

Abstract
Serine carboxypeptidase (SCP) is one of the largest groups of enzymes catalyzing proteolysis for functional protein maturation. To date, little is known about the function of SCPs in rice. In this study, we present a comprehensive analysis of the gene structure and expression profile of 59 rice SCPs. SCP46 is dominantly expressed in developing seeds, particularly in embryo, endosperm and aleurone layers, and could be induced by ABA. Functional characterization revealed that knock-down of SCP46 resulted in smaller grain size and enhanced seed germination. Furthermore, scp46 seed germination became less sensitive to the ABA inhibition than the Wild-type did; suggesting SCP46 is involved in ABA signaling. As indicated by RNA-seq and qRT-PCR analysis, numerous grain filling and seed dormancy related genes, such as SP, VP1 and AGPs were down-regulated in scp46. Yeast-two-hybrid assay also showed that SCP46 interacts with another ABA-inducible protein DI19-1. Taken together, we suggested that SCP46 is a master regulator of grain filling and seed germination, possibly via participating in the ABA signaling. The results of this study shed novel light into the roles of SCPs in rice.
Introduction
Proteolysis catalyzed by peptidase is an important post-translational modification for functional protein maturation. Based on the location of the cleavage site within the substrate, peptidases are divided into endopeptidase and exopeptidase. Endopeptidase splits internal peptide bonds within the protein, whereas exopeptidase can only detach the terminal amino acids of the protein chain. According to the terminus position of the peptide studied, an exopeptidase can be further classified into either an aminopeptidase or carboxypeptidase. Aminopeptidases usually cleave one or two residues each time from the N-terminus. On the contrary, carboxypeptidases cleave peptides from the C-terminus at a speed of one residue each time. Serine carboxypeptidase (SCP) is a type of acid carboxypeptidase whose optimum hydrolysis activity is achieved under acidic pH range. Hayashi et al (1973) named this group of enzymes as “serine carboxypeptidase” because they could be inhibited by a specific serine protease inhibitor DFP [1]. In protein structure, SCPs have a conserved Ser-Asp-His catalytic triad. Though these residues are dispersed in the primary amino acid sequence, the triad residues are physically aggregated in the protein tertiary structure to create a nucleophilic serine for substrate hydrolysis [2]. SCPs also have another conserved oxyanion hole motif, whose function is to stabilize the enzyme-substrate interaction during the substrate protein hydrolysis. Given the special enzymatic features in protein hydrolysis, SCPs have been widely used as molecular biology tool enzymes for determination, exchanging and synthesis of the C-terminal sequence in peptides [3].
Recently, increasing evidences have shown that SCPs play crucial roles in protein transport, targeting and processing. In humans, defects in SCP are associated with severe diseases ranging from obesity to epilepsy to neurodegeneration [4,5]. Besides humans, SCPs and SCP-like (SCPL) genes are widely distributed in animals, bacterium, fungi as well as higher plants [6–13]. A genome survey conducted by Tripathi et al (2006) revealed 54 and 66 SCPL genes in Arabidopsis and rice respectively [11]. By using the same hidden Markov models but different criteria, Feng et al (2009) identified 54 and 71 SCPL genes in Arabidopsis and rice. A large number of the Arabidopsis SCPLs showed rich alternative splicing models, while this phenomenon barely occurred in rice SCPLs [2]. The Arabidopsis SCP members displayed divergent spatial-expression patterns though they were phylogenetically closely related, suggesting versatile roles of SCPs in plant development [7]. Indeed, when ECS1 (Extra Carpels and Seeds), encoding a serine carboxypeptidase, was ectopically expressed in Arabidopsis, the carpel and silique seed number were significantly increased. Genetic analysis of ecs1 and bri1-5 (BRassinosteroid Insensitive 1–5) implied an important role of ECS1 in BR (brassinosteroid) signaling [14]. Additionally, another Arabidopsis SCP protein BRS1 was found to be involved in early event of BR signaling [15]. In germinating wheat grains, carboxypeptidase III (CPIII) is highly accumulated in the scutellum at 2–3 days after imbibition, indicating a role of CPIII in mobilization of starchy-endosperm proteins. Moreover, in situ nuclear DNA fragmentation was detected in cells with high CPIII expression, thus the authors proposed that CPIII may also be involved in programmed cell death during the development of the vascular tissue in wheat [16]. Unlike its wheat ortholog CPIII, PsCP from pea (Pisum sativum L.) does not participate in the mobilization of storage nutrients, but a predominant expression pattern in seed and selective hormone-induced, in particular GA-induced, expression pattern in seedlings still hinted of a very important function of PsCP in reproductive and vegetative developing tissues in dicotyledoneous plants [17]. Bienert et al (2012) isolated two extracellular serine carboxypeptidase III genes from tobacco (Nicotiana tabacum), NtSCP1 and NtSCP2. Both of these genes displayed a constitutive expression among all the checked tissues, but only purified His-tagged NtSCP1 had carboxypeptidase activity in vitro. Functional characterization revealed that NtSCP1 regulates cell elongation in flower and hypocotyls. Unfortunately, this study did not further explore the relationship between NtSCP1 and phytohormone GA, as there were already some cases showing SCPs like CPIII and PsCP are highly associated with GA controlled cell elongation [18]. In addition to the peptidase function, the emerging roles of SCPLs as acyltransferases were gradually recognized [12,19,20]. SCPL enzymes were shown to catalyze the region specific formation of diacylglucose (1,2-di-O-acylglucose) using the glucose esters 1-O-acylglucose as acyl donors [21]. So far, several publications have reported that SCPL acyltransferases are involved in secondary metabolism, herbicide detoxication, development, biotic and abiotic stress responses in a broad range of plant species, which greatly enhanced understanding of SCPs and SCPLs [7,12,22–26].
Rice (Oryza sativa L.) is one of the major crops in the world, feeding over half of the global population. Besides its economical importance, rice is also an ideal model plant for gene functional studies due to its released genome sequence, ample genetic resources, mature transformation system and co-linearity with other grasses [27]. In the past few decades, over 2000 rice genes have been cloned and functionally characterized, representing a tremendous progress in plant molecular biology research (http://www.ricedata.cn/gene/) [28]. Regarding the SCPs in rice, despite scientific exploration of the spatial- and hormone-induced expression pattern of the family members in the 1990s, the knowledge on rice SCPs remains rather poor with only a few exceptions over the last 20 years [29]. For example, OsBISCPL1 encoding a putative SCPL was found to positively regulate rice resistance against bacterial pathogens by elevating defense genes such as PR1, PR2, PR5 and PDF1.2. There was evidence showing that OsBISCPL1 may be related to the oxidative stress responses as well [30]. Li et al (2011) cloned another rice SCP gene GS5 which is a major QTL site determining rice grain size. [31]. In this study, we systematically analyzed the gene structure, tissue expression profile of rice SCP family. Employing a reverse genetic strategy, we revealed that SCP46, a seed-dominantly expressed gene, controls rice grain filling and seed germination by involvement in the ABA signaling. The results of this research shed novel light into the function of SCPs in rice.
Results
Rice SCP genes exhibit diverse expression patterns
A genome survey of the rice genome annotation project (RGAP) database (http://rice.plantbiology.msu.edu/) [32] resulted in the identification of 59 SCP genes in rice. By following the nomenclature of RGAP, all these SCPs were named from OsSCP1 to 66 with some numbers skipped due to the updated genome annotation. Table 1 presents the comprehensive information of this rice gene family. The 59 SCPs are distributed in different loci of the 12 rice chromosomes in the form of a single gene or gene cluster. The gene length of SCPs varies from 600bp to 33 kb, while the protein lengths are in the range of 300 to 500 amino acids with a few extreme exceptions (Table 1). Additionally, we downloaded 40 Maize SCPs from the MaizeGDB (http://www.maizegdb.org/) [33]. To investigate the evolutionary relationship of rice and maize SCP genes, the amino acid sequences of all the 99 genes were used as an input for the construction of the SCP phylogenic map by using ClustalW (MEGA7.0). The results suggested that the SCPs are largely diversified (Fig 1). The maize 2G123815 was the closest ortholog to GS5, indicating that this gene could also be functionally a seed development regulator. On the other hand, we found that some clades barely contain any maize SCPs, implying that this subgroup is highly specified in rice. CREP (Collection of Rice Expression Profiles) (http://crep.ncpgr.cn/) collected the expression profiles of rice genes in 39 tissues from 2 elite varieties [34]. To gain an overall view of the SCP expression profiles, we downloaded the expression data in 8 typical tissues and stages from CREP including callus, leaf, sheath, root, panicle, 7 DAP endosperm, 14 DAP endosperm and 21 DAP endosperm, and displayed the profile as a heat map in Fig 2A. As a result, SCP genes showed divergent expression patterns. Some members such as SCP11 and 26 were constitutively expressed in all the tissues. However, we did find several SCPs showed highly tissue specific expression patterns. For example, OsSCP3 was dominantly expressed in leaf, while OsSCP10, 16 and 24 were specifically expressed in panicles. As the research focus in our lab is rice seed development, we also attempted to find out the seed-specific or -dominant SCP genes. It turned out that the expression level in endosperms was at least 4 folds higher than in other tissues for SCP6, 46 and 56, which suggested a seed-dominant expression pattern and intrigued us to further explore their biological function in rice seed development. The divergent expression profiles of SCPs indicated versatile roles of this gene family in rice growth and development.
