

Abstract
The actions of cannabis are mediated by receptors that are part of an endogenous cannabinoid system. The endocannabinoid system (ECS) consists of the naturally occurring ligands N-arachidonoylethanolamine (anandamide) and 2-arachidonoylglycerol (2-AG), their biosynthetic and degradative enzymes, and the cannabinoid receptors CB1 and CB2. The ECS is a widely distributed transmitter system that controls gut functions peripherally and centrally. It is an important physiologic regulator of gastrointestinal motility. Polymorphisms in the gene encoding CB1 (CNR1) have been associated with some forms of irritable bowel syndrome. The ECS is involved in the control of nausea and vomiting and visceral sensation. The homeostatic role of the ECS also extends to the control of intestinal inflammation. We review the mechanisms by which the ECS links stress and visceral pain. CB1 in sensory ganglia controls visceral sensation, and transcription of CNR1 is modified through epigenetic processes under conditions of chronic stress. These processes might link stress with abdominal pain. The ECS is also involved centrally in the manifestation of stress, and endocannabinoid signaling reduces the activity of hypothalamic–pituitary–adrenal pathways via actions in specific brain regions—notably the prefrontal cortex, amygdala, and hypothalamus. Agents that modulate the ECS are in early stages of development for treatment of gastrointestinal diseases. Increasing our understanding of the ECS will greatly advance our knowledge of interactions between the brain and gut and could lead to new treatments for gastrointestinal disorders.
Keywords: CB1 receptor, CB2 receptor, fatty acid amide hydrolase, anandamide, 2-AG, enteric nervous system, HPA axis, stress, nausea, visceral pain, epigenetics
Cannabis has been used for millennia to treat the symptoms of inflammatory and functional disorders of the gastrointestinal (GI) tract1, including abdominal pain, cramps, diarrhea, nausea, and vomiting2-6. Currently, cannabis is used legally and illegally by patients seeking relief from a vast array of symptoms—many of GI origin. The availability and use of medical cannabis is increasing in the USA, Canada, and elsewhere, however, although cannabis is widely used for the treatment of nausea and abdominal pain, there is only limited evidence to support these uses7.
About 25 years ago researchers discovered the cannabinoid receptor 1 (CB1) and cannabinoid receptor 2 (CB2). Activation of these receptors accounts for the majority of the actions of the main psychoactive constituent of cannabis Δ9-tetrahydrocannabinol (THC)8, 9. The CB1 and CB2, their ligands N-arachidonoylethanolamine (anandamide) and 2-arachidonoylglycerol (2-AG), and the enzymes that synthesize and degrade them are the major components of the endocannabinoid system (ECS)9. Our knowledge of the ECS has increased exponentially in recent years and we now appreciate its roles in the brain–gut axis and gut pathophysiology. The actions of the ECS appear to be largely homeostatic, contributing importantly to the regulation of motility and inflammation in the GI tract. However, activation of CB1 in the intestinal epithelium also contributes in what appears to be a maladaptive fashion to the development of metabolic disease and obesity10. In the central nervous system (CNS), the ECS is involved in the pathophysiology of stress11, 12.
We review the fundamental properties of the ECS and describe its role in peripheral mechanisms of visceral pain, focusing on the dorsal root ganglia (DRG), which contain the cell bodies of the sensory nerves that innervate the gut. In the ganglia that control visceral sensation, the gene encoding CB1 (CNR1) is regulated epigenetically under conditions of chronic stress— this finding might provide a mechanism that links stress with visceral pain. The ECS also controls nausea and vomiting, and yet paradoxically cannabis has been associated with a hyperemesis syndrome. New findings reveal ways in which the ECS can be manipulated to alter the regulation of gut motility and inflammation. Despite the challenges of targeting the ECS therapeutically, increasing our understanding of how the ECS regulates the brain-gut axis could provide new insights into mechanisms and treatments of GI diseases.
Cannabinoids and the Endocannabinoid System
Cannabis contains dozens of different cannabinoids, terpenes, pinenes, flavonoids, and other compounds—nearly 500 different chemicals13, 14. The best known for their biological activities are THC and cannabidiol, but others, such as cannabichromene, Δ9-tetrahydrocannabivarin, cannabigerol, and cannabidivarin are being increasingly studied for a variety of actions, including those in the GI tract15, 16. The best-known receptors for these molecules include CB1 and CB2 (receptors for THC and most other cannabinoids), transient receptor potential vanilloid channels (which many cannabinoids interact with), G protein-coupled receptor 55 (GPR55, which binds some cannabinoids) and the 5-hydroxytryptamine receptor 1A (which is a receptor for cannabidiol). It is unlikely that all the receptors that mediate the actions of phytocannabinoids are known17, 18. Cannabis is active when inhaled, consumed, administered by oro-mucosal spray or rectal suppository, but smoking is the most frequent method of delivery. Cannabinoids reach their target organs in different amounts or at different times, depending on their route of administration and metabolism.
Endocannabinoids are synthesized from membrane lipids in response to specific signals—they are not stored in vesicles like most other neurotransmitters and hormones19 (Figure 1). Anandamide is normally synthesized from N-arachidonoylphosphatidylethanolamine (NAPE), which is formed by the transfer of arachidonic acid from the sn-1 position of a donor phospholipid to phosphatidylethanolamine by N–acyltransferase. Hydrolysis of NAPE by an N-acylphosphatidylethanolamine-hydrolyzing phospholipase D (NAPE-PLD) produces anandamide20. There are, however, alternative pathways of anandamide synthesis21. The principal enzyme for the degradation of anandamide is fatty acid amide hydrolase (FAAH)22. FAAH and NAPE-PLD are found in the GI tract and CNS20, 23.
Figure 1. Distribution of enzymes involved in the biosynthesis and degradation of endocannabinoids in the nervous system.
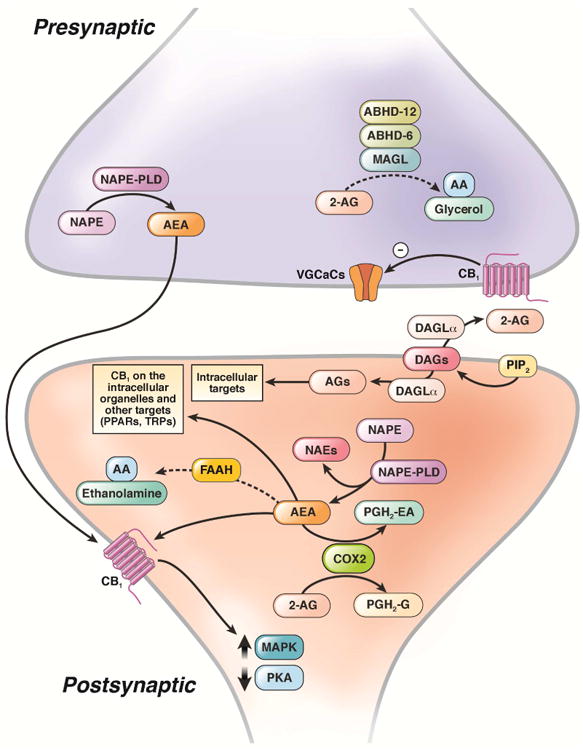
The role of 2-AG as a dominant retrograde synaptic transmitter and putative roles of anandamide (AEA) as an anterograde and intracellular transmitter. Anandamide is synthesized from NAPE by a specific phospholipase D (NAPE-PLD) and is degraded by FAAH. 2-AG is synthesized by DAGL-α and degraded by MAGL and ABHD-6 and ABHD-12. Anandamide and 2-AG can also be metabolized by COX2, which is shown in post-synaptic neurons but can also occur presynaptically. AA, arachidonic acid; AGs, 2-acylglycerols; DAGs, diacylglycerols; ER, endoplasmic reticulum; MAPK, mitogen-activated protein kinases; PIP2, phosphoinositide bisphosphate; PKA, protein kinase A; PPARs, peroxisome proliferator-activated receptors; TRPs, transient receptor potential channels; VGCCs, voltage-gated calcium channels. Dashed lines denote inactivation. Adapted from Di Marzo et al.118
Synthesis of 2-AG requires the activation of a phosphoinositol phospholipase C, which hydrolyzes inositol phospholipids at the sn-2 position to produce diacylglycerol (DAG)21. The hydrolysis of DAG via sn-1-selective diacylglycerol lipases (DAGL)-α (and potentially DAGL-β) then leads to the formation of 2-AG21. There are also other pathways for synthesis of 2-AG. 2-AG is mostly degraded by monoacylglycerol lipase (MAGL), but in mouse brain, about 15% of the degradation is by the enzymes α/β -hydrolase domain-containing protein-6 (ABHD-6) and ABHD-1224. Studies are needed to determine the relative activities of the enzymes that degrade 2-AG in the human brain and the GI tract. 2-AG can also be oxygenated by cyclooxygenase-2 (COX2) to form biologically active prostaglandin glyceryl esters, which regulate inflammation25, 26.
Endocannabinoids signal via CB1 and CB2, which are present in the GI tract. These receptors have been detected in enteric nerves and the epithelium, and CB1 has been detected on some enteroendocrine cells. CB1 appears to be the receptor that is most active under physiologic conditions3, 27. Anandamide is a natural ligand of the transient receptor potential cation channel subfamily V member 1 (TRPV1)28.
CB1 is one of the most abundant G protein-coupled receptors in the brain, highlighting its importance in the control of central neurotransmission. It is also highly expressed in the enteric nervous system, on all classes of enteric neuron except inhibitory motor neurons27. The widespread distribution of CB1 could account for the broad range of activities of cannabinoid ligands and varied effects of cannabis. However, this widespread distribution also poses a considerable challenge to therapies that target CB1—ligands can have a large variety of effects on the central and peripheral nervous systems.
