

Abstract
Ketamine, a pediatric anesthetic, is a noncompetitive N-methyl-D-aspartic acid (NMDA) receptor antagonist. Studies show that ketamine is neurotoxic in developing mammals and zebrafish. In both mammals and zebrafish, acetyl L-carnitine (ALCAR) has been shown to be protective against ketamine toxicity. Ketamine is known to modulate the serotonergic system in mammals. Here, we measured the levels of serotonin (5-HT) and its metabolite, 5-hydroxyindoleacetic acid (5-HIAA) in the embryos exposed to ketamine in the presence and absence of ALCAR. Ketamine, at lower doses, did not produce significant changes in the 5-HT or 5-HIAA levels in 3 dpf (day post-fertilization) embryos. However, 2 mM ketamine (internal embryo exposure levels comparable to human anesthetic plasma concentration) significantly reduced 5-HT level, and 5-HIAA was not detectable indicating that 5-HT metabolism was abolished. In the presence or absence of 2 mM ketamine, ALCAR by itself did not significantly alter 5-HT or 5-HIAA levels compared to the control. Ratios of metabolite/5-HT indicated that 2 mM ketamine inhibited 5-HT metabolism to 5-HIAA whereas lower doses (0.1–0.3 mM) of ketamine did not have any effect. ALCAR reversed the effects of 2 mM ketamine not only by restoring 5-HT and 5-HIAA levels but also 5-HT turnover rate to control levels. Whole mount immunohistochemical studies showed that 2 mM ketamine reduced the serotonergic area in the brain whereas ALCAR expanded it with increased axonal sprouting and branching. These results indicate that ketamine and ALCAR have opposing effects on the zebrafish serotonergic system.
Keywords: Ketamine, Zebrafish, Acetyl L-carnitine, Serotonin, Metabolism, HPLC/EC
1. Introduction
Ketamine, a pediatric anesthetic, is a noncompetitive antagonistof the N-methyl-D-aspartic acid (NMDA) receptor [22]. Althougha dissociative anesthetic [10], ketamine renders analgesic andamnesic effects and its potential anti-depressant properties are alsobeing investigated [3,6,30,36]. A number of studies on rodents and non-human primates have shown that treatment with high doses of ketamine or exposure for long durations during susceptible periods of development can induce neuroapoptosis [14,16,39]. Recently, ketamine’s role as an anti-depressant has also been documented [29,38].
Acetyl L-carnitine (ALCAR) belongs to the family of carnitines, a group of naturally occurring compounds that is essential for β-oxidation of fatty acids in mitochondria to generate ATP [7]. ALCAR effectively prevents mitochondrial injury resulting from oxidative damage [33]. It is also suggested that carnitines have neuroprotective effects on conditions caused by mitochondrial dysfunction and oxidative stress and possibly in neurodegenerative disorders, such as Parkinson’s disease [5]. Moreover, carnitines can neutralize toxic acyl–CoA production in the mitochondria [41] that occurs in various CNS disorders, such as Parkinson’s disease [32]. ALCAR is also effective in reducing age-dependent progressive mitochondrial dysfunction and can restore mitochondrial membrane potential, metabolic oxygen consumption and β-oxidation of fatty acids [15]. For neurons at early developmental stages, co-administration of ALCAR significantly diminished reactive oxygen species (ROS) generation and provided significant protection of neurons from ketamine-induced neurodegeneration [26]. Additionally, ALCAR has been shown to protect neurons from inhalation anesthetic-induced neurotoxicity in rat cortical neurons [52]. In zebrafish embryos, ALCAR offers protection against ketamine-induced cardiotoxicity and neurotoxicity [11,19].
Serotonin (5-hydroxytryptamine; or 5-HT) and monoamine oxidase (MAO) are involved in a number of physiological functions and pathological conditions. Zebrafish serotonergic system and its development are similar to those of higher order vertebrates [37]. Therefore, zebrafish embryos/larvae can be a good model system to study the 5-HT system. For its small size, prolific reproductive capacity and easy maintenance, zebrafish possess the typical complexity of vertebrate systems, and therefore, have been used in several areas of research with the prospect of extrapolating findings to mammals including humans [8,20,24,34].
In rats, ketamine inhibits the uptake of serotonin (5-HT) both in vitro [4] and in vivo [27]. In the rat brain, ketamine inhibits 5-HT synthesis and metabolism in vivo [28]. Ketamine exerts its antidepressant effects only at lower doses while higher doses induce anesthesia [29]. Zebrafish embryos have a functional 5-HT system with MAO mRNA expression in the brain beginning at 24 h post-fertilization (hpf) and enzyme activity at 42 hpf [37]. In mammalian models and patients, the role of 5-HT in depressive disorders and the modulation of the 5-HT system by ketamine have been reported [46,49,50]. In the present study, we examined whether ketamine and ALCAR evoke a response in the 5-HT system in the zebrafish embryos in terms of 5-HT biosynthesis and metabolism.
2. Materials and methods
2.1. Animals
Adult wild type (WT) zebrafish (Danio rerio, AB strain) were obtained from the Zebrafish International Resource Center (www.zirc.org) (Eugene, OR, USA). The fish were kept in fish tanks (Aquatic Habitats, FL, USA) at the NCTR/FDA zebrafish facility containing buffered water (pH 7.5) at 28 °C, and were fed daily live brine shrimp and Zeigler dried flake food (Zeiglers, Gardeners, PA, USA). Each 3 l tank housed 8 adult males or 8 females. Handling and maintenance of zebrafish were in compliance with the NIH Guide for the care and use of laboratory animals and were approved by the NCTR/FDA IACUC. The day–night cycle was maintained at 14:10 h. For in-system breeding, crosses of males and females were set up the previous day with partitions that were taken off the following morning at the time of light onset at 7:30 AM to stimulate spawning and fertilization. Fertilized zebrafish eggs were collected from the bottom of the tank as soon as they were laid. The eggs were placed in Petri dishes and washed thoroughly with buffered egg water (reverse osmosis water containing 60 mg sea salt (Crystal Sea®, Aquatic Eco-systems, Inc., Apopka, FL, USA) per liter of water (pH 7.5) and then allowed to develop in an incubator at 28.5 °C for later use.
