic advantages they accrued through their own educational and occupational attainments by partnering with socially advantaged mates.
Fig. 5.
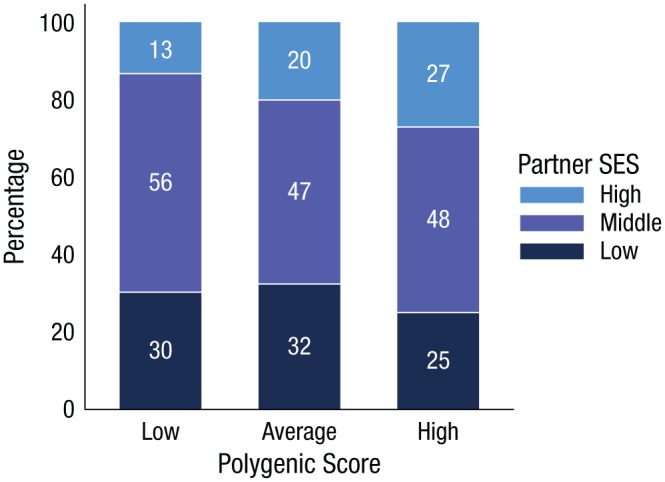
Association between polygenic score and partner’s socioeconomic status (SES). The graph shows the percentages (inside bars) of members who had low-, middle-, and high-SES partners, separately for Dunedin Study members with low polygenic scores (≥ 1 SD below the mean; n = 119), average polygenic scores (within 1 SD of the mean; n = 504), and high polygenic scores (≥ 1 SD above the mean; n = 136). Partners’ SES was defined according to whether they had completed a university degree and whether their income was above the national sex-specific median: High-SES partners had a university education and an above-median income, middle-SES partners met only one of these criteria, and low-SES partners met neither criterion.
Dunedin Study members with higher polygenic scores were not more satisfied with their lives
A higher polygenic score predicted conventional indicators of success: educational achievement, occupational prestige, financial security, even securing a socioeconomically successful partner. Yet some conceptualizations of success extend beyond the realms of material and social attainment. We therefore tested whether the polygenic score predicted Dunedin Study members’ self-rated satisfaction with life at age 38. It did not (r = .04, p = .189).
Genetic associations with pathways to socioeconomic success were not accounted for by Dunedin Study members’ social origins
Because of evidence that Dunedin Study children’s polygenic scores were associated with their families’ socioeconomic circumstances (r = .13, p < .001), Part 2 analyses presented in this section were repeated with statistical adjustment for the SES of Dunedin Study members’ families when the members were children. Genetic associations were largely independent of childhood SES. Complete results are included in Table S3 in the Supplemental Material.
Part 3: What personal characteristics helped children with higher polygenic scores achieve social and economic success?
The pattern of findings described previously suggests that the genetics uncovered in GWASs of educational attainment contribute to certain underlying characteristics that influence not only educational success, but also success in broader social and economic domains of life. We tested three different characteristics that might function as mediators of genetic influence on success in multiple life domains. These characteristics were higher cognitive ability, stronger noncognitive skills, and overall better physical health.
Children with higher polygenic scores performed better on IQ tests and exhibited a more rapid pace of cognitive development during childhood
Children with higher polygenic scores did not score significantly higher than their peers on the Peabody test at age 3 (r = .05, p = .133), but thereafter they showed an increasing cognitive advantage (r = 0.13 for Stanford-Binet IQ at age 5; r = .13–.19 for WISC-R IQ at ages 7–13; all ps < .001; Fig. 6a).
Fig. 6.

Association between polygenic score and cognitive ability. The plotted points in (a) show the magnitude of the effect of a 1-SD increase in polygenic score on standardized IQ (1 IQ point = 1/15 of 1 SD) measured at ages 3, 5, 7, 9, 11, and 13. Error bars indicate 95% confidence intervals. Cognitive ability was measured with the Peabody Picture Vocabulary Test (Dunn, 1965) at age 3, the Stanford-Binet Intelligence Scale (Terman & Merrill, 1960) at age 5, and Wechsler Intelligence Scales for Children–Revised (WISC-R; Wechsler, 1974) at ages 7–13. In (b), mental age is graphed as a function of chronological age for children with high polygenic scores (≥ 1 SD above the mean; n = 159) and those with low polygenic scores (≥ 1 SD below the mean; n = 147). The shaded areas show 95% confidence intervals. Mental age was measured with the WISC-R.