Table 1. Comprehensive information of SCPs in rice.
| Gene | Locus ID | Gene length (bp) | CDS length (bp) | Protein length (AA) | Domain length | Molecular weight (KDa) | PI | Exon | Intron |
|---|---|---|---|---|---|---|---|---|---|
| OsSCP1 | LOC_Os01g06490 | 2734 | 1416 | 471 | 412 | 52.53 | 5.2508 | 9 | 8 |
| OsSCP2 | LOC_Os01g11670 | 1686 | 1290 | 429 | 392 | 46.26 | 7.0798 | 2 | 1 |
| OsSCP3 | LOC_Os01g22980 | 4263 | 1442 | 480 | 427 | 54.32 | 6.6402 | 10 | 9 |
| OsSCP4 | LOC_Os01g43890 | 2066 | 1359 | 452 | 394 | 48.08 | 7.148 | 1 | 0 |
| OsSCP5 | LOC_Os01g61690 | 3631 | 1365 | 454 | 415 | 50.54 | 6.9461 | 9 | 8 |
| OsSCP6 | LOC_Os02g02320 | 4869 | 1503 | 500 | 399 | 55.45 | 6.0623 | 9 | 8 |
| OsSCP7 | LOC_Os02g26480 | 9003 | 1512 | 503 | 429 | 56.85 | 5.6285 | 14 | 13 |
| OsSCP8 | LOC_Os02g42310 | 7255 | 1458 | 485 | 427 | 53.92 | 6.3211 | 10 | 9 |
| OsSCP9 | LOC_Os02g46260 | 5212 | 1443 | 480 | 428 | 53.62 | 5.7362 | 14 | 13 |
| OsSCP10 | LOC_Os02g55130 | 7227 | 1422 | 473 | 427 | 52.34 | 9.3639 | 9 | 8 |
| OsSCP11 | LOC_Os03g09190 | 3522 | 1398 | 465 | 410 | 51.78 | 6.6214 | 6 | 5 |
| OsSCP12 | LOC_Os03g26920 | 2377 | 1512 | 503 | 425 | 55.96 | 4.7272 | 8 | 7 |
| OsSCP13 | LOC_Os03g26930 | 3430 | 1437 | 478 | 409 | 53.52 | 6.5811 | 8 | 7 |
| OsSCP14 | LOC_Os03g27480 | 3912 | 1413 | 470 | 414 | 50.98 | 7.5554 | 12 | 11 |
| OsSCP15 | LOC_Os03g27510 | 4805 | 1014 | 337 | 259 | 36.82 | 9.4207 | 11 | 10 |
| OsSCP16 | LOC_Os03g27530 | 6926 | 1173 | 390 | 291 | 42.84 | 6.7185 | 11 | 10 |
| OsSCP17 | LOC_Os03g27550 | 626 | 237 | 78 | 44 | 8.53 | 9.7602 | 3 | 2 |
| OsSCP18 | LOC_Os03g27590 | 5445 | 1413 | 470 | 424 | 50.79 | 4.7504 | 12 | 11 |
| OsSCP19 | LOC_Os03g52040 | 5692 | 1452 | 483 | 431 | 53.88 | 7.3067 | 11 | 10 |
| OsSCP20 | LOC_Os03g52070 | 7317 | 1317 | 438 | 384 | 48.56 | 6.9292 | 11 | 10 |
| OsSCP21 | LOC_Os03g52080 | 4148 | 1203 | 400 | 138 | 44.81 | 7.1324 | 13 | 12 |
| OsSCP22 | LOC_Os04g09720 | 8641 | 1404 | 467 | 422 | 51.69 | 6.0767 | 14 | 13 |
| OsSCP23 | LOC_Os04g25560 | 6011 | 1515 | 504 | 458 | 56.11 | 6.4622 | 14 | 13 |
| OsSCP24 | LOC_Os04g32540 | 3666 | 1413 | 470 | 422 | 51.45 | 6.5161 | 8 | 7 |
| OsSCP25 | LOC_Os04g44410 | 4606 | 1407 | 468 | 417 | 52.02 | 9.0621 | 7 | 6 |
| GS5/OsSCP26 | LOC_Os05g06660 | 4560 | 1452 | 483 | 428 | 53.73 | 6.509 | 10 | 9 |
| OsSCP28 | LOC_Os05g18604 | 33199 | 1452 | 483 | 419 | 53.77 | 7.1201 | 10 | 9 |
| OsSCP29 | LOC_Os05g50570 | 1574 | 1341 | 446 | 395 | 46.97 | 7.6887 | 1 | 0 |
| OsSCP30 | LOC_Os05g50580 | 1938 | 1356 | 451 | 400 | 47.76 | 6.4078 | 1 | 0 |
| OsSCP31 | LOC_Os05g50600 | 1473 | 1329 | 442 | 391 | 47.37 | 7.0264 | 1 | 0 |
| OsSCP32 | LOC_Os06g08720 | 7467 | 1455 | 484 | 419 | 53.41 | 5.6715 | 10 | 9 |
| OsSCP33 | LOC_Os06g13410 | 647 | 576 | 191 | 63 | 20.11 | 8.8431 | 2 | 1 |
| OsSCP34 | LOC_Os06g13420 | 2193 | 876 | 291 | 41 | 31.37 | 10.6293 | 4 | 3 |
| OsSCP35 | LOC_Os06g32740 | 4493 | 3273 | 1090 | 62 | 125.08 | 6.2913 | 6 | 5 |
| OsSCP36 | LOC_Os06g32770 | 2318 | 1593 | 531 | 438 | 59.14 | 7.7865 | 2 | 1 |
| OsSCP37 | LOC_Os06g36570 | 756 | 756 | 252 | 148 | 27.34 | 8.8198 | 1 | 0 |
| OsSCP38 | LOC_Os06g51370 | 2152 | 1503 | 501 | 431 | 55.48 | 6.5071 | 3 | 2 |
| OsSCP39 | LOC_Os07g29620 | 3827 | 1575 | 525 | 400 | 57.35 | 5.2912 | 9 | 8 |
| OsSCP40 | LOC_Os07g46350 | 4983 | 1575 | 525 | 423 | 56.15 | 5.4391 | 7 | 6 |
| OsSCP41 | LOC_Os08g44640 | 5339 | 1407 | 469 | 423 | 51.46 | 6.3128 | 12 | 11 |
| OsSCP42 | LOC_Os09g28830 | 3725 | 1335 | 445 | 337 | 49.06 | 8.6758 | 8 | 7 |
| OsSCP43 | LOC_Os09g28840 | 4851 | 1809 | 603 | 261 | 66.78 | 6.2419 | 11 | 10 |
| OsSCP44 | LOC_Os10g01110 | 3115 | 1383 | 461 | 410 | 51.34 | 7.1007 | 8 | 7 |
| OsSCP46 | LOC_Os10g01134 | 6317 | 1419 | 473 | 416 | 52.13 | 5.3897 | 3 | 2 |
| OsSCP47 | LOC_Os10g39560 | 3212 | 1383 | 461 | 389 | 51.27 | 5.4377 | 10 | 9 |
| OsSCP48 | LOC_Os11g10750 | 4046 | 1398 | 466 | 421 | 51.71 | 6.5081 | 10 | 9 |
| OsSCP50 | LOC_Os11g24180 | 15020 | 1377 | 459 | 403 | 51.07 | 5.2605 | 12 | 11 |
| OsSCP51 | LOC_Os11g24290 | 4028 | 690 | 230 | 164 | 25.27 | 4.4332 | 7 | 6 |
| OsSCP52 | LOC_Os11g24320 | 3720 | 486 | 162 | 57 | 17.55 | 4.5268 | 5 | 4 |
| OsSCP53 | LOC_Os11g24340 | 4668 | 1188 | 396 | 333 | 44.72 | 7.6011 | 9 | 8 |
| OsSCP55 | LOC_Os11g24374 | 24718 | 1182 | 394 | 347 | 43.87 | 6.5019 | 12 | 11 |
| OsSCP56 | LOC_Os11g24510 | 8005 | 1440 | 480 | 429 | 53.36 | 5.8082 | 13 | 12 |
| OsSCP58 | LOC_Os11g27170 | 19261 | 1425 | 475 | 400 | 52.96 | 5.4265 | 12 | 11 |
| OsSCP60 | LOC_Os11g27264 | 11827 | 1452 | 484 | 430 | 53.86 | 5.7842 | 14 | 13 |
| OsSCP62 | LOC_Os11g27329 | 31581 | 1077 | 359 | 258 | 39.52 | 5.6281 | 14 | 13 |
| OsSCP63 | LOC_Os11g31980 | 3853 | 1419 | 473 | 410 | 52.61 | 6.5171 | 10 | 9 |
| OsSCP64 | LOC_Os11g42390 | 6132 | 1389 | 463 | 369 | 51.35 | 7.9475 | 8 | 7 |
| OsSCP65 | LOC_Os12g15470 | 5108 | 1566 | 522 | 447 | 56.77 | 7.7368 | 14 | 13 |
| OsSCP66 | LOC_Os12g39170 | 6120 | 1362 | 454 | 402 | 50.48 | 5.9073 | 10 | 9 |
Fig 1. Phylogenetic analysis of rice and maize SCPs.
