

Abstract
Background:
Mammalian target of rapamycin (mTOR) complex 1 (mTORC1) acts as a downstream effector of phosphatidyl-inositol-3 kinase, which is frequently hyperactivated in glioblastoma multiforme and links to cell signaling in cellular proliferation, differentiation, metabolism, and survival. Although many studies have suggested the importance of mTORC1 in tumorigenesis, its role remains unclear in brain tumors other than glioblastoma.
Methods:
In the present study, we evaluated the activation of mTORC1 in 24 cases of primary central nervous system lymphoma (PCNSL).
Results:
Immunohistochemical analysis showed overexpression of Rheb, which is immediately upstream of mTORC1, in 20 cases of PCNSL. Immunohistochemical analysis also showed overexpression of phospho-4E-BP1 (Thr37/46) and phospho-S6 (Ser235/236), which are increased after mTORC1 activation as mTORC1 downstream effectors in 17 and 21 cases, respectively.
Conclusion:
Our data suggest that abnormal activation of the mTORC1 signaling pathway may cause tumor growth in patients with PCNSL.
Keywords: mTOR, phospho-4E-BP1, phospho-S6, primary central nervous system lymphoma, Rheb
INTRODUCTION
Mammalian target of rapamycin (mTOR) signaling impacts most major cellular functions, giving it an outsized role in regulating basic cell behaviors such as growth and proliferation.[19] Dysregulated mTORC1 signaling pathway drives tumorigenesis through the coordinated phosphorylation of proteins that directly regulate protein synthesis, cell-cycle progression, and metabolism, and of transcription factors that regulate the expression of genes involved in these processes.[30] The mTORC1 signaling pathway is inappropriately activated and inhibition of mTORC1 improves progression-free survival in patients with renal-cell carcinoma, pancreatic neuroendocrine tumor, breast cancer, and lymphangioleiomyomatosis.[3,11,25,38] In the brain, abnormally increased activation of mTORC1 contributes not only to benign tumor diseases such as tuberous sclerosis and Lhermitte–Duclos disease but also to malignant tumors such as glioblastoma multiforme; mTORC1 inhibitors have also been used to reduce the volume of subependymal giant-cell astrocytomas (SEGA) in patients with tuberous sclerosis.[1,6,14,18,20] These suggest that mTORC1 inhibitors may be potential alternatives to conventional chemotherapies in patients with brain tumors in which the mTORC1 signaling pathway is abnormally activated, and yet, mTORC1 activation in human brain tumors other than gliomas has not been well studied.
A small G protein Ras homologue enriched in brain (Rheb), regulated by a heterodimer consisting of tuberous sclerosis 1 (TSC1) and TSC2, directly interacts with mTORC1 and strongly stimulates its kinase activity when it is converted into its active GTP-bound state [Figure 1].[19] Overexpression of Rheb activates mTORC1, resulting in cell growth, cell cycle progression, and tumorigenesis.[23,27,32] Eukaryotic translation initiation factor 4E (eIF4E) binding protein 1 (4E-BP1) inhibits translation initiation by interfering with the interaction between the mRNA cap-binding protein eIF4E and eIF4G1, and consequently blocks cell growth and proliferation.[22,33] The phosphorylation of 4E-BP1 by activated mTORC1 prevents its binding to eIF4E, enabling it to participate in the formation of the eIF4F complex that is required for the initiation of cap-dependent translation [Figure 1].[19] S6, a ribosomal protein, is a component of the 40 S ribosomal subunit that is phosphorylated by the p70 ribosomal S6 kinases, S6K1 and S6K2 [Figure 1]. S6K1 and S6K2 activation is directly regulated by mTORC1 [Figure 1]. In diseases caused by the hyperactivation of mTORC1, for example, SEGA and Lhermitte-Duclos disease, phosphorylated S6 (p-S6) is highly expressed as a downstream effector of mTORC1.[1,4,7] Because it is difficult to directly measure mTORC1 activity, expression of downstream and/or upstream proteins has been measured as an alternative method.[19] In particular, p-S6 expression measured with immunohistochemistry or Western blotting tends to correlate with mTORC1 activation and appears to be associated with tumor sensitivity to mTORC1 inhibitors such as rapamycin and its analogs.[5,15,36,37]
Figure 1.
