

Abstract
This review is dedicated to the antimicrobial metabolite-producing Pseudoalteromonas strains. The genus Pseudoalteromonas hosts 41 species, among which 16 are antimicrobial metabolite producers. To date, a total of 69 antimicrobial compounds belonging to 18 different families have been documented. They are classified into alkaloids, polyketides, and peptides. Finally as Pseudoalteromonas strains are frequently associated with macroorganisms, we can discuss the ecological significance of antimicrobial Pseudoalteromonas as part of the resident microbiota.
Keywords: Pseudoalteromonas, antimicrobial metabolites, alkaloid, polyketide, non ribosomal peptide, genome mining, marine host-associated microbiota, probiotic
1. Introduction
Last October, we celebrated the 20th anniversary of the genus Pseudoalteromonas having been split from Alteromonas [1]. The genus Pseudoalteromonas includes Gram-negative, heterotrophic, and aerobic bacteria with a polar flagellum and has a GC content comprised between 38% and 50% [2]. It belongs to the order Alteromonadales in the γ-Proteobacteria class. The Pseudoalteromonas strains require a seawater base for growth and are therefore true marine bacteria. Nowadays, 41 species are assigned to this genus and over 3772 Pseudoalteromonas strains are currently listed in the NCBI taxonomy browser. Allowing for rare exceptions, Pseudoalteromonas strains are associated with healthy animals or algae. To date, few strains are known as pathogenic or opportunistic. Flavobacterium piscicida, reclassified as P. piscicida by Gauthier et al., is involved in flavobacteriosis in farm fish [3], while P. agarivorans NW4327 was recently reported as pathogenic for the sponge Rhopaloeides odorabile [4].
This bacterial genus is of great interest to the scientific community because of (i) its prolific metabolite-producing capacity and (ii) its usual association with macroorganisms [5], leading to a suspected and sometimes documented ecological significance. These two properties may, in fact, be interconnected. According to the hologenome theory [6,7,8,9,10], the holobiont is composed of the host and its associated microbial communities, named microbiota. Such a superorganism gains genetic plasticity and flexibility and therefore appears better equipped to face and adapt to environmental variations. The microbiota is supposed to play a critical role in holobiont homeostasy through its metabolic activities. Moreover, the microbial shielding of the microbiota may defend the holobiont from pathogen settlement and therefore participate in the host protection. The abundance of antimicrobial metabolite described from Pseudoalteromonas strains points them out as a key partner in marine invertebrate holobionts.
This review focuses on the antimicrobial compounds produced by Pseudoalteromonas strains. It presents a chemical inventory of the Pseudoalteromonas antimicrobial metabolites known to date. Their bioactivity will not be discussed because of the lack of a standardized panel of target microorganisms. Indeed, although significant efforts have been made regarding testing procedures, the use of various microbial species and strains is not suitable for a rigorous comparison of antimicrobial activity and potency. Finally, the potential ecological significance of the antimicrobial metabolite-producing Pseudoalteromonas is revisited in the light of antimicrobial activity and the origin of the bioactive strains.
2. Antimicrobial Metabolites from Pseudoalteromonas Species
In marine microorganisms, secondary metabolites are mostly comprised of nitrogenated (56%), acetate- (30%), and isoprene-derived (13%) molecules [11]. On the basis of their biosynthetic pathways, they are classified as alkaloids or peptides, polyketides, and terpenoids [12]. This review will adopt the same classification system for the antimicrobial metabolites produced by Pseudoalteromonas strains that way, the antimicrobial metabolites from Pseudoalteromonas known to date (n = 69) can be categorized as alkaloids (n = 32, 46%), polyketides (n = 20, 29%), and peptides (n = 17, 25%) (Figure 1). The antimicrobial strains that produce these metabolites are affiliated with at least 16 different species. In comparison, only six of the 126 species composing the dominant marine bacterial family, Vibrionaceae, are known to produce 29 antimicrobial identified compounds [13].
Figure 1.

Neighbor-joining tree indicating the phylogenetic relationships inferred from partial 16S rDNA gene sequences (1196 nt) of Pseudoalteromonas strains producing either alkaloids (green), polyketides (red), or peptides (blue). Empty circles represent strains producing unidentified antimicrobial compounds. Bootstrap values (expressed as percentage of 1000 replications) > 50% are shown at branching points. The proteobacteria Escherichia coli 2012K11 (position 167-1356) was used as outgroup. Bar, 0.01 substitutions per nucleotide.
Among the Pseudoalteromonas genus, the metabolite-producing capacity has usually been associated with pigmentation. The antimicrobial metabolite-producing Pseudoalteromonas species did not derogate since most of them (>80%) are pigmented and the bioactive metabolites produced have been shown to cause this appearance [14]. For example, P. tunicata CCUG 26757 and P. rubra DSM 6842 strains produced various pigments involved in their antibacterial and antifungal activity [15,16,17,18,19,20,21]. However, such a correlation is abusive. Indeed pigmentation is the result of the presence of conjugated double bonds in the chemical structure of the antimicrobial metabolite. This structural feature is not a prerogative for antimicrobial activity, as exemplified by bioactive non-ribosomal peptides that are colorless. Hence, this observation should not be laid down as a rule. It is important to keep in mind that non-pigmented Pseudoalteromonas strains are also potential bioactive metabolite producer even though it is less frequently observed.
2.1. Alkaloids
Alkaloids are biosynthesized from amino acids. They are predominantly made of heterocyclic incorporated basic amine nitrogen. Mostly derived from α-amino acids such as Phe, Tyr, Trp, Lys, and Orn, the biosynthesis pathways of alkaloids remain poorly understood [22]. Alkaloids represent the majority of the antimicrobial metabolites from Pseudolteromonas, having been isolated from a vast number of strains belonging to seven species (Figure 1). The antimicrobial alkaloid metabolites from Pseudoalteromonas are divided into non-halogenated (Table 1) and halogenated compounds (Table 2).