2.2. Reagents
Ketamine hydrochloride was purchased from Vedco, Inc. (St. Joseph, MO, USA). Acetyl L-carnitine (ALCAR) and a polyclonal anti-serotonin antibody were purchased from Sigma (St. Louis, MO, USA). All other reagents used in this study were purchased from Sigma (St. Louis, MO, USA) unless mentioned otherwise. Fluorescently labeled secondary antibody (Cy3-conjugated goat anti-rabbit antibody) was purchased from Jackson ImmunoResearch Laboratories (West Grove, PA, USA).
2.3. Treatment of zebrafish embryos with ketamine and acetyl L-carnitine
For treatment with ketamine and ALCAR, 52 hpf embryos were used. Fifty embryos were placed in individual 60 mm Petri dishes for each treatment. Ketamine and ALCAR treatments at various doses continued for 24 h (static exposures). Untreated control groups were examined in parallel. At 76 hpf (more than 72 hpf), zebrafish embryos are also called larvae. For consistency, we chose to use the term ‘embryo’ for 76 hpf zebrafish throughout this manuscript.
2.4. Determination of 5-HT and 5-HIAA by high-performance liquid chromatography with electrochemical detection (HPLC/EC)
Post-exposure to ketamine and ALCAR, zebrafish embryos (76 hpf) (50/sample) were homogenized in 100 ml of 0.2 μM perchloric acid containing 1 μM dihydroxybenzylamine (DHBA) as internal standard and centrifuged at 10,000 × g (10 min at 4 °C). Supernatant was removed and added to a filter tube containing a 0.2 μM membrane and spun at 4000 × g (4 min at 4 °C). The resulting filtrate was used for HPLC analysis of monoamines. A 25 μl aliquot was directly injected into an HPLC/EC detection system connected to BASi liquid chromatography pump (West Lafayette, IN, USA), a Supelco Supelcoil LC-18 (7.5 cm × 4.6 mm) analytical column, an amperometric detector and an LC-17 oxidative flow cell (BASi) with a glassy carbon electrode versus an Ag–AgCl reference electrode which was maintained at a potential of 0.75 V. The HPLC mobile phase consisted of potassium phosphate pH 3.0, methanol and heptane-sulfonic acid [1] and chromatograms were recorded and integrated using a BASi HPLC system with Epsilon Integrator. Concentrations of 5-HT and 5-HIAA were calculated against series of internally prepared, known standards [17]. Ratios of 5-HIAA/5-HT were calculated as estimated 5-HT release and turnover.
2.5. Immunocytochemistry for 5-HT (serotonergic) neurons in the brain
Embryos (n = 15/exposure group) post-exposure to ketamine and ALCAR were fixed with 4% paraformaldehyde in phosphate buffer, pH 7.3, overnight at 4 °C. Fixed embryos were washed 3 × 5 min each in phosphate buffer saline (PBS), pH 7.3 followed by permeabilization in acetone at −20 °C for 10 min. To reduce nonspecific binding by the anti-serotonin antibody, embryos were incubated in PBS plus 10% goat serum plus 1% Triton-X 100 (blocking buffer) for 1 h at room temperature. The embryos were incubated overnight at 4 °C with the anti-serotonin polyclonal antibody (1:500 dilution) in the blocking buffer. Following antibody incubation, embryos were washed 4 × 25 min each with the blocking buffer to remove excess primary antibody. Embryos were then incubated overnight at 4 °C in the blocking buffer containing Cy3-conjugated goat anti-rabbit anti-serum as the secondary antibody (1:100 dilution). Subsequent to multiple washes for 2 h in blocking buffer and a final wash with PBS for 30 min, labeled embryos were visualized and images acquired using a Nikon Eclipse Ni-U upright microscope.
2.6. Statistical analyses
In order to determine 5-HT and 5-HIAA levels, data from biological replicates were averaged. For all pairwise multiple comparison procedures, the ANOVA was used for data analysis with overall significance level set at P = < 0.05.
3. Results
3.1. Ketamine dose-dependently modulates 5-HT and 5-HIAA levels in the zebrafish embryos
Zebrafish embryos at 52 hpf were exposed to 0.1–0.3 mM ketamine for 24 h. In these 76 hpf embryos, none of the three doses (0.1, 0.2 and 0.3 mM) altered 5-HT or 5-HIAA levels when compared to the control (Fig. 1A–B). Our earlier studies demonstrated that exposure of embryos to 2 mM ketamine resulted in an internal exposure of ~8 μM [44]. This concentration is comparable to human anesthetic blood levels (2.2 μg/ml). Ketamine at 2 mM significantly reduced 5-HT level and its metabolite 5-HIAA was not detectable (Fig. 2A–B). These results indicated that 2 mM ketamine adversely affects 5-HT synthesis and turnover.
Fig. 1.