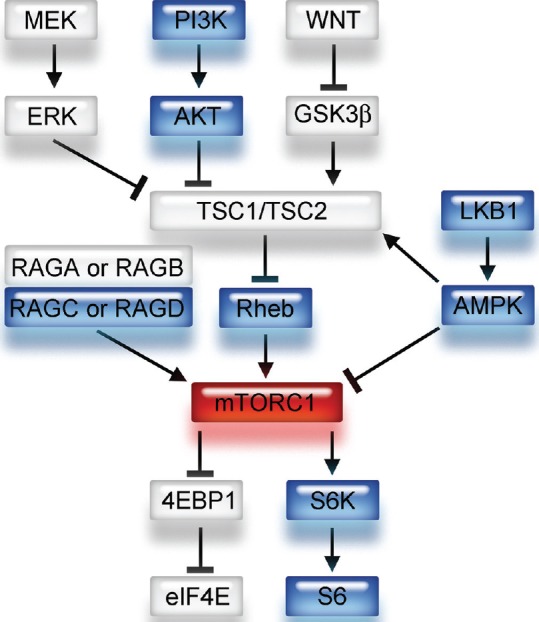
Schema of upstream and downstream signaling pathways of mTORC1. mTORC1 has several upstream pathways. Note that Rheb is a direct upstream component and 4E-BP1 and S6 are downstream components of the mTORC1 signaling pathway. GTP-bound Rheb activates mTORC1 and contributes to the phosphorylation of 4E-BP1 and S6
Primary central nervous system lymphomas (PCNSL) are estimated to account for approximately 3% of all CNS tumors. Despite high chemosensitivity and radiosensitivity and remissions being frequently of short duration, treatment of PCNSL remains challenging.[9] Elucidation of activated signaling pathways in PCNSL might contribute to better understanding of the mechanism of early remissions of PCNSL after treatments. In the present study, we examined mTORC1 activation in human PCNSLs using immunohistochemistry for its upstream and downstream markers, Rheb, phospho-4E-BP1 (p-4E-BP1), and p-S6. Our data demonstrate that the mTORC1 signaling pathway is activated in many cases of PCNSL, suggesting that abnormal mTORC1 activation may cause growth of PCNSL.
MATERIALS AND METHODS
This retrospective study was approved by the Institutional Review Boards of Shiga University of Medical Science and was performed in accordance with the ethical standards laid down in the 1975 Declaration of Helsinki and its later amendments.
Tissue samples of consecutive cases of PCNSL (n = 27) obtained from January 1, 1998 to March 31, 2015 were collected from the surgical pathology files of the Shiga University of Medical Science Hospital, Shiga, Japan. All samples were obtained during the first surgery before chemotherapy and radiotherapy.
Sections of Formalin-fixed, paraffin-embedded tissues were stained with hematoxylin and eosin. Immunohistochemistry was performed for Rheb, which is an upstream regulator of mTORC1, and for p-4E-BP1 (Thr37/46) and p-S6 (Ser235/236), which are downstream effectors of mTORC1 [Figure 1] (all antibodies were purchased from Cell Signaling Technology, Danvers, MA). We performed antigen retrieval with 0.01 M citrate buffer (pH 6.0) for 16 min in a microwave oven. After peroxidase activity was quenched with 3% hydrogen peroxide in water, biotin and avidin binding sites were blocked with Avidin/Biotin Blocking Kit (Vector Laboratories, Burlingame, CA) according to the manufacturer's protocol. Primary antibodies (Rheb at 1:250, p-4E-BP1 at 1:250, p-S6 at 1:250) were diluted in 5% normal goat serum in tris-buffered saline and applied for 24 h at 4°C, followed by biotinylated secondary antibodies (Vector) at 1:250 dilution for 2 h and avidin-biotin complex (Elite ABC; Vector) for 1 h at room temperature. Diaminobenzidine tetrahydrochloride was used as the enzyme substrate to visualize specific antibody localization. Slides were counterstained with hematoxylin. With having mTORC1 activation at high frequency because of TSC inactivation, SEGA were used for a positive control.[4] Tissue samples of consecutive cases of SEGA (n = 3), which were surgically removed from tuberous sclerosis patients because of mass effect and/or hydrocephalus, were collected from the surgical pathology files of our hospital. This positive control was processed concurrently with each tested tumor. For a negative antibody control, the tumors were tested without the primary antibodies with each tumor. Expressions of Rheb, p-4E-BP1, and p-S6 were also evaluated in normal human brain (cerebral cortex and cerebral medullary substance) tissues (n = 4) and normal human lymph node tissues (n = 4), which were autopsy-derived tissues purchased from Capital Bioscience (Gaithersburg, MD), ProSci (Poway, CA), US Biomax (Rockville, MD), Alpha Diagnostic International (San Antonio, TX), or BioChain (Newark, CA).