Table 1.
Non-halogenated compounds from Pseudoalteromonas species with antimicrobial activities.
| Producers | Products | Sensitive Microorganisms | Ref. | ||
|---|---|---|---|---|---|
| Affiliated Species | Strain | Name | Structure | ||
| P. peptidolytica | J010 | Korormicin, R: (CH2)7CH3 |
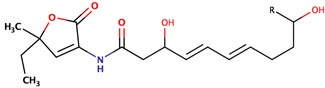 |
Gram-negative | [27,28,29] |
| Korormicin G | 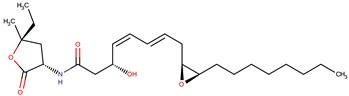 |
||||
| Korormicin J R = CH2CH3 |
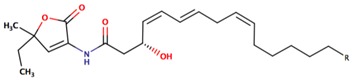 |
||||
| Korormicin K R = (CH2)2CH3 | |||||
| P. tunicata | CCUG 267547 | Tambjamine | 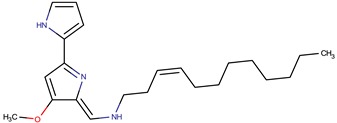 |
Staphylococcus aureus, Escherichia coli, Candida albicans, Malassezia furfur | [20,21,30,31,32,33,34,35] |
| Tambjamine YP1 | 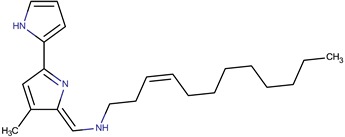 |
||||
| P. rubra | DSM 6842 | Prodigiosin | 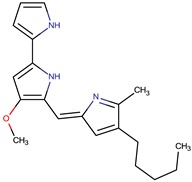 |
Staphylococcus aureus, Escherichia coli, Candida albicans | [15,16,17] |
| Cycloprodigiosin | 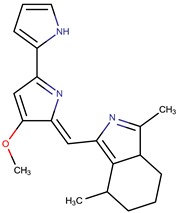 |
||||
| 2-(p-H-benzyl) prodigiosin | 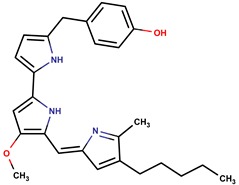 |
||||
| P. luteoviolacea | NCIMB 2035 | Violacein R1 = OH, R2 = H |
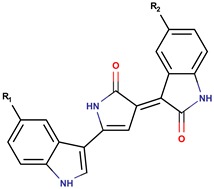 |
Staphylococcus aureus, Bacilus subtilis, Bacillus megaterium, Photobacterium sp., fungi | [15,36,37] |
| Oxyviolacein R1 = R2 = OH | |||||
| Deoxyviolacein R1 = R2 = H | |||||
| Indolmycin | 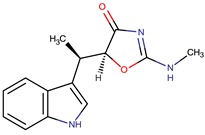 |
Staphylococcus aureus | [15,38] | ||
|
P. rubra, P. piscicida |
DSM 6842 A1-J11 | pentyl-quinolinone R: (CH2)3CH3 |
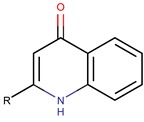 |
Staphylococcus aureus, Vibrio anguillarum, Vibrio harveyi, Candida albicans | [15,39,40,41,42] |
| heptyl-quinolinone R: (CH2)5CH3 | |||||
| nonyl-quinolinone R: (CH2)7CH3 | |||||
| P. piscicida | NJ6-3-1 | Norharman | 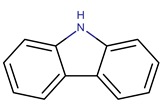 |
Bacillus subtilis, Staphylococcus aureus, Agrobacterium tumefaciens, Escherichia coli, Saccharomyces cerevisiae | [43] |
| P. flavipulchra | JG1 | n-hydroxy benzoisoxazolone, 2′-deoxyadenosine |
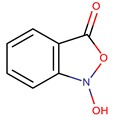 |
Gram-positive and Gram-negative | [44] |
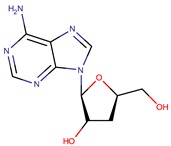 | |||||
| P. issachenkonni | non-referenced | Isatin | 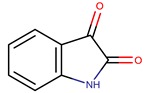 |
Candida albicans | [45,46,47] |
| P. piscicida | OT59 | Alteramide A | 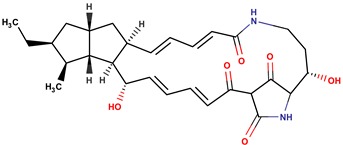 |
Fungi | [48] |
Table 2.
Halogenated compounds from Pseudoalteromonas species with antimicrobial activities.
