

Graphical abstract
Introduction
Engineered nanoparticle platforms have been developed intensely in recent years, yielding significantly broadened applications from interrogating novel biology to new therapies. The mammalian immune system has emerged as perhaps the most powerful physiological system where nanoparticle design parameters have drastic implications for nanoparticle fate and function. In particular, the innate immune system is a major concern due to its role as the first-line defense against foreign invaders, responsible for initiating and maintaining the innate immune response as well as priming the adaptive immune response. Understanding how nanoparticles are perceived from a biological perspective is crucial to informing their design with different immunogenic, immunosuppressive, or stealth properties. Nanoparticles are particularly susceptible to innate immune responses such as opsonization and recognition by receptors that enable phagocytic uptake or evoke strong inflammatory responses due to the recognition of various “danger” signals. In this review, we will summarize some of the key advances in our understanding of how physical and biochemical parameters of nanoparticles interact with innate immunity, and note potential opportunities and emerging frontiers for controlling and evaluating nanoparticle-immune interactions.
Innate Immunity: The First Line of Host Response
The innate immune system comprises diverse cells and mechanisms to enable defense against pathogens, as well as maintenance of homeostasis by functions including clearance of cell debris, rapid recall of adaptive responses, and facilitating wound healing. The process of opsonization coats microbes or other particles with antibodies, circulating serum proteins, and/or complement proteins; one major function of opsonization is to clear pathogens from the body.
One of the most basic parts of the innate immune system that is at once incredibly simplistic and highly complicated and organized is the complement system. The complement system is often one of the first responders to pathogens. Complement involves a unique set of proteins that transform through a cascade of reactions from inactive to active forms, ultimately leading to clearance of pathogens. There are three pathways to the complement system: the classical, lectin, and alternative pathways. In the classical pathway, C1q, one of the three subunits of the first component of complement, binds to antibody-antigen complexes and initiates the cascade. In the lectin pathway, mannose binding lectin binds to sugars on pathogens. In the alternative pathway, activation can be spontaneous due to adsorption of complement on pathogen or materials surfaces. Not surprisingly, materials introduced into the body tend to activate the complement system by the alternative pathway due to adsorption and binding to surfaces. The complement cascade involves activation, amplification, progression, and regulation. An excellent review of the complement system is in Zipfle and Skerka 1. Complement activation can happen quite rapidly. Following injury, changes in complement factor have been documented to occur over hours and can continue for days 2, 3. Very few papers look at changes in critical players in the complement cascade over time periods of less than an hour, but for infusible nanomedicines, this time period may be critical.
To facilitate clearance, opsonization enhances the activity of innate immune cells including phagocytosis (uptake) and activation of effector functions. Innate immune cells include a diverse set of circulating and tissue-resident cells that include granulocytes (neutrophils, basophils, eosinophils, mast cells) and antigen-presenting cells (APCs) such as monocytes, macrophages, and dendritic cells. Uptake duties are performed by both neutrophils and APCs; particles that are phagocytosed are degraded and interrogated by the ingesting cell. Interrogation is achieved by a diverse array of receptors that can directly facilitate pathogen recognition, uptake, and generate an immune response. These receptors and mechanisms include pattern classical pattern recognition receptors (such as Toll-like receptors (TLRs) and C-type lectin receptors (CLRs)) as well as emerging pathways such as inflammasomes and autophagy, and the different receptors can be expressed on either cell membranes (TLRs, CLRs) or in the cytosol (NOD-like receptors, NLRs; RIG-I-like receptors (RLRs) (reviewed extensively in 4). Each family member binds specific molecular patterns often referred to as danger signals, consisting of molecules with structures and/or subcellular localization that are different from the host and often conserved among foreign microbes. Examples include TLR4 recognition of extracellular bacterial lipopolysaccharide, NLR/inflammasome recognition of cytosolic bacterial DNA or peptidoglycan, and RLR recognition of cytosolic viral RNA. A major endpoint for these pathways is the activation of effector functions, including the secretion of inflammatory cytokines, chemokines, histamine, and various other molecules as well as the activation of cellular processes such as apoptosis. It is notable that all of these receptors typically recognize microbes or other particulates in the low micrometer to sub-micron size range, so it is unsurprising that engineered nanoparticles are subject to surveillance and interrogation by these pathways. It is becoming increasingly clear that the host response to nanoparticles, particularly host tolerance of nanoparticle therapies and therapeutic efficacy, are dependent upon their interactions with the innate immune system.
Clinically Relevant Innate Immune Mechanisms for Nanomedicines
One of the most well-known nanomedicines is DOXIL for cancer therapy. DOXIL is a liposomal formulation of doxorubicin. It must be infused at a very slow rate compared to other chemotherapy agents to avoid a complement-mediated response; DOXIL is infused at a rate of 1 mg/ml over the course of hours compared to a bolus over 3-10 minutes for doxorubicin. Even with this slow infusion rate, approximately 10% of patients have a response that leads to serious cardiac side effects during the first administrations that can be dose limiting 5. It should be noted that with repeated administrations, the patient response is lessened.
DOXIL is far from the only nanomedicine with a complement-associated infusion reaction. Some iron oxide formulations for MRI contrast have been withdrawn from the clinic for complement activation issues 6. Furthermore, complement-associated reactions are seen in a small number of patients who get intravenous nanoparticle iron infusions for severe anemia.
A diverse range of nanomaterials from intravenously infused poly(lactic-co-glycolic acid) nanoparticles 7 to carbon nanotubes 8 have been associated with activation of the complement cascade either by the alternative pathway or a combination of the alternative pathway and direct absorption of C1q on the particles 8. Many of the newer nanomedicines currently in clinical trials have the potential to trigger the complement cascade complicating administration.
In a similar vein, nanoemulsions are oil-in-water systems stabilized by surfactants and are currently the only approved adjuvants for use in human vaccines. Their mechanisms of action remained undetermined for decades, but recent studies have begun to identify key components of that trigger innate immunity. In particular, inflammasomes appear to be critically important. Inflammasomes are a relatively new arm of innate immunity that are responsible for driving acute inflammation during infection, and can cause autoimmune disorders if unchecked. Notably, crystals such as silica, aluminum salt, or monosodium urate (cause of gout), are known to activate inflammasome cascades, increasing their immunogenicity via inflammatory cytokines as well as prostaglandin E2 (antibody-promoting) from macrophages 9-13. The over-arching mechanism has been suggested to be lysosomal disruption, such that any internalized material causing this effect (even in a “sterile” context) would be sufficient to activate the NLRP3 inflammasome 11. Consistent with this, another study indicated that small silver nanoparticles (30-1000 nm diameter) effectively induced lysosomal destabilization and inflammasome activation in macrophages 14. Macrophage NLRP3 is also activated by titania and silica nanoparticles 15. Intriguingly, this study also demonstrated that nanoparticle-mediated activation of inflammasomes may be independent of uptake, a paradigm that is distinct from the hypothesis of lysosomal disruption and seems to circumvent the dogma indicating uptake is required for immune recognition and signaling of particulates. Regardless, it is abundantly clear that nanomedicines have significant clinical implications that depend upon interactions with innate immunity.