This pattern of findings indicates genetic influence over the developmental process through which children accumulate cognitive abilities, a hypothesis suggested by previous twin research on intelligence (Plomin, 2012) but, to our knowledge, still untested in molecular data. To test hypotheses about polygenic influence on the course of cognitive development, data from repeated WISC-R assessments were analyzed. This analysis focused on mental-age scores, rather than IQ scores, because, whereas IQ scores are age-corrected in order to allow comparisons between a child and the population of children of the same chronological age (e.g., a student’s score is in the 66th percentile for his or her age), mental-age scores express the child’s level of performance as the chronological age for which his or her score is normative (e.g., a 10-year-old student might have a mental age of 12). Mental age can be used to monitor a child’s intraindividual development over time (e.g., a 10-year-old child with an unstandardized IQ score equal to the average unstandardized score for 12-year-olds would have a mental age of 12; Lezak, Howieson, Loring, Hannay, & Fischer, 2004).
Growth-curve modeling tested whether the cognitive development of children with higher polygenic scores differed from that of their peers (see the Supplemental Material). The model intercept captured the cohort mean mental age at a chronological age of 7 years (b = 7). The linear-slope term captured average annual change in mental age (b = 1). Model terms were statistically significant (p < .001). We tested genetic influence on growth by modeling intercept and slope terms of the growth curve as functions of the polygenic score and covariates. Polygenic-score coefficients measure the effect of a 1-standard-deviation difference in polygenic score on mental age at chronological age 7 (intercept), and on the linear change per year in mental age from chronological age 7 to 13 (linear slope).
Children with higher polygenic scores tended to have older mental ages at the chronological age-7 baseline (intercept: b = 0.13, SE = 0.04, p < .001), and they exhibited a faster pace of cognitive development through age 13 years (slope: b = 0.05, SE = 0.01, p < .001; Fig. 6b). Taken together, these effects mean that a child with a genetic score 1 standard deviation above the mean would, by the age of 13 years, accrue a 6-month advantage in cognitive development relative to the population norm.
Children with higher polygenic scores had stronger noncognitive skills
In addition to cognitive abilities, so-called noncognitive skills influence individuals’ attainments (Heckman, 2006). Genetic associations were tested for two noncognitive skills, self-control and interpersonal skill.
As described previously (Moffitt et al., 2011), dossiers of children’s self-control skills were compiled from observational ratings and from parent and teacher reports when the children were between ages 3 and 11 years old, and from self-reports when the children were 11 years old. Children with higher polygenic scores tended to show better self-control skills across their first decade of life (r = .10, p = .001).
Children’s interpersonal skill was measured from reports by trained research staff on behavioral observations of the Dunedin Study members at ages 3, 5, 7, and 9 years. At each age, children were given binary ratings if they impressed the staff as being friendly, confident, cooperative, or communicative. These ratings were used to form an interpersonal skill scale (see the Supplemental Material). Children with higher polygenic scores were rated as having better interpersonal skill (r = .10, p = .004).
Genetic associations with children’s cognitive abilities and noncognitive skills were independent of their social origins
Analysis of childhood psychological characteristics was repeated with statistical adjustment for the SES of the children’s families. Genetic associations were found to be independent of childhood SES. Complete results are included in Table S3 in the Supplemental Material.
Cognitive abilities and noncognitive skills mediated genetic influences on educational and socioeconomic attainments
Genetic associations with cognitive and noncognitive skills suggest that these characteristics could explain why children with higher polygenic scores went on to achieve higher educational and socioeconomic attainments. Mediation analyses tested whether cognitive abilities and noncognitive skills accounted for genetic associations with life attainments (see Figs. S7 and S8 and Table S4 in the Supplemental Material). Cognitive ability, self-control, and interpersonal skill were all statistically significant mediators of genetic associations with educational and socioeconomic outcomes. Together, cognitive abilities and noncognitive skills accounted for about 60% of the genetic association with educational attainment and about 47% of the genetic association with the adult-attainment-factor score (p < .001 for both).