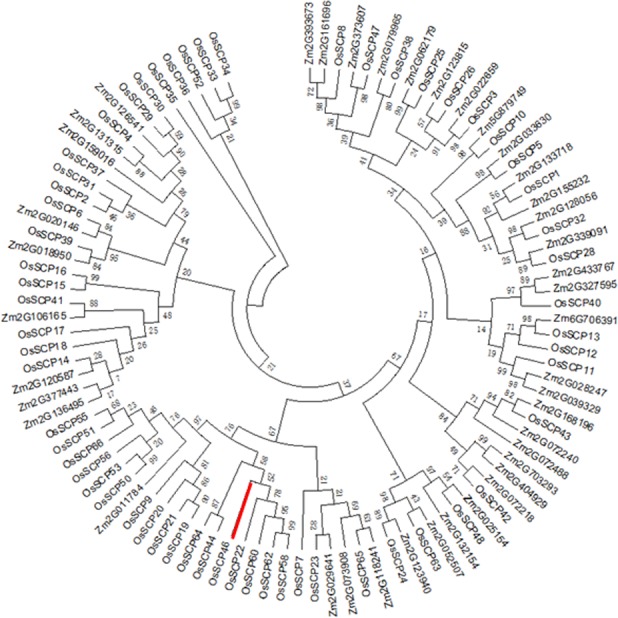
The branch length scale bar indicates the evolutionary distance. Red line corresponds to SCP46.
Fig 2. Expression analysis of SCPs in tissues and response to phytohormone treatments.

(A) Hierarchical cluster display of the expression profile of rice SCPs in callus, leaf, shealth, root, panicle, 7, 14 and 21 DAP seeds. Color bar on the top represents the log2 expression values. Blue, white and Purple indicate the low, medium and high expression values respectively. (B) Schematical representation of the gene structure of SCP46. (C) qRT-PCR analysis of SCP46 in various rice tissues. Different characters indicate a statistically significant difference at P<0.05. (D-F) qRT-PCR analysis of SCP46 in response to ABA (D), BR (E) and GA (F) treatment at 5 different time points. ** indicates P<0.01 by t-test in comparison with 0h. (G-J) mRNA in-situ hybridization assay of SCP46 in seeds at early (G), medium (H) and late stages (I); (J) shows the negative control by using sense probe for hybridization. Em: embryo; En: endosperm; PL: pericap layer; AL: aleuron layer; Bar = 1.5 mm.
SCP46 is dominantly expressed in embryo, endosperm and aleurone layer of developing seeds
A spatial- or temporal-expression pattern usually indicates the critical roles of the genes in the corresponding biological process. According to the microarray data from CREP, SCP46 is dominantly expressed in rice developing seeds. Due to the alternative splicing models, SCP46 is predicted to have 2 isoforms, both of which consist of 3 exons and 2 introns (Fig 2B). To validate the expression profile of SCP46, we performed quantitative RT-PCR to examine the transcriptional level of SCP46 in ten various rice tissues and stages. As shown in Fig 2B, the SCP46 mRNA abundances were extremely low in callus, leaf, root, stem, old panicles and unpollinated pistils, but dominantly expressed in developing seeds. In particular, we found that the SCP46 transcription level increased with the development of seeds, and reached a maximum level at 25 DAP, suggesting that SCP46 may have an important function in rice grain filling and seed maturation. In addition, we checked the transcription level of SCP46 in response to the application of 100μM ABA, 100μM GA and 10μM BR at 0, 3, 6, 12 and 24 hours respectively. Upon ABA treatment, the SCP46 transcription level was unchanged at 3h, but sharply increased to 18 fold and 51 fold at 6h and 12h respectively, and then decreased back to a low level at 24h, when compared with the control (Fig 2D). Such an ABA-induced expression pattern strongly suggested that SCP46 may be involved in ABA signaling in rice. On the contrary, we observed an opposite trend of SCP46 in response to BR (Fig 2E). The BR treatment significantly repressed the gene transcription level at 6h and 12h while increased the level at 24h, which is in accordance to the antagonistic role of BR to ABA [35]. To our surprise, we did not observe any featured expression pattern by the GA induction (Fig 2F), though GA is also antagonistic to ABA, indicating that SCP46 is functionally irrelevant to GA response.
To gain an in-depth view of the SCP46 profile, we further conducted RNA in-situ hybridization experiment in developing seeds of Nipponbare at various stages. In the early developing seeds (2–3 DAP, 1–2 mm in length), SCP46 was mainly detected in the embryo and the inner layer of the seed coat (Fig 2G). Signals were also observed on cellularized endosperm cells, but in a weaker manner than the embryo (Fig 2G). With the development of seeds (4–5 DAP, 3–4 mm in length), SCP46 was strongly accumulated in the embryo, endosperm and particularly in the aleurone layers. However, only weak expression of SCP46 was detected in other parts of the seeds such as the pericap layer (Fig 2H). This type of expression pattern extended to the later stage (10–15 DAP, 6-7mm in length), except that the SCP46 signal gradually weakened as the stage progressed (Fig 2I). No signals were detected when a sense probe was used as a negative control (Fig 2J). Given that sucrose and other photosynthesis assimilates are transported through the aleurone layers to the embryo sac cavity [36], a high expression in aleurone layer suggested that SCP46 may be involved in rice grain filling.
Knock-down of SCP46 affected rice grain size and seed germination
To dissect the biological roles of SCP46 in rice seed development, we generated 10 independent SCP46 RNAi suppressing lines in the background of Kitaake (Oryza sativa, ssp.japonica), of which 6 lines showed substantial reduction of SCP46 and similar phenotype in T0 generation (S1 Fig). For a better description of the plant phenotypes, the data of two representative T0 lines scp46-1 and scp46-2 are presented here. Our qRT-PCR results showed that the SCP46 expression levels were significantly reduced to 5% and 15% of the wild-type plants respectively, suggesting a substantial reduction of the gene (P<0.01) (Fig 3C). Through a whole-life-cycle phenotype characterization, no obvious phenotypes were observed in the RNAi lines, except that both scp46-1 and scp46-2 showed significant reduction of grain size when compared with the wild-type. Under our growth conditions, the grain length of Kitaake was about 7.28 mm, whereas the grain length of scp46-1 and scp46-2 decreased to 6.51mm and 6.7mm respectively (Fig 3B and 3E). Meanwhile, the grain width of scp46-1 and scp46-2 also had a 7% and 8.4% reduction when compared with wild-type (Fig 3A and 3F). As the consequence of the smaller grain size, the 1000-grain-weights of RNAi lines were only 77.0–83.6% of that of the wild-type (Fig 3D). Furthermore, the high expression in embryo as well as the ABA-induced expression pattern intrigued us to test the function of SCP46 in seed germination. In the MS medium with or without ABA added, both the seeds of scp46-1 and scp46-2 exhibited faster germination. When there is no ABA added in the MS medium, 79.7% of the wild-type seeds germinated in 72 hours, whereas 85.3% and 86.3% of the seeds were able to germinate for scp46-1 and scp46-2 respectively at the same time point (Fig 3G and 3H). The difference was further magnified when ABA was added into the medium. Under the 2 μM ABA treatment, there were 29.6% and 32.7% more of the scp46 seeds were germinated than the wild-type at 72 hours. The 5 μM ABA treatment experiment exhibited a similar result as 2 μM ABA, though it took a longer time for all the seeds to germinate (Fig 3G and 3H). The enhanced germinability of SCP46 RNAi seeds suggested that this gene may be involved in ABA signaling, and knock-down of SCP46 repressed the rice sensitivity to ABA. In the following T1 generation, we consistently observed the similar phenotypes of SCP46 RNAi seeds for 10 tested lines, thus we concluded that these phenotypes were attributed to the knock-down of SCP46 (S2 Fig).