Immunohistochemical staining quantification was categorized into two groups [low expressors, negative or focally positive (Figure 2a: Focally positive) in less than 25% of tumor cells; high expressors, positive in 25% (Figure 2b: 25% positive) or more tumor cells] as described in previous reports in which ≥25% or ≥20% was used as high expression.[1,15] Scoring of staining on whole tissue sections was performed by an independent observer (AS).
Figure 2.
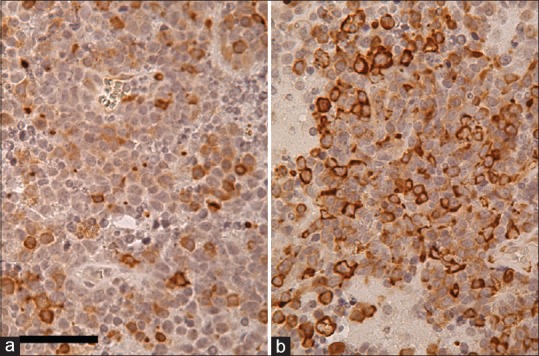
Examples of scoring of immunohistochemical staining: focally positive (low expresser) (a) and 25% positive (barely high expresser) (b). Scale bar = 50 μm in (a)
RESULTS
In the normal brain tissues, Rheb was strongly expressed in neuronal soma in the cortex as “Ras homologue enriched in brain,” and weakly or not expressed in glial cells [Figure 3a]. p-4E-BP1 was expressed in neither neurons nor glial cells [Figure 3b]. p-S6 was weakly or not expressed in neurons, and not expressed in glial cells [Figure 3c]. In the normal lymph node tissues, neither Rheb nor p-4E-BP1 was expressed and p-S6 was expressed in a few lymphocytes [Figure 3d-f]. In the tissues of SEGA, Rheb, p-4E-BP1, and p-S6 were expressed moderately in many tumor cells [Figure 3g-i]. The results of the immunohistochemical analysis of Rheb, p-4E-BP1, and p-S6 in PCNSLs are shown in Table 1. In three cases, because samples were small and contained less than 600 tumor cells per section, the result were omitted. All PCNSLs are diffuse large B-cell lymphomas (DLBCL). Rheb was highly expressed in 20 cases of PCNSL (n = 24) [Figure 4a]. High expression of p-4E-BP1 and p-S6 were also observed in 17 cases and 21 cases, respectively (n = 24) [Figure 4b and c; Table 1]. In 12 specimens, all three mTORC1 activation markers were highly expressed, suggesting that mTORC1 had absolutely been activated in these specimens [Table 1]. In 7 specimens, Rheb and a downstream marker were highly expressed, suggesting that mTORC1 had probably been activated [Table 1]. However, it remains unclear why only one downstream had been highly activated. Three specimens highly expressed two downstream markers but not Rheb, suggesting that mTORC1 had been activated probably by other upstream effector than Rheb [Table 1]. In 1 specimen, only p-4E-BP1 was highly expressed, suggesting that mTORC1 might have been activated with activation of neither Rheb nor S6K-S6 signaling pathways [Table 1]. In 1 specimen, only Rheb was highly expressed, suggesting that mTORC1 had not been activated [Table 1]. These results suggest that mTORC1 had been highly activated in at least half of cases of PCNSL.
Figure 3.