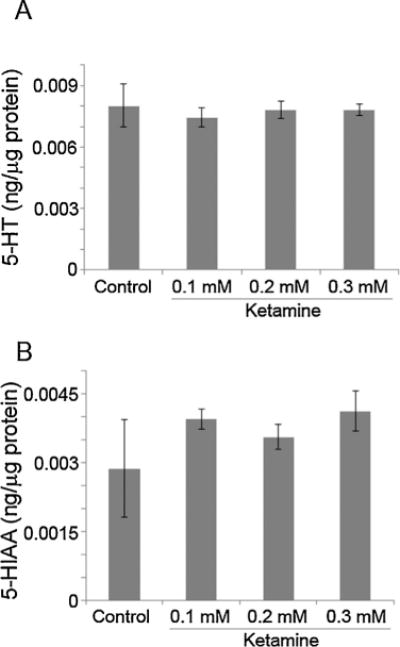
Effects of low doses of ketamine on 5-HT and 5-HIAA levels in zebrafish embryos.
Two day old embryos (52 hpf) were treated with 0.1–0.3 mM ketamine. Twenty four hours post exposure, 76 hpf embryos were processed for 5-HT (A) as well as its metabolite, 5-HIAA (B) measurements using HPLC/EC. Values are expressed in ng/μg protein in the embryo extract as mean ± SD. One-way ANOVA with post-hoc analyses were used to obtain statistical significance of variation in the 5-HT and 5-HIAA levels between the experimental groups as well as with the control.
Fig. 2.
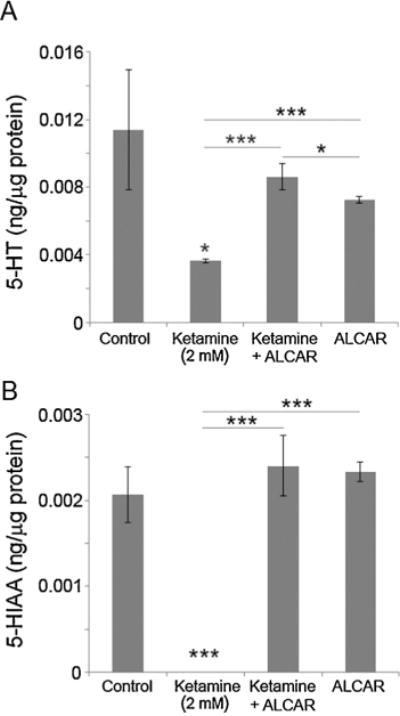
Effects of 2 mM ketamine (internal exposure comparable to human anesthetic plasma concentrations) and ALCAR on 5-HT and 5-HIAA levels in zebrafish embryos.
Embryos at 52 hpf were treated with 2 mM ketamine, 2 mM ketamine + 0.5 mM ALCAR and 0.5 mM ALCAR alone. Twenty four hours post exposure 76 hpf embryos were processed for 5-HT (A) as well as its metabolites, 5-HIAA (B) measurements using HPLC/EC. Values are expressed in ng/μg protein in the embryo extract as mean ± SD. One-way ANOVA with post-hoc analyses were used to obtain statistical significance (*P < 0.05; **P < 0.001; ***P < 0.0001) of variation in the 5-HT and its metabolites, 5-HIAA levels between the experimental groups as well as with the control.
3.2. Lower doses of ketamine do not affect 5-HT metabolism to 5-HIAA
The metabolite (5-HIAA)/5-HT ratios were used to determine the rate of 5-HT metabolism in zebrafish embryos. At low doses (0.1–0.3 mM), ketamine did not significantly alter 5-HT metabolism (Fig. 3A). In embryos treated with 2 mM ketamine, since 5-HIAA levels were not detectable, it is likely that at this dose, ketamine inhibited 5-HT turnover (Figs. 2B and 3B).
Fig. 3.
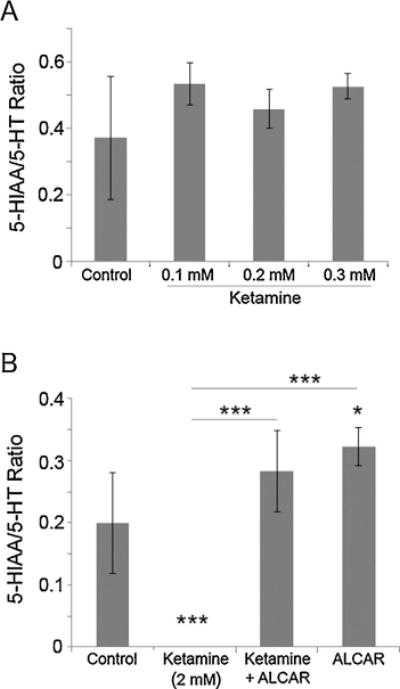
Effects of ketamine and ALCAR on 5-HT metabolism in zebrafish embryos. Embryos at 52 hpf were treated with lower doses of ketamine (0.1–0.3 mM) as well as 2 mM ketamine (internal exposure equivalent to human anesthetic dose), 2 mM ketamine + 0.5 mM ALCAR and 0.5 mM ALCAR alone. Twenty four hours post exposure 76 hpf embryos were processed for 5-HT as well as its metabolites, 5-HIAA measurements using HPLC/EC. The rate of 5-HT metabolism was calculated using the 5-HIAA/5-HT ratios. The ratios for the embryos treated with 0.1–0.3 mM ketamine (A) or treated with 2 mM ketamine, 2 mM ketamine + 0.5 mM ALCAR and 0.5 mM ALCAR alone (B) are presented. Values are expressed as mean ± SD. One-way ANOVA with post-hoc analyses were used to obtain statistical significance (*P < 0.05; **P < 0.001; ***P < 0.0001) of variation between the experimental groups as well as with the control.