CB1 is predominantly found on neurons, most abundantly in the gut. CB2 is found on enteric neurons27, and also expressed by immune and epithelial cells in the GI tract29, 30. Only a few studies have examined the role of CB2 in interactions between the brain and gut. One of the most interesting examined visceral pain. Oral administration of probiotic strains of Lactobacillus that reduced visceral sensitivity were found to upregulate CB2 (and mu opioid receptors) in the intestinal epithelium of rats. Blocking CB2 attenuates the reduction in visceral sensory thresholds caused by the alteration in the gut microbiome31. Little is known about the mechanisms of this fascinating effect.
Similarly, chronic stress and antibiotic-induced dysbiosis upregulated CNR2 mRNA (encodes CB2) in the GI tract32. Although these are interesting observations, a clinical trial of the same Lactobacillus strains that reduced visceral sensitivity in rats found that in humans, these bacteria reduced expression of CB233. However, the bacteria still appeared to reduce visceral sensation and upregulate mu opioid receptors. Studies are needed to determine whether this finding is specific only to this probiotic or whether CB2 regulates visceral sensation in humans.
A Peripheral Mechanism Linking Chronic Stress to Pain
Many patients with chronic visceral pain find that it is exacerbated by stress. There is increasing evidence that the ECS modulates chronic stress-associated increases in abdominal pain (visceral hyperalgesia)3, 34 (Figure 2).
Figure 2. Effects of chronic stress on peripheral endocannabinoid pathways in visceral primary afferent neurons.
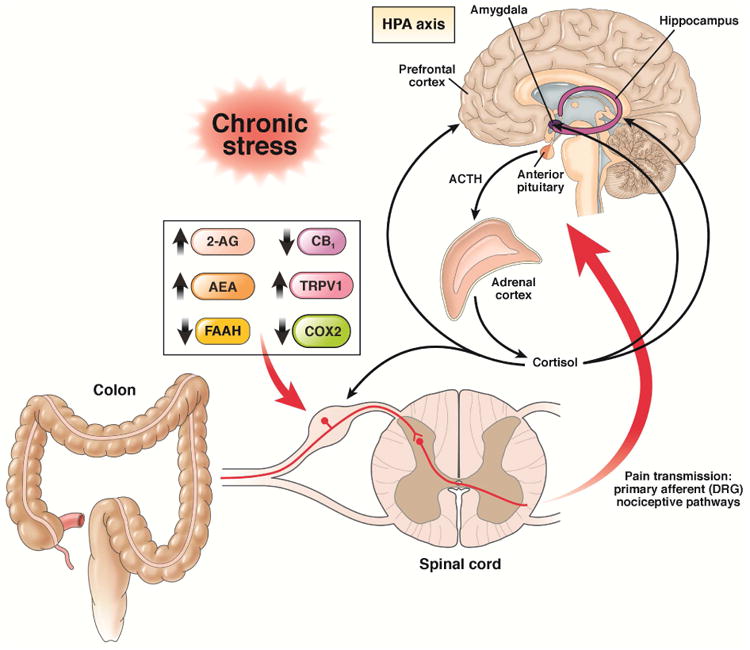
Under conditions of chronic stress, levels of 2-AG and AEA increase and endocannabinoid degradation enzymes COX-2 and FAAH are decreased in nociceptive DRG neurons that innervate the colon and pelvis. Along with this, levels of CB1 are reduced and there is an increase in TRPV1 expression and phosphorylation in nociceptive primary afferent neurons. These effects are mediated by corticosteroids from the HPA pathway.
CB1 localizes to the cell bodies of DRG neurons. Under baseline conditions, most of these are intermediate and larger-diameter neurons35, 36. There is, however, a population of small-diameter nociceptive neurons that express CB1 along with TRPV1 and contain peptides such as substance P and calcitonin gene-related peptide36, 37. When there is inflammation at their peripheral terminals, the proportion of CB1-containing neurons increases and a much higher proportion co-localize with TRPV138. Activation of CB1 inhibits pain perception (nociception), whereas activation of TRPV1 increases pain perception39. CB1 interacts with and inhibits TRPV1 directly and indirectly, via the cyclic AMP–protein kinase A pathway40.
In rats, chronic, intermittent water-avoidance stress was associated with reciprocal changes in levels of 2-AG and anandamide (increased) and endocannabinoid degradation enzymes COX2 and FAAH (decreased). In these rats, stress was also associated with a decrease in levels of CB1 and an increase in TRPV1 expression and phosphorylation in nociceptive lumbosacral primary afferent neurons innervating the colon and pelvis, but not in those innervating the lower extremities41. The reciprocal changes in CB1 and TRPV1 were reproduced by exposure of control sensory neurons with anandamide. In contrast, exposure of control DRGs in vitro with the CB1 and CB2 agonist WIN 55,212-2 decreased the levels of TRPV1 and its phosphorylation. Treatment of stressed rats with WIN 55,212-2 or the TRPV1 antagonist capsazepine prevented visceral hyperalgesia41.
Linkage of the CB1–TRPV1 pathway to hypothalamic-pituitary-adrenal (HPA) stress was confirmed in experiments demonstrating that administration of the glucocorticoid receptor antagonist RU-486 to stressed rats prevented the reciprocal changes in CB1 and TRPV1 expression and visceral hyperalgesia. Control rats given serial injections of corticosterone, which mimicked chronic stress, developed visceral hyperalgesia, had increases in anandamide content and expression of TRPV1, and decreased levels of CB1. These changes were prevented by RU-48642. Taken together these findings indicate that chronic stress induces visceral hyperalgesia that involves differential, region-specific changes in endovanilloid (an endogenous ligand for TRPV1) and endocannabinoid pathways in sensory neurons innervating the pelvic viscera. These observations could provide a link between chronic stress and visceral hyperalgesia and lead to new therapeutic approaches. They also raise the question, what sustains these chronic changes? There is evidence that these could be mediated by epigenetic changes in sensory neurons.
Epigenetic regulation of chronic stress-induced visceral hyperalgesia
Epigenetic modifications produce long-term effects on gene expression without altering the DNA sequence (Figure 3). Interest in epigenetic regulatory pathways has emerged rapidly because of their apparent significance in key physiological processes. Important epigenetic alterations include DNA methylation, catalyzed by DNA methyltransferases (DNMTs) such as DNMT1, which is responsible for the maintenance of methylation patterns, and DNMT3a and DNMT3b, which are responsible for de novo methylation patterns. Methylation typically silences genes, whereas histone acetylation, catalyzed by acetyltransferases, typically activates genes43-45. DNA methylation and histone acetylation are considered to be stable yet reversible; the enzymes that control these processes can be induced or inhibited by biochemical and environmental factors.
Figure 3. Effects of chronic stress on epigenetic regulation of the gene encoding CB1 (CNR1).
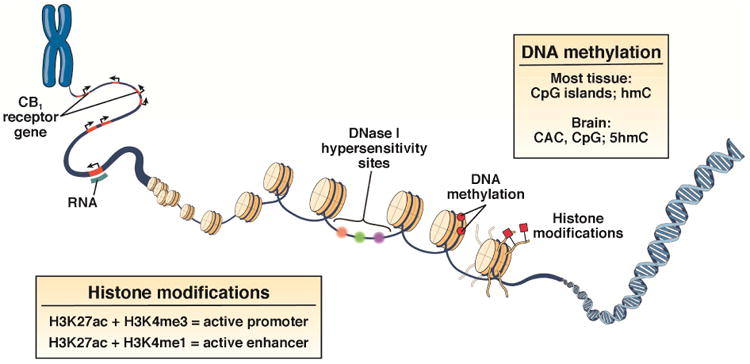
Chromosomes are located in chromatin-bound territories in the nucleus. Euchromatin is characterized by DNase 1 hypersensitivity and specific combinations of histone marks that define active genomic regulatory elements, such as promoters H3K27ac + H3K4me3, and enhancers H3K27ac + H3K4me1. An enhancer can either increase or decrease transcription. Recent research demonstrates that, in brain, the DNA sequence CAC is a common site of methylation, in contrast to other tissues where CpG is most often methylated. Additionally, in brain, 5-hydroxymethylcytosine (5hmC), a reactive species, is methylated. In contrast, in the periphery, methylcytosine (hmC) is a common site for methylation. DNA methylation is catalyzed by DNMTs. Chronic stress is associated with increased levels of DNMT1-mediated methylation of CRN1, resulting in reduced expression. H3K4me3, histone H3 trimethyl Lys4; H3K27ac, histone H3 Lys27 acetylation; H3Kme1, histone H3 monomethyl Lys4. Adapted from Wiley et al.117
Chronic stress was associated with increased methylation of the Cnr1 gene promoter by DNMT1, resulting in reduced levels of CB1 in the sensory neurons that innervate the pelvic organs, including the colon, but not neurons that innervate the lower extremities46 (Figure 3). The glucocorticoid receptor (encoded by Nr3c1) is a transcription factor that also regulates the Cnr1 gene. Chronic stress was associated with increased DMNT1-mediated methylation of the Nr3c1 promoter and reduced expression of this gene in nociceptive primary afferent neurons. Also, chronic stress increases expression of the histone acetyltransferase EP300 and promotes acetylation of histones in the Trpv1 promoter, leading to increased levels of TRPV1 in these neurons.
Region-specific knockdown of DNMT1 and EP300 gene expression in visceral primary afferent neurons innervating the pelvis reduced DNA methylation and histone acetylation, respectively, and prevented chronic stress-induced increases in visceral pain. These observations indicate that chronic stress promotes DNA methylation and down-regulation of anti-nociceptive Cnr1 and concurrent increase in histone acetylation of pro-nociceptive Trpv1 in a region- and cell-specific manner, resulting in visceral hyperalgesia.