Complement and Complications
The big question is what are the implications for activating the complement cascade with intravenously delivered nanomedicines. Administration of nanomedicines that activate the complement system, cleaving C3 to C3a, can trigger degranulation of cells including endothelial and mast cells leading to allergic-like responses. This can cause a host of responses termed complement-activation related pseudoallergy (CARPA) 16-18. CARPA is a hypersensitivity reaction that can be life threatening. The symptoms of CARPA include: increase in heart rate, hypotension, flushing of the skin (erythema), decreased cardiac output, decreased pulmonary pressures, and decreased blood gas levels 16-18. Not every person will have a life-threatening response, but based on the DOXIL data, approximately 10% of patients will have a serious enough response to require halting administration 5. Genetic differences are known to contribute to the complement response to nanomedicines, but it is not currently possible to predict a patient's response to an intravenous infusion of a nanomaterial 19.
Cytokine storms are another major concern in nanoparticle toxicity, and may be driven both by endotoxin contamination of nanoparticle preparations 20 as well as uncontrolled activation of various cells of the innate immune system by the biomaterial itself. Cytokine storms, similar to CARPA, are life-threatening, acute events that can occur during organ transplant, infection or infusion of therapeutics 21, 22. Symptoms of cytokine storm are shared with CARPA such as edema, erythema, fever, and hypotension. Additionally, organ and local tissue pathological damage occurs due to altered perfusion, intense local inflammation or fibrosis. Multiple organ failure can be the result of high systemic levels of cytokines and recruitment of immune cells, and organ dysfunction can persist in non-fatal cases 22. An increasing array of nanomedicines are entering clinical trials and even receiving FDA approval (e.g., the recent breakthrough approval of MM-398, a nanoliposomal formulation of irinotecan), so the clinical impact of immune responses to on-market nanomaterials (partially summarized in Table 1) presents an important issue for preclinical and clinical investigation.
Table 1. Clinically used nanomaterials that activate the innate immune system.
| Material | Activation Pathway | References |
|---|---|---|
| DOXIL | Alternative pathway activation shown to be activated | 5, 16, 23, 24 |
| Iron oxide nanoparticles for MRI contrast (Feridex) | Alternative pathway activated. C3b covalently attaches to hydroxyls and amines on the surfaces of the particles 25 | 6 |
| PEGylated PLA-based nanoparticles (BIND-014) | Unknown, but hypersensitivity reactions reported in phase II trial (18 of 40 patients with 3 experiencing grade 3 or 4 reaction) | 26 |
| Nanoemulsions (alum, MF59, AS04) | Inflammasomes, TLR4 | 11, 14, 15 |
Mangement of An Uncontrolled Immune Response
Immune hyperactivation and toxic responses such as cytokine storm and CARPA can be modulated by a number of approaches. Drugs that reduce the hyperactive immune response or address toxic symptoms include immunosuppresants (dexamethasone, ibuprofen, acetaminophen) or antihistamines 27. The infusion rate is critical to the CARPA response, and as discussed with DOXIL, slower infusions can reduce the response significantly. The CARPA response is also reduced with repeated administrations 24. To that end, if infusion of a nanomedicine is anticipated, an empty carrier can be infused to help ameliorate the hypersensitivity reaction before the delivery of the nanomedicine 27. This approach may alleviate CARPA, and potentially other innate immune pathways by inducing tolerance or exhaustion (such as endotoxin tolerance 28).
Even with all of these measures, some patients still have significant reactions that limit treatment. Furthermore, for some therapies, such as intravenously administered hemostatic agents 29-34, the methods to limit CARPA responses would not be possible. If one administers a hemostatic agent too slowly in response to an injury, bleed out may occur before the nanomaterial is present in sufficient quantities to be effective.
Critical Features of Biomaterials for Innate Immune Recognition
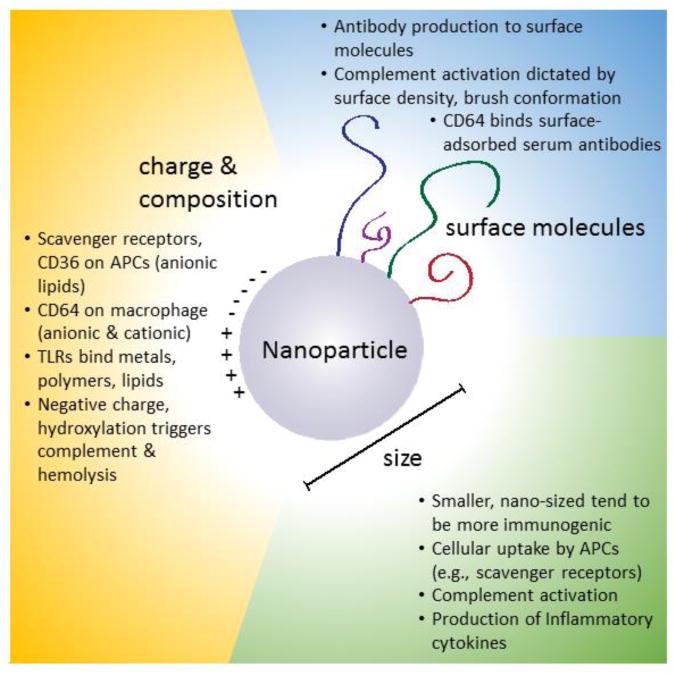
If one cannot administer a nanomedicine in a way avoids the hypersensitivity reaction of CARPA, can one engineer the nanomaterials to reduce complement activation? Potentially, yes. The molecular features on nanomedicines matter: surface charge, corona molecules, size, and shape of the nanomaterials have all been shown to matter for complement activation 35-37.
While groups have begun to investigate features of nanoparticles that impact immune activation 36, there has been no systematic study that provides clear guidelines to direct the design of the next generation of nanomedicines that are immune neutral. Nonetheless, there is a wealth of data that provides strong evidence for the materials features that matter. One of the critical things to consider when looking at the literature is the sometimes stark differences between in vitro and in vivo findings. When it comes to the properties of nanomaterials, what we design, no matter how elegant, is not what we achieve in vitro or in vivo at biological steady-state because all materials interact with biology. We design surfaces that can modulate those interactions, but those interactions ultimately direct the biological responses, whether they be absorption to hydrophobic regions, charged regions, regions of texture, or regions of molecular motifs. Furthermore, it is critical to remember that the interactions in vitro and in vivo are not always the same 38. Despite caveats including unpredictable adsorption of a range of biological molecules, it is still possible to direct and tailor biological interactions in a reproducible and predictable manner 38.