| Producers | Products | Sensitive Microorganisms | Ref. | ||
|---|---|---|---|---|---|
| Affiliated Species | Strain | Name | Structure | ||
| P. luteoviolacea P. peptidolytica | I-L-33 J010 | Tetrabromopyrrole | 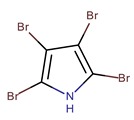 |
Staphylococcus aureus, Enterobacter aerogenes, Escherichia coli, Photobacterium fisheri, Photobacterium mandapamensis, Photobacterium phosphoreum, Pseudomonas aeruginosa, Candida albicans | [27,49] |
| P. luteoviolacea | I-L-33 | Hexa-bromo-2,2′-bipyrrole | 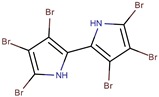 |
Photobacterium sp. | [23,49] |
| 4′-((3,4,5-tribromo-1H-pyrrol-2-yl)methyl) phenol | 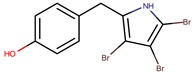 |
Staphylococcus aureus | [27] | ||
| P. luteoviolacea | 2ta16 | 2,4,6-Tribromophenol | 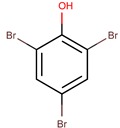 |
Bacteria and fungi | [50,51] |
| 2,6-dibromophenol | 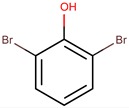 |
Fungi | [50,52] | ||
| P. luteoviolacea P. phenolica | I-L-33 D5047 | Pentabromopseudilin | 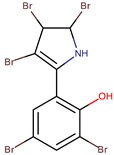 |
Staphylococcus aureus Photobacterium phosophoreum | [23,49,53,54,55] |
| P. phenolica | D5047 | 2,3,5,7-tetrabromobenzofuro [3,2-b]pyrrole | 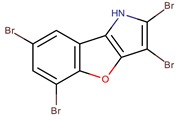 |
Staphylococcus aureus, Candida albicans | [55] |
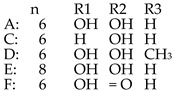
| P. peptidolytica | J010 | Korormicin H, F, I H: R1: (CH2)2CH3 R2: OH R3: Cl F: R1: (CH2)2CH3 R2: Br R3: OH I: R1: (CH2)2CH3 R2: Cl R3: OH |
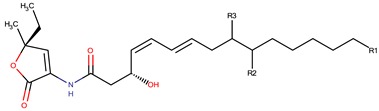 |
Gram-negative | [27] |
| P. flavipulchra | JG1 | 6-bromoindolyl-3-acetic acid | 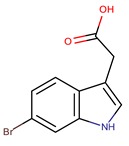 |
Gram-positive and Gram-negative | [44] |
A number of halogenated alkaloids is produced by marine animals, bacteria, and fungi [23]. Halogenation has now been observed in many natural product biosynthetic pathways and can play a significant role in establishing the bioactivity of a compound [24,25]. Only brominated and chlorinated residues are incorporated into antimicrobial halogenated alkaloids produced by Pseudoalteromonas species. They are associated with pyrrole or benzene ring(s) or aliphatic side chain. The bromoperoxidase enzyme catalyzes the oxidation of bromide (Br−) to the electrophilic bromonium cation (Br+), which in turn attacks hydrocarbons and results in the production of brominated alkaloids [23,26]. Similarly, chloroperoxidases are responsible for the production of chlorinated alkaloids. P. luteoviolacea produces four of the 10 brominated alkaloids with antimicrobial activity discovered among Pseudoalteromonas strains. Other strains such as P. peptidolytica, P. flavipulchra, and P. phenolica are also described to produce alkaloids with bromine, with P. peptidolytica being the only species to produce chlorinated alkaloids.
2.2. Polyketides
Polyketides constitute a rich and diverse family of natural compounds from prokaryotes and eukaryotes. By definition, all polyketides are synthesized by condensation of acetyl (also referred to as ketides) or malonyl units via specific enzymes called PolyKetide Synthases (PKS). These enzymes catalyze the condensation of activated acyl derivatives (acyl-CoA) on an existing ketide linked to the enzyme via a thioester bond [56]. Several categories of PKS are described within bacteria, depending on (i) the linear/iterative functioning of the enzyme; and (ii) the way acyl-coA is incorporated (involving or not an Acyl Carrier Protein ACP) [57]. Hence, to date, three PKS types have been recognized. Type I PKS are sequential multifunctional enzymes while type II PKS are iterative and multienzyme complexes. In contrast to type I and II PKS, type III PKS are iterative but do not involve ACP. Until the late 1990s, type III was thought to be plant-specific but recent developments have highlighted that they are also found in microorganisms [58]. PKS have been shown to yield many bioactive molecules, especially antibiotics such as erythromycin [59] and tetracycline [60]. Within the genus Pseudoalteromonas, only three different species produce nine groups of polyketides exhibiting antimicrobial activity (Table 3). Indeed, polyketides are at the origin of numerous antimicrobial phenols and, in association with NRPS and Fatty Acid Synthase (FAS) systems, of the well-known thiomarinols produced by P. luteoviolacea [61] (Table 3).
Table 3.
Polyketides produced by Pseudoalteromonas species with antimicrobial activities.
| Producers | Products | Sensitive Microorganisms | Ref. | ||
|---|---|---|---|---|---|
| Affiliated Species | Strain | Name | Structure | ||
| P. phenolica | O-BC30T | MC21-A | 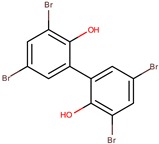 |
Staphylococcus aureus, Escherichia coli | [55,62] |
| D5047 | 4,4′,6-tribromo-2,2′-biphenol | 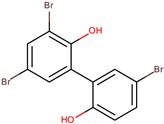 |
|||
| P. luteoviolacea | Non-referenced | 2,4-dibromo-6-chlorophenol | 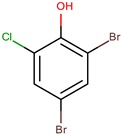 |
Staphylococcus aureus | [63] |
| I-L-33 | 4-hydroxy benzaldehyde (2) | 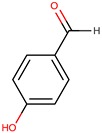 |
Gram-positive Gram-negative | [49] | |
| n-propyl-3-hydroxybenzoate (3) | 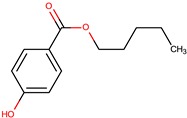 |
||||
| P. luteoviolacea | SANK 73390 | Thiomarinols A,C,D,E,F | 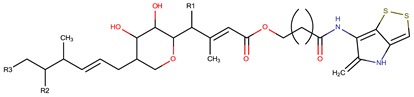 |
Gram-positive Gram-negative | [61,64,65,66,67] |
 |
|||||
| Thiomarinol B | 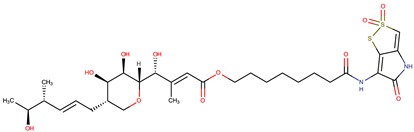 |
||||
| Thiomarinol G | |
||||
| Xenorhabdin | 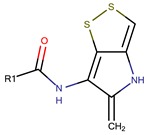 |
||||
| R1: decanoyl | |||||
| R1: dodecanoyl | |||||
| R1: E-dec-3-enoyl | |||||
| R1: Z-dec-4-enoyl | |||||
| R1: E-tetradecenoyl | |||||
| R1: Z-hexadecenoyl | |||||
| P. flavipulchra | JG1 | p-hydroxybenzoic acid | 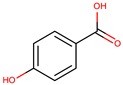 |
Bacillus subtilis, Aeromonas hydrophila, Photobacterium damselae, Vibrio anguillarum, Vibrio harveyi | [44] |
| trans-cinnamic acid | 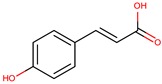 |
||||
2.3. Non Ribosomally Synthesized Peptides
Non Ribosomal Peptides Synthetases (NRPS) represent a wide group of multimodular enzymes that polymerize amino acids as well as fatty acids, and α-hydroxy acids [68]. NRPS can also incorporate unusual amino acids (non-proteinogenic) and hence generate a great chemodiversity. Basically, NRPS are divided into modules, each one being responsible for binding one specific amino acid to the peptidic chain. To do so, each module consists of at least three domains. The Adenylation domain (A) activates the amino acid. Then, the Peptidyl Carrier Protein (PCP, also referred to as thiolation domain T) transfers it to the Condensation domain, which incorporates the new amino acid residue into the rest of the peptide via a peptidic bond. For the last module, a final ThioEsterase domain (TE), breaks the NRPS–Peptide bond and releases the product [69]. Furthermore, several tailoring modules can modify the structure of the peptide [70]. As for PKS, the mechanism of synthesis is usually linear but can also be iterative [69,71]. NRPS are frequently associated with PKS and even FAS to generate hybrid molecules, such as the bromoalterochromides produced by several Pseudoalteromonas species (Table 4) [72,73]. Actually, many NRPS products are hybrid, meaning these products can be classified as polyketides and non-ribosomal peptides at the same time.