3.3. ALCAR accelerates 5-HT metabolism alone and reverses ketamine-induced reduction in 5-HT level and its metabolism
In zebrafish embryos (52 hpf) treated for 24 h, ALCAR (0.5 mM) alone or in the presence of 2 mM ketamine did not significantly alter 5-HT levels compared to the controls (Fig. 2A), suggesting that ALCAR can reverse ketamine-induced reduction in 5-HT. ALCAR itself did not alter 5-HIAA levels and restored 5-HIAA levels in 2 mM ketamine-treated embryos to control levels (Fig. 2B), indicating that ketamine and ALCAR have opposing effects on 5-HT metabolism.
The 5-HIAA/5-HT ratio, used to determine the rate of 5-HT metabolism, showed that in embryos treated with 2 mM ketamine, 5-HT metabolism was completely inhibited (Fig. 3B). However, ALCAR (0.5 mM) significantly increased the rate of 5-HT metabolism to 5-HIAA and restored the turnover rate in ketamine-treated embryos to control levels (Fig. 3B). These results indicated that ALCAR and ketamine have opposing effects on the 5-HT system in zebrafish embryos.
3.4. Ketamine and ALCAR differentially modulate 5-HT neurons in the brain
In order to determine whether exposure to ketamine and ALCAR alone or in combination produces changes in the serotonergic (5-HT) neurons in the developing zebrafish brain, control and treated embryos were labeled with an antibody that recognizes serotonin (5-HT). Embryos (n = 15/treatment group) were treated at 52 hpf for 24 h. At 76 hpf, the embryos were processed for whole mount immunohistochemistry using a serotonin-specific polyclonal antibody. Immunofluorescent images (both ventral and dorsal views of the head region) clearly showed serotonin-positive neurons in the brains (Fig. 4). Ketamine at 0.1–0.3 mM doses showed immunostaining patterns similar to the control (data not shown). However, compared to the control group (Fig. 4A,E), embryos treated with 2 mM ketamine showed reduced area of serotonin labeling although the fluorescence intensity of the labeled neurons was not different (Fig. 4B,F). ALCAR in the presence of 2 mM ketamine also reduced the area and intensity of serotonin labeling when viewed ventrally (Fig. 4C) compared to the control but showed increased area of positive staining dorsally (Fig. 4G). It also resulted in axonal branching (Fig. 4G) not detected in the ketamine-treated embryos (Fig. 4B,F). These results indicated that ALCAR prevented ketamine-induced reduction in the serotonin-positive areas in the brain. In ventral views, the brains of embryos treated with ALCAR alone did not show any serotonin-positive neurons (Fig. 4D) at the same location as in the other groups (Fig. 4A–C), but there were serotonin-positive neurons with their extensions and sprouts spread out to the forebrain and midbrain areas. As observed in dorsal views, however, there were increased serotonin-positive cells than any other group including the control (Fig. 4E–G) residing in a broad range of brain areas and the axonal branching was extensive (Fig. 4H) than in any other groups. These studies suggest that the neuronal progenitor proliferation, migration, maturation and axonogenesis may have been differentially affected by ketamine and ALCAR.
Fig. 4.
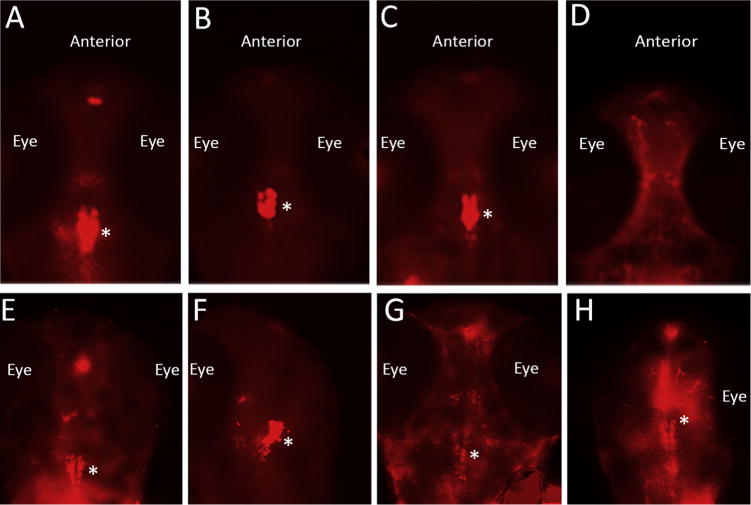
Effects of 2 mM and ALCAR on the serotonergic neurons in the zebrafish embryo brain.
Embryos at 52 hpf were treated with 2 mM ketamine, 2 mM ketamine + 0.5 mM ALCAR and 0.5 mM ALCAR alone. Twenty four hours post exposure, 76 hpf embryos were processed for whole mount immunohistochemistry using an antibody against serotonin (5-HT). Ventral views (upper panel) and dorsal views (lower panel) of the head region (anterior side up) show serotonergic neurons in the brain (major population marked by *) and their axonal branching, if any, in the Control (A, E), and the experimental groups, 2 mM ketamine (B, F), 2 mM ketamine + 0.5 mM ALCAR (C, G), 0.5 mM ALCAR (D, H). The location of the eye is indicated. The areas of positive labeling and fluorescence intensity indicate variations between the control and the treated embryos.