Size
Size matters in a multitude of ways. As the particle size increases, the surface area per mass decreases. Particles greater in size than a few microns show significantly reduced complement activation 39. Size is often considered at the tissue physiology level when studying trafficking and systemic distribution. However, size also has a significant impact on direct cellular interactions of biomaterials with the immune system. The role of size is vastly different at the macroscale (>100 μm) and microscale (>1 μm) compared to the nanoscale (1-1000 nm), with the foreign body response dominating objects larger than the nanoscale. A notable recent study demonstrated that extremely large spherical implants (>1.5 mm diameter) can reduce foreign body responses 40. For nanoparticles, processes such as opsonization and uptake have a greater impact on biomaterial-immune outcomes.
Particle size has a significant impact on the ability for uptake of nanoparticles by many cells, particularly those of the innate immune system. APCs are a crucial component of immune surveillance and determination of foreign versus self identity. They also play a major role in the elimination of circulating foreign bodies and interpreting whether to initiate an immune response. The uptake of nanoparticles by APCs such as macrophages and dendritic cells (DCs) is of interest when addressing issues of circulation life and nanoparticle immunotoxicity.
Poly(lactic-co-glycolic acid) nanoparticles were found to be significantly more immunogenic than microparticles, delivering their cargo for antigen processing and presentation to DCs both in vitro and in vivo 41. Titania nanoparticles (∼10 and 20 nm) produced inflammatory cytokines and activation of DCs in vitro when treating a human immune tissue construct; by comparison, microscale particles elicited minimal or no responses 42. Gold and silver nanoparticles of varying sizes were also tested with macrophages for uptake, cytotoxicity, and induction of inflammatory cytokines IL-1, IL-6, and TNF-α 43. Small gold nanoparticles (2-4 nm) elicited vastly increased levels (all 3 cytokines) and prolonged kinetics (IL-1, IL-6) of inflammatory gene expression when compared to medium (5-7 nm) and large (20-40 nm) nanoparticles. Gold nanoparticles had significant effects on macrophages compared to silver nanoparticles of similar size, potentially due to increased uptake/retention of gold nanoparticles. Toxicity was found to be size-dependent, with small gold nanoparticles more toxic than larger ones. Importantly, cytotoxicity can shape of immune responses via triggering distinct types of cell death that can be more or less immunogenic 44. A conflicting study demonstrated that gold nanoparticles alone did not induce cytokines from macrophages, and that small ∼4 nm gold nanoparticles actually inhibited TLR9 signaling significantly compared to large ∼45 nm gold nanoparticles 44. Size clearly elicits innate immune effects, but appears context dependent.
Scavenger receptors (SRs) are a group of receptors expressed on many APCs that bind lipoproteins and other macromolecules to clear them from circulation. The affinity of SRs for nanoparticles may be somewhat related to negative charge and surface adsorbed protein (both properties that can correlate with size). Class A SRs are one major pathway for nanoparticle internalization 45.The family member MARCO has been extensively characterized as a major receptor for particle interactions. Initially thought to only bind microparticles, MARCO has now been shown to directly bind and internalize nanoparticles as small as 20 nm 46. MARCO engagement is known to activate immune responses, notably the secretion of inflammatory cytokines 47. However, the immunologic consequences of MARCO engagement appear to be extremely context dependent, as demonstrated by the successful suppression of autoimmunity using antigen-coupled, inert polymeric nanoparticles ∼500 nm 48.
Overall, smaller nanoparticles seem to have increased uptake and induce more immunogenic and cytotoxic responses in APCs. Indeed, ∼40-50 nm polystyrene beads proved to be highly immunogenic and reliant upon interactions with DCs in vivo 49. These authors hypothesized that the nano-size range bears similarity to viral particles, and thus may be an inherent, conserved immunogenic signal. Supporting the concept of immunogenic size, in vitro experiments showed that DCs cultured with 40 nm gold nanoparticles secreted inflammatory cytokines TNF-α, IL-6, IL-12, and GM-CSF; 20 nm gold nanoparticles induced no inflammatory activity 50. The in vivo consequences of size are likely specific for anatomic location and cellular interactions. For example, ∼400 nm poly(lactic-co-glycolic) nanoparticles incubated in vitro with a macrophage cell line show no immunostimulatory response 51. Therefore, the design and fate of nanoparticles that are phagocytosed by APCs are crucial to consider. Part of the size-dependent responses and differential behaviors may be attributed to differences in physical properties including available surface area and zeta potential, both of which can be influenced by size. To that end, there has been research looking at the role of nanoparticle surface charge and chemistry on activation showing that making surfaces more negative and increasing hydroxylation of the surfaces can increase complement activation 52, an important consideration for immunological engineering of particles.
Surface properties and modification
Not surprisingly, the surface molecules on nanomaterials, particularly those that are water soluble, play an important role in complement activation. Surface modification of poly(lactic-co-glycolic acid) nanoparticles with glucosamine has been shown to reduce complement activation 53. Dextrans can reduce complement activation at high surface coverage where there is limited ability for C3 to adsorb 54. A similar effect is seen with high PEGylation, and the density and organization of PEG matters. Brush conformation for PEG is critical to avoid activating complement and this requires PEG of specific molecular weight and density on the surface of the particle 55. Another way to think about this is that PEGylation for stealth may require very specific tuning to achieve the optimal effect of increased half-life with minimal immunotoxicity, particularly with repeated doses of PEGylated nanoparticles 56. This is at least partially due to anti-PEG antibodies. Anti-PEG antibodies are even reported to develop in response to empty ∼100 nm PEGylated liposomes, suggesting an inherent immunogenicity of PEG that may accelerate or promote opsonization and complement activation upon repeated dosing 57, 58. It is quite possible that with higher densities of PEG promoting brush conformation that these antibodies to PEG may be slower to form, but this hypothesis remains to be tested.
Receptor-based recognition of components, charge
All biomaterials have the potential to be recognized by specific receptors through interactions with their protein coronas or directly though the material itself. Innate immune cells have diverse receptor families that recognize potential danger signals, particulate and opsonized complexes, and patterns unique to foreign microbes that initiate inflammatory signaling and phagocytic uptake; these include TLRs, SRs, and inflammsomes. Rigotti et al. identified the first well-defined receptors (class B SRs, SR-B1 and CD36) that potently bind anionic lipids and lipoproteins, in particular phosphatidylserine and phosphatidylinositol liposomes 59. These receptors are preferentially expressed on APCs such as macrophages, and perform a native immunologic function: binding of inner-leaflet plasma membrane phosphatidylserine on apoptotic cells to induce anti-inflammatory molecules as well as reduce APC activation. Indeed, the negatively charged phosphatidylserine-containing liposomes have successfully been used to reduce inflammation in models of delayed phase edema 60. Functionalized polystyrene nanoparticles (both cationic and anionic) in the presence of human serum are largely internalized by human macrophages via CD64, the high affinity Fc receptor for IgG antibodies 61. CD64 activation can serve as immunogenic or immunosuppressive depending on context. Intrinsic lipoprotein receptors also internalize nanomaterials into APCs after they acquisition of a surface corona of serum components.