Children with higher polygenic scores were no healthier than their peers
Genetic associations with adult attainments might also result from general benefits to physical integrity that make individuals healthier as children, setting them up for success later in life (Case, Fertig, & Paxson, 2005). Dunedin Study members’ health was measured from repeated clinical assessments of motor development, growth and obesity, cardiovascular and pulmonary functioning, and infections and injuries when the children were between the ages of 3 and 11 years (see the Supplemental Material). Dunedin Study members with higher polygenic scores were no healthier in childhood than their peers (r = .01, p = .806). Together with the aforementioned lack of association between the polygenic score and walking, feeding, and potty training, this finding suggests that GWASs of educational attainment have not identified a set of genetic influences on overall robust functioning of the body’s physical systems.
As a second test of the physical-robustness hypothesis, we analyzed the genetics of human height. Like education, human height is known to be related to socioeconomic attainments (Case & Paxson, 2008). For this analysis, we substituted a polygenic score derived from GWASs of human height for the education polygenic score in our original analysis predicting life attainments. We used published results from a large-scale GWAS of human height (Wood et al., 2014) to calculate height polygenic scores for Dunedin Study members. As expected, Dunedin Study members’ polygenic scores for height were correlated with their measured stature (r = .54, p < .001). However, even though taller members did tend to do better in life (adult-attainment factor: r = .13, p = .011), we observed no association between the polygenic score for height and life attainments measured by the adult-attainment factor (r = .00, p = .952).
Discussion
This article describes how genetic discoveries made in GWAS analysis of educational attainment were related to the courses of human lives. We studied a population-representative birth cohort followed over the course of four decades. Findings showed that genome-wide DNA-sequence differences identified from GWASs and summarized in a “polygenic score” were associated with basic processes of human social and economic success. Three points are important in interpreting the substance of these findings. First, genetic associations between the polygenic score and adult socioeconomic success were not fully accounted for by educational attainment. Second, children’s socioeconomic origins were correlated with their polygenic scores; however, genetic associations with adult socioeconomic success, with the developmental and behavioral pathways to such success, and with the psychological characteristics we studied were mostly independent of children’s socioeconomic origins. Third, across the board, effect sizes were small in magnitude.
The primary finding was that polygenic scores derived from a GWAS of educational attainment predicted life outcomes well beyond schooling. Dunedin Study members with higher polygenic scores were geographically mobile in search of professional opportunities, they built more successful careers, they secured higher social status mates, and they built stronger financial foundations for retirement. From childhood to midlife, Dunedin Study members’ genetic inheritance predicted their social mobility. Even among children born into socially disadvantaged homes, those with higher polygenic scores achieved more. Achievements of children with higher polygenic scores were enabled in part by a suite of psychological traits already evident from early life. Dunedin Study members with higher polygenic scores talked earlier, did better on cognitive tests beginning at 5 years old, and showed a more rapid pace of cognitive development, and they developed better self-control and interpersonal skills. Collectively, these childhood psychological characteristics accounted for about half of the genetic association with social success in adulthood. Strikingly, the same genetic differences that predicted children’s cognitive, emotional, and social functioning were not related to their attainment of nonverbal milestones or their physical health.
The substance of these findings is bolstered by evidence that GWAS discoveries for educational attainment are not genetic artifacts of a socially privileged class. Because children born into better-off families are more likely to earn advanced degrees (Breen & Jonsson, 2005), a GWAS of educational attainment could have identified the genetics of better-off families rather than the genetics of a propensity to succeed. GWAS discoveries could be no more than markers of socially advantaged ancestry. Consistent with such a possibility, both previous studies (Conley et al., 2015; Domingue et al., 2015; Krapohl & Plomin, 2016) and the current study found that children born into better-off homes had higher polygenic scores. But two findings suggest that the genetic associations are nonspurious. First, studies that compare siblings within the same family (who share identical ancestries) find that the sibling with the higher polygenic score tends to complete more years of schooling (Domingue et al., 2015; Rietveld, Conley, et al., 2014). Second, our study shows that polygenic scores also influence changes in social position within a single generation, thereby suggesting a mechanism to explain the gene-environment correlation in which children of socially advantaged families tend to have higher polygenic scores.