Endocannabinoids, Chronic Stress, and Pain
The ECS is involved in the CNS response to stress and pain (Figure 4). Stress evokes changes in the levels of anandamide and 2-AG, reducing AEA and increasing 2-AG47. Chronic stress causes downregulation or loss of CB1. These changes are widespread and occur throughout the brain47. The functional consequences of this are the manifestation of the stress response, including activation of the HPA stress response and anxiety47. Increased 2-AG signaling correlates with the termination and adaptation of the activity of the HPA axis and contributes to changes in pain perception. These effects presumably occur via remaining CB1 receptors, whose sensitivity may be increased.
Figure 4. Effects of chronic stress on the endocannabinoid system in the brain.
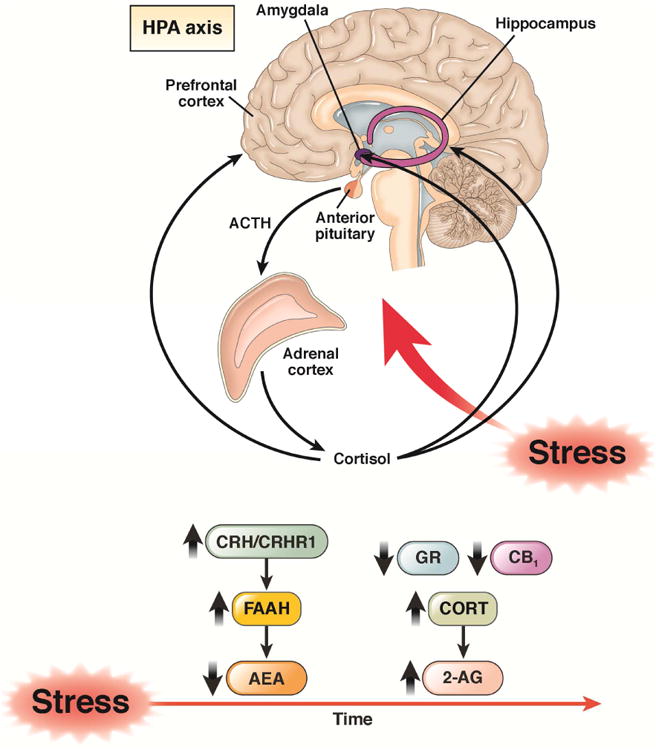
Chronic stress is characterized by a sustained reduction in levels of AEA, associated with increases in the degradative enzyme FAAH. These effects are mediated by CRH via the CRH 1 receptor (CRHR1). Corticosterone (in rodents; corticosterol in humans; CORT) appear to mediate the upregulation of CRH. The increased level of CORT are associated with increases in 2-AG. Chronic stress is associated with reduced levels of CB1 and the glucocorticoid receptor (GR) in the brain. Adapted from Morena et al.47 and Krugers et al.119
In support of the observation that endocannabinoids that act via the CB1 have an analgesic effect on pain signaling, inhibitors of endocannabinoid degradation reduce stress-related hyperalgesia48. Increasing anandamide signaling with URB597, a FAAH inhibitor, reduced chronic stress-induced anxiety and thermal hyperalgesia in mice. Endocannabinoid modulation by FAAH and MAGL also act on the analgesic circuitry in the periaqueductal grey (PAG) region of the brainstem to promote analgesia49. The PAG is activated in response to noxious visceral stimulation50 and contributes to the descending inhibitory neural pathways that gate peripheral sensory input to the CNS. The individual or combined inhibition of FAAH and/or MAGL increased tonic disinhibition within the PAG—an effect that is consistent with the promotion of analgesia. Supporting the concept that the combined inhibitor increases efficacy, the dual inhibitor of FAAH and MAGL (JZL195) was more effective in reducing allodynia (central pain sensitization following painful, often repetitive, stimulation) than selective FAAH or MAGL inhibitors. The dual inhibitor also had a larger therapeutic window than cannabinoid receptor agonists in a mouse model of neuropathic pain 51 or in animal models of visceral pain52.
The endocannabinoid system appears to be sensitive to early-life stress, in a sex- and region-specific manner53. Early-life stress is an important factor in the development of irritable bowel syndrome (IBS) and is associated with epigenetic changes that lead to visceral hypersensitivity54. Male and female Wistar rats raised under standard conditions or exposed to maternal deprivation demonstrated changes in the ECS. Specifically, expression of genes in the cannabinoid system (Cnr1, Cnr2a, Cnr2b and GPR55, which encode enzymes involved in synthesis and degradation of endocannabinoids) were measured in adolescent rats in different regions of brain (frontal cortex, ventral and dorsal striatum, dorsal hippocampus, and amygdala). Maternal deprivation increased the expression of all the endocannabinoid genes surveyed in the frontal cortex in male rats, whereas in female rats, increased expression was noted only in the hippocampus. These observations have potentially important implications on the differential sex-specific effects stress on pain perception and the mechanisms by which early-life affects these responses.
Diurnal changes are a feature of visceral sensation and correlate with changes in the HPA axis55, which is dysregulated in patients with IBS56. The ECS regulates basal and circadian HPA axis activation. For example, the hypothalamic content of anandamide is highest at the times of 07:00 and 11:00 and low between 15:00 and 03:0057. In vivo studies demonstrate that low doses of CB1 agonists other than THC reduced basal and stress-induced HPA pathway responses in rodents58, 59. In persons who were either naıve to cannabis or infrequent users, acute consumption of cannabis60 or THC61, 62 increased secretion of cortisol. The stimulatory effect of THC administration on cortisol levels was lower in chronic cannabis users, supporting the development of tolerance 63, 64. Some studies65, 66 found that chronic cannabis users had higher basal levels of cortisol than infrequent users. Stress-induced activation of the HPA stress response was reported to be reduced in adult and adolescent chronic users of cannabis66, 67. Taken together, these observations indicate that chronic cannabis use might dysregulate basal, circadian, and stress-regulated HPA stress responses in a complex manner.
The ECS suppresses the activity of the HPA pathway via distinct actions within the prefrontal cortex, amygdala, hypothalamus, and hippocampus12. The region-specific action of the ECS in the CNS could account for the observed effects of cannabinoids on anxiety, appetite, nausea, and pain.
Endocannabinoids in the Control of Nausea and Vomiting
Cannabis is an effective anti-emetic and is used in the clinic, although it is not a first-line therapeutic agent4. However, unlike most commonly used anti-emetics, cannabis also prevents or limits the sensation of nausea. The brainstem circuitry of vomiting is well established4. The ECS is involved in the control of the emetic reflex, as shown by the fact that antagonists of CB1 cause vomiting in humans and exacerbate it animals. Interestingly, not only do CB1 agonists block emesis, but so do exogenous and endogenous agonists of CB268. Since CB2 activation is not psychotropic, this observation potentially opens up new avenues for therapeutic interventions.
Unlike vomiting, the brain circuitry responsible for evoking nausea is incompletely understood, but is being elucidated through the use of functional magnetic resonance imaging in humans and through the use of animal models4, 69, 70. These studies have associated nausea with activation of many complex neural circuits, involving brainstem, limbic, interoceptive, somatosensory, and cognitive neural networks69, 70. In a rat model of nausea, when levels of endocannabinoid were increased systemically by administration of a FAAH inhibitor, nausea was reduced. However, this effect is not mediated by CB171, 72, since it was not blocked by an antagonist. In contrast, inhibition of MAGL, which increases levels of 2-AG, is mediated by CB1. Studies of humans and animals have found the insular cortex to be involved in the perception of nausea (human studies) and physiologic responses to nausea (animal studies)69, 70, 73. Activation of the ECS in the insular cortex can regulate the expression of nausea in animals; interestingly, it appears that 2-AG, rather than anandamide, mediates this regulatory control system71, 74 (Figure 5). Thus MAGL inhibitors, but not FAAH inhibitors, when administered into the visceral insular cortex, block nausea in animals71, 74. Further studies are needed to elucidate how the selectivity occurs in this system, when both endocannabinoid ligands are agonists of CB1, but the process could involve locations of the receptors and/or metabolism of endocannabinoids in this brain region. It will also be important to determine whether agents that alter the ECS system, beyond cannabis, can be developed for treatment of patients with nausea.
Figure 5. Role of endocannabinoids in control of nausea and vomiting.
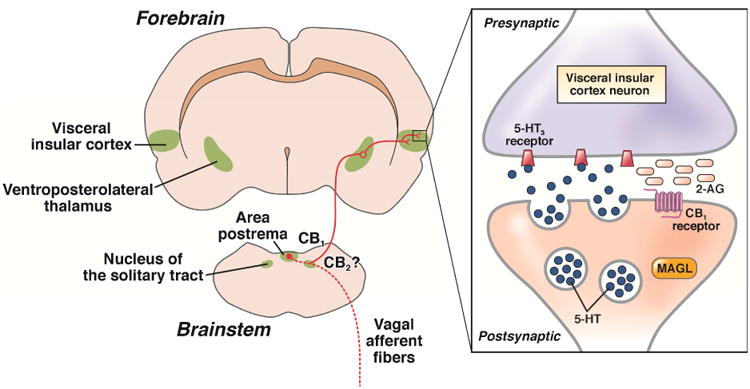
In the brainstem regions involved in the control of vomiting (area postrema and nucleus of the solitary tract), activation of CB1 and possibly CB2 attenuates the emetic reflex by reducing the release of excitatory transmitters. In the forebrain, release of 2-AG in the visceral insular cortex inhibits, in a retrograde manner, release of neurotransmitters including serotonin (5-HT), which acts at 5-HT3 receptors to produce nausea. Note that to date this model has been studied in the rat and that in humans the CB1 agonist Δ9-tetrahydrocannabinol reduces nausea. 5-HT3 receptor antagonists are excellent antiemetics but are less effective at reducing nausea. Adapted from Sticht et al.120
The cannabinoid hyperemesis syndrome (CHS) was first described in 200475. CHS is a syndrome of cyclic episodes of nausea, vomiting, and abdominal pain associated with chronic cannabis usage. CHS is frequently confused with cyclic vomiting syndrome, which has similar features76. An interesting and poorly understood feature of the syndrome is that patients report reduced symptoms following hot baths or showers75, 77, 78. Typically, standard anti-emetics are ineffective in treating CHS79.