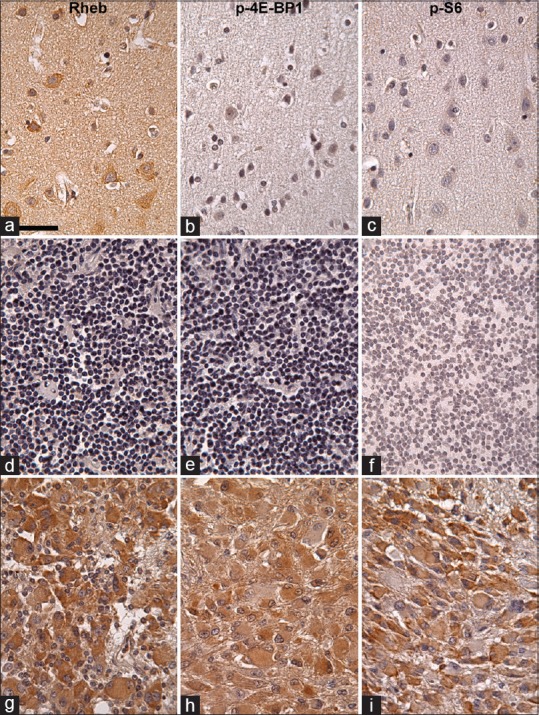
Rheb (a, d, g), p-4E-BP1 (b, e, h), and p-S6 (c, f, i) immunohistochemistry in the normal human cerebral cortex (a-c), normal lymph node (d-f), and SEGA (g-i) counterstained with hematoxylin. Most of the neuronal somas are strongly Rheb-positive in cytoplasm, not p-4E-BP1 positive, and mildly or not p-S6 positive (a-c). Glial cells are Rheb- p-4E-BP1-, and p-S6-negative (a-c). In normal lymph node, Rheb, p-4E-BP1, or p-S6 is not expressed (d-f). In SEGA, Rheb, p-4E-BP1, and p-S6 are moderately expressed in tumor cells, suggesting that the mTORC1 signaling pathway is activated in these cells. Scale bar = 50 μm in (a)
Table 1.
Rheb, p-4E-BP1 and p-S6 expression in PCNSLs
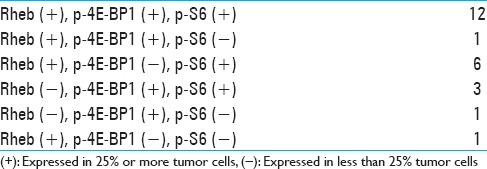
Figure 4.
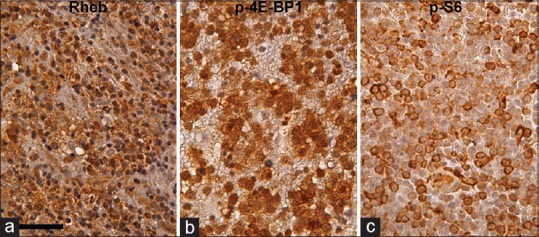
Rheb (a), p-4E-BP1 (b), and p-S6 (c) immunohistochemistry in PCNSL counterstained with hematoxylin. Most of the tumor cells are Rheb-positive in cytoplasm. Furthermore, some of the tumor cells are strongly p-4E-BP1 and p-S6 positive, suggesting that the mTORC1 signaling pathway is activated in these tumor cells. Scale bar = 50 μm in (a)
DISCUSSION
The present report describes activation of the mTORC1 signaling pathway in many cases of PCNSL. Both proliferation regulator p-4E-BP1 and cell size regulator p-S6 were highly expressed in half of cases of PCNSL, suggesting that mTORC1 activation contributes to not only rapid proliferation but also hypertrophy of the tumor cells of PCNSL.[10,29]
Lymphomas and mTORC1
Numerous studies have shown that the mTORC1 signaling pathway plays important roles in the biology of malignant cells, including deregulation of the cell cycle, suppression of apoptotic machinery and resistance to chemotherapies. The mTORC1 signaling pathway has been reported to be activated in systemic lymphomas, for example, Hodgkin lymphoma, anaplastic large cell lymphoma, diffuse large B-cell lymphoma, etc.[8] In Hodgkin lymphoma, treatment with rapamycin, an mTORC1 inhibitor, results in G1-S cell-cycle arrest and enhances the cytotoxicity of other chemotherapeutic agents against Hodgkin/Reed-Sternberg cells.[17] In other lymphomas, rapamycin treatment has also been shown to induce G1-S cell-cycle arrest and to enhance the cytotoxicity of chemotherapeutic agents.[8] PCNSL is distributed along the spectrum of systemic DLBCL including the activated B-cell (ABC), the germinal center B-cell (GCB), and the non-ABC/non-GCB subtype, suggesting that PCNSL possibly shares characteristics, for example, hyperactivated signaling pathways, with systemic DLBCL.[26] Our results suggest that the mTORC1 signaling pathway is activated in not only systemic lymphomas but also “primary central nervous system” lymphoma and may cause the growth of these tumors.