Table 4.
Peptidic compounds produced by Pseudoalteromonas species with antimicrobial activities.
| Producer | Compound | Sensitive Microorganisms | Ref. | ||
|---|---|---|---|---|---|
| Affiliated Species | Strain | Name | Structure | ||
|
P. rubra P. flavipulchra P. maricaloris |
DSM 6842 NCIMB 2033 KMM 636 |
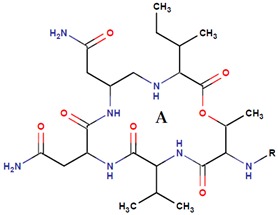
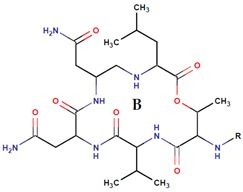
Bromoalterochromide A, R = i B, R = i Dibromoalterochromide A, R = ii B, R = ii |
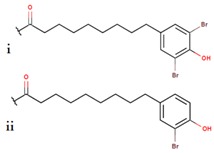 |
Bacillus subtilis, Staphylococcus aureus, Enterococcus faecium, Vibrio anguillarum, Candida albicans | [15,74] |
 |
 |
||||
| P. maricaloris | KMM 636 | cyclo-(isoleucyl-prolyl-leucyl-alanyl) | Bacteria and fungi | [75] | |
2.4. Bacteriocins and Bacteriocin-Like Inhibitory Substances (BLIS)
Bacteriocins and Bacteriocin-Like Inhibitory Substances (BLIS) are proteinaceous antibacterial compounds. They are ribosomally synthesized and exhibit a narrow spectrum of activity, generally limited to closely related strains. In Pseudoalteromonas, most of them are large proteins (MW > 100 kDa) (Table 5). Although they fell under this classification, most of them exhibited enzymatic activity ((l)-amino acid oxidase (LAAO)) activity. In many cases, these LAAOs were considered to be a flavin adenine dinucleotide (FAD)-containing homodimeric protein, [76]. LAAOs are widely found in Pseudoalteromonas and provide an important ecological function in marine environments [77].
Table 5.
Bacteriocins and BLIS produced by Pseudoalteromonas species.
| Producer | Compound | Sensitive Microorganisms | Ref. | |||
|---|---|---|---|---|---|---|
| Affiliated Species | Strain | Name | Structural Information | |||
| P. luteoviolacea | CPMOR-1 | l-amino acid oxidase | 110 kDa | Bacillus subtilis, Staphylococcus | [78] | |
| epidermidis, Escherichia coli | ||||||
| P. tunicata | D2 | AlpP | l-Lysine oxidase | 190 kDa | Gram positive Gram negative |
[79] |
| P. flavipulchra | C2 | l-amino acid oxidase | 60 kDa | Staphylococcus aureus | [80] | |
| JG1 | PfaP | 694 amino acids | 77 kDa | Vibrio anguillarum | [81] | |
| P. luteoviolacea | 9k-V10 | 100 kDa | Gram positive Gram negative |
[82] | ||
| 9k-V9 | 50 kDa | Vibrio parahaemolyticus | [41,42] | |||
| P. prydzensis | hCg-6 | BLIS | Gram negative | |||
| hCg-42 | BLIS | Gram negative | [71,83] | |||
| DIT44, DIT46, DIT9 | Amphiphilic compounds | <3 kDa | Vibrio parahaemolyticus | [84] | ||
2.5. Uncharacterized Chemistry of Antimicrobial Metabolites Produced by Pseudoalteromonas Species
Numerous Pseudoalteromonas strains have shown an antimicrobial activity but the bioactive compound(s) remain unidentified [55,85,86,87]. During a global marine research cruise, 15 strains related to P. ruthenica have exhibited antibacterial activity against Vibrio anguillarum [15]. Three bioactive fractions were detected after ethyl acetate extraction and RP-HPLC splitting. The compounds could not be assigned to any known secondary metabolites produced by pigmented Pseudoalteromonas species [15]. Moreover, P. tunicata strains inhibiting a variety of common fouling organisms were shown to produce a polar, heat-stable compound [88] and a heat-sensitive 3–10 kDa compound [89]. A cell-free supernatant of a P. piscicida strain isolated from a crustacean also showed an antifungal activity [90]. More recently, an antimicrobial anionic protein with an 87 kDa molecular weight was isolated from a culture of P. piscida [91]. This hydrophobic compound showed a high content of serine as well as aspartic and glutamic acids, but its amino acid sequence was not defined. Finally, two yellow-pigmented strains of P. citrea and P. aurantia were shown to exert an antimicrobial activity against bacteria, but their chemical nature has not been elucidated yet [92,93].