Fig 3. Phenotypical characterization of scp46 grain size and seed germination.
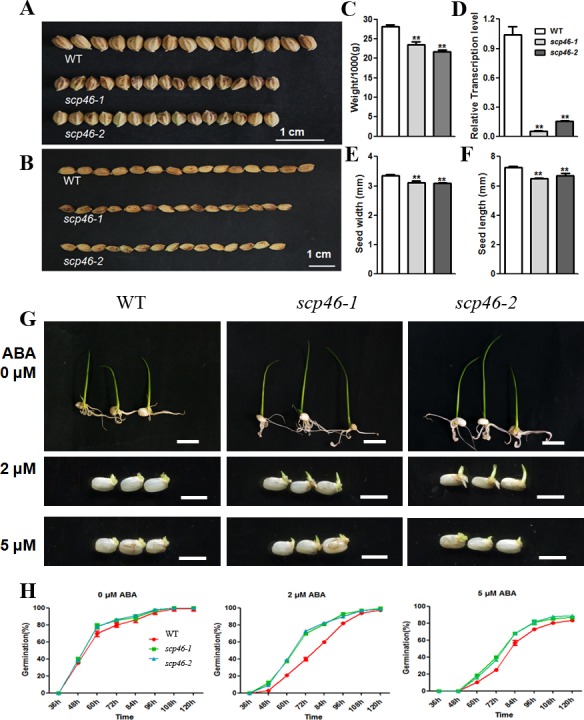
(A-B) Comparison of the width (A) and length (B) of scp46 and WT grains; (C) qRT-PCR analysis of SCP46 expression level in the seeds of RNAi and WT lines. 1000-grain-weight (D), grain length (E) and width (F) of RNAi and WT lines. (G) Seed germination of scp46 and WT seeds under 0, 2 and 5 μM ABA. Pictures were taken at 120 hours after the germination assay. (H) Germination time courses of scp46 and WT seeds. Asterisks indicate the significance of differences between Wild-type and scp46 lines as determined by Student’s t test analysis: ** P<0.01.
SCP46 regulates grain filling and seed germination related genes
To find out the genes regulated by SCP46, RNA-seq experiment was performed for the Wild-type and scp46-1 developing seeds at 7 DAP by using Illumina HiSeqTM 2000 platform. For Wild-type and scp46-1, 14,244,606 and 12,086,473 high quality (Q30>92%), clean reads were generated respectively, out of which 93.9% and 93.8% could be mapped into the rice genome. FPKM (Fragments Per Kilobase of transcript per Million fragments mapped) was employed to evaluate the gene transcriptional abundance. As a result, a total of 1835 genes were found to be differentially expressed between the RNAi plant and Wild-type, including 808 genes which showed over 2 fold up-regulation and 1027 genes were down-regulated in scp46-1 (|log 2Ratio|≥1; FDR<0.001) (S1 Table). Among these DEGs (Differentially Expressed Genes), several have been known to be functionally relevant to rice seed development. For example, SPK (Suc synthase Protein Kinase) (LOC_Os10g39420), OsGLN1 (Glutamine synthesiase I) (LOC_Os03g12290), OsPPDKA (Pyruvate orthophosphate dikinase A) (LOC_Os03g31750) and OsEIN2 (Ethylene Insensitive 2) (LOC_Os07g06130) are important regulator of rice grain filling, while VP1 (Viviparous 1) (LOC_Os01g68370), OsRACK1 (Receptor for activated C kinase 1) (LOC_Os01g53294) and OsMT2b (Metallothionein2b) (LOC_Os05g02070) are involved in the control of seed germination. A KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway analysis of the DEGs revealed that high ratios of our DEGs were involved in the plant hormone and signal transduction, ribosome and various metabolism pathways (S3 Fig). In particular, we found that 12 DEGs are related to starch and sucrose metabolism, and 20 DEGs are related to photosynthesis antenna protein complex or photosynthesis process (S3 Fig). Given that photosynthesis and starch metabolism are two of the key processes for grain filling, the KEGG pathway result strongly supported our conclusion that SCP46 regulates grain filling in rice.
To validate the RNA-seq data, 10 DEGs were selected for qRT-PCR verification. As shown in Fig 4A, the qRT-PCR result exhibited a similar tendency in the regulation pattern with the RNA-seq experiments, though the extent of expression change may slightly vary from gene to gene, suggesting that the RNA-seq data is highly reliable in this study. Given that RNA-seq results sometimes may ignored the DEGs with minor but statistically significant differences, we also examined the mRNA abundance of 16 other reported grain filling related genes which were not considered differentially expressed in DEG analysis except OsPHO2. These genes included rice starch synthesis rate limiting enzyme genes AGPs (ADP-glucose pyrophosphorylase), starch synthesases. As a result, 11 of the tested genes were found to be down-regulated in both scp46-1 and scp46-2, which well explained the reduced grain size in scp46 plants (Fig 4B).
Fig 4. Expression analysis of SCP46 regulated genes by qRT-PCR.
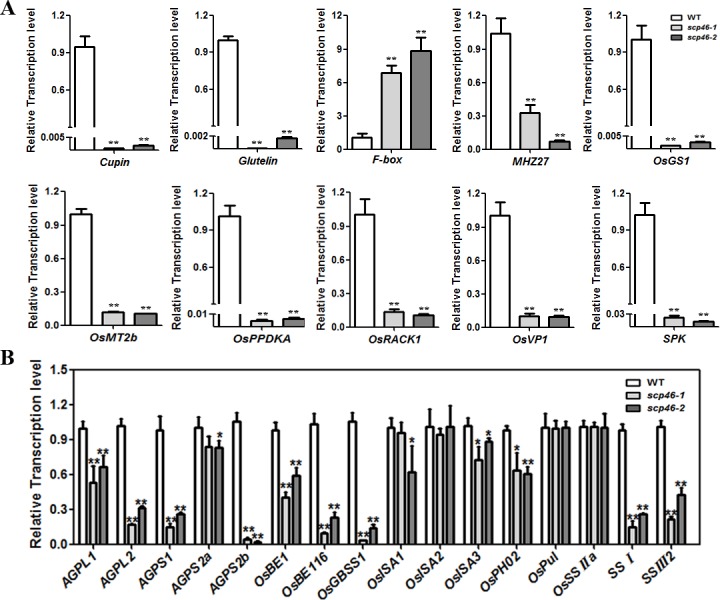
(A) qRT-PCR validation of the DEGs revealed by RNA-seq experiments; (B) the expression change of rice grain filling genes in scp46 and WT. 7 DAP seed cDNA was used as templates for this analysis. All values are based on three technical repeats and presented as means±SE.* indicates P<0.05, ** indicates P<0.01 by t-test.
SCP46 interacts with Dehydration-Induced 19–1 in yeast
To search for the SCP46 interactive proteins, the full coding sequence of SCP46 was fused with GAL4 DNA-BD domain, and used as a bait to screen a cDNA expression library constructed from Nipponbare young seedling mRNA. A total of 8 million colonies were screened and resulted in the identification of 9 unique SCP46 interactive proteins (Table 2). Although the 9 proteins are functionally unknown, the DI19-1 (Dehydration-Induced 19–1) protein attracted our interest. DI19 is a group of transcription factors containing two unusual, but evolutionally conserved Cys2-His2 (C2H2) putative zinc finger domains [37]. DI19 proteins are responsive to various abiotic stresses like dehydration, heat and salinity. Though Arabidopsis DI19s are not induced by ABA, the members in cotton, maize and rice have been reported to be involved in ABA signaling [38–40]. DI19-1 was constitutively expressed in all the tested tissues. However, we found that the expression of DI19-1 gradually increased with the development of seeds and reached a plateau in 25 DAP (Fig 5A), which is similar to the tendency of SCP46. On the other hand, we attempted to examine the DI19-1 expression in Wild-type and scp46 lines in response to ABA treatment. In the Wild-type, DI19-1 was induced by ABA treatment at 24 hours, indicating a role of it in ABA signaling as well. Meanwhile, DI19-1 displayed a lower expresion level at each time point in scp46s, but very similar expression tendencies as in the Wild-type, suggesting that Knock-down of SCP46 down-regulated the expression of DI19-1. To confirm the SCP46-DI19-1 interaction in yeast, we did a reciprocal hybridization by fusing SCP46 with AD and DI19-1 with BD domain respectively. As we expected, yeast co-transformed with pGBK-DI19-1 and pGAD-SCP46 could grow on the quart drop-out medium as the positive control did. Meanwhile, no colonies were visible in the selection medium for all the negative controls, suggesting the interaction between SCP46 and DI19-1 is reliable and specific in yeast.