Upstream markers of mTORC1 activation
In the present study, we evaluated mTORC1 activation using immunohistochemistry for Rheb, p-S6, and p-4E-BP1 because mTORC1 activity cannot be evaluated directly. Although many previous reports used expression of phospho-Akt (p-Akt) (Ser473) as an upstream marker of mTORC1 activation, recent studies have shown that there are several other upstream pathways of mTORC1, for example, mitogen-activated protein kinase signaling pathway, AMP-activated protein kinase signaling pathway, WNT signaling pathway, etc., [Figure 1].[31] It has also been reported that mTORC1 activation induces negative feedback on PI3K and Akt, and consequently reduces the level of p-Akt.[35] Rheb, the downstream effector of TSC1/TSC2, directly interacts with mTORC1 and strongly stimulates its kinase activity, and its increased expression causes mTORC1 activation in some tumors such as lymphoma.[19,23,27] Instead of evaluating all upstream signaling pathways, we used this proximal activator of mTORC1 as an upstream marker of mTORC1 activation [Figure 1]. Although GDP-bound Rheb, into which TSC converts GTP-bound Rheb, provides a less effective stimulus to mTORC1 than GTP-bound Rheb does, endogenous Rheb binds preferentially to GTP even under basal conditions.[13] Thus, increase in the expression of Rheb has the potential to activate mTORC1 and promote tumorigenesis.[21,23,27] In the present study, Rheb was highly expressed in 20 cases, in which both p-4E-BP1 and p-S6 were highly expressed in 12 cases and one of them was highly expressed in 7 cases, suggesting that overexpression of Rheb may contribute to mTORC1 activation in PCNSL.
Downstream markers of mTORC1 activation
mTORC1 directly phosphorylates translational regulators 4E-BP1 and S6K1, which, in turn, promote protein synthesis.[19] The phosphorylation of 4E-BP1 prevents its binding with the cap-binding protein eIE4, enabling eIF4E to participate in the formation of the eIF4F complex and resulting in the initiation of cap-dependent translation.[19] Consequently, activated eIF4E preferentially drives the translation of mRNA for pro-tumorigenic genes, including cell cycle regulators.[39] In contrast to 4E-BP1 relating with cell proliferation and tumorigenesis, S6 is a determinant of cell size, but is not dispensable for cancer formation.[10,29] Although S6 is phosphorylated by p-S6K after mTORC1 activation and p-S6 is commonly used as a downstream marker of mTORC1 activation, the exact role of p-S6 in tumorigenesis remains unclear. In the present study, p-4E-BP1 and p-S6 were expressed in many PCNSL cells, possibly contributing to the proliferation and hypertrophy of tumor cells of PCNSL as a result of the activation of mTORC1. In some specimens, only one downstream pathway was activated [Table 1]. Although it remains unclear what mechanisms determine selective downstream activation, the activation patterns of downstream pathways might predict what drug is effective in treatment of each case of PCNSL.
mTORC1 activation in brain tumors
mTORC1 signaling pathway is activated in several brain tumors, in particular in gliomas, and increased expression of p-4E-BP1 and/or p-S6 in the tumors has been reported to correlate with enhancing proliferation and low survival.[2,24,28] mTORC1 is also activated in pediatric low-grade glioma and pilocytic astrocytoma.[12] In neurofibromatosis-2 (NF2)-associated meningioma and schwannoma, abnormally activated mTORC1 contributes to those aberrant growth and proliferation because NF2/Merlin does not work as negative regulator of mTORC1 in them.[16] The present study showed that mTORC1 is activated in PCNSL, suggesting that dysregulated mTORC1 may contribute to proliferation and hypertrophy of tumor cells of PCNSL. The next question is how we can apply this result to clinical practice. Widely used inhibitors of mTORC1 are rapamycin and its analogs, however, mTORC1 has been reported to have rapamycin-resistant functions that are required for cell growth and proliferation and that are mediated by phosphorylation of 4E-BP1.[34] When we take advantage of the abnormal activation of mTORC1 for the treatment of PCNSL, we should use mTORC1 inhibitors that can suppress not only S6 but also 4E-BP1 phosphorylation, which may contribute to suppress tumor cell growth and proliferation and be more effective than the treatment with rapamycin and its analogs.