Little is known about the mechanism(s) of this syndrome—they might involve toxic metabolite(s) from the cannabis plant or progressive high exposure to the ligand may lead to down-regulation of cannabinoid receptors and loss of the endocannabinoid anti-emetic pathway4. CHS has not been reported in patients taking only a pure THC product such as Marinol on a scheduled regimen. Genetic factors might also affect susceptibility to CHS. Patients who experience hyperemesis from chronic cannabis use may have a genetic variation in their hepatic drug-transforming enzymes that results in excessive levels of cannabis metabolites that promote emesis80. The cutaneous (cutaneous steal) and splanchnic (vasodilation) circulations have been proposed as contributing factors to abdominal pain and nausea81. Currently, the only effective treatment of CHS is a drug holiday—cessation of cannabis use to allow clearance of putative toxic metabolites and/or normalization of CB receptor levels and function.
Cannabinoids and Control of GI Motility
Cannabis reduces cramping and slows GI motility. These actions prompted detailed studies of the role of the ECS in the control of motility and specifically in IBS (Figure 6). Variants of the CNR1 and FAAH genes have been observed in patients with diarrhea-predominant and alternating forms of IBS82-84. Levels of FAAH mRNA are reduced in intestinal tissues of patients with constipation-predominant IBS85. These genetic polymorphisms and alterations in gene expression are associated with alterations in GI motility and sensation, supporting the pathophysiologic significance of alterations in the ECS in the gut.
Figure 6. Effects of endocannabinoids in the GI tract are mediated by CB1 and CB2.
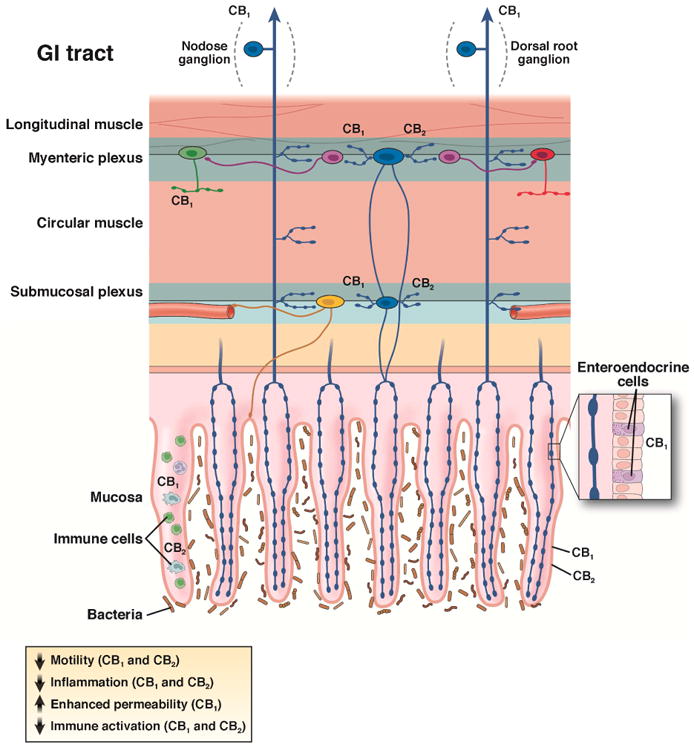
Cannabinoid receptors are widely distributed in the GI tract. CB1 is expressed by all classes of cholinergic enteric neurons in the myenteric and submucosal plexuses (primary afferent neurons (blue), interneurons (purple), secretomotor (yellow), and excitatory motor neurons (green), but not on inhibitory motor neurons (red). CB1 is also found on some enteroendocrine cells and in the epithelium. Extrinsic vagal and spinal primary afferent neurons express CB1, which is regulated by feeding (vagal) and stress (spinal), respectively. CB2 is expressed by enteric neurons and immune cells in the GI tract. Under conditions of inflammation and injury, CB2 is upregulated in the epithelium and its function on enteric neurons increases. The classes of enteric neurons that express CB2 have not been determined. The effects of activating cannabinoid receptors in the GI tract include a reduction in motility, reduced inflammation, and reduced immune activation. Adapted from Lomax et al.121
In mice, disruption of Cnr1 increases motility. Consistent with this, patients given CB1 antagonists report diarrhea3. Remarkably, disruption of Cnr1 specifically in the vagal system (afferent and efferent neurons) also increased GI motility86, but it is not clear whether this was due only to changes in gastric emptying. Nonetheless, the ECS in the gut appears to be a potential therapeutic target for IBS and other functional bowel disorders.
Researchers have studied whether peripherally restricted cannabinoids or locally produced endocannabinoids alters gut motility. Potent agonists of CB1 without central actions have been examined in rats and mice with increased GI motility caused by stress, as well as in control animals87-89. Interestingly, agonists were more potent in stressed than control mice, and extremely low doses (which had no effects in control mice) were found to normalize accelerated small- and large-intestinal motility87. There is no clear explanation for these observations, but possibilities include either an increase numbers or affinity of CB1, or the coupling of the second messenger systems to the effector responses.
Naturally released endocannabinoids might potentially be used to control GI motility. Drugs that increase or reduce the local levels of endocannabinoids, which would have activities in only the specific areas where they are produced, could theoretically have limited off-target actions. Proof of concept for this idea come from studies using DAGL and FAAH inhibitors. Inhibitors of DAGL have been examined in models of constipation; they effectively normalized slowed transit in opioid-induced constipation and in congenital slow-transit constipation90. Conversely, inhibiting the degradation of endocannabinoids, by blocking FAAH, has antidiarrheal and anti-nociceptive effects in models of accelerated motility and in mice with oil of mustard-induced colitis, respectively91, 92. In these studies, increasing levels of endocannabinoids in the gut was not associated with cannabinoid-like side effects such as reduced ambulatory locomotion. However, it should be noted that these examples are both acute studies—chronic administration can have negative effects because of the physiological role of the ECS. For example, inhibiting FAAH may promote cardiovascular injury because of the actions of the ECS in the heart including atherosclerotic plaque formation and myocardial tissue injury93-95. Nonetheless, peripherally restricted cannabinoids and modulators of endocannabinoid synthesis and/or degradation might be used for short-term treatment of symptoms of IBS and other functional bowel disorders.
Regulation of Intestinal Inflammation by Cannabinoids and Fatty Acid Ethanolamides
The homeostatic role of the also ECS extends to the control of intestinal inflammation (Figure 6). Although exogenous cannabinoids can reduce colitis, one therapeutic strategy has been to increase levels of endocannabinoids by inhibiting their degradation. Inhibiting FAAH, which increases the levels of anandamide and other fatty acid ethanolamides, blocks the development of colitis in mouse models of inflammatory bowel disease96-98. Evidence in support of the homeostatic role of this system comes from studies of FAAH-knockout mice, which develop a significantly milder colitis than control mice96. The anti-inflammatory effects of FAAH inhibition are mediated by CB1 and CB2. Inhibitors of MAGL also block the development of colitis in mice99, but more extensive investigations of the homeostatic role of MAGL are required.
These studies raised the question of whether colitis develops, in part, through the loss of this endogenous anti-inflammatory system. Although polymorphisms in FAAH have not been directly associated with risk for inflammatory bowel disese100, patients with Crohn's disease homozygous for the mutation in FAAH encoding the amino acid change Pro129Thr were more likely to develop severe disease associated with fistulae and extra-intestinal manifestations. Patients with ulcerative colitis who are homozygous for this FAAH variant had earlier onset of disease than patients without this variant100. Patients with acute ulcerative colitis have an increase in FAAH-reactive immune cells in the lamina propria23, but FAAH expression has been reported to be increased or reduced in different studies98, 101. Similarly, in patients with colitis, levels of anandamide have also been reported to be either increased or reduced102, 103. Hence further studies are needed to reconcile these disparate findings.
Recently, a dual FAAH and COX inhibitor was evaluated in mice with colitis104. This interesting compound reduced the severity of colitis by increasing levels of anandamide, which acted on CB1, and also increased levels of the fatty acid ethanolamides palmitoylethanolamide (PEA) and oleoylethanolamide, which act via peroxisome proliferator-activated receptor-α. Levels of PEA are increased in intestinal tissues of patients with ulcerative colitis105. PEA is a powerful anti-inflammatory agent that reduces features of colitis in mice and secretion of inflammatory cytokines106. PEA binds to peroxisome proliferator-activated receptor-α and inhibits the expression of S100B and toll-like receptor 4 on enteric glia, to reduce inflammation induced by nuclear factor-κB107. This is a complex system, because PEA, like anandamide, affects enteric glia and neurons in the gut, via different receptors on each cell type. PEA is not only degraded by FAAH but also N-acylethanolamine–hydrolyzing acid amidase108. An N-acylethanolamine–hydrolyzing acid amidase inhibitor was shown to markedly reduce colitis, providing yet another therapeutic target for study108.
Although our discussion has focused on the GI tract, administration of the cannabinoid agonist CB13 to the CNS was reported to block the development of colitis109. The mechanism of this process has not been elucidated, but this remarkable finding indicates the importance of central and peripheral CB receptors in modulating gut functions and inflammation.