We acknowledge limitations. First, our study concerned a single birth cohort of European descent in one country, New Zealand. The extent to which findings generalize to other birth cohorts growing up under other circumstances needs to be tested. Although New Zealand has levels of social inequality similar to those in the United States and Great Britain (after-tax Gini coefficients: New Zealand, .33; United Kingdom, .34; United States, .37; Wikipedia, 2015), international comparisons will prove informative (Tucker-Drob & Bates, 2016), including in settings in which inequality is engineered to be low (Firkowska et al., 1978). Second, the measurement of the human genome we studied is necessarily preliminary. We studied a polygenic score based on the best available information about genetic correlates of educational success. But future GWASs with larger sample sizes are expected to yield a more precise set of genetic correlates. Replication checks with subsequent iterations of the polygenic score for education are needed. Third, follow-up of social and economic outcomes in our study is right censored, extending through the fourth decade of life, but not beyond. Extension of findings into longitudinal cohort studies of older adults is needed to clarify the extent of genetic associations into the second half of the life course. Finally, the set of outcomes, pathways, and traits that we studied is not comprehensive. Studies of other samples that use different measurement batteries are needed to expand our understanding of how genetic correlates of educational attainment relate to human life courses.
In light of these limitations, our study contributes to public and scientific conversation about genetic discoveries regarding educational attainment in five ways. First, GWAS discoveries regarding educational attainment are not only about education. They are discoveries about socioeconomic success more broadly (although perhaps not about satisfaction with life). Education accounted for under half of the relationship between genes and adult socioeconomic attainments, suggesting that the mechanisms of genetic influence are not limited to success in schooling and do not depend on it.
Second, the psychological mediators of genetic associations with socioeconomic success involve more than what IQ tests measure as intelligence. Multivariate twin research suggests that the heritability of educational attainment reflects genetic influences on noncognitive skills as well as intelligence (Krapohl et al., 2014). We found molecular evidence to support this hypothesis: Children’s polygenic scores for educational attainment were correlated with their noncognitive self-control and interpersonal skills as well as with their IQ scores. Our “top-down” approach, working from an adult phenotype backward in development toward a DNA sequence, yielded findings that suggest behavioral mechanisms for genetic influences on educational attainment.
Third, children with higher polygenic scores grew apart from their peers along coherent developmental trajectories that began to form even before they entered school. Dunedin Study members with higher polygenic scores began to talk at a younger age. Subsequently, they learned to read before many of their peers did. This early success was followed by loftier academic aspirations and attainments extending into adulthood. These findings support the logic of interventions to promote early literacy, particularly those focusing on early language development (Talbot, 2015).
In addition, and more speculatively, the life-course analysis that we report also suggests that GWAS findings for educational attainment may provide a clue to the genetic roots of life-history differences in free-living humans. Unlike education, which is a relatively modern human experience, patterns of migration, mate selection, and resource acquisition and management are ancient human behaviors that plausibly bear the imprint of our species’ evolutionary history. The finding that GWAS discoveries for education predict these ancient behaviors suggests a window into genetic regulation of humans’ strategies to survive and reproduce. Our data cannot reveal whether frequencies of education-associated genotypes reflect some Darwinian fitness strategy. Rather, the data suggest that individuals whose genomes carry more education-associated alleles are forging life histories that achieve success in the modern world, and the pathways to this success include some that would be familiar to our ancestors.
Fourth, the current findings lend weight to earlier twin-study observations that genes shape not just behavior, but also the environment that contextualizes and constrains behavioral choices (Plomin & Bergeman, 1991). The molecular realization of such gene-environment correlations creates opportunities for social theory and research. Results reported in this study suggest that by incorporating DNA sequence into studies of status attainment, migration, assortative mating, and financial behavior, social scientists may be able to frame novel “sociogenomic” research questions. For example, do public programs to build human capital (such as improving teacher salaries or providing universal access to prekindergarten education) change the ways in which genes influence life attainments? If so, are the returns greater for programs that magnify genetic influences or for programs that reduce them? Do the genetics of educational attainment relate to social gradients in midlife health and aging? If so, how is this process shaped by health-care costs, quality, and access? As concerns about economic inequality increase, are genes linked with socioeconomic success becoming concentrated within social and geospatial elites? If so, is this process influenced by exogenous shocks such as natural disasters, policy shifts such as multinational trade and border agreements, or cultural changes in equality of opportunity?
Finally, our findings shed light on the stakes of the public conversation about sociogenomic discoveries that is now emerging. For the general public, the significance of new knowledge about how to measure and interpret DNA sequence is uncertain and hotly debated, even in the field of biomedicine, in which clinical applications of genetic discoveries are already possible (Khoury & Evans, 2015; Lander, 2015; Roberts et al., 2012). At present, genetic prediction of educational outcomes and life success in general is far from sensitive or specific enough to recommend any translational application. Although there is movement toward improving the predictive power of polygenic scores through increased GWAS sample sizes and improved genomic measurements, a precision medicine-type approach to human capital development remains well out of reach. And yet debate is already under way about the possibility for genetic testing to someday be used in forecasting human potential. Policy action may be needed to regulate the ethical use of genomic information in school admissions and tracking decisions, and such actions should be informed by realistic estimates of the magnitude of genetic effects.