CONCLUSION
The mTORC1 signaling pathway is activated in PCNSL, which may contribute to proliferation and hypertrophy of tumor cells of PCNSL.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
Footnotes
Contributor Information
Naoki Nitta, Email: nnitta@belle.shiga-med.ac.jp.
Satoshi Nakasu, Email: neuro-nakasu@kfx.biglobe.ne.jp.
Ayako Shima, Email: simagoro@gmail.com.
Kazuhiko Nozaki, Email: noz@belle.shiga-med.ac.jp.
REFERENCES
- 1.Abel TW, Baker SJ, Fraser MM, Tihan T, Nelson JS, Yachnis AT, et al. Lhermitte-Duclos disease: A report of 31 cases with immunohistochemical analysis of the PTEN/AKT/mTORpathway. J Neuropathol Exp Neurol. 2005;64:341–9. doi: 10.1093/jnen/64.4.341. [DOI] [PubMed] [Google Scholar]
- 2.Annovazzi L, Mellai M, Caldera V, Valente G, Tessitore L, Schiffer D. mTOR, S6 and AKT expression in relation to proliferation and apoptosis/autophagy in glioma. Anticancer Res. 2009;29:3087–94. [PubMed] [Google Scholar]
- 3.Baselga J, Campone M, Piccart M, Burris HA, 3rd, Rugo HS, Sahmoud T, et al. Everolimus in postmenopausal hormone-receptor-positive advanced breast cancer. N Engl J Med. 2012;366:520–9. doi: 10.1056/NEJMoa1109653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chan JA, Zhang H, Roberts PS, Jozwiak S, Wieslawa G, Lewin-Kowalik J, et al. Pathogenesis of tuberous sclerosis subependymal giant cell astrocytomas: Biallelic inactivation of TSC1 or TSC2 leads to mTOR activation. J Neuropathol Exp Neurol. 2004;63:1236–42. doi: 10.1093/jnen/63.12.1236. [DOI] [PubMed] [Google Scholar]
- 5.Cho D, Signoretti S, Dabora S, Regan M, Seeley A, Mariotti M, et al. Potential histologic and molecular predictors of response to temsirolimus in patients with advanced renal cell carcinoma. Clin Genitourin Cancer. 2007;5:379–85. doi: 10.3816/CGC.2007.n.020. [DOI] [PubMed] [Google Scholar]
- 6.Choe G, Horvath S, Cloughesy TF, Crosby K, Seligson D, Palotie A, et al. Analysis of the phosphatidylinositol 3’-kinase signaling pathway in glioblastoma patients in vivo. Cancer Res. 2003;63:2742–6. [PubMed] [Google Scholar]
- 7.Crino PB, Nathanson KL, Henske EP. The tuberous sclerosis complex. N Engl J Med. 2006;355:1345–56. doi: 10.1056/NEJMra055323. [DOI] [PubMed] [Google Scholar]
- 8.Drakos E, Rassidakis GZ, Medeiros LJ. Mammalian target of rapamycin (mTOR) pathway signalling in lymphomas. Expert Rev Mol Med. 2008;10:e4. doi: 10.1017/S1462399408000586. [DOI] [PubMed] [Google Scholar]
- 9.Hoang-Xuan K, Bessell E, Bromberg J, Hottinger AF, Preusser M, Rudà R, et al. Diagnosis and treatment of primary CNS lymphoma in immunocompetent patients: Guidelines from the European Association for Neuro-Oncology. Lancet Oncol. 2015;16:e322–32. doi: 10.1016/S1470-2045(15)00076-5. [DOI] [PubMed] [Google Scholar]
- 10.Hsieh AC, Costa M, Zollo O, Davis C, Feldman ME, Testa JR, et al. Genetic dissection of the oncogenic mTOR pathway reveals druggable addiction to translational control via 4EBP-eIF4E. Cancer Cell. 2010;17:249–61. doi: 10.1016/j.ccr.2010.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hudes G, Carducci M, Tomczak P, Dutcher J, Figlin R, Kapoor A, et al. Temsirolimus, interferon alfa, or both for advanced renal-cell carcinoma. N Engl J Med. 2007;356:2271–81. doi: 10.1056/NEJMoa066838. [DOI] [PubMed] [Google Scholar]