Clinical Application of ECS Manipulation
The significant role of the ECS in the GI tract makes it an appealing therapeutic target. However, the widespread distribution of the ECS and its extensive activities throughout the body8 make it a challenge to selectively alter in the gut, without affecting other organ systems. Agonists of CB1 and CB2 (such as dronabinol and nabilone) are effective in treatment of nausea and vomiting and stimulating appetite, but are limited by their central side effects. Clinical studies using other CB receptor ligands have also proved disappointing. CB1 antagonists were thought to be promising for the treatment of obesity and metabolic syndrome110, 111, but were withdrawn from the market after some patients developed serious psychiatric side effects. Peripherally restricted agonist of CB1 and CB2 were not found to be effective in patients with pain, and produced CNS and cardiovascular side effects112. A selective agonist of CB2 was also examined for its analgesic potential and found to be rather ineffective, though well tolerated113.
FAAH inhibitors appeared to be promising in preclinical studies, but human studies did not demonstrate efficacy, although these agents increased levels of anandamide in the target tissues114. Findings from preclinical studies have indicated that inhibitors of FAAH can promote cardiovascular injury, because of the role of the ECS in the heart93-95, raising significant concerns about their chronic use, even if they are found to be effective. Most of these adverse effects are due to activation of CB1, as CB2 seems to protect the heart 15.
These studies paint a somewhat bleak picture for translation of promising preclinical findings to the clinic. Because the ECS is of such widespread importance, any systemic alterations involving the ECS will almost certainly produce challenges related to the drug target. Findings from clinical studies remind us that that is important to increase our understanding of the role of the ECS in maintenance of health and development of disease, and to better appreciate how this system interacts with other key regulatory systems.
However, based on what is known about the role of the ECS in the control of the GI tract in health and disease there are a number of important directions to consider in the development of therapeutic strategies to modulate the ECS. First, peripherally restricted CB1 antagonists are likely to increase gut motility and possibly reduce intestinal permeability—features that could make them useful therapeutics, especially given their beneficial effects in patients with diabetes116. Agonists of CB2 could reduce inflammation in the GI tract and limit abnormally accelerated motility. Another approach could be to use cannabinoid ligands that are not absorbed and act only on the epithelium31.
Future Directions
The ECS is an important regulator of intestinal function and the brain-gut axis. It is a widely distributed homeostatic system that generally inhibits neural activity in pathways involved in the physiological regulation of the GI tract. Important gut functions including visceral sensation, pain, motility, and inflammation are regulated by the ECS. However, there are many areas where we lack a detailed understanding of the distribution and function of the various components of the ECS, notably the biosynthetic enzymes for anandamide and 2-AG.
Genetic and/or epigenetic alterations that affect activities of the ECS could be involved in the pathogenesis of IBS. As we have outlined, alterations of the ECS may provide a link between stress and visceral pain, a well-recognized feature of many GI conditions. Inflammation of the gut alters the expression of degradative enzymes of the ECS, leading to marked changes in the local levels of these pleiotropic lipid mediators. Far more work is required to better understand the involvement of the ECS in GI diseases.
The glucocorticoid receptor regulates expression of CNR1, and chronic stress is associated with increased DMNT1-mediated methylation at the promoters of genes that encode the glucocorticoid receptor (NR3C1) and CB1 (CNR1), resulting in decreased expression of the glucocorticoid receptor and CB1. Chronic stress-induced changes in this pathway are region- and cell-specific and might be targeted by therapeutic agents. Chronic stress and chronic cannabis use affect ECS regulation of the circadian HPA pathway, so chronic stress may affect potential circadian and ultradian transcriptional regulation of CNR1 in a region- and cell-specific manner. Cortisol (human) and corticosterone (rodent) have been shown to promote circadian and ultradian bursts of transcriptional activity in many genes, including the CLOCK gene family; this rhythm is disrupted in patients with stress-related mental health, GI, or immune disorders117. Epigenetic factors determine how the glucocorticoid receptor interacts with target genes, including CNR1. Dysregulation of the central and peripheral the glucocorticoid receptor regulome has potentially significant consequences for stress-related disorders that affect the brain-gut axis and ECS.
Cannabinoids are prescribed for GI conditions, particularly nausea and vomiting, as an appetite stimulant and for the treatment of visceral pain. There is evidence that direct or indirect activation of CB1, and possibly CB2, inhibits visceral sensitivity and pain in rodents. Central side effects such as drowsiness and heightened awareness have been reported. Recently developed ligands of CB2 and peripherally acting ligands of CB1 and CB2 have the potential to alleviate GI symptoms without CNS side effects. Future research will focus on the development of compounds that either act directly at the CB receptors or alter ligand availability in a region-specific manner to minimize side effects on brain function. FAAH, MAGL, and N-acylethanolamine–hydrolyzing acid amidase appear to be promising therapeutic targets based on findings from animal studies, but their widespread physiologic functions could limit our ability to alter their activity for treatment of chronic GI conditions.
With the increasing use of cannabis in Western society, there is a need to better understand the effects of the many components of this complex plant on gut functions in health and disease. A full understanding of the ECS will advance our knowledge of the brain–gut axis and could lead to new avenues for the treatment of many GI disorders.
Acknowledgments
KAS is the Crohn's Colitis Canada Chair in Inflammatory Bowel Disease Research at the University of Calgary. KAS receives research funding from the Canadian Institutes of Health Research. JWW receives research funding from the NIH 5R01DK098205 and Takeda, Pharmaceuticals.
Abbreviations
- 2-AG
2-arachidonoylglycerol
- ABHD
α/β -hydrolase domain-containing protein
- AEA
anandamide
- CB
cannabinoid
- CHS
cannabis hyperemesis syndrome
- CNS
central nervous system
- COX
cyclooxygenase
- DAG
diacylglycerol
- DAGL
diacylglycerol lipase
- DNMT
DNA methyltransferase
- DRG
dorsal root ganglia
- ECS
endocannabinoid system
- FAAH
fatty acid amide hydrolase
- GI
gastrointestinal
- HPA
hypothalamic-pituitary-adrenal
- MAGL
monoacylglycerol lipase
- NAPE
N-arachidonoylphosphatidylethanolamine
- NAPE-PLD
N-acylphosphatidylethanolamine-hydrolyzing phospholipase D
- PAG
periaqueductal grey
- PEA
palmitoylethanolamide
- THC
Δ9-tetrahydrocannabinol
- TRPV
transient receptor potential vanilloid
Footnotes
Disclosures: KAS is a consultant for FAAH Pharma. JWW has no conflicts of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Grinspoon L, Bakalar JB. Marihuana, the forbidden medicine. New Haven: Yale University Press; 1997. [Google Scholar]
- 2.Aviello G, Romano B, Izzo AA. Cannabinoids and gastrointestinal motility: animal and human studies. Eur Rev Med Pharmacol Sci. 2008;12(1):81–93. [PubMed] [Google Scholar]
- 3.Izzo AA, Sharkey KA. Cannabinoids and the gut: new developments and emerging concepts. Pharmacol Ther. 2010;126:21–38. doi: 10.1016/j.pharmthera.2009.12.005. [DOI] [PubMed] [Google Scholar]
- 4.Sharkey KA, Darmani NA, Parker LA. Regulation of nausea and vomiting by cannabinoids and the endocannabinoid system. Eur J Pharmacol. 2014;722:134–46. doi: 10.1016/j.ejphar.2013.09.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Abalo R, Vera G, Lopez-Perez AE, et al. The gastrointestinal pharmacology of cannabinoids: focus on motility. Pharmacology. 2012;90:1–10. doi: 10.1159/000339072. [DOI] [PubMed] [Google Scholar]
- 6.Alhouayek M, Muccioli GG. The endocannabinoid system in inflammatory bowel diseases: from pathophysiology to therapeutic opportunity. Trends Mol Med. 2012;18:615–25. doi: 10.1016/j.molmed.2012.07.009. [DOI] [PubMed] [Google Scholar]
- 7.Whiting PF, Wolff RF, Deshpande S, et al. Cannabinoids for Medical Use: A Systematic Review and Meta-analysis. JAMA. 2015;313:2456–73. doi: 10.1001/jama.2015.6358. [DOI] [PubMed] [Google Scholar]
- 8.Maccarrone M, Bab I, Biro T, et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol Sci. 2015;36:277–96. doi: 10.1016/j.tips.2015.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Di Marzo V, De Petrocellis L. Why do cannabinoid receptors have more than one endogenous ligand? Philos Trans R Soc Lond B Biol Sci. 2012;367:3216–28. doi: 10.1098/rstb.2011.0382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cani PD. Crosstalk between the gut microbiota and the endocannabinoid system: impact on the gut barrier function and the adipose tissue. Clin Microbiol Infect. 2012;18(4):50–3. doi: 10.1111/j.1469-0691.2012.03866.x. [DOI] [PubMed] [Google Scholar]