4. Discussion
Several studies have documented ketamine’s toxic effects on the zebrafish embryos and larvae [11,12,18,19,24,35]. Previously, we have shown [44] that the amount of ketamine accumulation (~8 μM) in the zebrafish embryos/larvae treated with 2 mM is closer to the lower range of human anesthetic range (plasma concentration ~10–100 μM) [13]. Here, we explored whether both the 2 mM dose as well as lower doses (0.1–0.3 mM) altered 5-HT system in zebrafish embryos. Low dose of ketamine can act as antidepressants in patients suffering from depressive disorder [29,38]. The role of the neurotransmitter, 5-HT, in the mammalian nervous system is well established [31] although over 90% of the 5-HT in the human body is produced outside of the central nervous system, especially in the intestine [9]. Our studies using an antibody against serotonin identified serotonergic neurons in the zebrafish embryo brains, but the whole embryo immunostaining did not specifically immunostain any intestinal regions (data not shown).
There are conflicting reports on the effect of ketamine on 5-HT turnover. In rodents, both decreased [42,51] and increased [21,45] 5-HT turnover following ketamine administration have been reported. The reason behind these variable results has been attributed to the fact that in most of these studies, only one dose of ketamine was used or the duration of exposure varied. In vivo studies on rats showed that ketamine decreased brain 5-HIAA in 30 min [27] and 2 h [51]. We evaluated the effect of ketamine on 5-HT content and turnover through measurements of whole body 5-HT and 5-HIAA levels since dissecting the head from the rest of the body of the larva for these measurements is not feasible. Our results showed no effect of ketamine at 0.1–0.3 mM doses on the 5-HT and 5-HIAA levels in the zebrafish embryos. In addition, 5-HIAA/5-HT ratios indicated that all low doses (0.1, 0.2, 0.3 mM) of ketamine did not affect 5-HT metabolism. However, at 2 mM (an internal exposure comparable to human anesthetic plasma levels), ketamine significantly reduced 5-HT level, and abolished its metabolism to 5-HIAA, an effect similar to what is seen in vivo in rats that exhibit reduced 5-HT content and metabolism upon ketamine exposure [27]. The finding that ketamine could inhibit 5-HT turnover in the zebrafish embryos may be of relevance since antidepressant-like properties of ketamine are observed in patients [6,29,38].
We have previously demonstrated that ALCAR reverses ketamine’s neurotoxic and cardiotoxic effects on zebrafish embryos [11,19]. ALCAR doses lower than 0.5 mM is ineffective against ketamine-induced toxicities in the zebrafish embryos (our unpublished data). In the mice brain, chronic ALCAR supplementation increased the levels of serotonin in the cortex, consistent with ALCAR’s potential efficacy for depressive symptoms [40]. In rats, a 7-day administration of ALCAR increased serotonin output in the nucleus accumbens shell [43]. In the present study, we investigated whether ALCAR altered ketamine’s effects on the 5-HT system in the zebrafish embryos. The results show that ALCAR reversed ketamine-induced decrease in 5-HT level and turnover. In our earlier studies, we reported that ketamine (2 mM) attenuated MAPK/ERK activity in the zebrafish embryos and ALCAR reversed this effect [19,44]. Moreover, it was shown in PC12 cells that 5-HT induced MAPK/ERK activity [23]. It would be interesting to examine whether ketamine-induced reduction in MAPK/ERK activity in the zebrafish embryos [19,44] is linked to the reduction in 5-HT levels.
In rats, it has been reported that ALCAR increased the level of 5-HIAA and acute ALCAR pre-treatment before 3,4-methylenedioxymethamphetamine (MDMA, ecstasy) injection prevented loss of 5-HT [2]. Moreover, chronic ALCAR treatment increased 5-HT level in the cerebral cortex and reduced 5-HT turnover [40]. In our studies on the zebrafish embryos, ALCAR enhanced 5-HT turnover and prevented loss of 5-HT and 5-HIAA observed in 2 mM ketamine-treated embryos. ALCAR’s effects on neurotransmitter (monoamine) regulations, especially serotonergic activities, are considered to have antidepressant actions in rodents [2,25,40,47,48]. Here, we show that ALCAR accelerated 5-HT metabolism. More importantly, it restored 5-HT level to control levels in ketamine-treated embryos suggesting a beneficial effect on the serotonergic system. Taken together, the data indicate the overall effects of ketamine and ALCAR on the serotonergic system and are comparable to mammalian and human data showing the modulation of the 5-HT system by ketamine [46,49,50] and ALCAR [2,25,40,47,48].
In summary, these studies show that zebrafish embryo model can help unveil the mode of action of drugs affecting the nervous system disorders involving the 5-HT system. Since we measured 5-HT and 5-HIAA levels in the whole embryos, further studies are warranted to determine whether ketamine and ALCAR alter the expression of TH and MAO enzymes at the transcriptional and translational levels, especially in the brain of the zebrafish embryos, in order to correlate these changes to brain disorders. It is also important to perform follow-up studies that would define a dose of ketamine (possibly between 0.3 mM and 2 mM), which can inhibit 5-HT metabolism but not synthesis.
5. Disclaimer
This document has been reviewed in accordance with United States Food and Drug Administration (FDA) policy and approved for publication. Approval does not signify that the contents necessarily reflect the position or opinions of the FDA, nor does mention of trade names or commercial products constitute endorsement or recommendation for use. The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the FDA.
HIGHLIGHTS.
Low doses of ketamine do not alter 5-HT and 5-HIAA levels in zebrafish embryos.
Anesthetic dose of ketamine reduces 5-HT levels and abolishes its metabolism to 5-HIAA.
ALCAR reverses the effects of ketamine on the 5-HT level and metabolism.
Anesthetic dose of ketamine reduces serotonin-positive cells in the brain.
Ketamine and ALCAR are being tested as anti-depressants, these results indicate their possible modes of action on the 5-HT system.