2.6. Genome Mining Strategies as a Tool to Discover Antibiotics in Pseudoalteromonas
The last two decades were marked by the onset of genomic area with cheaper and faster technologies for genome sequencing. To date, 29,000 complete bacterial genomes have been deciphered and another 31,000 bacterial genome sequences are still in progress under way (GOLD database: http://www.genomesonline.org; date of access: May 2016). This has led to the development of genome-based strategies to discover new drugs and antibiotics [94]. As a consequence, bioinformatic tools have been developed to improve the metabolite-pathway analysis, allowing access to putative metabolites and therefore to the investigation of the biotechnological potential of bacterial strains. Thanks to this genome mining approach, the metabolome can be predicted. Furthermore, bioinformatic tools enable the identification of new and potentially novel compounds via the expression of silent genes. Indeed, it is estimated that less than 10% of secondary metabolite gene clusters are expressed in sufficient amount for detection in current lab conditions [95].
As NRPS, PKS, and hybrid NRPS-PKS pathways exhibit repeated motifs in genomic nuclear sequences, they have led to the development of powerful bioinformatics programs such as antiSMASH [96], NapDos [97], SBSPKS [98], or Np.searcher [99]. Moreover, the description of a specificity-conferring code in the NRPS A-domain [100] has resulted in the development of specialised chemical structure prediction tools, e.g., NRPS predictor2 [101] and NRPS substrate predictor [102]. Another tool dedicated to ribosomally-produced peptides (bacteriocins), BAGEL3 [103] can be useful in a genome mining approach focusing on the discovery of new antimicrobial compounds. The reader is referred to the recent reviews focusing on genomics strategies defined to discover microbial natural products [104,105,106,107]. Combined with this, experimental plans have also been established to characterize potential secondary metabolites according to in silico analyses: isotope labelling, gene knockout, heterologous expression, or transcription activation. For more information on these strategies, the reader is referred to [95,106,108,109].
Despite an increasing number of available Pseudoalteromonas genomes and an increased knowledge about their bioactive compounds, only a few studies based on genome mining have been dedicated to the discovery of new antibacterial metabolites [110,111]. A consequent genomic study of 21 antimicrobial marine bacteria, of which seven belonged to the Pseudoalteromonas genus, highlighted the potential of pigmented Pseudoalteromonas strain as producers of secondary metabolites. Genomic analyses corroborated biochemical results e.g., Photobacterium halotolerans S2753 was previously described to produce holomycin [13] and the gene cluster responsible for its biosynthesis was underlined in this study. Concerning Pseudoalteromonas, despite a deeper analysis, no potential gene or gene cluster was identified on the bacterial genome of Pseudoalteromonas ruthenica [110]. As for biochemical approaches, all efforts made into genome analysis failed to characterize anti-Vibrio and Staphylococcus compounds in this strain. Added to this, the same team recently applied a coupled metabolomics and genomics workflow to determine the biosynthetic potential of Pseudoalteromonas luteoviolaceae [112]. Combined methods allowed rapid identification of new antibiotics and their biosynthetic pathways. Papaleo and co-workers conducted a study on four Antartic strains, two of which belonged to the genus Pseudoalteromonas, having inhibitory activity against Burkholderia cepacia complex [111]. As antimicrobial activity appeared to be shared and related to microbial volatile organic compounds, they conducted a genomic comparative analysis to point possible common secondary metabolite producer genes. Few candidates (11) were shown to be involved in secondary metabolite biosynthesis, transport, and catabolism, and further analysis had to be performed to clarify the implication of these genes in the antimicrobial activity.
As shown by the small number of reports, genome mining is in its infancy for the genus Pseudoalteromonas; however, pioneering studies on this topic highlight the use of the genomics approach in bioprospecting. However, as underlined by these same studies, it should be kept in mind that the reality of these virtual metabolites is to be established. They have to be structurally and functionally characterized. Therefore, the genome mining tools should be considered as tools to orientate metabolite discovery and not as an end in themselves. Nevertheless, further analyses should be carried out to characterize the different pathways and/or the natural products. The recent development of generic tools for Pseudoalteromonas genetic manipulation [113], as well as more specific ones [77,114,115,116], will be helpful in this endeavor.
3. Ecological Significance in Marine Life
To date, of the 41 Pseudoalteromonas species described, 16 (39%) have been shown to produce antimicrobial compounds. When investigating the ecological niche occupied or the isolation origin of these bioactive Pseudoalteromonas, we observed that these antimicrobial metabolite-producing Pseudoalteromonas were directly associated with macroorganisms except for P. phenolica, isolated from seawater (Table 6). Therefore, there is a great temptation to connect Pseudoalteromonas antimicrobial properties with its host association. The antimicrobial metabolite producing Pseudoalteromonas strains may form a microbial shield and as a result contribute to the protection of their host against pathogens [111].
Table 6.