- 11.Hill MN, Patel S, Campolongo P, et al. Functional interactions between stress and the endocannabinoid system: from synaptic signaling to behavioral output. J Neurosci. 2010;30:14980–6. doi: 10.1523/JNEUROSCI.4283-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hill MN, Tasker JG. Endocannabinoid signaling, glucocorticoid-mediated negative feedback, and regulation of the hypothalamic-pituitary-adrenal axis. Neuroscience. 2012;204:5–16. doi: 10.1016/j.neuroscience.2011.12.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Russo EB. Taming THC: potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br J Pharmacol. 2011;163:1344–64. doi: 10.1111/j.1476-5381.2011.01238.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gertsch J, Pertwee RG, Di Marzo V. Phytocannabinoids beyond the Cannabis plant - do they exist? Br J Pharmacol. 2010;160:523–9. doi: 10.1111/j.1476-5381.2010.00745.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.De Petrocellis L, Orlando P, Moriello AS, et al. Cannabinoid actions at TRPV channels: effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol (Oxf) 2012;204:255–66. doi: 10.1111/j.1748-1716.2011.02338.x. [DOI] [PubMed] [Google Scholar]
- 16.Izzo AA, Capasso R, Aviello G, et al. Inhibitory effect of cannabichromene, a major non-psychotropic cannabinoid extracted from Cannabis sativa, on inflammation-induced hypermotility in mice. Br J Pharmacol. 2012;166:1444–60. doi: 10.1111/j.1476-5381.2012.01879.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hill AJ, Williams CM, Whalley BJ, et al. Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacol Ther. 2012;133:79–97. doi: 10.1016/j.pharmthera.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 18.Di Marzo V, Piscitelli F. The Endocannabinoid System and its Modulation by Phytocannabinoids. Neurotherapeutics. 2015;12:692–8. doi: 10.1007/s13311-015-0374-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Katona I, Freund TF. Multiple functions of endocannabinoid signaling in the brain. Annu Rev Neurosci. 2012;35:529–58. doi: 10.1146/annurev-neuro-062111-150420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Di Marzo V, Fontana A, Cadas H, et al. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature. 1994;372:686–91. doi: 10.1038/372686a0. [DOI] [PubMed] [Google Scholar]
- 21.Blankman JL, Cravatt BF. Chemical probes of endocannabinoid metabolism. Pharmacol Rev. 2013;65:849–71. doi: 10.1124/pr.112.006387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Deutsch DG, Ueda N, Yamamoto S. The fatty acid amide hydrolase (FAAH) Prostaglandins Leukot Essent Fatty Acids. 2002;66:201–10. doi: 10.1054/plef.2001.0358. [DOI] [PubMed] [Google Scholar]
- 23.Marquez L, Suarez J, Iglesias M, et al. Ulcerative colitis induces changes on the expression of the endocannabinoid system in the human colonic tissue. PLoS One. 2009;4:e6893. doi: 10.1371/journal.pone.0006893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Blankman JL, Simon GM, Cravatt BF. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem Biol. 2007;14:1347–56. doi: 10.1016/j.chembiol.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hermanson DJ, Gamble-George JC, Marnett LJ, et al. Substrate-selective COX-2 inhibition as a novel strategy for therapeutic endocannabinoid augmentation. Trends Pharmacol Sci. 2014;35:358–67. doi: 10.1016/j.tips.2014.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alhouayek M, Muccioli GG. COX-2-derived endocannabinoid metabolites as novel inflammatory mediators. Trends Pharmacol Sci. 2014;35:284–92. doi: 10.1016/j.tips.2014.03.001. [DOI] [PubMed] [Google Scholar]
- 27.Trautmann SM, Sharkey KA. The Endocannabinoid System and Its Role in Regulating the Intrinsic Neural Circuitry of the Gastrointestinal Tract. Int Rev Neurobiol. 2015;125:85–126. doi: 10.1016/bs.irn.2015.10.002. [DOI] [PubMed] [Google Scholar]
- 28.Starowicz K, Nigam S, Di Marzo V. Biochemistry and pharmacology of endovanilloids. Pharmacol Ther. 2007;114:13–33. doi: 10.1016/j.pharmthera.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 29.Wright K, Rooney N, Feeney M, et al. Differential expression of cannabinoid receptors in the human colon: cannabinoids promote epithelial wound healing. Gastroenterology. 2005;129:437–53. doi: 10.1016/j.gastro.2005.05.026. [DOI] [PubMed] [Google Scholar]
- 30.Wright KL, Duncan M, Sharkey KA. Cannabinoid CB2 receptors in the gastrointestinal tract: a regulatory system in states of inflammation. Br J Pharmacol. 2008;153:263–70. doi: 10.1038/sj.bjp.0707486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rousseaux C, Thuru X, Gelot A, et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med. 2007;13:35–7. doi: 10.1038/nm1521. [DOI] [PubMed] [Google Scholar]
- 32.Aguilera M, Vergara P, Martinez V. Stress and antibiotics alter luminal and wall-adhered microbiota and enhance the local expression of visceral sensory-related systems in mice. Neurogastroenterol Motil. 2013;25:e515–29. doi: 10.1111/nmo.12154. [DOI] [PubMed] [Google Scholar]
- 33.Ringel-Kulka T, Goldsmith JR, Carroll IM, et al. Lactobacillus acidophilus NCFM affects colonic mucosal opioid receptor expression in patients with functional abdominal pain - a randomised clinical study. Aliment Pharmacol Ther. 2014;40:200–7. doi: 10.1111/apt.12800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Storr MA, Sharkey KA. The endocannabinoid system and gut-brain signalling. Curr Opin Pharmacol. 2007;7:575–82. doi: 10.1016/j.coph.2007.08.008. [DOI] [PubMed] [Google Scholar]
- 35.Hohmann AG, Herkenham M. Localization of central cannabinoid CB1 receptor messenger RNA in neuronal subpopulations of rat dorsal root ganglia: a double-label in situ hybridization study. Neuroscience. 1999;90:923–31. doi: 10.1016/s0306-4522(98)00524-7. [DOI] [PubMed] [Google Scholar]
- 36.Bridges D, Rice AS, Egertova M, et al. Localisation of cannabinoid receptor 1 in rat dorsal root ganglion using in situ hybridisation and immunohistochemistry. Neuroscience. 2003;119:803–12. doi: 10.1016/s0306-4522(03)00200-8. [DOI] [PubMed] [Google Scholar]
- 37.Ahluwalia J, Urban L, Capogna M, et al. Cannabinoid 1 receptors are expressed in nociceptive primary sensory neurons. Neuroscience. 2000;100:685–8. doi: 10.1016/s0306-4522(00)00389-4. [DOI] [PubMed] [Google Scholar]
- 38.Amaya F, Shimosato G, Kawasaki Y, et al. Induction of CB1 cannabinoid receptor by inflammation in primary afferent neurons facilitates antihyperalgesic effect of peripheral CB1 agonist. Pain. 2006;124:175–83. doi: 10.1016/j.pain.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 39.Malik Z, Baik D, Schey R. The role of cannabinoids in regulation of nausea and vomiting, and visceral pain. Curr Gastroenterol Rep. 2015;17:429. doi: 10.1007/s11894-015-0429-1. [DOI] [PubMed] [Google Scholar]
- 40.Turcotte D, Le Dorze JA, Esfahani F, et al. Examining the roles of cannabinoids in pain and other therapeutic indications: a review. Expert Opin Pharmacother. 2010;11:17–31. doi: 10.1517/14656560903413534. [DOI] [PubMed] [Google Scholar]
- 41.Hong S, Fan J, Kemmerer ES, et al. Reciprocal changes in vanilloid (TRPV1) and endocannabinoid (CB1) receptors contribute to visceral hyperalgesia in the water avoidance stressed rat. Gut. 2009;58:202–10. doi: 10.1136/gut.2008.157594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hong S, Zheng G, Wu X, et al. Corticosterone mediates reciprocal changes in CB 1 and TRPV1 receptors in primary sensory neurons in the chronically stressed rat. Gastroenterology. 2011;140:627–637 e4. doi: 10.1053/j.gastro.2010.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bestor TH. The DNA methyltransferases of mammals. Hum Mol Genet. 2000;9:2395–402. doi: 10.1093/hmg/9.16.2395. [DOI] [PubMed] [Google Scholar]
- 44.Miranda TB, Jones PA. DNA methylation: the nuts and bolts of repression. J Cell Physiol. 2007;213:384–90. doi: 10.1002/jcp.21224. [DOI] [PubMed] [Google Scholar]
- 45.Karlic R, Chung HR, Lasserre J, et al. Histone modification levels are predictive for gene expression. Proc Natl Acad Sci U S A. 2010;107:2926–31. doi: 10.1073/pnas.0909344107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hong S, Zheng G, Wiley JW. Epigenetic regulation of genes that modulate chronic stress-induced visceral pain in the peripheral nervous system. Gastroenterology. 2015;148:148–157 e7. doi: 10.1053/j.gastro.2014.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Morena M, Patel S, Bains JS, et al. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology. 2016;41:80–102. doi: 10.1038/npp.2015.166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lomazzo E, Bindila L, Remmers F, et al. Therapeutic potential of inhibitors of endocannabinoid degradation for the treatment of stress-related hyperalgesia in an animal model of chronic pain. Neuropsychopharmacology. 2015;40:488–501. doi: 10.1038/npp.2014.198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lau BK, Drew GM, Mitchell VA, et al. Endocannabinoid modulation by FAAH and monoacylglycerol lipase within the analgesic circuitry of the periaqueductal grey. Br J Pharmacol. 2014;171:5225–36. doi: 10.1111/bph.12839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang Z, Bradesi S, Maarek JM, et al. Regional brain activation in conscious, nonrestrained rats in response to noxious visceral stimulation. Pain. 2008;138:233–43. doi: 10.1016/j.pain.2008.04.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Adamson Barnes NS, Mitchell VA, Kazantzis NP, et al. Actions of the dual FAAH/MAGL inhibitor JZL195 in a murine neuropathic pain model. Br J Pharmacol. 2016;173:77–87. doi: 10.1111/bph.13337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sakin YS, Dogrul A, Ilkaya F, et al. The effect of FAAH, MAGL, and Dual FAAH/MAGL inhibition on inflammatory and colorectal distension-induced visceral pain models in Rodents. Neurogastroenterol Motil. 2015;27:936–44. doi: 10.1111/nmo.12563. [DOI] [PubMed] [Google Scholar]