References
- 1.Ali SF, David SN, Newport GD. Age-related susceptibility to MPTP-induced neurotoxicity in mice. Neurotoxicology. 1993;14:29–34. [PubMed] [Google Scholar]
- 2.Alves E, Binienda Z, Carvalho F, Alves CJ, Fernandes E, de Lourdes Bastos M, Tavares MA, Summavielle T. Acetyl-L-carnitine provides effective in vivo neuroprotection over 3,4-methylenedioximethamphetamine-induced mitochondrial neurotoxicity in the adolescent rat brain. Neuroscience. 2009;158:514–523. doi: 10.1016/j.neuroscience.2008.10.041. [DOI] [PubMed] [Google Scholar]
- 3.Aroni F, Iacovidou N, Dontas I, Pourzitaki C, Xanthos T. Pharmacological aspects and potential new clinical applications of ketamine: reevaluation of an old drug. J Clin Pharmacol. 2009;49:957–964. doi: 10.1177/0091270009337941. [DOI] [PubMed] [Google Scholar]
- 4.Azzaro AJ, Smith DJ. The inhibitory action of ketamine HC1 on [3H]5-hydroxytryptamine accumulation by rat brain synaptosomal-rich fractions: comparison with [3H]catecholamine and [3H]gamma-aminobutyric acid uptake. Neuropharmacology. 1977;16:349–356. doi: 10.1016/0028-3908(77)90072-7. [DOI] [PubMed] [Google Scholar]
- 5.Beal MF. Bioenergetic approaches for neuroprotection in Parkinson’s disease. Ann Neurol. 2003;53(Suppl 3):S39–S47. doi: 10.1002/ana.10479. Discussion S47–S38. [DOI] [PubMed] [Google Scholar]
- 6.Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS, Krystal JH. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry. 2000;47:351–354. doi: 10.1016/s0006-3223(99)00230-9. [DOI] [PubMed] [Google Scholar]
- 7.Bieber LL. Carnitine. Annu Rev Biochem. 1988;57:261–283. doi: 10.1146/annurev.bi.57.070188.001401. [DOI] [PubMed] [Google Scholar]
- 8.Briggs JP. The zebrafish: a new model organism for integrative physiology. Am J Physiol Regul Integr Comp Physiol. 2002;282:R3–R9. doi: 10.1152/ajpregu.00589.2001. [DOI] [PubMed] [Google Scholar]
- 9.Buffa R, Capella C, Fontana P, Usellini L, Solcia E. Types of endocrine cells in the human colon and rectum. Cell Tissue Res. 1978;192:227–240. doi: 10.1007/BF00220741. [DOI] [PubMed] [Google Scholar]
- 10.Corssen G, Miyasaka M, Domino EF. Changing concepts in pain control during surgery: dissociative anesthesia with CI-581. A progress report. Anesth Analg. 1968;47:746–759. [PubMed] [Google Scholar]
- 11.Cuevas E, Trickler WJ, Guo X, Ali SF, Paule MG, Kanungo J. Acetyl L-carnitine protects motor neurons and Rohon–Beard sensory neurons against ketamine-induced neurotoxicity in zebrafish embryos. Neurotoxicol Teratol. 2013;39:69–76. doi: 10.1016/j.ntt.2013.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Felix LM, Antunes LM, Coimbra AM. Ketamine NMDA receptor-independent toxicity during zebrafish (Danio rerio) embryonic development. Neurotoxicol Teratol. 2014;41:27–34. doi: 10.1016/j.ntt.2013.11.005. [DOI] [PubMed] [Google Scholar]
- 13.Geisslinger G, Hering W, Thomann P, Knoll R, Kamp HD, Brune K. Pharmacokinetics and pharmacodynamics of ketamine enantiomers in surgical patients using a stereoselective analytical method. Br J Anaesth. 1993;70:666–671. doi: 10.1093/bja/70.6.666. [DOI] [PubMed] [Google Scholar]
- 14.Haberny KA, Paule MG, Scallet AC, Sistare FD, Lester DS, Hanig JP, Slikker W., Jr Ontogeny of the N-methyl-D-aspartate (NMDA) receptor system and susceptibility to neurotoxicity. Toxicol Sci. 2002;68:9–17. doi: 10.1093/toxsci/68.1.9. [DOI] [PubMed] [Google Scholar]
- 15.Hagen TM, Moreau R, Suh JH, Visioli F. Mitochondrial decay in the aging rat heart: evidence for improvement by dietary supplementation with acetyl-L-carnitine and/or lipoic acid. Ann N Y Acad Sci. 2002;959:491–507. doi: 10.1111/j.1749-6632.2002.tb02119.x. [DOI] [PubMed] [Google Scholar]
- 16.Ikonomidou C, Bosch F, Miksa M, Bittigau P, Vockler J, Dikranian K, Tenkova TI, Stefovska V, Turski L, Olney JW. Science. Vol. 283. New York, N.Y: 1999. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain; pp. 70–74. [DOI] [PubMed] [Google Scholar]
- 17.Imam SZ, el-Yazal J, Newport GD, Itzhak Y, Cadet JL, Slikker W, Jr, Ali SF. Methamphetamine-induced dopaminergic neurotoxicity: role of peroxynitrite and neuroprotective role of antioxidants and peroxynitrite decomposition catalysts. Ann N Y Acad Sci. 2001;939:366–380. doi: 10.1111/j.1749-6632.2001.tb03646.x. [DOI] [PubMed] [Google Scholar]