- 12.Hütt-Cabezas M, Karajannis MA, Zagzag D, Shah S, Horkayne-Szakaly I, Rushing EJ, et al. Activation of mTORC1/mTORC2 signaling in pediatric low-grade glioma and pilocytic astrocytoma reveals mTOR as a therapeutic target. Neuro Oncol. 2013;15:1604–14. doi: 10.1093/neuonc/not132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Im E, von Lintig FC, Chen J, Zhuang S, Qui W, Chowdhury S, et al. Rheb is in a high activation state and inhibits B-Raf kinase in mammalian cells. Oncogene. 2002;21:6356–65. doi: 10.1038/sj.onc.1205792. [DOI] [PubMed] [Google Scholar]
- 14.Inoki K, Li Y, Zhu T, Wu J, Guan KL. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat Cell Biol. 2002;4:648–57. doi: 10.1038/ncb839. [DOI] [PubMed] [Google Scholar]
- 15.Iwenofu OH, Lackman RD, Staddon AP, Goodwin DG, Haupt HM, Brooks JS. Phospho-S6 ribosomal protein: A potential new predictive sarcoma marker for targeted mTOR therapy. Mod Pathol. 2008;21:231–7. doi: 10.1038/modpathol.3800995. [DOI] [PubMed] [Google Scholar]
- 16.James MF, Han S, Polizzano C, Plotkin SR, Manning BD, Stemmer-Rachamimov AO, et al. NF2/merlin is a novel negative regulator of mTOR comple×1, and activation of mTORC1 is associated with meningioma and schwannoma growth. Mol Cell Biol. 2009;29:4250–61. doi: 10.1128/MCB.01581-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jundt F, Raetzel N, Müller C, Calkhoven CF, Kley K, Mathas S, et al. A rapamycin derivative (everolimus) controls proliferation through down-regulation of truncated CCAAT enhancer binding protein {beta} and NF-{kappa} B activity in Hodgkin and anaplastic large cell lymphomas. Blood. 2005;106:1801–7. doi: 10.1182/blood-2004-11-4513. [DOI] [PubMed] [Google Scholar]
- 18.Krueger DA, Care MM, Holland K, Agricola K, Tudor C, Mangeshkar P, et al. Everolimus for subependymal giant-cell astrocytomas in tuberous sclerosis. N Engl J Med. 2010;363:1801–11. doi: 10.1056/NEJMoa1001671. [DOI] [PubMed] [Google Scholar]
- 19.Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell. 2012;149:274–93. doi: 10.1016/j.cell.2012.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liaw D, Marsh DJ, Li J, Dahia PL, Wang SI, Zheng Z, et al. Germline mutations of the PTEN gene in Cowden disease, an inherited breast and thyroid cancer syndrome. Nat Genet. 1997;16:64–7. doi: 10.1038/ng0597-64. [DOI] [PubMed] [Google Scholar]
- 21.Lu ZH, Shvartsman MB, Lee AY, Shao JM, Murray MM, Kladney RD, et al. Mammalian target of rapamycin activator RHEB is frequently overexpressed in human carcinomas and is critical and sufficient for skin epithelial carcinogenesis. Cancer Res. 2010;70:3287–98. doi: 10.1158/0008-5472.CAN-09-3467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Martineau Y, Azar R, Bousquet C, Pyronnet S. Anti-oncogenic potential of the eIF4E-binding proteins. Oncogene. 2013;32:671–7. doi: 10.1038/onc.2012.116. [DOI] [PubMed] [Google Scholar]
- 23.Mavrakis KJ, Zhu H, Silva RL, Mills JR, Teruya-Feldstein J, Lowe SW, et al. Tumorigenic activity and therapeutic inhibition of Rheb GTPase. Genes Dev. 2008;22:2178–88. doi: 10.1101/gad.1690808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.McBride SM, Perez DA, Polley MY, Vandenberg SR, Smith JS, Zheng S, et al. Activation of PI3K/mTOR pathway occurs in most adult low-grade gliomas and predicts patient survival. J Neurooncol. 2010;97:33–40. doi: 10.1007/s11060-009-0004-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McCormack FX, Inoue Y, Moss J, Singer LG, Strange C, Nakata K, et al. Efficacy and safety of sirolimus in lymphangioleiomyomatosis. N Engl J Med. 2011;364:1595–606. doi: 10.1056/NEJMoa1100391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Montesinos-Rongen M, Siebert R, Deckert M. Primary lymphoma of the central nervous system: Just DLBCL or not? Blood. 2009;113:7–10. doi: 10.1182/blood-2008-04-149005. [DOI] [PubMed] [Google Scholar]