Supplementary Material
Acknowledgments
We thank the Dunedin Study members, along with their parents, teachers, partners, and peer informants, and study founder Phil Silva.
Footnotes
Action Editor: Ian H. Gotlib served as action editor for this article.
Declaration of Conflicting Interests: The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding: The Dunedin Multidisciplinary Health and Development Research Unit is supported by the New Zealand Health Research Council and the New Zealand Ministry of Business, Innovation and Employment (MBIE). This research was supported by National Institute on Aging Grants R01-AG032282, R01-AG048895, and 1R01-AG049789, United Kingdom Medical Research Council Grant MR/K00381X, and United Kingdom Economic and Social Research Council Grant ES/M010309/1. Additional support was provided by National Institute on Aging Grant P30-AG028716, by Eunice Kennedy Shriver National Institute of Child Health and Human Development Grant R21-HD078031, and by the Jacobs Foundation. D. W. Belsky is supported by an Early-Career Research Fellowship from the Jacobs Foundation.
Supplemental Material: Additional supporting information can be found at http://pss.sagepub.com/content/by/supplemental-data
Open Practices: The analysis plan for this study can be found at https://docs.google.com/viewer?a=v&pid=sites&srcid=ZGVmYXVsdGRvbWFpbnxkdW5lZGluZXJpc2tjb25jZXB0cGFwZXJzfGd4Ojc5NTA0YjNhNjMzYzY3YmE. The data are not publicly available because (a) the size of the sample and the nature of the data might make it possible to identify participants and (b) consent for broad sharing of the data was not obtained from the participants. However, the Supplemental Material provides details about possible data sharing. The complete Open Practices Disclosure for this article can be found at http://pss.sagepub.com/content/by/supplemental-data.
References
- 1000 Genomes Project. (2016). 1000 Genomes project data. Retrieved from http://www.1000genomes.org/
- Baron R. M., Kenny D. A. (1986). The moderator–mediator variable distinction in social psychological research: Conceptual, strategic, and statistical considerations. Journal of Personality and Social Psychology, 51, 1173–1182. doi: 10.1037/0022-3514.51.6.1173 [DOI] [PubMed] [Google Scholar]
- Belsky D. W., Moffitt T. E., Caspi A. (2013). Genetics in population health science: Strategies and opportunities. American Journal of Public Health, 103(Suppl. 1), S73–S83. doi: 10.2105/AJPH.2012.301139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breen R., Jonsson J. O. (2005). Inequality of opportunity in comparative perspective: Recent research on educational attainment and social mobility. Annual Review of Sociology, 31, 223–243. doi: 10.1146/annurev.soc.31.041304.122232 [DOI] [Google Scholar]
- Breen R., Salazar L. (2011). Educational assortative mating and earnings inequality in the United States. American Journal of Sociology, 117, 808–843. doi: 10.1086/661778 [DOI] [Google Scholar]
- Case A., Fertig A., Paxson C. (2005). The lasting impact of childhood health and circumstance. Journal of Health Economics, 24, 365–389. doi: 10.1016/j.jhealeco.2004.09.008 [DOI] [PubMed] [Google Scholar]
- Case A., Paxson C. (2008). Stature and status: Height, ability, and labor market outcomes. Journal of Political Economy, 116, 499–532. doi: 10.1086/589524 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chabris C. F., Lee J. J., Cesarini D., Benjamin D. J., Laibson D. I. (2015). The fourth law of behavior genetics. Current Directions in Psychological Science, 24, 304–312. doi: 10.1177/0963721415580430 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conley D., Domingue B., Cesarini D., Dawes C. T., Rietveld C. A., Boardman J. (2015). Is the effect of parental education on offspring biased or moderated by genotype? Sociological Science, 2, 82–105. doi: 10.15195/v2.a6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Zeeuw E. L., van Beijsterveldt C. E. M., Glasner T. J., Bartels M., Ehli E. A., Davies G. E., . . . Boomsma D. I. (2014). Polygenic scores associated with educational attainment in adults predict educational achievement and ADHD symptoms in children. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 165B, 510–520. doi: 10.1002/ajmg.b.32254 [DOI] [PubMed] [Google Scholar]