- 18.Kanungo J, Cuevas E, Ali SF, Paule MG. Ketamine induces motor neuron toxicity and alters neurogenic and proneural gene expression in zebrafish. J Appl Toxicol. 2013;33:410–417. doi: 10.1002/jat.1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kanungo J, Cuevas E, Ali SF, Paule MG. L-Carnitine rescues ketamine-induced attenuated heart rate and MAPK (ERK) activity in zebrafish embryos. Reprod Toxicol. 2012;33:205–212. doi: 10.1016/j.reprotox.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kanungo J, Cuevas E, Ali SF, Paule MG. Zebrafish model in drug safety assessment. Curr Pharm Des. 2014;20:5416–5429. doi: 10.2174/1381612820666140205145658. [DOI] [PubMed] [Google Scholar]
- 21.Kari HP, Davidson PP, Kohl HH, Kochhar MM. Effects of ketamine on brain monoamine levels in rats. Res Commun Chem Pathol Pharmacol. 1978;20:475–488. [PubMed] [Google Scholar]
- 22.Kohrs R, Durieux ME. Ketamine: teaching an old drug new tricks. Anesth Analg. 1998;87:1186–1193. doi: 10.1097/00000539-199811000-00039. [DOI] [PubMed] [Google Scholar]
- 23.Koizumi K, Nakajima H. Serotonin induces the migration of PC12 cells via the serotonin receptor 6/cAMP/ERK pathway. Biomed Reports. 2014;2:29–33. doi: 10.3892/br.2013.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lantz-McPeak S, Guo X, Cuevas E, Dumas M, Newport GD, Ali SF, Paule MG, Kanungo J. Developmental toxicity assay using high content screening of zebrafish embryos. J Appl Toxicol. 2015;35:261–272. doi: 10.1002/jat.3029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Levine J, Kaplan Z, Pettegrew JW, McClure RJ, Gershon S, Buriakovsky I, Cohen H. Effect of intraperitoneal acetyl-L-carnitine (ALCAR) on anxiety-like behaviours in rats. Int J Neuropsychopharmacol. 2005;8:65–74. doi: 10.1017/S1461145704004596. [DOI] [PubMed] [Google Scholar]
- 26.Liu F, Patterson TA, Sadovova N, Zhang X, Liu S, Zou X, Hanig JP, Paule MG, Slikker W, Jr, Wang C. Ketamine-induced neuronal damage and altered N-methyl-D-aspartate receptor function in rat primary forebrain culture. Toxicol Sci. 2013;131:548–557. doi: 10.1093/toxsci/kfs296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Martin LL, Bouchal RL, Smith DJ. Ketamine inhibits serotonin uptake in vivo. Neuropharmacology. 1982;21:113–118. doi: 10.1016/0028-3908(82)90149-6. [DOI] [PubMed] [Google Scholar]
- 28.Martin LL, Smith DJ. Ketamine inhibits serotonin synthesis and metabolism in vivo. Neuropharmacology. 1982;21:119–125. doi: 10.1016/0028-3908(82)90150-2. [DOI] [PubMed] [Google Scholar]
- 29.Miller JT. Two cellular hypotheses explaining ketamine’s antidepressant actions: direct inhibition and disinhibition. Neuropharmacology. 2015 doi: 10.1016/j.neuropharm.2015.07.028. [DOI] [PubMed] [Google Scholar]
- 30.Niesters M, Khalili-Mahani N, Martini C, Aarts L, van Gerven J, van Buchem MA, Dahan A, Rombouts S. Effect of subanesthetic ketamine on intrinsic functional brain connectivity: a placebo-controlled functional magnetic resonance imaging study in healthy male volunteers. Anesthesiology. 2012;117:868–877. doi: 10.1097/ALN.0b013e31826a0db3. [DOI] [PubMed] [Google Scholar]
- 31.Olivier B. Serotonin: a never-ending story. Eur J Pharmacol. 2015;753:2–18. doi: 10.1016/j.ejphar.2014.10.031. [DOI] [PubMed] [Google Scholar]
- 32.Orth M, Schapira AH. Mitochondrial involvement in Parkinson’s disease. Neurochem Int. 2002;40:533–541. doi: 10.1016/s0197-0186(01)00124-3. [DOI] [PubMed] [Google Scholar]
- 33.Palacios HH, Yendluri BB, Parvathaneni K, Shadlinski VB, Obrenovich ME, Leszek J, Gokhman D, Gasiorowski K, Bragin V, Aliev G. Mitochondrion-specific antioxidants as drug treatments for alzheimer disease. CNS Neurol Disord Drug Targets. 2011;10:149–162. doi: 10.2174/187152711794480474. [DOI] [PubMed] [Google Scholar]
- 34.Parng C, Seng WL, Semino C, McGrath P. Zebrafish: a preclinical model for drug screening. Assay Drug Dev Technol. 2002;1:41–48. doi: 10.1089/154065802761001293. [DOI] [PubMed] [Google Scholar]
- 35.Riehl R, Kyzar E, Allain A, Green J, Hook M, Monnig L, Rhymes K, Roth A, Pham M, Razavi R, Dileo J, Gaikwad S, Hart P, Kalueff AV. Behavioral and physiological effects of acute ketamine exposure in adult zebrafish. Neurotoxicol Teratol. 2011;33:658–667. doi: 10.1016/j.ntt.2011.05.011. [DOI] [PubMed] [Google Scholar]
- 36.Rowland LM. Subanesthetic ketamine: how it alters physiology and behavior in humans. Aviat Space Env Med. 2005;76:C52–C58. [PubMed] [Google Scholar]