A striking study by Schmidt et al. demonstrated that TLR4 directly mediated allergic reactions to the metal nickel 62. The nature of the binding was a chelating pocket for the metal in the binding site of TLR4. A similar study described the potential direct triggering of TLR3 by gold in DCs 63. Quantum dots with either carboxyl or PEG functionalized surfaces were both shown to activate inflammatory responses in macrophages, and these were abrogated by TLR4 inhibition 64. In contrast to those studies, gold nanoparticles were shown to provoke no inherent immune response in macrophages; instead, they directly bound lysosomal HMGB1 and inhibited TLR9 signaling 65. These surprisingly specific metal-receptor interactions leave open the possibility that many innate immune effects of metal-based nanoparticles may be mediated by specific receptor recognition and signaling.
Cationic liposomes favorably interact with DCs 66 for uptake and, compared to macrophages, had significantly slower uptake of phosphatidylserine liposomes. This may be attributed to the concept that DCs ingest apoptotic cell bodies for debris clearance. While these differences appear significant for cell uptake in vitro, differentially charged and targeted nanoparticles yielded similar immune responses after in vivo administration 67. Cationic sensing may also involve TLRs. For example, cationic polylysine, dextran, and gelatin were all shown to at least partially stimulate MyD88-dependent sensing, generating Th1-type in cultured peritoneal macrophages and co-localization of biomaterials with TLR4 68.
In addition to lipids, polymers can also be recognized by a variety of immune cells. Polymeric carrier subunits can be haptenated (a process where small molecules complex with carriers to create an immunogen) to generate IgM antibodies by directly activating B cells 69. This study utilized fluorescein as a hapten and determined that commonly used polymers including Ficoll, dextran, cellulose, and polyvinyl alcohol could all be made immunogenic after haptens exceeded thresholds for molecular weight and valency. Polyanyhydride nanoparticles directly triggered TLR-2, -4, and -5 to promote Th1 responses in DCs 70. Interestingly, this study showed that pre-immunization with empty nanoparticles protected mice against a lethal Salmonella challenge, suggesting increased CD8+ T cell responsiveness caused by nanoparticles alone. In fact, diverse TLRs (TLR-4, in addition to TLR2 and -6) can be engaged by polymers on DCs 71, suggesting a potential universal recognition mechanism for polymers. Cumulative evidence above and structural information on the nature of TLR recognition indicate that the exposed hydrophobic regions of polymers or adsorbed proteins may be a main driver of these interactions. Hydrophobicity is hypothesized to serve as a conserved type of danger signal that may be detected as either abnormally folded/exposed protein regions or in various nanoparticle materials 72.
By further determining what receptors can directly recognize biomaterials, and the molecular and biophysical determinants of recognition, there remain abundant opportunities to tune either biomaterial composition or properties to avoid recognition or to actively monitor and suppress receptor recognition or immune response during nanoparticle administration.
The charge on the surface of a nanoparticle has a significant effect on complement activation. Negatively-charged liposomes increase complement activation and CARPA in vivo 17. More negative charge increases both hemolysis and complement activation on a range of particles in vitro 35-37. Increasing hydroxylation of surfaces on more negatively charged particles increases complement activation in vitro suggesting, based on the work looking at the role of hydroxyls on surfaces 52, that complement activation may be due, at least in part, to the presence of hydroxyl molecules that can covalently bind to C3b 25. As summarized in Table 2, critical features of nanomaterials that are found to dictate innate immune interactions include size, surface molecule coating, and net charge, and these properties all interact with immunity via diverse pathways that are context-dependent in vitro and in vivo.
Table 2. Critical features of nanomaterials with respect to the innate immune response.
| Biomaterials feature | Innate immune response | In vitro or in vivo | References |
|---|---|---|---|
| Size (smaller increases response) |
|
Both in vitro and in vivo with very similar responses | 39-44 |
| Surface molecules |
|
In vitro and in vivo | 53;54, 56-58 |
| Charge |
|
In vitro, the effects are often magnified with negatively charged particles, but in vivo, both positively and negatively charged particles are associated with receptor binding | 35-37, 52, 59-61 |
Bio-Inspired Solutions for Shielding Nanomaterials from Immunity
In addition to the potential pharmacological solutions indicated above, there may be multiple natural strategies for reducing the immune toxicity of nanoparticles. The increasing potential for anti-PEG immune responses also suggests alternate strategies are needed for shielding nanoparticles from immune detection and responses. Here we present a few emerging strategies that may shield nanoparticles from immune detection.
Calcium phosphate nanoparticles are most commonly known for their ability to deliver biomacromolecules across cell membranes in vitro, notably nucleic acids. However, amorphous nanoparticles made of calcium phosphate have recently been posited to have a critical, endogenous role in generating tolerance to gut microbial products 73. In this study, the authors investigated amorphous calcium phosphate particles and their interactions with Peyer's patches, lymphoid tissue structures found in the gut. These structures serve as a crucial interface between gut luminal contents and the immune system, with an overlying epithelial barrier and underlying tissue rich with cells of the innate immune system including APCs. Previous work determined that Peyer's patches have high uptake and transport of orally delivered nanoparticles ∼200 nm and smaller 74. Powell et al. demonstrated that amorphous calcium phosphate particle uptake is mediated almost completely by the highly specialized epithelial M cells, and that endocytic trafficking occurred without enzymatic degradation before trans-epithelial delivery to tissue APCs. Further, amorphous calcium phosphate particles incorporated peptidoglycan, a constitutive product of gut commensal microbes. The co-delivery of peptidoglycan signaling to APCs induced the expression of the tolerogenic molecule PD-L1. These results indicate that the gut nanomineral pathway may be a homeostatic mechanism for tuning gut innate immunity. Given the tolerogenic fate of this pathway, targeting nanoparticles to the gut using nanominerals may provide a new immune silent pathway for stealth nanoparticle uptake and systemic delivery after oral or gastrointestinal delivery. Determining the biochemical and physical properties required to take advantage of this pathway should be a focus of future work to fully utilize the tolerogenic transcytosis of the gut.
Another emerging strategy for creating stealth materials uses biomimetic or bio-derived coatings. Red blood cell (RBC) membrane-coated nanoparticles have been developed to reduce circulating alloantibody concentrations in in a model of hemolytic transfusion 75. The protein CD47 has been identified as one crucial component of RBC membranes that results in increased circulation and decreased macrophage uptake compared to PEGylation 76, 77. CD47-based self-stealthing has been further developed, including the identification of minimal peptides to inhibit phagocytosis 78. In addition, other properties of RBC membranes are suggested to result in inherently neutral immune processing, with no detectable anti-RBC protein responses arising in a vaccine nanotoxin preparation 79. Nanoparticles have also been coated with leukocyte membranes, yielding similar stealthing but enhanced receptor-ligand interactions 80. Coating with cancer cell membranes yields antigen presentation for effective vaccination as well as homotypic cancer-cell fusion of coated nanoparticles in situ 81. These developing techniques may greatly enhance our ability to stealth nanoparticles from the immune system and modulate their function, mitigating issues such as the emergence of anti-PEG immunity and targeting uptake/delivery.