- Domingue B. W., Belsky D. W., Conley D., Harris K. M., Boardman J. D. (2015). Polygenic influence on educational attainment. AERA Open, 1(3), 1–13. doi: 10.1177/2332858415599972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudbridge F. (2013). Power and predictive accuracy of polygenic risk scores. PLoS Genetics, 9(3), Article e1003348. doi: 10.1371/journal.pgen.1003348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunn L. (1965). The Peabody Picture Vocabulary Test. Minneapolis, MN: American Guidance Service. [Google Scholar]
- Euesden J., Lewis C. M., O’Reilly P. F. (2015). PRSice: Polygenic Risk Score software. Bioinformatics, 31, 1466–1468. doi: 10.1093/bioinformatics/btu848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firkowska A. N., Ostrowska A., Sokolowska M., Stein Z., Susser M., Wald I. (1978). Cognitive development and social policy. Science, 200, 1357–1362. doi: 10.1126/science.663616 [DOI] [PubMed] [Google Scholar]
- Heckman J. J. (2006). Skill formation and the economics of investing in disadvantaged children. Science, 312, 1900–1902. doi: 10.1126/science.1128898 [DOI] [PubMed] [Google Scholar]
- Henig R. M. (2015, December 11). Are there genes for intelligence—And is it racist to ask? Retrieved from http://news.nationalgeographic.com/2015/12/151211-genetics-intelligence-racism-science/
- Howie B. N., Donnelly P., Marchini J. (2009). A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genetics 5(6), Article e1000529. doi: 10.1371/journal.pgen.1000529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoury M. J., Evans J. P. (2015). A public health perspective on a national precision medicine cohort: Balancing long-term knowledge generation with early health benefit. The Journal of the American Medical Association, 313, 2117–2118. doi: 10.1001/jama.2015.3382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krapohl E., Plomin R. (2016). Genetic link between family socioeconomic status and children’s educational achievement estimated from genome-wide SNPs. Molecular Psychiatry, 21, 437–443. doi: 10.1038/mp.2015.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krapohl E., Rimfeld K., Shakeshaft N. G., Trzaskowski M., McMillan A., Pingault J.-B., . . . Plomin R. (2014). The high heritability of educational achievement reflects many genetically influenced traits, not just intelligence. Proceedings of the National Academy of Sciences, USA, 111, 15273–15278. doi: 10.1073/pnas.1408777111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lander E. S. (2015). Cutting the Gordian helix—regulating genomic testing in the era of precision medicine. The New England Journal of Medicine, 372, 1185–1186. doi: 10.1056/NEJMp1501964 [DOI] [PubMed] [Google Scholar]
- Lezak D. M., Howieson D. B., Loring D. W., Hannay H. J., Fischer J. S. (2004). Neuropsychological assessment (4th ed.). New York, NY: Oxford University Press. [Google Scholar]
- Moffitt T. E., Arseneault L., Belsky D., Dickson N., Hancox R. J., Harrington H., . . . Caspi A. (2011). A gradient of childhood self-control predicts health, wealth, and public safety. Proceedings of the National Academy of Sciences, USA, 108, 2693–2698. doi: 10.1073/pnas.1010076108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavot W., Diener E. (1993). Review of the Satisfaction With Life Scale. Psychological Assessment, 5, 164–172. [Google Scholar]
- Plomin R. (2012). Genetics: How intelligence changes with age. Nature, 482, 165–166. doi: 10.1038/482165a [DOI] [PubMed] [Google Scholar]
- Plomin R., Bergeman C. S. (1991). The nature of nurture: Genetic influence on “environmental” measures. Behavioral & Brain Sciences, 14, 414–427. doi: 10.1017/S0140525X00070588 [DOI] [Google Scholar]
- Poulton R., Moffitt T. E., Silva P. A. (2015). The Dunedin Multidisciplinary Health and Development Study: Overview of the first 40 years, with an eye to the future. Social Psychiatry & Psychiatric Epidemiology, 50, 679–693. doi: 10.1007/s00127-015-1048-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preacher K. J., Hayes A. F. (2008). Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behavior Research Methods, 40, 879–891. doi: 10.3758/BRM.40.3.879 [DOI] [PubMed] [Google Scholar]
- Preacher K. J., Kelley K. (2011). Effect size measures for mediation models: Quantitative strategies for communicating indirect effects. Psychological Methods, 16, 93–115. doi: 10.1037/A0022658 [DOI] [PubMed] [Google Scholar]