- 37.Sallinen V, Sundvik M, Reenila I, Peitsaro N, Khrustalyov D, Anichtchik O, Toleikyte G, Kaslin J, Panula P. Hyperserotonergic phenotype after monoamine oxidase inhibition in larval zebrafish. J Neurochem. 2009;109:403–415. doi: 10.1111/j.1471-4159.2009.05986.x. [DOI] [PubMed] [Google Scholar]
- 38.Serafini G, Howland RH, Rovedi F, Girardi P, Amore M. The role of ketamine in treatment-resistant depression: a systematic review. Curr Neuropharmacol. 2014;12:444–461. doi: 10.2174/1570159X12666140619204251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Slikker W, Jr, Zou X, Hotchkiss CE, Divine RL, Sadovova N, Twaddle NC, Doerge DR, Scallet AC, Patterson TA, Hanig JP, Paule MG, Wang C. Ketamine-induced neuronal cell death in the perinatal rhesus monkey. Toxicol Sci. 2007;98:145–158. doi: 10.1093/toxsci/kfm084. [DOI] [PubMed] [Google Scholar]
- 40.Smeland OB, Meisingset TW, Borges K, Sonnewald U. Chronic acetyl-L-carnitine alters brain energy metabolism and increases noradrenaline and serotonin content in healthy mice. Neurochem Int. 2012;61:100–107. doi: 10.1016/j.neuint.2012.04.008. [DOI] [PubMed] [Google Scholar]
- 41.Stumpf DA, Parker WD, Jr, Angelini C. Carnitine deficiency, organic acidemias, and Reye’s syndrome. Neurology. 1985;35:1041–1045. doi: 10.1212/wnl.35.7.1041. [DOI] [PubMed] [Google Scholar]
- 42.Sung YF, Frederickson EL, Holtzman SG. Effects of intravenous anesthetics on brain monoamines in the rat. Anesthesiology. 1973;39:478–487. doi: 10.1097/00000542-197311000-00004. [DOI] [PubMed] [Google Scholar]
- 43.Tolu P, Masi F, Leggio B, Scheggi S, Tagliamonte A, De Montis MG, Gambarana C. Effects of long-term acetyl-L-carnitine administration in rats: I. Increased dopamine output in mesocorticolimbic areas and protection toward acute stress exposure. Neuropsychopharmacology. 2002;27:410–420. doi: 10.1016/S0893-133X(02)00306-8. [DOI] [PubMed] [Google Scholar]
- 44.Trickler WJ, Guo X, Cuevas E, Ali SF, Paule MG, Kanungo J. Ketamine attenuates cytochrome p450 aromatase gene expression and estradiol-17beta levels in zebrafish early life stages. J Appl Toxicol. 2014;34:480–488. doi: 10.1002/jat.2888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Vargiu L, Stefanini E, Musinu C, Saba G. Possible role of brain serotonin in the central effects of ketamine. Neuropharmacology. 1978;17:405–408. doi: 10.1016/0028-3908(78)90014-x. [DOI] [PubMed] [Google Scholar]
- 46.Vinckier F, Gaillard R, Palminteri S, Rigoux L, Salvador A, Fornito A, Adapa R, Krebs MO, Pessiglione M, Fletcher PC. Confidence and psychosis: a neuro-computational account of contingency learning disruption by NMDA blockade. Mol Psychiatry. 2015 doi: 10.1038/mp.2015.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang SM, Han C, Lee SJ, Patkar AA, Masand PS, Pae CU. A review of current evidence for acetyl-L-carnitine in the treatment of depression. J Psychiatr Res. 2014;53:30–37. doi: 10.1016/j.jpsychires.2014.02.005. [DOI] [PubMed] [Google Scholar]
- 48.Wang W, Lu Y, Xue Z, Li C, Wang C, Zhao X, Zhang J, Wei X, Chen X, Cui W, Wang Q, Zhou W. Rapid-acting antidepressant-like effects of acetyl-L-carnitine mediated by PI3K/AKT/BDNF/VGF signaling pathway in mice. Neuroscience. 2015;285:281–291. doi: 10.1016/j.neuroscience.2014.11.025. [DOI] [PubMed] [Google Scholar]
- 49.Yamamoto S, Ohba H, Nishiyama S, Harada N, Kakiuchi T, Tsukada H, Domino EF. Subanesthetic doses of ketamine transiently decrease serotonin transporter activity: a PET study in conscious monkeys. Neuropsychopharmacology. 2013;38:2666–2674. doi: 10.1038/npp.2013.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yamanaka H, Yokoyama C, Mizuma H, Kurai S, Finnema SJ, Halldin C, Doi H, Onoe H. A possible mechanism of the nucleus accumbens and ventral pallidum 5-HT1B receptors underlying the antidepressant action of ketamine: a PET study with macaques. Transl Psychiatry. 2014;4:e342. doi: 10.1038/tp.2013.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ylitalo P, Saarnivaara L, Ahtee L. Effect of ketamine anaesthesia on the content of monoamines and their metabolites in the rat brain. Acta Anaesthesiol Scand. 1976;20:216–220. doi: 10.1111/j.1399-6576.1976.tb05032.x. [DOI] [PubMed] [Google Scholar]
- 52.Zou X, Sadovova N, Patterson TA, Divine RL, Hotchkiss CE, Ali SF, Hanig JP, Paule MG, Slikker W, Jr, Wang C. The effects of L-carnitine on the combination of, inhalation anesthetic-induced developmental, neuronal apoptosis in the rat frontal cortex. Neuroscience. 2008;151:1053–1065. doi: 10.1016/j.neuroscience.2007.12.013. [DOI] [PubMed] [Google Scholar]