Marine macroorganisms that host antimicrobial Pseudoalteromonas species. (Green, blue and black point out algae, invertebrates, and vertebrates, respectively).
| Antimicrobial Pseudoalteromonas sp. | Marine Host | Ref. |
|---|---|---|
| P. atlantica | [87,117] | |
| P. citrea | [118,119,120] | |
| P. elyakovii | [117,121,122,123] | |
| P. flavipulchra | [80,124] | |
| P. haloplanktis | [125] | |
| P. issachenkonii | [126] | |
| P. luteoviolacea | [37,78,127,128] | |
| P. maricaloris | [129,130] | |
| P. peptidolytica | [131] | |
| P. phenolica | Unassociated marine macroorganisms | |
| P. piscicida | [90,132,133] | |
| P. prydzensis | [83,117,134] | |
| P. rubra | [16,135] | |
| P. ruthenica | [136,137] | |
| P. tunicata | [32,138,139] | |
| P. undina | Dicentrarchus labrax, Sparus aurata | [140] |
A plethora of antimicrobial metabolite-producing strains of Pseudoalteromonas has been isolated in association with marine invertebrates (Table 6). The most documented hosts belong to Porifera (sponges in particular) and Cnidaria. The former are known to house great bacterial diversity. Pseudoalteromonas strains producing antimicrobial metabolite in vitro have been isolated from animals living in tropical [141], temperate [87,142], and cold seawater [111]. In Cnidarians, the role of Pseudoalteromonas in host defense and health has been hypothesized facing the antimicrobial metabolite producing strains of Pseudoalteromonas in corals [6,48,143,144]. Such associations with antimicrobial metabolite producing strains of Pseudoalteromonas strains are less described in the other Phyla except for molluscs (mainly bivalves [84,145,146] and crustaceans [147]).
The suggested role of such bacterial strains has led to the hologenome theory and concept [6,7,8,9,10], in which the holobiont is considered as the true evolutionary unit. Therefore antimicrobial metabolite producing strains of Pseudoalteromonas may play a key role in microbiota shaping and microbial shielding of marine invertebrates. Using next-generation sequencing methods, the impact of biotic (pathogen or probiotic) and/or abiotic stresses onto microbiota may be clarified. Such a strategy was recently applied to hemolymph microbiome of oysters [148]. An abiotic stress (temperature) was shown to provoke significant modifications of the microbiome composition while a biotic one (Vibrio sp. infection) did not. This microbiome stability has supported the hypothetical role of microbiota in host defense. Exploiting antimicrobial-producing Pseudoalteromonas spp. as tools to shape the marine host-associated microbiota along with high-throughput sequencing of host-associated microbiota may elucidate the role of Pseudoalteromonas in host defense.
The molecular dialog between antimicrobial-producing Pseudoalteromonas and the host immune system is another area that should be investigated. Whether in invertebrates, vertebrates, or algae, macroorganisms have Pattern-Recognition Receptors (PRR) at their disposal to detect and recognize microbial components known as Pathogen-Associated Molecular Patterns (PAMP). In Gram-negative bacteria such as Pseudoalteromonas, one of the most potent PAMPs is the LipoPolySaccharide (LPS). Few studies completely defined LPS structures of Pseudoalteromonas species [149]. However, it appears that most Pseudoalteromonas LPS known to date are composed of a pentaacylated Lipid A instead of a hexaacylated one [149,150,151,152,153,154]. Such a structural difference may provide an advantage to evade PRRs-mediated recognition. Indeed, LPS from various Pseudoalteromonas strains were shown to elicit a low immune response and are capable of modulating immune responses in their hosts [150,153].
Finally, in vivo experiments are required to determine the ecological significance of the association between Pseudoalteromonas and their source macroorganisms. The major issue is to determine whether antimicrobial-producing Pseudoalteromonas provide a real benefit to their host, especially in the context of pathogenic events. In such a case, Pseudoalteromonas could stand as a next generation of probiotics for marine aquaculture.
4. Conclusions
The genus Pseudoalteromonas has a high, if not the highest, proportion of species producing antimicrobial metabolites in the marine bacterial world. However, this bacterial genus is still underexplored at the biotechnological level. Furthermore, the vast majority of Pseudoalteromonas spp. have not been found to exhibit pathogenicity. Almost all known antimicrobial-producing Pseudoalteromonas spp. originate from healthy marine macroorganisms, suggesting that these strains may participate in the host’s homeostasis. It would be simplistic to link only the associated microbiota and the ensuing microbial shield with the presence of Pseudoalteromonas strains. Obviously, the metabolite richness and the genetic plasticity of the symbiont–host relationship result from metabolic and therefore microbial diversity. Nevertheless, the involvement of antimicrobial-producing Pseudoalteromonas in microbiota shaping and protection of their host should not be under-estimated, or neglected. In any event, the genus Pseudoalteromonas appears as the first or at least one of the leading antimicrobial providers in the marine microbiota. Therefore, Pseudoalteromonas strains offer real potential to develop the next generation of marine probiotics and their use as probiotics in aquaculture should be further investigated.