What is critical at this point for understanding the relationship between molecular design and immune activation is taking all of this data and working through the specifics of features like surface charge and density of corona molecules to be able to go beyond saying something does or does not lead to immune activation to determine specific ranges for these values to direct the design of nanomedicines. Towards this goal, future work should focus on increasing our ability to screen and predict immunotoxicity of various nanomaterials in a variety of assays. Particularly useful would be prognostic signatures or biomarkers that would direct the design and choice of nanomaterials.
Models for Assessing Immune Activation by Nanomaterials
Activation of the complement cascade can be measured both in vitro and in vivo. There are a classic set of assays in vitro in human blood predicated on the measurement of one or more of the complement proteins. Traditionally, researchers looking at complement activation on biomaterials have focused on changes in C3, C3a, and C5a following incubation of the material in human serum 52. To distinguish between the classical and alternative pathways, one incubates the serum with EDTA or EGTA and then adds the nanoparticles of interest. The classical pathway is both Ca and Mg dependent whereas the alternative pathway is only Mg dependent 55. Assuming that most nanomaterials activate complement via the alternative pathway, incubation with EGTA will not affect complement activation but EDTA will. The data regarding the ratio of C3 to C3a and C5a are complemented by measurements of SC5b-9 (terminal membrane attack complex) following incubation of serum with the nanomaterial. Ultimately, though, one would like to be able to look at the complement response not only in vitro but in vivo to correlate it with the in vitro findings particularly with regards to complications such as the CARPA response.
The CARPA response exists both in rats and pigs, but the response in rats is far reduced in intensity and only seen at high doses. Pigs, like people, exhibit the response at very modest doses of nanoparticles (0.1 mg/kg) 82. Thus, there is great value in also looking at the porcine response to nanomaterials for changes in complement. The challenge is that there are far fewer ELISAs that are sensitive to porcine complement. Thus, an alternative used regularly in the field is the complement hemolytic activity test (CH50 test) in which the incubated serum is added to antibody-sensitized sheep erythrocytes 82, 83. It is not as sensitive as ELISAs but provides a welcome alternative to quantifying complement in vitro and in vivo in the porcine model.
Many other immunoassays could be used to assess immunotoxicity of nanoparticles 84. The development of high-throughput, multiplex panels for quantitation of bioanalytes (e.g., cytokines, chemokines, antibodies, other proteins and biomolecules) makes for an ideal diagnostic platform, both preclinically and clinically 85-88. Prior to administration of therapy, patients could be challenged with low doses of biomaterials to determine whether there is potential for a hyperactive immune response by measuring circulating protein levels. Other potential diagnostics could directly test the activity of immune cells in blood ex vivo or cultured with biomaterials using cytokine output and/or other functional outcomes (cell proliferation, upregulation of activation-associated genes or proteins) 89, 90; other alternatives could use a more traditional grading approach based on gross clinical symptoms 91. Ultimately, these diagnostics before administration or at the time of reaction could help identify targets for intervention; as an example, IL-6 was successfully utilized to alleviate cytokine storm in a tumor immunotherapy clinical case 92.
Conclusions
The clinical application of nanomedicines and biomaterials is expanding quickly. In concert, many studies have investigated potential complications for nanoparticles in clinical implementation, in particular their potential for immune recognition and immunotoxicity. While diverse mechanisms and pathways have been implicated in responding to nanoparticle properties and composition, relatively little has been definitively shown about the specific structural and biophysical designs that elicit various immune reactions; future studies need to more exhaustively determine the design parameters causing these interactions. A number of promising strategies are emerging, in particular bio-inspired and bio-mimetics, which may enable a more advanced approach for stealthing of nanomedicines than traditional approaches such as PEGylation. These new approaches will be complemented by and directly benefit from more advanced studies on how precise molecular and physical properties guide immune outcomes, and rational, precise ways these properties can be modulated to create immune stealth or neutral outcomes for novel therapies.
Figure 1. Features of nanoparticles associated with their innate immune responses.
Acknowledgments
This work was supported in part by the NIH Director's New Innovator Award, DP2OD007338.
References
- 1.Zipfel PF, Skerka C. Nature reviews Immunology. 2009;9:729–740. doi: 10.1038/nri2620. [DOI] [PubMed] [Google Scholar]
- 2.Brennan FH, Gordon R, Lao HW, Biggins PJ, Taylor SM, Franklin RJ, Woodruff TM, Ruitenberg MJ. J Neurosci. 2015;35:6517–6531. doi: 10.1523/JNEUROSCI.5218-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vakeva A, Morgan BP, Tikkanen I, Helin K, Laurila P, Meri S. Am J Pathol. 1994;144:1357–1368. [PMC free article] [PubMed] [Google Scholar]