Table 2. Interactive proteins of SCP46 identified by yeast two hybrid assay.
| Locus ID | Protein annotation |
|---|---|
| LOC_Os02g06890 | ubiquitin thioesterase OTU1 |
| LOC_Os02g10190 | uncharacterized protein |
| LOC_Os05g05950 | translocase of chloroplast 159 |
| LOC_Os05g09740 | acid phosphatase 1 |
| Os05g0334400 | chaperone protein DnaJ |
| LOC_Os05g48800 | Dehydration-induced 19–1 |
| LOC_Os06g47890 | adagio-like protein 1 |
| LOC_Os07g01990 | uncharacterized protein |
| LOC_Os09g03610 | flowering time control protein FCA |
All the interactive protein annotation was extracted from RGAP (http://rice.plantbiology.msu.edu/)
Fig 5. Expressoin profile of DI19-1 and yeast-two-hybrid assay of SCP46-DI19-1 interaction.
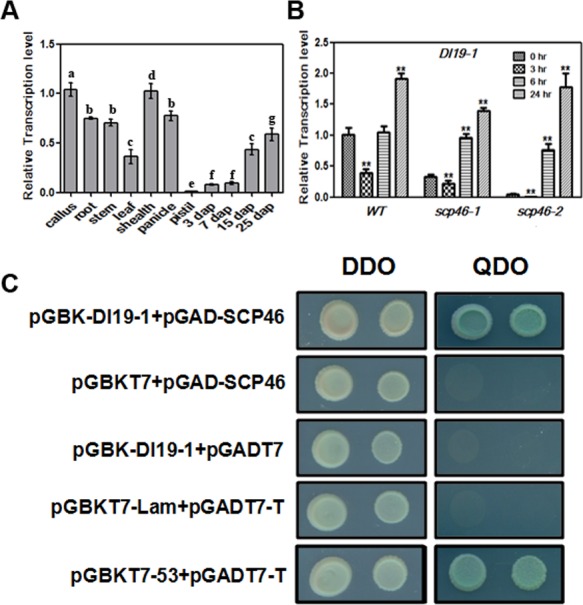
(A) Expression profile of DI19-1 in different rice tissues; Different characters indicate a statistically significant difference at P<0.05. (B) SCP46 expression in response to ABA treatment; * indicates P<0.05, ** indicates P<0.01 by t-test in comparison with 0h. (C) yeast-two-hybrid assay of SCP46-DI19-1 interaction. pGBKT7-Lam+pGADT7-T and pGBKT7-53+pGADT7-T were used as negative and positive controls as instructed by the Kit. DDO: double drop out medium (SD/-Trp-Leu); QDO: quarter drop out medium (SD/-Trp-Leu-Ade-His/+X-α-Gal).
Discussion
Proteolysis is not only a key mechanism to maintain the balance of protein synthesis and degradation, but also a major type of post-translational modification for functional protein maturation by cleaving amino acids off the protein precursors, thus altering the protein structures or biochemical activities [11]. Serine proteases are one of the largest groups of proteolytic enzymes found in plants and are associated with several essential physiological pathways. In rice, one the most important food crop in the world, at least 59 SCPs were found with detailed gene structures information in this study. Unfortunately, the function of only a few SCPs has been characterized till now. A well-documented example is that GS5/OsSCP26 was found to be a major QTL regulating grain width, filling and weight [31]. Sequence association analysis of the GS5 promoter region revealed three haplotypes controlling the grain width in 51 rice accessions from a wide geographic range, which suggested that natural variation in GS5 contributes to grain size diversity in rice. A followed up study further demonstrated that 2 SNPs (Single Nucleotide Polymorphisms) in upstream of the gene elevated the ABA-induced expression of effective GS5 in developing young panicles, promoted the mitotic division in lemma/palea via a BR signaling pathway, and eventually resulted in wider grains [41]. Despite the understanding of GS5, the function of the rest of the OsSCPs other than GS5 remains untouched. Generally, two strategies have been routinely used to predict the roles of unknown genes. One is to establish a phylogenic relationship of the candidate genes with other potential gene family members by amino acid sequence alignment, because the knowledge from homologs or orthologs from other related species are usually applicable in the candidate genes. Interestingly, we noticed that the subgroup where SCP46 located in consists 17 rice proteins but only one maize protein (2G011784). The expansion of rice members suggests more complicated functions of these SCPs in rice than maize. Meanwhile, the close phylogenetic relationship between SCP46 and 2G011784 also implies similar function of 2G011784 in maize seed development. Another way is to gain clues from the temporal- or spatial-expression profiles, as a featured expression pattern is usually indicative of the gene function in certain biological processes. Due to the limited knowledge of SCPs in plants, we relied more on the expression profile strategy to start our research on OsSCPs in this study. Interestingly, several OsSCPs displayed tissue-specific or dominant patterns, including OsSCP3 which is dominantly expressed in leaf, OsSCP10, 16 and 24 which are specifically expressed in panicle, and SCP6, 46 and 56 which are dominantly accumulated in developing seeds, suggesting crucial roles of these SCP genes in the development of corresponding tissues. Besides the tissue profile, examining the expression profile of OsSCPs under different hormone treatments or in various biological processes would be very helpful for their functional characterization.
In this study, a reverse genetic strategy starting from the tissue-specific expression pattern was employed to study the functions of OsSCPs. The microarray data together with our qRT-PCR results both clearly showed that SCP46 is dominantly expressed in developing seeds, especially in the late stages. Moreover, the RNA in-situ hybridization emphasized the accumulation of SCP46 in embryo, endosperm and aleurone layers. It is therefore rational for us to put our research focus of SCP46 function in rice grain filling. Not surprisingly, the grain size was significantly reduced when SCP46 was suppressed. The consistent phenotype observed on multiple independent RNAi lines for two consecutive generations strongly indicated that SCP46 controls rice grain size possibly via regulation of the grain fillings. Our conclusions were also supported by the RNA-seq result, in which several key coordinators of grain filling, such as SPK (LOC_Os10g39420), OsEIN2 (LOC_Os07g06130), OsGLN1 (LOC_Os03g12290) and OsPPDKA (LOC_Os03g31750) were down-regulated in scp46 lines [42–45]. SPK (LOC_Os10g39420) encoding a calmodulin-like domain protein kinase was 6.3 fold down-regulated in scp46-1. SPK is specifically expressed in the endosperm of immature seeds. Antisense SPK transformants produced defected seeds with no starch accumulated. More evidences showed that SPK controls the activity of sucrose synthase by catalyzing phosphorylation at the conserved RxxS domain, hence to regulate sucrose synthesis, which is the initial step for the biosynthesis of storage starch [44]. OsEIN2 is another down-regulated gene in SCP46 RNAi plants. OsEIN2 participates in ethylene signaling. Grain length was promoted in OsEIN2-transgenic plants and 1000-grain weight was reduced in osein2 mutants [45]. OsGLN1 encodes a cytosolic glutamine synthetase. It was reported that OsGLN1 regulates rice nitrogen and carbon assimilation, which finally affects the plant growth rate and grain filling [42]. Literature also described that regulatory phosphorylation of OsPPKDA might be involved in the rice seed development [43]. In addition to the regulators, rice starch synthesis genes and storage protein genes were affected by SCP46 as well. Our RNA-seq DEG analysis together with the qRT-PCR results identified at least 9 glutelin genes, 5 cupin genes and 16 late embryogenesis genes for storage proteins synthesis. Meanwhile, all the 5 AGP complex members, which are the rate limiting enzymes of starch synthesis, were repressed in SCP46 RNAi plants too.