- 53.Marco EM, Echeverry-Alzate V, Lopez-Moreno JA, et al. Consequences of early life stress on the expression of endocannabinoid-related genes in the rat brain. Behav Pharmacol. 2014;25:547–56. doi: 10.1097/FBP.0000000000000068. [DOI] [PubMed] [Google Scholar]
- 54.Moloney RD, Stilling RM, Dinan TG, et al. Early-life stress-induced visceral hypersensitivity and anxiety behavior is reversed by histone deacetylase inhibition. Neurogastroenterol Motil. 2015;27:1831–6. doi: 10.1111/nmo.12675. [DOI] [PubMed] [Google Scholar]
- 55.Gschossmann JM, Buenger L, Adam B, et al. Diurnal variation of abdominal motor responses to colorectal distension and plasma cortisol levels in rats. Neurogastroenterol Motil. 2001;13:585–9. doi: 10.1046/j.1365-2982.2001.00293.x. [DOI] [PubMed] [Google Scholar]
- 56.Chang L, Sundaresh S, Elliott J, et al. Dysregulation of the hypothalamic-pituitary-adrenal (HPA) axis in irritable bowel syndrome. Neurogastroenterol Motil. 2009;21:149–59. doi: 10.1111/j.1365-2982.2008.01171.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liedhegner ES, Sasman A, Hillard CJ. Brain region-specific changes in N-acylethanolamine contents with time of day. J Neurochem. 2014;128:491–506. doi: 10.1111/jnc.12495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Patel S, Roelke CT, Rademacher DJ, et al. Endocannabinoid signaling negatively modulates stress-induced activation of the hypothalamic-pituitary-adrenal axis. Endocrinology. 2004;145:5431–8. doi: 10.1210/en.2004-0638. [DOI] [PubMed] [Google Scholar]
- 59.Saber-Tehrani A, Naderi N, Hosseini Najarkolaei A, et al. Cannabinoids and their interactions with diazepam on modulation of serum corticosterone concentration in male mice. Neurochem Res. 2010;35:60–6. doi: 10.1007/s11064-009-0030-9. [DOI] [PubMed] [Google Scholar]
- 60.Cone EJ, Johnson RE, Moore JD, et al. Acute effects of smoking marijuana on hormones, subjective effects and performance in male human subjects. Pharmacol Biochem Behav. 1986;24:1749–54. doi: 10.1016/0091-3057(86)90515-0. [DOI] [PubMed] [Google Scholar]
- 61.Kleinloog D, Liem-Moolenaar M, Jacobs G, et al. Does olanzapine inhibit the psychomimetic effects of Delta(9)-tetrahydrocannabinol? J Psychopharmacol. 2012;26:1307–16. doi: 10.1177/0269881112446534. [DOI] [PubMed] [Google Scholar]
- 62.Klumpers LE, Beumer TL, van Hasselt JG, et al. Novel Delta(9) -tetrahydrocannabinol formulation Namisol(R) has beneficial pharmacokinetics and promising pharmacodynamic effects. Br J Clin Pharmacol. 2012;74:42–53. doi: 10.1111/j.1365-2125.2012.04164.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.D'Souza DC, Ranganathan M, Braley G, et al. Blunted psychotomimetic and amnestic effects of delta-9-tetrahydrocannabinol in frequent users of cannabis. Neuropsychopharmacology. 2008;33:2505–16. doi: 10.1038/sj.npp.1301643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ranganathan M, Braley G, Pittman B, et al. The effects of cannabinoids on serum cortisol and prolactin in humans. Psychopharmacology (Berl) 2009;203:737–44. doi: 10.1007/s00213-008-1422-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.King GR, Ernst T, Deng W, et al. Altered brain activation during visuomotor integration in chronic active cannabis users: relationship to cortisol levels. J Neurosci. 2011;31:17923–31. doi: 10.1523/JNEUROSCI.4148-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Somaini L, Manfredini M, Amore M, et al. Psychobiological responses to unpleasant emotions in cannabis users. Eur Arch Psychiatry Clin Neurosci. 2012;262:47–57. doi: 10.1007/s00406-011-0223-5. [DOI] [PubMed] [Google Scholar]
- 67.van Leeuwen AP, Creemers HE, Greaves-Lord K, et al. Hypothalamic-pituitary-adrenal axis reactivity to social stress and adolescent cannabis use: the TRAILS study. Addiction. 2011;106:1484–92. doi: 10.1111/j.1360-0443.2011.03448.x. [DOI] [PubMed] [Google Scholar]
- 68.Van Sickle MD, Duncan M, Kingsley PJ, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310:329–32. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]
- 69.Napadow V, Sheehan JD, Kim J, et al. The brain circuitry underlying the temporal evolution of nausea in humans. Cereb Cortex. 2013;23:806–13. doi: 10.1093/cercor/bhs073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sclocco R, Kim J, Garcia RG, et al. Brain Circuitry Supporting Multi-Organ Autonomic Outflow in Response to Nausea. Cereb Cortex. 2014 doi: 10.1093/cercor/bhu172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sticht MA, Limebeer CL, Rafla BR, et al. Endocannabinoid regulation of nausea is mediated by 2-arachidonoylglycerol (2-AG) in the rat visceral insular cortex. Neuropharmacology. 2016;102:92–102. doi: 10.1016/j.neuropharm.2015.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Parker LA, Rock EM, Sticht MA, et al. Cannabinoids suppress acute and anticipatory nausea in preclinical rat models of conditioned gaping. Clin Pharmacol Ther. 2015;97:559–61. doi: 10.1002/cpt.98. [DOI] [PubMed] [Google Scholar]
- 73.Parker LA. Conditioned flavor avoidance and conditioned gaping: rat models of conditioned nausea. Eur J Pharmacol. 2014;722:122–33. doi: 10.1016/j.ejphar.2013.09.070. [DOI] [PubMed] [Google Scholar]
- 74.Sticht MA, Limebeer CL, Rafla BR, et al. Intra-visceral insular cortex 2-arachidonoylglycerol, but not N-arachidonoylethanolamide, suppresses acute nausea-induced conditioned gaping in rats. Neuroscience. 2015;286:338–44. doi: 10.1016/j.neuroscience.2014.11.058. [DOI] [PubMed] [Google Scholar]
- 75.Allen JH, de Moore GM, Heddle R, et al. Cannabinoid hyperemesis: cyclical hyperemesis in association with chronic cannabis abuse. Gut. 2004;53:1566–70. doi: 10.1136/gut.2003.036350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Lee LY, Abbott L, Moodie S, et al. Cyclic vomiting syndrome in 28 patients: demographics, features and outcomes. Eur J Gastroenterol Hepatol. 2012;24:939–43. doi: 10.1097/MEG.0b013e328354fc83. [DOI] [PubMed] [Google Scholar]
- 77.Chang YH, Windish DM. Cannabinoid hyperemesis relieved by compulsive bathing. Mayo Clin Proc. 2009;84:76–8. doi: 10.4065/84.1.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Soriano-Co M, Batke M, Cappell MS. The cannabis hyperemesis syndrome characterized by persistent nausea and vomiting, abdominal pain, and compulsive bathing associated with chronic marijuana use: a report of eight cases in the United States. Dig Dis Sci. 2010;55:3113–9. doi: 10.1007/s10620-010-1131-7. [DOI] [PubMed] [Google Scholar]
- 79.Wallace EA, Andrews SE, Garmany CL, et al. Cannabinoid hyperemesis syndrome: literature review and proposed diagnosis and treatment algorithm. South Med J. 2011;104:659–64. doi: 10.1097/SMJ.0b013e3182297d57. [DOI] [PubMed] [Google Scholar]
- 80.Darmani NA. Cannabinoid-Induced Hyperemesis: A Conundrum—From Clinical Recognition to Basic Science Mechanisms. Pharmaceuticals. 2010;3:2163–2177. doi: 10.3390/ph3072163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Bolognesi M, Di Pascoli M, Verardo A, et al. Splanchnic vasodilation and hyperdynamic circulatory syndrome in cirrhosis. World J Gastroenterol. 2014;20:2555–63. doi: 10.3748/wjg.v20.i10.2555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Camilleri M, Kolar GJ, Vazquez-Roque MI, et al. Cannabinoid receptor 1 gene and irritable bowel syndrome: phenotype and quantitative traits. Am J Physiol Gastrointest Liver Physiol. 2013;304:G553–60. doi: 10.1152/ajpgi.00376.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Park JM, Choi MG, Cho YK, et al. Cannabinoid receptor 1 gene polymorphism and irritable bowel syndrome in the Korean population: a hypothesis-generating study. J Clin Gastroenterol. 2011;45:45–9. doi: 10.1097/MCG.0b013e3181dd1573. [DOI] [PubMed] [Google Scholar]
- 84.Camilleri M, Carlson P, McKinzie S, et al. Genetic variation in endocannabinoid metabolism, gastrointestinal motility, and sensation. Am J Physiol Gastrointest Liver Physiol. 2008;294:G13–9. doi: 10.1152/ajpgi.00371.2007. [DOI] [PubMed] [Google Scholar]