- 4.Brubaker SW, Bonham KS, Zanoni I, Kagan JC. Annual review of immunology. 2015;33:257–290. doi: 10.1146/annurev-immunol-032414-112240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Goram AL, Richmond PL. Pharmacotherapy. 2001;21:751–763. doi: 10.1592/phco.21.7.751.34572. [DOI] [PubMed] [Google Scholar]
- 6.Banda NK, Mehta G, Chao Y, Wang G, Inturi S, Fossati-Jimack L, Botto M, Wu L, Moghimi S, Simberg D. Particle and fibre toxicology. 2014;11:64. doi: 10.1186/s12989-014-0064-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fornaguera C, Caldero G, Mitjans M, Vinardell MP, Solans C, Vauthier C. Nanoscale. 2015;7:6045–6058. doi: 10.1039/c5nr00733j. [DOI] [PubMed] [Google Scholar]
- 8.Andersen AJ, Wibroe PP, Moghimi SM. Adv Drug Deliv Rev. 2012;64:1700–1705. doi: 10.1016/j.addr.2012.05.005. [DOI] [PubMed] [Google Scholar]
- 9.Li H, Willingham SB, Ting JP, Re F. Journal of immunology (Baltimore, Md : 1950) 2008;181:17–21. doi: 10.4049/jimmunol.181.1.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kool M, Petrilli V, De Smedt T, Rolaz A, Hammad H, van Nimwegen M, Bergen IM, Castillo R, Lambrecht BN, Tschopp J. Journal of immunology (Baltimore, Md : 1950) 2008;181:3755–3759. doi: 10.4049/jimmunol.181.6.3755. [DOI] [PubMed] [Google Scholar]
- 11.Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL, Fitzgerald KA, Latz E. Nature immunology. 2008;9:847–856. doi: 10.1038/ni.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cassel SL, Eisenbarth SC, Iyer SS, Sadler JJ, Colegio OR, Tephly LA, Carter AB, Rothman PB, Flavell RA, Sutterwala FS. Proc Natl Acad Sci U S A. 2008;105:9035–9040. doi: 10.1073/pnas.0803933105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kuroda E, Ishii KJ, Uematsu S, Ohata K, Coban C, Akira S, Aritake K, Urade Y, Morimoto Y. Immunity. 2011;34:514–526. doi: 10.1016/j.immuni.2011.03.019. [DOI] [PubMed] [Google Scholar]
- 14.Kusaka T, Nakayama M, Nakamura K, Ishimiya M, Furusawa E, Ogasawara K. PLoS One. 2014;9:e92634. doi: 10.1371/journal.pone.0092634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yazdi AS, Guarda G, Riteau N, Drexler SK, Tardivel A, Couillin I, Tschopp J. Proc Natl Acad Sci U S A. 2010;107:19449–19454. doi: 10.1073/pnas.1008155107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Szebeni J, Bedocs P, Rozsnyay Z, Weiszhar Z, Urbanics R, Rosivall L, Cohen R, Garbuzenko O, Bathori G, Toth M, Bunger R, Barenholz Y. Nanomedicine. 2012;8:176–184. doi: 10.1016/j.nano.2011.06.003. [DOI] [PubMed] [Google Scholar]
- 17.Szebeni J, Bedocs P, Csukas D, Rosivall L, Bunger R, Urbanics R. Adv Drug Deliv Rev. 2012;64:1706–1716. doi: 10.1016/j.addr.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 18.Johnson RA, Simmons KT, Fast JP, Schroeder CA, Pearce RA, Albrecht RM, Mecozzi S. J Pharm Sci. 2011;100:2685–2692. doi: 10.1002/jps.22488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Moghimi SM, Wibroe PP, Helvig SY, Farhangrazi ZS, Hunter AC. Adv Drug Deliv Rev. 2012;64:1385–1393. doi: 10.1016/j.addr.2012.05.010. [DOI] [PubMed] [Google Scholar]
- 20.Vallhov H, Qin J, Johansson SM, Ahlborg N, Muhammed MA, Scheynius A, Gabrielsson S. Nano Lett. 2006;6:1682–1686. doi: 10.1021/nl060860z. [DOI] [PubMed] [Google Scholar]
- 21.Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, Katze MG. Microbiology and molecular biology reviews : MMBR. 2012;76:16–32. doi: 10.1128/MMBR.05015-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Suntharalingam G, Perry MR, Ward S, Brett SJ, Castello-Cortes A, Brunner MD, Panoskaltsis N. N Engl J Med. 2006;355:1018–1028. doi: 10.1056/NEJMoa063842. [DOI] [PubMed] [Google Scholar]
- 23.Szebeni J. Critical Reviews™ in Therapeutic Drug Carrier Systems. 1998;15 [PubMed] [Google Scholar]
- 24.Szebeni J, Muggia F, Gabizon A, Barenholz Y. Adv Drug Deliv Rev. 2011;63:1020–1030. doi: 10.1016/j.addr.2011.06.017. [DOI] [PubMed] [Google Scholar]
- 25.Janssen BJ, Huizinga EG, Raaijmakers HC, Roos A, Daha MR, Nilsson-Ekdahl K, Nilsson B, Gros P. Nature. 2005;437:505–511. doi: 10.1038/nature04005. [DOI] [PubMed] [Google Scholar]
- 26.Natale R, Socinski M, Hart L, Lipaytov O, Spigel D, Gershenhorn B, Weiss G, Kazmi S, Karaseva N, Gladkov O, Moiseyenko V, Summa J, Youssoufian H, Otterson G. presented in part at the EORTC-NCI0AACR Annual Symposium; Barcelona, Spain. 2014. [Google Scholar]
- 27.Szebeni J, Bedocs P, Urbanics R, Bunger R, Rosivall L, Toth M, Barenholz Y. J Control Release. 2012;160:382–387. doi: 10.1016/j.jconrel.2012.02.029. [DOI] [PubMed] [Google Scholar]
- 28.Fan H, Cook JA. Journal of endotoxin research. 2004;10:71–84. doi: 10.1179/096805104225003997. [DOI] [PubMed] [Google Scholar]
- 29.Kheirabadi BS, Mace JE, Terrazas IB, Fedyk CG, Estep JS, Dubick MA, Blackbourne LH. J Trauma. 2010;68:269–278. doi: 10.1097/TA.0b013e3181c97ef1. [DOI] [PubMed] [Google Scholar]
- 30.Okamura Y, Fukui Y, Kabata K, Suzuki H, Handa M, Ikeda Y, Takeoka S. Bioconjugate Chemistry. 2009;20:1958–1965. doi: 10.1021/bc900325w. [DOI] [PubMed] [Google Scholar]
- 31.Shoffstall AJ, Everhart LM, Varley ME, Soehnlen ES, Shick AM, Ustin JS, Lavik EB. Biomacromolecules. 2013 doi: 10.1021/bm400619v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brown AC, Stabenfeldt SE, Ahn B, Hannan RT, Dhada KS, Herman ES, Stefanelli V, Guzzetta N, Alexeev A, Lam WA, Lyon LA, Barker TH. Nat Mater. 2014;13:1108–1114. doi: 10.1038/nmat4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chan LW, Wang X, Wei H, Pozzo LD, White NJ, Pun SH. Sci Transl Med. 2015;7 doi: 10.1126/scitranslmed.3010383. 277ra229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chan LW, White NJ, Pun SH. Bioconjug Chem. 2015;26:1224–1236. doi: 10.1021/acs.bioconjchem.5b00070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Dash BC, Rethore G, Monaghan M, Fitzgerald K, Gallagher W, Pandit A. Biomaterials. 2010;31:8188–8197. doi: 10.1016/j.biomaterials.2010.07.067. [DOI] [PubMed] [Google Scholar]