Acknowledgments
Thanks are given to Hervé Bourdon for critical reading of the manuscript. This work was partly funded by the region Bretagne (ExMim project, ARED 8347 and IMA No. 14/D1215). Clément Offret was supported by a “Quimper-communauté” grant for his Ph.D. thesis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Gauthier G., Gauthier M., Christen R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. Int. J. Syst. Bacteriol. 1995;45:755–761. doi: 10.1099/00207713-45-4-755. [DOI] [PubMed] [Google Scholar]
- 2.Ivanova E.P., Kiprianova E.A., Mikhailov V.V., Levanova G.F., Garagulya A.D., Gorshkova N.M., Vysotskii M.V., Nicolau D.V., Yumoto N., Taguchi T., et al. Phenotypic diversity of Pseudoalteromonas citrea from different marine habitats and emendation of the description. Int. J. Syst. Evol. Microbiol. 1998;48:247–256. doi: 10.1099/00207713-48-1-247. [DOI] [PubMed] [Google Scholar]
- 3.Austin B. Environment and Aquaculture in Developing Countries. ICLARM; Penang, Malaysia: 1993. Environmental issues in the control of bacterial diseases of farmed fish; pp. 237–251. [Google Scholar]
- 4.Choudhury J.D., Pramanik A., Webster N.S., Llewellyn L.E., Gachhui R., Mukherjee J. The Pathogen of the Great Barrier Reef Sponge Rhopaloeides odorabile is a New Strain of Pseudoalteromonas agarivorans Containing Abundant and Diverse Virulence-Related Genes. Mar. Biotechnol. 2015;17:463–478. doi: 10.1007/s10126-015-9627-y. [DOI] [PubMed] [Google Scholar]
- 5.Bowman J.P. Bioactive Compound Synthetic Capacity and Ecological Significance of Marine Bacterial Genus Pseudoalteromonas. Mar. Drugs. 2007;5:220–241. doi: 10.3390/md504220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rosenberg E., Sharon G., Zilber-Rosenberg I. The hologenome theory of evolution contains Lamarckian aspects within a Darwinian framework. Environ. Microbiol. 2009;11:2959–2962. doi: 10.1111/j.1462-2920.2009.01995.x. [DOI] [PubMed] [Google Scholar]
- 7.Zilber-Rosenberg I., Rosenberg E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008;32:723–735. doi: 10.1111/j.1574-6976.2008.00123.x. [DOI] [PubMed] [Google Scholar]
- 8.Rosenberg E., Zilber-Rosenberg I. Microbes Drive Evolution of Animals and Plants: The Hologenome Concept. mBio. 2016;7:e01395-15. doi: 10.1128/mBio.01395-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Theis K.R., Dheilly N.M., Klassen J.L., Brucker R.M., Baines J.F., Bosch T.C.G., Cryan J.F., Gilbert S.F., Goodnight C.J., Lloyd E.A., et al. Getting the Hologenome Concept Right: An Eco-Evolutionary Framework for Hosts and Their Microbiomes. mSystems. 2016;1:e00028-16. doi: 10.1128/mSystems.00028-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Brucker R.M., Bordenstein S.R. The capacious hologenome. Zool. Jena Ger. 2013;116:260–261. doi: 10.1016/j.zool.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 11.Kelecom A. Secondary metabolites from marine microorganisms. An. Acad. Bras. Ciênc. 2002;74:151–170. doi: 10.1590/S0001-37652002000100012. [DOI] [PubMed] [Google Scholar]
- 12.Faulkner D.J. Marine natural products. Nat. Prod. Rep. 2001;18:1–49. doi: 10.1039/b006897g. [DOI] [PubMed] [Google Scholar]
- 13.Mansson M., Gram L., Larsen T.O. Production of bioactive secondary metabolites by marine Vibrionaceae. Mar. Drugs. 2011;9:1440–1468. doi: 10.3390/md9091440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huang Y.-L., Li M., Yu Z., Qian P.-Y. Correlation between pigmentation and larval settlement deterrence by Pseudoalteromonas sp. sf57. Biofouling. 2011;27:287–293. doi: 10.1080/08927014.2011.562978. [DOI] [PubMed] [Google Scholar]
- 15.Vynne N.G., Mansson M., Nielsen K.F., Gram L. Bioactivity, chemical profiling, and 16S rRNA-based phylogeny of Pseudoalteromonas strains collected on a global research cruise. Mar. Biotechnol. 2011;13:1062–1073. doi: 10.1007/s10126-011-9369-4. [DOI] [PubMed] [Google Scholar]
- 16.Fehér D., Barlow R.S., Lorenzo P.S., Hemscheidt T.K. A 2-Substituted Prodiginine, 2-(p-Hydroxybenzyl)prodigiosin, from Pseudoalteromonas rubra. J. Nat. Prod. 2008;71:1970–1972. doi: 10.1021/np800493p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lattasch H., Thomson R.H. A revised structure for cycloprodigiosin. Tetrahedron Lett. 1983;24:2701–2704. doi: 10.1016/S0040-4039(00)87981-2. [DOI] [Google Scholar]
- 18.Gerber N.N. Prodigiosin-like Pigments from Actinomadura (Nocardia) pelletieri and Actinomadura madurae. Appl. Microbiol. 1969;18:1–3. doi: 10.1128/am.18.1.1-3.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gerber N.N. Prodigiosin-like pigments. CRC Crit. Rev. Microbiol. 1975;3:469–485. doi: 10.3109/10408417509108758. [DOI] [PubMed] [Google Scholar]
- 20.Franks A., Haywood P., Holmström C., Egan S., Kjelleberg S., Kumar N. Isolation and structure elucidation of a novel yellow pigment from the marine bacterium Pseudoalteromonas tunicata. Molecules. 2005;10:1286–1291. doi: 10.3390/10101286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Holmström C., James S., Egan S., Kjelleberg S. Inhibition of common fouling organisms by marine bacterial isolates ith special reference to the role of pigmented bacteria. Biofouling. 1996;10:251–259. doi: 10.1080/08927019609386284. [DOI] [PubMed] [Google Scholar]
- 22.De Luca V., Salim V., Thamm A., Masada S.A., Yu F. Making iridoids/secoiridoids and monoterpenoid indole alkaloids: Progress on pathway elucidation. Curr. Opin. Plant Biol. 2014;19:35–42. doi: 10.1016/j.pbi.2014.03.006. [DOI] [PubMed] [Google Scholar]
- 23.Gribble G.W. The diversity of naturally occurring organobromine compounds. Chem. Soc. Rev. 1999;28:335–346. doi: 10.1039/a900201d. [DOI] [Google Scholar]