Because of the high accumulation in the embryo and inducible pattern to ABA treatment of SCP46, we attempted to observe the performance of SCP46 RNAi seeds during germination. In pure MS medium, scp46 seeds showed weaker dormancy than the Wild-type. Accordingly, we found that VP1 (LOC_Os01g68370) and OsRACK1A (LOC_Os01g53294) were repressed by the knock-down of SCP46. VP1 has been known to regulate seed maturation and dormancy by interacting with TRAB1 and mediating ABA-induced transcription [46]. Meanwhile, OsRACK1A positively regulates seed germination and negatively regulates the responses of seed germination to exogenous ABA, which is similar to the phenotype observed in OsSCP46 RNAi seeds [47]. Moreover, the germinability differences between scp46 and Wild-type seeds were further magnified when ABA was applied to the medium, strongly indicating that SCP46 is involved in the ABA signaling, and the enhanced germination was due to the reduced ABA sensitivity. To detect proteins interacting with SCP46, we did yeast-two-hybrid screening and found an ABA-inducible protein DI19-1 could bind with SCP46. However, though both proteins shared similar ABA-induced pattern, it is unlikely that DI19-1 acts as a substrate of SCP46 proteolysis because DI19-1 was found to be down-regulated in the two scp46 lines as revealed by qRT-PCR (Fig 5B). Presumably, DI19-1 works with SCP46 and other unknown factors as a protein complex in proteolysis in seed development.
ABA is an isoprenoid hormone playing critical roles in various plant biological processes, including grain filling and germination [48]. Early in 1993, scientists found that ABA could control the rate of assimilate accumulation from source organ to sink organ by promoting unloading of assimilates from sieve tube to sink apoplast and uptake of unloaded assimilates by storage cells [49]. However, the effects of ABA on grain filling are dosage-dependent, as also numerous literatures described that high concentration and hypersensitivity of the ABA could inhibit grain fillings and lead to smaller seeds [50,51]. On the other hand, ABA has been well-known for its critical roles in seed dormancy and germination [52,53]. Disruption of maize ABA signaling elements VP1 gives rise to pre-harvest sprouting (PHS) [54]. PHS in modern hexaploid wheat varieties and the sprouting susceptible rice varieties was ascribed to the missplicing of wheat Vp1 genes and rice Vp1 counterpart [55,56]. It was also reported that the mutant of ABI3, the close ortholog of VP1 in Arabidopsis, showed insensitivity to ABA, reduced grain filling and activated germination, which mimicked the phenotype of vp1 and scp46 observed in this study [57,58]. More interestingly, it happened that rice VP1 was significantly down-regulated in the scp46 as indicated by both DEG and qRT-PCR results. Given the down-regulation of OsVP1 and similar phenotype observed in Arabidopsis, we proposed that OsVP1 may be involved in the SCP46–regulated rice grain filling and seed germination.
Materials and Methods
Sequence alignments and phylogenetic analysis
The information regarding the SCP genomic sequence, CDS sequence and protein sequence was downloaded from the Rice Genome Annotation Project (http://rice.plantbiology.msu.edu/). The sequence of maize SCPL gene family was downloaded from MaizeGDB (http://www.maizegdb.org/). The numbers of the SCPL gene family were analyzed by ClustalW (MEGA 7.0). The phylogenetic tree was constructed in MEGA7.0 using the Maximum Likelihood. The evolutionary distances were computed using the Jones-Taylor-Thornton (JTT) model, and the substitutions type was amino acid. There were about 1,000 replicates performed in each analysis in the bootstrap test to obtain confidence support. The ML tree was searched using the Nearest-Neighbor-Interchange (NNI). The branch length scale bar indicates the evolutionary distance.
Plant materials and growth condition
Nipponbare, Kitaake (Oryza sativa, ssp japonica) and all the transgenic plants used in this study were grown in the experimental field of China National Rice Research Institute in the summer of 2014 and 2015. The callus, root, stem, leaf, sheath, pistil and seeds in different developmental stages of Nipponbare, and 7 DAP seeds in SCP46 RNAi and Kitaake were put into liquid nitrogen immediate after harvest and stored at -80°C for total RNA isolation. For the phytohormone treatment, 14-day-old Nipponbare young seedlings with intact roots were kept in a beaker with water containing 100μM ABA (Sigma, St Louise, U.S.A.), 100μM GA (Sigma, St Louise, U.S.A.) and 10μM BR (Sigma, St Louise, U.S.A.) respectively, the whole seedling samples were collected at the corresponding time points and immediately used for RNA isolation. ABA treatment on 14-day-old Kitaake and scp46 young seedlings was conducted by the same method as above.
RNA in-situ hybridization
Nipponbare seeds at different developmental stages were fixed in diethyl pyrocarbonate (DEPC)-treated 70% FAA fix solution (70% ethanol, 5% glacial acetic acid and 3.7% formaldehyde) overnight at 4°C, then dehydrated through a series of ethanol solutions, infiltrated with xylene and embedded in paraffin. The embedded tissues were cut into 9-μm-thick sections. The SCP46 probe was PCR amplified by primer set In-SCP46F+In-SCP46R. After being cloned into pGEM-T Easy vector (Promega, Madison, U.S.A.), the segmental sequence was transcribed in vitro to synthesize sense and antisense probes from either T7 or SP6 promoter using the Digoxigenin RNA labeling kit (Roche, Basilea, Swiss Confederation). The sense strand was used as a negative reference. The hybridization and immunological detection were conducted as described by Zhang et al.2010 [59].
Vector construction
To construct the SCP46 RNAi construct, a 300 bp-long, unique segment of SCP46 coding sequence was amplified by the primer set SCP46RNAif and SCP46RNAir. The PCR fragment was firstly cloned into pENTR/D by TOPO cloning kit (Invitrogen, Carlsbad, U.S.A.). The intermediate vector was then linearized by EcoRV cut and finally cloned into pANDA vector by attL×attR recombination reaction [60].
For the SCP46 bait construct, the entire SCP46 CDS fragment was amplified by PCR using BamHI and SalI linker primers and cloned into vector pGBKT7 by ligation. The DI19-1 CDS fragment was amplified by PCR using BamHI and EcoRI linker primers, then digested and linked with the pGADT7 vector. All the primers used are listed in S2 Table.
Phenotypic analysis of rice transgenic plant seeds
The length, width and thousand-grain-weight of WT and scp46 lines were examined by a seed phenotyping system (Wangsheng, Hangzhou, China), and the data was analyzed by student t-test in Microsoft Office Excel 2010. For the seed germination assay, Kitaake and scp46 seeds were firstly surface sterilized by washing in 70% ethanol for 1 minutes and 50% bleach for 30 minutes. After being flushed with autoclaved water for several times, the seeds were placed on Murashige and Skoog medium containing 0, 2 and 5 μM ABA respectively, and kept in a growth chamber (28°C, 14 hours daytime and 10 hours night time). Seeds which formed green shoots were considered as germinated. Seed germination was scored every 12 hours for 5 days. For each medium with different ABA concentration, at least 300 seeds (100 seeds with three replicates) of each line were tested. PCR with a pair of hygromycin primers was conducted for all the seedlings to exclude the T-DNA negative lines for the germination assay.
RNA-seq
7 DAP rice seed total RNA was analyzed by Nanodrop 2000 spectrophotometer and Agilent 2100 Bioanalyzer to assure the proper purity, concentration and integrity. The qualified RNA was preceded for sequencing library construction as described by Hou et al (2015) [61]. The high-throughput sequencing was performed using the Illumina HiSeq™ 2000 platform when the quality of library was up to standard. The generated clean reads were aligned with reference sequences of rice in RGAP (http://rice.plantbiology.msu.edu/). Gene expression changes between the samples were analyzed by SOAP aligner/SOAP2 software. The KEGG pathway analysis of the DEGs was ultimately done by Biomarker technologies Co., Ltd., Beijing, China.
RNA isolation and qRT-PCR
The RNA of all the tissues except developing seeds was extracted by Trizol (Invitrogen, Carlsbad, U.S.A.) according to the manufacturer's instructions. For developing seeds RNA extraction, a modified SDS-trizol method was applied as described by an online protocol (http://www.ag.arizona.edu/research/larkinslab/protocols/RNA%20extraction%20from%20endosperm%20-SDS-Trizol%20combo.pdf). Briefly, 0.5 grams of grinded seed powders were treated with 300 μL SDS RNA extraction buffer (50mM Tris.Cl pH 8.0, 5mM EDTA pH 8.0, 150mM LiCl and 1% SDS), 300 μL Phenol (pH 8.0): Chloroform = 1:1, the RNA in supernatant was then extracted by Trizol as for other tissues. The total RNA was quantified by Nanodrop spectrometer and 2 μg of each was used for reverse transcription by using M-MLV reverse transcriptase (Takara) with oligo(dT)20 (Invitrogen, Carlsbad, U.S.A.) as primer. A total reaction volume of ten microliter (5 μL THUNDERBIRD SYBR® qPCR Mix [Toyobo], 1 μL cDNA, 0.2μL primers, and 3.8μL water) was performed on the CFX96 touch realtime PCR detection system (Bio-rad, Hercules, U.S.A.) with three technical replicates. Expression was assessed by evaluating threshold cycle (CT) values. The relative expression level of tested genes was normalized to ubiquitin gene and calculated by the 2-ΔΔCT method [62]. The sequence of all the primers used in this study can be found in S2 Table.