- 36.Pham CT, Thomas DG, Beiser J, Mitchell LM, Huang JL, Senpan A, Hu G, Gordon M, Baker NA, Pan D, Lanza GM, Hourcade DE. Nanomedicine. 2013 doi: 10.1016/j.nano.2013.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Robbins GR, Roberts RA, Guo H, Reuter K, Shen T, Sempowski GD, McKinnon KP, Su L, DeSimone JM, Ting JP. Nanomedicine. 2015 doi: 10.1016/j.nano.2014.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hadjidemetriou M, Al-Ahmady Z, Mazza M, Collins RF, Dawson K, Kostarelos K. ACS Nano. 2015 doi: 10.1021/acsnano.5b03300. [DOI] [PubMed] [Google Scholar]
- 39.Pacheco PM, Le B, White D, Sulchek T. Nano LIFE. 2013;3:1341001. doi: 10.1142/S1793984413410018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Veiseh O, Doloff JC, Ma M, Vegas AJ, Tam HH, Bader AR, Li J, Langan E, Wyckoff J, Loo WS, Jhunjhunwala S, Chiu A, Siebert S, Tang K, Hollister-Lock J, Aresta-Dasilva S, Bochenek M, Mendoza-Elias J, Wang Y, Qi M, Lavin DM, Chen M, Dholakia N, Thakrar R, Lacik I, Weir GC, Oberholzer J, Greiner DL, Langer R, Anderson DG. Nat Mater. 2015;14:643–651. doi: 10.1038/nmat4290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Silva AL, Rosalia RA, Varypataki E, Sibuea S, Ossendorp F, Jiskoot W. Vaccine. 2015;33:847–854. doi: 10.1016/j.vaccine.2014.12.059. [DOI] [PubMed] [Google Scholar]
- 42.Schanen BC, Karakoti AS, Seal S, Drake DR, 3rd, Warren WL, Self WT. ACS Nano. 2009;3:2523–2532. doi: 10.1021/nn900403h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yen HJ, Hsu SH, Tsai CL. Small (Weinheim an der Bergstrasse, Germany) 2009;5:1553–1561. doi: 10.1002/smll.200900126. [DOI] [PubMed] [Google Scholar]
- 44.Pan Y, Neuss S, Leifert A, Fischler M, Wen F, Simon U, Schmid G, Brandau W, Jahnen-Dechent W. Small (Weinheim an der Bergstrasse, Germany) 2007;3:1941–1949. doi: 10.1002/smll.200700378. [DOI] [PubMed] [Google Scholar]
- 45.Wang H, Wu L, Reinhard BM. ACS Nano. 2012;6:7122–7132. doi: 10.1021/nn302186n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kanno S, Furuyama A, Hirano S. Toxicological sciences : an official journal of the Society of Toxicology. 2007;97:398–406. doi: 10.1093/toxsci/kfm050. [DOI] [PubMed] [Google Scholar]
- 47.Coller SP, Paulnock DM. Journal of leukocyte biology. 2001;70:142–148. [PubMed] [Google Scholar]
- 48.Getts DR, Martin AJ, McCarthy DP, Terry RL, Hunter ZN, Yap WT, Getts MT, Pleiss M, Luo X, King NJ, Shea LD, Miller SD. Nat Biotechnol. 2012;30:1217–1224. doi: 10.1038/nbt.2434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fifis T, Gamvrellis A, Crimeen-Irwin B, Pietersz GA, Li J, Mottram PL, McKenzie IF, Plebanski M. Journal of immunology (Baltimore, Md : 1950) 2004;173:3148–3154. doi: 10.4049/jimmunol.173.5.3148. [DOI] [PubMed] [Google Scholar]
- 50.Niikura K, Matsunaga T, Suzuki T, Kobayashi S, Yamaguchi H, Orba Y, Kawaguchi A, Hasegawa H, Kajino K, Ninomiya T, Ijiro K, Sawa H. ACS Nano. 2013;7:3926–3938. doi: 10.1021/nn3057005. [DOI] [PubMed] [Google Scholar]
- 51.Nicolete R, dos Santos DF, Faccioli LH. International immunopharmacology. 2011;11:1557–1563. doi: 10.1016/j.intimp.2011.05.014. [DOI] [PubMed] [Google Scholar]
- 52.Thomas SN, van der Vlies AJ, O'Neil CP, Reddy ST, Yu SS, Giorgio TD, Swartz MA, Hubbell JA. Biomaterials. 2011;32:2194–2203. doi: 10.1016/j.biomaterials.2010.11.037. [DOI] [PubMed] [Google Scholar]
- 53.Thasneem YM, Sajeesh S, Sharma CP. Colloids Surf B Biointerfaces. 2013;108:337–344. doi: 10.1016/j.colsurfb.2013.03.001. [DOI] [PubMed] [Google Scholar]
- 54.Vauthier C, Persson B, Lindner P, Cabane B. Biomaterials. 2011;32:1646–1656. doi: 10.1016/j.biomaterials.2010.10.026. [DOI] [PubMed] [Google Scholar]
- 55.Hamad I, Al-Hanbali O, Hunter AC, Rutt KJ, Andresen TL, Moghimi SM. ACS Nano. 2010;4:6629–6638. doi: 10.1021/nn101990a. [DOI] [PubMed] [Google Scholar]
- 56.Li SD, Huang L. J Control Release. 2010;145:178–181. doi: 10.1016/j.jconrel.2010.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wang X, Ishida T, Kiwada H. J Control Release. 2007;119:236–244. doi: 10.1016/j.jconrel.2007.02.010. [DOI] [PubMed] [Google Scholar]
- 58.Ishida T, Wang X, Shimizu T, Nawata K, Kiwada H. J Control Release. 2007;122:349–355. doi: 10.1016/j.jconrel.2007.05.015. [DOI] [PubMed] [Google Scholar]
- 59.Rigotti A, Acton SL, Krieger M. J Biol Chem. 1995;270:16221–16224. doi: 10.1074/jbc.270.27.16221. [DOI] [PubMed] [Google Scholar]
- 60.Ramos GC, Fernandes D, Charao CT, Souza DG, Teixeira MM, Assreuy J. Br J Pharmacol. 2007;151:844–850. doi: 10.1038/sj.bjp.0707302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lunov O, Syrovets T, Loos C, Beil J, Delacher M, Tron K, Nienhaus GU, Musyanovych A, Mailander V, Landfester K, Simmet T. ACS Nano. 2011;5:1657–1669. doi: 10.1021/nn2000756. [DOI] [PubMed] [Google Scholar]
- 62.Schmidt M, Raghavan B, Muller V, Vogl T, Fejer G, Tchaptchet S, Keck S, Kalis C, Nielsen PJ, Galanos C, Roth J, Skerra A, Martin SF, Freudenberg MA, Goebeler M. Nature immunology. 2010;11:814–819. doi: 10.1038/ni.1919. [DOI] [PubMed] [Google Scholar]
- 63.Rachmawati D, Alsalem IW, Bontkes HJ, Verstege MI, Gibbs S, von Blomberg BM, Scheper RJ, van Hoogstraten IM. Toxicology in vitro : an international journal published in association with BIBRA. 2015;29:363–369. doi: 10.1016/j.tiv.2014.10.010. [DOI] [PubMed] [Google Scholar]