- 85.Fichna J, Wood JT, Papanastasiou M, et al. Endocannabinoid and cannabinoid-like fatty acid amide levels correlate with pain-related symptoms in patients with IBS-D and IBS-C: a pilot study. PLoS One. 2013;8:e85073. doi: 10.1371/journal.pone.0085073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Vianna CR, Donato J, Jr, Rossi J, et al. Cannabinoid receptor 1 in the vagus nerve is dispensable for body weight homeostasis but required for normal gastrointestinal motility. J Neurosci. 2012;32:10331–7. doi: 10.1523/JNEUROSCI.4507-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Keenan CM, Storr MA, Thakur GA, et al. AM841, a covalent cannabinoid ligand, powerfully slows gastrointestinal motility in normal and stressed mice in a peripherally restricted manner. Br J Pharmacol. 2015;172:2406–18. doi: 10.1111/bph.13069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Abalo R, Chen C, Vera G, et al. In vitro and non-invasive in vivo effects of the cannabinoid-1 receptor agonist AM841 on gastrointestinal motor function in the rat. Neurogastroenterol Motil. 2015 doi: 10.1111/nmo.12668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Cluny NL, Keenan CM, Duncan M, et al. Naphthalen-1-yl-(4-pentyloxynaphthalen-1-yl)methanone (SAB378), a peripherally restricted cannabinoid CB1/CB2 receptor agonist, inhibits gastrointestinal motility but has no effect on experimental colitis in mice. J Pharmacol Exp Ther. 2010;334:973–80. doi: 10.1124/jpet.110.169946. [DOI] [PubMed] [Google Scholar]
- 90.Bashashati M, Nasser Y, Keenan CM, et al. Inhibiting endocannabinoid biosynthesis: a novel approach to the treatment of constipation. Br J Pharmacol. 2015;172:3099–111. doi: 10.1111/bph.13114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fichna J, Salaga M, Stuart J, et al. Selective inhibition of FAAH produces antidiarrheal and antinociceptive effect mediated by endocannabinoids and cannabinoid-like fatty acid amides. Neurogastroenterol Motil. 2014;26:470–81. doi: 10.1111/nmo.12272. [DOI] [PubMed] [Google Scholar]
- 92.Bashashati M, Storr MA, Nikas SP, et al. Inhibiting fatty acid amide hydrolase normalizes endotoxin-induced enhanced gastrointestinal motility in mice. Br J Pharmacol. 2012;165:1556–71. doi: 10.1111/j.1476-5381.2011.01644.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Mukhopadhyay P, Horvath B, Rajesh M, et al. Fatty acid amide hydrolase is a key regulator of endocannabinoid-induced myocardial tissue injury. Free Radic Biol Med. 2011;50:179–95. doi: 10.1016/j.freeradbiomed.2010.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hoyer FF, Khoury M, Slomka H, et al. Inhibition of endocannabinoid-degrading enzyme fatty acid amide hydrolase increases atherosclerotic plaque vulnerability in mice. J Mol Cell Cardiol. 2014;66:126–32. doi: 10.1016/j.yjmcc.2013.11.013. [DOI] [PubMed] [Google Scholar]
- 95.Lenglet S, Thomas A, Soehnlein O, et al. Fatty acid amide hydrolase deficiency enhances intraplaque neutrophil recruitment in atherosclerotic mice. Arterioscler Thromb Vasc Biol. 2013;33:215–23. doi: 10.1161/ATVBAHA.112.300275. [DOI] [PubMed] [Google Scholar]
- 96.Massa F, Marsicano G, Hermann H, et al. The endogenous cannabinoid system protects against colonic inflammation. J Clin Invest. 2004;113:1202–9. doi: 10.1172/JCI19465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Storr MA, Keenan CM, Emmerdinger D, et al. Targeting endocannabinoid degradation protects against experimental colitis in mice: involvement of CB1 and CB2 receptors. J Mol Med (Berl) 2008;86:925–36. doi: 10.1007/s00109-008-0359-6. [DOI] [PubMed] [Google Scholar]
- 98.Salaga M, Mokrowiecka A, Zakrzewski PK, et al. Experimental colitis in mice is attenuated by changes in the levels of endocannabinoid metabolites induced by selective inhibition of fatty acid amide hydrolase (FAAH) J Crohns Colitis. 2014;8:998–1009. doi: 10.1016/j.crohns.2014.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Alhouayek M, Lambert DM, Delzenne NM, et al. Increasing endogenous 2-arachidonoylglycerol levels counteracts colitis and related systemic inflammation. FASEB J. 2011;25:2711–21. doi: 10.1096/fj.10-176602. [DOI] [PubMed] [Google Scholar]
- 100.Storr M, Emmerdinger D, Diegelmann J, et al. The role of fatty acid hydrolase gene variants in inflammatory bowel disease. Aliment Pharmacol Ther. 2009;29:542–51. doi: 10.1111/j.1365-2036.2008.03910.x. [DOI] [PubMed] [Google Scholar]
- 101.Suarez J, Romero-Zerbo Y, Marquez L, et al. Ulcerative colitis impairs the acylethanolamide-based anti-inflammatory system reversal by 5-aminosalicylic acid and glucocorticoids. PLoS One. 2012;7:e37729. doi: 10.1371/journal.pone.0037729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.D'Argenio G, Valenti M, Scaglione G, et al. Up-regulation of anandamide levels as an endogenous mechanism and a pharmacological strategy to limit colon inflammation. FASEB J. 2006;20:568–70. doi: 10.1096/fj.05-4943fje. [DOI] [PubMed] [Google Scholar]
- 103.Di Sabatino A, Battista N, Biancheri P, et al. The endogenous cannabinoid system in the gut of patients with inflammatory bowel disease. Mucosal Immunol. 2011;4:574–83. doi: 10.1038/mi.2011.18. [DOI] [PubMed] [Google Scholar]
- 104.Sasso O, Migliore M, Habrant D, et al. Multitarget fatty acid amide hydrolase/cyclooxygenase blockade suppresses intestinal inflammation and protects against nonsteroidal anti-inflammatory drug-dependent gastrointestinal damage. FASEB J. 2015;29:2616–27. doi: 10.1096/fj.15-270637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Darmani NA, Izzo AA, Degenhardt B, et al. Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and neuropathic conditions: review of the available pre-clinical data, and first human studies. Neuropharmacology. 2005;48:1154–63. doi: 10.1016/j.neuropharm.2005.01.001. [DOI] [PubMed] [Google Scholar]
- 106.Borrelli F, Romano B, Petrosino S, et al. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br J Pharmacol. 2015;172:142–58. doi: 10.1111/bph.12907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Esposito G, Capoccia E, Turco F, et al. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-alpha activation. Gut. 2014;63:1300–12. doi: 10.1136/gutjnl-2013-305005. [DOI] [PubMed] [Google Scholar]
- 108.Alhouayek M, Bottemanne P, Subramanian KV, et al. N-Acylethanolamine-hydrolyzing acid amidase inhibition increases colon N-palmitoylethanolamine levels and counteracts murine colitis. FASEB J. 2015;29:650–61. doi: 10.1096/fj.14-255208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Fichna J, Bawa M, Thakur GA, et al. Cannabinoids alleviate experimentally induced intestinal inflammation by acting at central and peripheral receptors. PLoS One. 2014;9:e109115. doi: 10.1371/journal.pone.0109115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kipnes MS, Hollander P, Fujioka K, et al. A one-year study to assess the safety and efficacy of the CB1R inverse agonist taranabant in overweight and obese patients with type 2 diabetes. Diabetes Obes Metab. 2010;12:517–31. doi: 10.1111/j.1463-1326.2009.01188.x. [DOI] [PubMed] [Google Scholar]
- 111.Topol EJ, Bousser MG, Fox KA, et al. Rimonabant for prevention of cardiovascular events (CRESCENDO): a randomised, multicentre, placebo-controlled trial. Lancet. 2010;376:517–23. doi: 10.1016/S0140-6736(10)60935-X. [DOI] [PubMed] [Google Scholar]
- 112.Kalliomaki J, Annas P, Huizar K, et al. Evaluation of the analgesic efficacy and psychoactive effects of AZD1940, a novel peripherally acting cannabinoid agonist, in human capsaicin-induced pain and hyperalgesia. Clin Exp Pharmacol Physiol. 2013;40:212–8. doi: 10.1111/1440-1681.12051. [DOI] [PubMed] [Google Scholar]
- 113.Ostenfeld T, Price J, Albanese M, et al. A randomized, controlled study to investigate the analgesic efficacy of single doses of the cannabinoid receptor-2 agonist GW842166, ibuprofen or placebo in patients with acute pain following third molar tooth extraction. Clin J Pain. 2011;27:668–76. doi: 10.1097/AJP.0b013e318219799a. [DOI] [PubMed] [Google Scholar]
- 114.Huggins JP, Smart TS, Langman S, et al. An efficient randomised, placebo-controlled clinical trial with the irreversible fatty acid amide hydrolase-1 inhibitor PF-04457845, which modulates endocannabinoids but fails to induce effective analgesia in patients with pain due to osteoarthritis of the knee. Pain. 2012;153:1837–46. doi: 10.1016/j.pain.2012.04.020. [DOI] [PubMed] [Google Scholar]
- 115.O'Sullivan SE. Endocannabinoids and the Cardiovascular System in Health and Disease. Handb Exp Pharmacol. 2015;231:393–422. doi: 10.1007/978-3-319-20825-1_14. [DOI] [PubMed] [Google Scholar]
- 116.Pacher P, Kunos G. Modulating the endocannabinoid system in human health and disease--successes and failures. FEBS J. 2013;280:1918–43. doi: 10.1111/febs.12260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Wiley JW, Higgins GA, Athey BD. Stress and glucocorticoid receptor transcriptional programming in time and space: Implications for the brain-gut axis. Neurogastroenterol Motil. 2016;28:12–25. doi: 10.1111/nmo.12706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Di Marzo V, Stella N, Zimmer A. Endocannabinoid signalling and the deteriorating brain. Nat Rev Neurosci. 2015;16:30–42. doi: 10.1038/nrn3876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Krugers HJ, Hoogenraad CC, Groc L. Stress hormones and AMPA receptor trafficking in synaptic plasticity and memory. Nat Rev Neurosci. 2010;11:675–81. doi: 10.1038/nrn2913. [DOI] [PubMed] [Google Scholar]
- 120.Sticht MA, Rock EM, Limebeer CL, et al. Endocannabinoid Mechanisms Influencing Nausea. Int Rev Neurobiol. 2015;125:127–62. doi: 10.1016/bs.irn.2015.09.001. [DOI] [PubMed] [Google Scholar]
- 121.Lomax AE, Linden DR, Mawe GM, et al. Effects of gastrointestinal inflammation on enteroendocrine cells and enteric neural reflex circuits. Auton Neurosci. 2006;126-127:250–7. doi: 10.1016/j.autneu.2006.02.015. [DOI] [PubMed] [Google Scholar]