Yeast-two-hybrid assay
The Matchmaker™ Gold Yeast Two-Hybrid system (Clontech, Dalian, China) was used to perform the yeast two-hybrid experiment. The rice young seedling cDNA library in Y2HGold strain was purchased from Clontech. Both the bait vector was transformed into yeast Y187 strain and mated with cDNA library according to the manufacturer’s instructions. The SCP46-DI19 yeast two hybrid assay was conducted by co-transformation of both plasmids into Y2HGold. The co-transformed yeast cell was cultured in SD/-Trp-Leu solid medium, and the interaction was confirmed by the colony growth in SD/-Trp-Leu-Ade-His solid medium with X-α-Gal.
Supporting Information
(TIF)
(TIF)
(TIF)
(XLSX)
(XLS)
Acknowledgments
We thank Dr. Hana Mujahid of Mississippi State University, U.S.A. for critical review of the manuscript, Ms. Xiaoling Qiu for genetic transformation of the rice plants.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Agricultural Sciences and Technologies Innovation Program of Chinese Academy of Agricultural Sciences (CAAS) to Rice Reproductive Developmental Biology Group (No grant numbers) (http://www.caas.cn/kjcxgczl/index.shtml), the “Elite Youth” program (CAAS) to JZ (No grant number) (http://www.caas.cn/rc/qnyc1/index.shtml), and the National Natural Science Foundation of China to JZ (grant number: 31401366) (http://www.nsfc.gov.cn/). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Hayashi R, Moore S, Stein WH (1974) Serine at the active center of yeast carboxypeptidase. Journal of Biological Chemistry 248: 8366–8369. [PubMed] [Google Scholar]
- 2.Feng Y, Yu C (2009) Genome-wide comparative study of rice and Arabidopsis serine carboxypeptidase-like protein families. Journal of Zhejiang University 35: 1–15. [Google Scholar]
- 3.Breddam K (1986) SERINE CARBOXYPEPTIDASES. A REVIEW. Carlsberg Res Commun 51: 83–128. [Google Scholar]
- 4.Sapio MR, Fricker LD (2014) Carboxypeptidases in disease: insights from peptidomic studies. Proteomics Clin Appl 8: 327–337. 10.1002/prca.201300090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Oda K (2012) New families of carboxyl peptidases: serine-carboxyl peptidases and glutamic peptidases. J Biochem 151: 13–25. 10.1093/jb/mvr129 [DOI] [PubMed] [Google Scholar]
- 6.Galjart NJ, Gillemans N, Harris A, van der Horst GT, Verheijen FW, Galjaard H, et al. (1988) Expression of cDNA encoding the human "protective protein" associated with lysosomal beta-galactosidase and neuraminidase: homology to yeast proteases. Cell 54: 755–764. [DOI] [PubMed] [Google Scholar]
- 7.Fraser CM, Rider LW, Chapple C (2005) An expression and bioinformatics analysis of the Arabidopsis serine carboxypeptidase-like gene family. Plant Physiol 138: 1136–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Feng Y, Xue Q (2006) The serine carboxypeptidase like gene family of rice (Oryza sativa L. ssp. japonica). Funct Integr Genomics 6: 14–24. [DOI] [PubMed] [Google Scholar]
- 9.Jones CG, Lycett GW, Tucker GA (1996) Protease inhibitor studies and cloning of a serine carboxypeptidase cDNA from germinating seeds of pea (Pisum sativum L.). Eur J Biochem 235: 574–578. [DOI] [PubMed] [Google Scholar]
- 10.Wajant H, Mundry KW, Pfizenmaier K (1994) Molecular cloning of hydroxynitrile lyase from Sorghum bicolor (L.). Homologies to serine carboxypeptidases. Plant Mol Biol 26: 735–746. [DOI] [PubMed] [Google Scholar]
- 11.Tripathi LP, Sowdhamini R (2006) Cross genome comparisons of serine proteases in Arabidopsis and rice. BMC Genomics 7: 200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mugford ST, Osbourn A (2010) Evolution of serine carboxypeptidase-like acyltransferases in the monocots. Plant Signal Behav 5: 193–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dal Degan F, Rocher A, Cameron-Mills V, von Wettstein D (1994) The expression of serine carboxypeptidases during maturation and germination of the barley grain. Proc Natl Acad Sci U S A 91: 8209–8213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wen J, Li J, Walker JC (2012) Overexpression of a serine carboxypeptidase increases carpel number and seed production in Arabidopsis thaliana. Food and Energy Security 1: 61–69. [Google Scholar]
- 15.Li J, Lease KA, Tax FE, Walker JC (2001) BRS1, a serine carboxypeptidase, regulates BRI1 signaling in Arabidopsis thaliana. Proc Natl Acad Sci U S A 98: 5916–5921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Domínguez F, González M, Cejudo FJ (2002) A germination-related gene encoding a serine carboxypeptidase is expressed during the differentiation of the vascular tissue in wheat grains and seedlings. Planta 215: 727–734. [DOI] [PubMed] [Google Scholar]
- 17.Cercos M, Urbez C, Carbonell J (2003) A serine carboxypeptidase gene (PsCP), expressed in early steps of reproductive and vegetative development in Pisum sativum, is induced by gibberellins. Plant Mol Biol 51: 165–174. [DOI] [PubMed] [Google Scholar]
- 18.Bienert MD, Delannoy M, Navarre C, Boutry M (2012) NtSCP1 from tobacco is an extracellular serine carboxypeptidase III that has an impact on cell elongation. Plant Physiol 158: 1220–1229. 10.1104/pp.111.192088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mugford ST, Milkowski C (2012) Serine carboxypeptidase-like acyltransferases from plants. Methods Enzymol 516: 279–297. 10.1016/B978-0-12-394291-3.00006-X [DOI] [PubMed] [Google Scholar]
- 20.Milkowski C, Strack D (2004) Serine carboxypeptidase-like acyltransferases. Phytochemistry 65: 517–524. [DOI] [PubMed] [Google Scholar]
- 21.Schaller A (2004) A cut above the rest: the regulatory function of plant proteases. Planta 220: 183–197. [DOI] [PubMed] [Google Scholar]
- 22.Lehfeldt C, Shirley AM, Meyer K, Ruegger MO, Cusumano JC, Viitanen PV, et al. (2000) Cloning of the SNG1 gene of Arabidopsis reveals a role for a serine carboxypeptidase-like protein as an acyltransferase in secondary metabolism. Plant Cell 12: 1295–1306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mugford ST, Qi X, Bakht S, Hill L, Wegel E, Papadopoulou K, et al. (2009) A serine carboxypeptidase-like acyltransferase is required for synthesis of antimicrobial compounds and disease resistance in oats. Plant Cell 21: 2473–2484. 10.1105/tpc.109.065870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Milkowski C, Baumert A, Schmidt D, Nehlin L, Strack D (2004) Molecular regulation of sinapate ester metabolism in Brassica napus: expression of genes, properties of the encoded proteins and correlation of enzyme activities with metabolite accumulation. Plant J 38: 80–92. [DOI] [PubMed] [Google Scholar]
- 25.Wolf AE, Dietz KJ, Schroder P (1996) Degradation of glutathione S-conjugates by a carboxypeptidase in the plant vacuole. FEBS Lett 384: 31–34. [DOI] [PubMed] [Google Scholar]
- 26.Liu L, Wang J, Zhang Z, Zhao J, Pan G (2013) Cloning and Expression Analysis of Serine Carboxypeptidases in Maize (Zea mays L.). Acta Agronomica Sinica 39: 164–171. [Google Scholar]
- 27.Zhang J, Guo D, Chang Y, You C, Li X, Dai X, et al. (2007) Non-random distribution of T-DNA insertions at various levels of the genome hierarchy as revealed by analyzing 13 804 T-DNA flanking sequences from an enhancer-trap mutant library. Plant J 49: 947–959. [DOI] [PubMed] [Google Scholar]
- 28.Xiao JH, Wu CY, Yuan M, Wang NL, Fang YR, Yang M, et al. (2015) The progress and perspective of rice functional genomics research in China (in Chinese). Chin Sci Bull 60: 1711–1722. [Google Scholar]
- 29.Washio K, Ishikawa K (1994) Organ-specific and hormone-dependent expression of genes for serine carboxypeptidases during development and following germination of rice grains. Plant Physiol 105: 1275–1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu H, Wang X, Zhang H, Yang Y, Ge X, Song F, et al. (2008) A rice serine carboxypeptidase-like gene OsBISCPL1 is involved in regulation of defense responses against biotic and oxidative stress. Gene 420: 57–65. 10.1016/j.gene.2008.05.006 [DOI] [PubMed] [Google Scholar]