- 64.Ho CC, Luo YH, Chuang TH, Yang CS, Ling YC, Lin P. Toxicology. 2013;308:1–9. doi: 10.1016/j.tox.2013.03.003. [DOI] [PubMed] [Google Scholar]
- 65.Tsai CY, Lu SL, Hu CW, Yeh CS, Lee GB, Lei HY. Journal of immunology (Baltimore, Md : 1950) 2012;188:68–76. doi: 10.4049/jimmunol.1100344. [DOI] [PubMed] [Google Scholar]
- 66.Foged C, Arigita C, Sundblad A, Jiskoot W, Storm G, Frokjaer S. Vaccine. 2004;22:1903–1913. doi: 10.1016/j.vaccine.2003.11.008. [DOI] [PubMed] [Google Scholar]
- 67.Arigita C, Bevaart L, Everse LA, Koning GA, Hennink WE, Crommelin DJ, van de Winkel JG, van Vugt MJ, Kersten GF, Jiskoot W. Infection and immunity. 2003;71:5210–5218. doi: 10.1128/IAI.71.9.5210-5218.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Chen H, Li P, Yin Y, Cai X, Huang Z, Chen J, Dong L, Zhang J. Biomaterials. 2010;31:8172–8180. doi: 10.1016/j.biomaterials.2010.07.056. [DOI] [PubMed] [Google Scholar]
- 69.Dintzis RZ, Okajima M, Middleton MH, Greene G, Dintzis HM. Journal of immunology (Baltimore, Md : 1950) 1989;143:1239–1244. [PubMed] [Google Scholar]
- 70.Tamayo I, Irache JM, Mansilla C, Ochoa-Reparaz J, Lasarte JJ, Gamazo C. Clinical and vaccine immunology : CVI. 2010;17:1356–1362. doi: 10.1128/CVI.00164-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Shokouhi B, Coban C, Hasirci V, Aydin E, Dhanasingh A, Shi N, Koyama S, Akira S, Zenke M, Sechi AS. Biomaterials. 2010;31:5759–5771. doi: 10.1016/j.biomaterials.2010.04.015. [DOI] [PubMed] [Google Scholar]
- 72.Seong SY, Matzinger P. Nature reviews Immunology. 2004;4:469–478. doi: 10.1038/nri1372. [DOI] [PubMed] [Google Scholar]
- 73.Powell JJ, Thomas-McKay E, Thoree V, Robertson J, Hewitt RE, Skepper JN, Brown A, Hernandez-Garrido JC, Midgley PA, Gomez-Morilla I, Grime GW, Kirkby KJ, Mabbott NA, Donaldson DS, Williams IR, Rios D, Girardin SE, Haas CT, Bruggraber SF, Laman JD, Tanriver Y, Lombardi G, Lechler R, Thompson RP, Pele LC. Nature nanotechnology. 2015;10:361–369. doi: 10.1038/nnano.2015.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Awaad A, Nakamura M, Ishimura K. Nanomedicine. 2012;8:627–636. doi: 10.1016/j.nano.2011.08.009. [DOI] [PubMed] [Google Scholar]
- 75.Copp JA, Fang RH, Luk BT, Hu CM, Gao W, Zhang K, Zhang L. Proc Natl Acad Sci U S A. 2014;111:13481–13486. doi: 10.1073/pnas.1412420111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hu CM, Zhang L, Aryal S, Cheung C, Fang RH, Zhang L. Proc Natl Acad Sci U S A. 2011;108:10980–10985. doi: 10.1073/pnas.1106634108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hu CM, Fang RH, Luk BT, Chen KN, Carpenter C, Gao W, Zhang K, Zhang L. Nanoscale. 2013;5:2664–2668. doi: 10.1039/c3nr00015j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rodriguez PL, Harada T, Christian DA, Pantano DA, Tsai RK, Discher DE. Science. 2013;339:971–975. doi: 10.1126/science.1229568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hu CM, Fang RH, Luk BT, Zhang L. Nature nanotechnology. 2013;8:933–938. doi: 10.1038/nnano.2013.254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Parodi A, Quattrocchi N, van de Ven AL, Chiappini C, Evangelopoulos M, Martinez JO, Brown BS, Khaled SZ, Yazdi IK, Enzo MV, Isenhart L, Ferrari M, Tasciotti E. Nature nanotechnology. 2013;8:61–68. doi: 10.1038/nnano.2012.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Fang RH, Hu CM, Luk BT, Gao W, Copp JA, Tai Y, O'Connor DE, Zhang L. Nano Lett. 2014;14:2181–2188. doi: 10.1021/nl500618u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Dezsi L, Fulop T, Meszaros T, Szenasi G, Urbanics R, Vazsonyi C, Orfi E, Rosivall L, Nemes R, Kok RJ, Metselaar JM, Storm G, Szebeni J. J Control Release. 2014;195:2–10. doi: 10.1016/j.jconrel.2014.08.009. [DOI] [PubMed] [Google Scholar]
- 83.Yu K, Lai BF, Foley JH, Krisinger MJ, Conway EM, Kizhakkedathu JN. ACS Nano. 2014;8:7687–7703. doi: 10.1021/nn504186b. [DOI] [PubMed] [Google Scholar]
- 84.Dobrovolskaia MA, Germolec DR, Weaver JL. Nature nanotechnology. 2009;4:411–414. doi: 10.1038/nnano.2009.175. [DOI] [PubMed] [Google Scholar]
- 85.Chowdhury F, Williams A, Johnson P. J Immunol Methods. 2009;340:55–64. doi: 10.1016/j.jim.2008.10.002. [DOI] [PubMed] [Google Scholar]
- 86.Tarrant JM. Toxicological sciences : an official journal of the Society of Toxicology. 2010;117:4–16. doi: 10.1093/toxsci/kfq134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Elsabahy M, Wooley KL. Chemical Society reviews. 2013;42:5552–5576. doi: 10.1039/c3cs60064e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Xu XJ, Tang YM, Song H, Yang SL, Xu WQ, Zhao N, Shi SW, Shen HP, Mao JQ, Zhang LY, Pan BH. The Journal of pediatrics. 2012;160:984–990. doi: 10.1016/j.jpeds.2011.11.046. e981. [DOI] [PubMed] [Google Scholar]
- 89.Romer PS, Berr S, Avota E, Na SY, Battaglia M, ten Berge I, Einsele H, Hunig T. Blood. 2011;118:6772–6782. doi: 10.1182/blood-2010-12-319780. [DOI] [PubMed] [Google Scholar]
- 90.Guo Y, Luan L, Rabacal W, Bohannon JK, Fensterheim BA, Hernandez A, Sherwood ER. Journal of immunology (Baltimore, Md : 1950) 2015;195:2353–2364. doi: 10.4049/jimmunol.1500300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Lee DW, Gardner R, Porter DL, Louis CU, Ahmed N, Jensen M, Grupp SA, Mackall CL. Blood. 2014;124:188–195. doi: 10.1182/blood-2014-05-552729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Teachey DT, Rheingold SR, Maude SL, Zugmaier G, Barrett DM, Seif AE, Nichols KE, Suppa EK, Kalos M, Berg RA, Fitzgerald JC, Aplenc R, Gore L, Grupp SA. Blood. 2013;121:5154–5157. doi: 10.1182/blood-2013-02-485623. [DOI] [PMC free article] [PubMed] [Google Scholar]