- Price A. L., Patterson N. J., Plenge R. M., Weinblatt M. E., Shadick N. A., Reich D. (2006). Principal components analysis corrects for stratification in genome-wide association studies. Nature Genetics, 38, 904–909. doi: 10.1038/ng1847 [DOI] [PubMed] [Google Scholar]
- Price A. L., Zaitlen N. A., Reich D., Patterson N. (2010). New approaches to population stratification in genome-wide association studies. Nature Reviews Genetics, 11, 459–463. doi: 10.1038/nrg2813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rietveld C. A., Conley D., Eriksson N., Esko T., Medland S. E., Vinkhuyzen A. A. E., . . . Social Science Genetics Association Consortium. (2014). Replicability and robustness of genome-wide-association studies for behavioral traits. Psychological Science, 25, 1975–1986. doi: 10.1177/0956797614545132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rietveld C. A., Esko T., Davies G., Pers T. H., Turley P., Benyamin B., . . . Koellinger P. D. (2014). Common genetic variants associated with cognitive performance identified using the proxy-phenotype method. Proceedings of the National Academy of Sciences, USA, 111, 13790–13794. doi: 10.1073/pnas.1404623111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rietveld C. A., Medland S. E., Derringer J., Yang J., Esko T., Martin N. W., . . . Koellinger P. D. (2013). GWAS of 126,559 individuals identifies genetic variants associated with educational attainment. Science, 340, 1467–1471. doi: 10.1126/science.1235488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts N. J., Vogelstein J. T., Parmigiani G., Kinzler K. W., Vogelstein B., Velculescu V. E. (2012). The predictive capacity of personal genome sequencing. Science Translational Medicine, 4(133), Article 133ra58. doi: 10.1126/scitranslmed.3003380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz C. R. (2013). Trends and variation in assortative mating: Causes and consequences. Annual Review of Sociology, 39, 451–470. doi: 10.1146/annurev-soc-071312-145544 [DOI] [Google Scholar]
- Scottish Council for Research in Education. (1976). Burt Word Reading Test (rev. ed.). London, England: Hodder & Stoughton. [Google Scholar]
- Sherry S. S., Ward M. M., Kholodov M., Baker J., Phan L., Smigielski E. E., Sirotkin K. (2001). dbSNP: The NCBI database of genetic variation. Nucleic Acids Research, 29, 308–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer J. D., Willett J. B. (2003). Applied longitudinal data analysis. New York, NY: Oxford University Press. [Google Scholar]
- Talbot M. (2015, January 12). The talking cure. The New Yorker. Retrieved from http://www.newyorker.com/magazine/2015/01/12/talking-cure
- Terman L. M., Merrill M. A. (1960). Stanford-Binet Intelligence Scale: Manual for the third revision. Oxford, England: Houghton Mifflin. [Google Scholar]
- Tucker-Drob E. M., Bates T. C. (2016). Large cross-national differences in gene × socioeconomic status interaction on intelligence. Psychological Science, 27, 138–149. doi: 10.1177/0956797615612727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward M. E., McMahon G., St Pourcain B., Evans D. M., Rietveld C. A., Benjamin D. J., . . . Timpson N. J. (2014). Genetic variation associated with differential educational attainment in adults has anticipated associations with school performance in children. PLoS ONE, 9(7), Article e100248. doi: 10.1371/journal.pone.0100248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wechsler D. (1974). Manual for the Wechsler Intelligence Scale for Children–Revised. New York, NY: Psychological Corporation. [Google Scholar]
- Wikipedia. (2014). Overseas experience. Retrieved from http://en.wikipedia.org/w/index.php?title=Overseas_experience&oldid=633298303
- Wikipedia. (2015). List of countries by income equality. Retrieved from https://en.wikipedia.org/w/index.php?title=List_of_countries_by_income_equality&oldid=686250962
- Wood A. R., Esko T., Yang J., Vedantam S., Pers T. H., Gustafsson S., . . . Frayling T. M. (2014). Defining the role of common variation in the genomic and biological architecture of adult human height. Nature Genetics, 46, 1173–1186. doi: 10.1038/ng.3097 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.