- 31.Li Y, Fan C, Xing Y, Jiang Y, Luo L, Sun L, et al. (2011) Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet 43: 1266–1269. 10.1038/ng.977 [DOI] [PubMed] [Google Scholar]
- 32.Kawahara Y, de la Bastide M, Hamilton JP, Kanamori H, McCombie WR, Ouyang S, et al. (2013) Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice (N Y) 6: 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lawrence CJ, Harper LC, Schaeffer ML, Sen TZ, Seigfried TE, Campbell DA, et al. (2008) MaizeGDB: The maize model organism database for basic, translational, and applied research. Int J Plant Genomics 2008: 496957 10.1155/2008/496957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang L, Xie W, Chen Y, Tang W, Yang J, Ye R, et al. (2010) A dynamic gene expression atlas covering the entire life cycle of rice. Plant J 61: 752–766. 10.1111/j.1365-313X.2009.04100.x [DOI] [PubMed] [Google Scholar]
- 35.Zhang S, Cai Z, Wang X (2009) The primary signaling outputs of brassinosteroids are regulated by abscisic acid signaling. Proc Natl Acad Sci U S A 106: 4543–4548. 10.1073/pnas.0900349106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bai AN, Lu XD, Li DQ, Liu JX, Liu CM (2016) NF-YB1-regulated expression of sucrose transporters in aleurone facilitates sugar loading to rice endosperm. Cell Res 26: 384–388. 10.1038/cr.2015.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Milla MA, Townsend J, Chang IF, Cushman JC (2006) The Arabidopsis AtDi19 gene family encodes a novel type of Cys2/His2 zinc-finger protein implicated in ABA-independent dehydration, high-salinity stress and light signaling pathways. Plant Mol Biol 61: 13–30. [DOI] [PubMed] [Google Scholar]
- 38.Li G, Tai FJ, Zheng Y, Luo J, Gong SY, Zhang ZT, et al. (2010) Two cotton Cys2/His2-type zinc-finger proteins, GhDi19-1 and GhDi19-2, are involved in plant response to salt/drought stress and abscisic acid signaling. Plant Mol Biol 74: 437–452. 10.1007/s11103-010-9684-6 [DOI] [PubMed] [Google Scholar]
- 39.Qin LX, Nie XY, Hu R, Li G, Xu WL, Li XB, et al. (2016) Phosphorylation of serine residue modulates cotton Di19-1 and Di19-2 activities for responding to high salinity stress and abscisic acid signaling. Sci Rep 6: 20371 10.1038/srep20371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang L, Yu C, Chen C, He C, Zhu Y, Huang W, et al. (2014) Identification of rice Di19 family reveals OsDi19-4 involved in drought resistance. Plant Cell Rep 33: 2047–2062. 10.1007/s00299-014-1679-3 [DOI] [PubMed] [Google Scholar]
- 41.Xu C, Liu Y, Li Y, Xu X, Xu C, Li X, et al. (2015) Differential expression of GS5 regulates grain size in rice. J Exp Bot 66: 2611–2623. 10.1093/jxb/erv058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tabuchi M, Sugiyama K, Ishiyama K, Inoue E, Sato T, Takahashi H, et al. (2005) Severe reduction in growth rate and grain filling of rice mutants lacking OsGS1;1, a cytosolic glutamine synthetase1;1. Plant J 42: 641–651. [DOI] [PubMed] [Google Scholar]
- 43.Chastain CJ, Heck JW, Colquhoun TA, Voge DG, Gu XY (2006) Posttranslational regulation of pyruvate, orthophosphate dikinase in developing rice (Oryza sativa) seeds. Planta 224: 924–934. [DOI] [PubMed] [Google Scholar]
- 44.Asano T, Kunieda N, Omura Y, Ibe H, Kawasaki T, Takano M, et al. (2002) Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: phosphorylation of sucrose synthase is a possible factor. Plant Cell 14: 619–628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ma B, He SJ, Duan KX, Yin CC, Chen H, Yang C, et al. (2013) Identification of rice ethylene-response mutants and characterization of MHZ7/OsEIN2 in distinct ethylene response and yield trait regulation. Mol Plant 6: 1830–1848. 10.1093/mp/sst087 [DOI] [PubMed] [Google Scholar]
- 46.Hobo T, Kowyama Y, Hattori T (1999) A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proc Natl Acad Sci U S A 96: 15348–15353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang D, Chen L, Li D, Lv B, Chen Y, Chen J, et al. (2014) OsRACK1 is involved in abscisic acid- and H2O2-mediated signaling to regulate seed germination in rice (Oryza sativa, L.). PLoS One 9: e97120 10.1371/journal.pone.0097120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hrabak EM, Chan CW, Gribskov M, Harper JF, Choi JH, Halford N, et al. (2003) The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol 132: 666–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kato T, Sakurai N, Kuraishi S (1993) The changes of endogenous abscisic acid in developing grain of two rice cultivars with different grain size. Japanese journal of crop science 62: 456–461. [Google Scholar]
- 50.Zhang H, Tan G, Yang L, Yang J, Zhang J, Zhao B, et al. (2009) Hormones in the grains and roots in relation to post-anthesis development of inferior and superior spikelets in japonica/indica hybrid rice. Plant Physiol Biochem 47: 195–204. 10.1016/j.plaphy.2008.11.012 [DOI] [PubMed] [Google Scholar]
- 51.Ahmadi A, Baker DA (1999) Effects of abscisic acid (ABA) on grain filling processes in wheat. Plant Growth Regulation 28: 187–197. [Google Scholar]
- 52.Shu K, Liu XD, Xie Q, He ZH (2016) Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol Plant 9: 34–45. 10.1016/j.molp.2015.08.010 [DOI] [PubMed] [Google Scholar]
- 53.Nakashima K, Yamaguchi-Shinozaki K (2013) ABA signaling in stress-response and seed development. Plant Cell Rep 32: 959–970. 10.1007/s00299-013-1418-1 [DOI] [PubMed] [Google Scholar]
- 54.Hoecker U, Vasil IK, McCarty DR (1995) Integrated control of seed maturation and germination programs by activator and repressor functions of Viviparous-1 of maize. Genes Dev 9: 2459–2469. [DOI] [PubMed] [Google Scholar]
- 55.McKibbin RS, Wilkinson MD, Bailey PC, Flintham JE, Andrew LM, Lazzeri PA, et al. (2002) Transcripts of Vp-1 homeologues are misspliced in modern wheat and ancestral species. Proc Natl Acad Sci U S A 99: 10203–10208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fan J, Niu X, Wang Y, Ren G, Zhuo T, Yang Y, et al. (2007) Short, direct repeats (SDRs)-mediated post-transcriptional processing of a transcription factor gene OsVP1 in rice (Oryza sativa). J Exp Bot 58: 3811–3817. [DOI] [PubMed] [Google Scholar]
- 57.Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, Goodman HM, et al. (1992) Isolation of the Arabidopsis ABI3 gene by positional cloning. Plant Cell 4: 1251–1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Nambara E, Naito S, Mccourt P (1992) A mutant of Arabidopsis which is defective in seed development and storage protein accumulations is a new abi3 allele. Plant Journal 2: 435–441. [Google Scholar]
- 59.Zhang J, Nallamilli BR, Mujahid H, Peng Z (2010) OsMADS6 plays an essential role in endosperm nutrient accumulation and is subject to epigenetic regulation in rice (Oryza sativa). Plant J 64: 604–617. 10.1111/j.1365-313X.2010.04354.x [DOI] [PubMed] [Google Scholar]
- 60.Miki D, Shimamoto K (2004) Simple RNAi vectors for stable and transient suppression of gene function in rice. Plant Cell Physiol 45: 490–495. [DOI] [PubMed] [Google Scholar]
- 61.Hou Y, Wang L, Wang L, Liu L, Li L, Sun L, et al. (2015) JMJ704 positively regulates rice defense response against Xanthomonas oryzae pv. oryzae infection via reducing H3K4me2/3 associated with negative disease resistance regulators. BMC Plant Biol 15: 286 10.1186/s12870-015-0674-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(TIF)
(TIF)
(TIF)
(XLSX)
(XLS)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.