- 27.Nardella C, Chen Z, Salmena L, Carracedo A, Alimonti A, Egia A, et al. Aberrant Rheb-mediated mTORC1 activation and Pten haploinsufficiency are cooperative oncogenic events. Genes Dev. 2008;22:2172–7. doi: 10.1101/gad.1699608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Riemenschneider MJ, Betensky RA, Pasedag SM, Louis DN. AKT activation in human glioblastomas enhances proliferation via TSC2 and S6 kinase signaling. Cancer Res. 2006;66:5618–23. doi: 10.1158/0008-5472.CAN-06-0364. [DOI] [PubMed] [Google Scholar]
- 29.Ruvinsky I, Sharon N, Lerer T, Cohen H, Stolovich-Rain M, Nir T, et al. Ribosomal protein S6 phosphorylation is a determinant of cell size and glucose homeostasis. Genes Dev. 2005;19:2199–211. doi: 10.1101/gad.351605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shaw RJ, Cantley LC. Ras, PI (3) K and mTOR signalling controls tumour cell growth. Nature. 2006;441:424–30. doi: 10.1038/nature04869. [DOI] [PubMed] [Google Scholar]
- 31.Shimobayashi M, Hall MN. Making new contacts: The mTOR network in metabolism and signalling crosstalk. Nat Rev Mol Cell Biol. 2014;15:155–62. doi: 10.1038/nrm3757. [DOI] [PubMed] [Google Scholar]
- 32.Stocker H, Radimerski T, Schindelholz B, Wittwer F, Belawat P, Daram P, et al. Rheb is an essential regulator of S6K in controlling cell growth in Drosophila. Nat Cell Biol. 2003;5:559–65. doi: 10.1038/ncb995. [DOI] [PubMed] [Google Scholar]
- 33.Thoreen CC, Chantranupong L, Keys HR, Wang T, Gray NS, Sabatini DM. A unifying model for mTORC1-mediated regulation of mRNA translation. Nature. 2012;485:109–13. doi: 10.1038/nature11083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Thoreen CC, Kang SA, Chang JW, Liu Q, Zhang J, Gao Y, et al. An ATP-competitive mammalian target of rapamycin inhibitor reveals rapamycin-resistant functions of mTORC1. J Biol Chem. 2009;284:8023–32. doi: 10.1074/jbc.M900301200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tremblay F, Marette A. Amino acid and insulin signaling via the mTOR/p70 S6 kinase pathway. A negative feedback mechanism leading to insulin resistance in skeletal muscle cells. J Biol Chem. 2001;276:38052–60. doi: 10.1074/jbc.M106703200. [DOI] [PubMed] [Google Scholar]
- 36.Wagner AJ, Malinowska-Kolodziej I, Morgan JA, Qin W, Fletcher CD, Vena N, et al. Clinical activity of mTOR inhibition with sirolimus in malignant perivascular epithelioid cell tumors: Targeting the pathogenic activation of mTORC1 in tumors. J Clin Oncol. 2010;28:835–40. doi: 10.1200/JCO.2009.25.2981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wolff N, Kabbani W, Bradley T, Raj G, Watumull L, Brugarolas J. Sirolimus and temsirolimus for epithelioid angiomyolipoma. J Clin Oncol. 2010;28:e65–8. doi: 10.1200/JCO.2009.26.3061. [DOI] [PubMed] [Google Scholar]
- 38.Yao JC, Shah MH, Ito T, Bohas CL, Wolin EM, Van Cutsem E, et al. Everolimus for advanced pancreatic neuroendocrine tumors. N Engl J Med. 2011;364:514–23. doi: 10.1056/NEJMoa1009290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zoncu R, Efeyan A, Sabatini DM. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol. 2011;12:21–35. doi: 10.1038/nrm3025. [DOI] [PMC free article] [PubMed] [Google Scholar]