- 24.Neumann C.S., Fujimori D.G., Walsh C.T. Halogenation strategies in natural product biosynthesis. Chem. Biol. 2008;15:99–109. doi: 10.1016/j.chembiol.2008.01.006. [DOI] [PubMed] [Google Scholar]
- 25.Runguphan W., Qu X., O’Connor S.E. Integrating carbon-halogen bond formation into medicinal plant metabolism. Nature. 2010;468:461–464. doi: 10.1038/nature09524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fer W. Natural Products Chemistry the Marine Environme. SCIENCE. 1982;215:19. doi: 10.1126/science.215.4535.923. [DOI] [PubMed] [Google Scholar]
- 27.Tebben J., Motti C., Tapiolas D., Thomas-Hall P., Harder T. A Coralline Algal-Associated Bacterium, Pseudoalteromonas Strain J010, Yields Five New Korormicins and a Bromopyrrole. Mar. Drugs. 2014;12:2802–2815. doi: 10.3390/md12052802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yoshikawa K., Takadera T., Adachi K., Nishijima M., Sanc H. Korormicin, a novel antibiotic specifically active against marine gram-negative bacteria, produced by a marine bacterium. J. Antibiot. 1997;50:949–953. doi: 10.7164/antibiotics.50.949. [DOI] [PubMed] [Google Scholar]
- 29.Yoshikawa K., Nakayama Y., Hayashi M., Unemoto T., Mochida K. Korormicin, an antibiotic specific for gram-negative marine bacteria, strongly inhibits the respiratory chain-linked Na+-translocating NADH: quinone reductase from the marine Vibrio alginolyticus. J. Antibiot. 1999;52:182–185. doi: 10.7164/antibiotics.52.182. [DOI] [PubMed] [Google Scholar]
- 30.Egan S., James S., Holmström C., Kjelleberg S. Correlation between pigmentation and antifouling compounds produced by Pseudoalteromonas tunicata. Environ. Microbiol. 2002;4:433–442. doi: 10.1046/j.1462-2920.2002.00322.x. [DOI] [PubMed] [Google Scholar]
- 31.Egan S., James S., Kjelleberg S. Identification and Characterization of a Putative Transcriptional Regulator Controlling the Expression of Fouling Inhibitors in Pseudoalteromonas tunicata. Appl. Environ. Microbiol. 2002;68:372–378. doi: 10.1128/AEM.68.1.372-378.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Holmström C., James S., Neilan B.A., White D.C., Kjelleberg S. Pseudoalteromonas tunicata sp. nov., a bacterium that produces antifouling agents. Int. J. Syst. Bacteriol. 1998;48:1205–1212. doi: 10.1099/00207713-48-4-1205. [DOI] [PubMed] [Google Scholar]
- 33.Holmström C., Kjelleberg S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999;30:285–293. doi: 10.1016/S0168-6496(99)00063-X. [DOI] [PubMed] [Google Scholar]
- 34.Pinkerton D.M., Banwell M.G., Garson M.J., Kumar N., de Moraes M.O., Cavalcanti B.C., Barros F.W.A., Pessoa C. Antimicrobial and Cytotoxic Activities of Synthetically Derived Tambjamines C and E–J, BE-18591, and a Related Alkaloid from the Marine Bacterium Pseudoalteromonas tunicata. Chem. Biodivers. 2010;7:1311–1324. doi: 10.1002/cbdv.201000030. [DOI] [PubMed] [Google Scholar]
- 35.Burke C., Thomas T., Egan S., Kjelleberg S. The use of functional genomics for the identification of a gene cluster encoding for the biosynthesis of an antifungal tambjamine in the marine bacterium Pseudoalteromonas tunicata. Environ. Microbiol. 2007;9:814–818. doi: 10.1111/j.1462-2920.2006.01177.x. [DOI] [PubMed] [Google Scholar]
- 36.Wang H., Wang F., Zhu X., Yan Y., Yu X., Jiang P., Xing X.-H. Biosynthesis and characterization of violacein, deoxyviolacein and oxyviolacein in heterologous host, and their antimicrobial activities. Biochem. Eng. J. 2012;67:148–155. doi: 10.1016/j.bej.2012.06.005. [DOI] [Google Scholar]
- 37.Yang L.H., Xiong H., Lee O.O., Qi S.-H., Qian P.-Y. Effect of agitation on violacein production in Pseudoalteromonas luteoviolacea isolated from a marine sponge. Lett. Appl. Microbiol. 2007;44:625–630. doi: 10.1111/j.1472-765X.2007.02125.x. [DOI] [PubMed] [Google Scholar]
- 38.Hurdle J.G., O’Neill A.J., Chopra I. Anti-staphylococcal activity of indolmycin, a potential topical agent for control of staphylococcal infections. J. Antimicrob. Chemother. 2004;54:549–552. doi: 10.1093/jac/dkh352. [DOI] [PubMed] [Google Scholar]
- 39.Wratten S.J., Wolfe M.S., Andersen R.J., Faulkner D.J. Antibiotic Metabolites from a Marine Pseudomonad. Antimicrob. Agents Chemother. 1977;11:411–414. doi: 10.1128/AAC.11.3.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Del Castillo C.S., Wahid M.I., Takeshi Y., Taizo S. Isolation and inhibitory effect of anti-Vibrio substances from Pseudoalteromonas sp. A1-J11 isolated from the coastal sea water of Kagoshima Bay. Fish. Sci. 2008;74:174–179. doi: 10.1111/j.1444-2906.2007.01507.x. [DOI] [Google Scholar]
- 41.Sakata T., Sakaguchi K., Kakimoto D. Antiobiotic production by marine pigmented bacteria. I. Antibacterial effect of Alteromonas luteoviolaceus. Mem. Fac. Fish. Kagoshima Univ. 1982;31:243–250. [Google Scholar]
- 42.Sakata T., Sakaguchi K., Kakimota D. Antibiotic production by marine pigmented bacteria. II. Purification and characterization of antibiotic substance of Alteromonas luteoviolacea. Mem. Fac. Fish. 1986;35:29–37. [Google Scholar]
- 43.Zheng L., Chen H., Han X., Lin W., Yan X. Antimicrobial screening and active compound isolation from marine bacterium NJ6-3-1 associated with the sponge Hymeniacidon perleve. World J. Microbiol. Biotechnol. 2005;21:201–206. doi: 10.1007/s11274-004-3318-6. [DOI] [Google Scholar]
- 44.Yu M., Wang J., Tang K., Shi X., Wang S., Zhu W.-M., Zhang X.-H. Purification and characterization of antibacterial compounds of Pseudoalteromonas flavipulchra JG1. Microbiol. Read. Engl. 2012;158:835–842. doi: 10.1099/mic.0.055970-0. [DOI] [PubMed] [Google Scholar]
- 45.Fujioka M., Wada H. The bacterial oxidation of indole. Biochim. Biophys. Acta. 1968;158:70–78. doi: 10.1016/0304-4165(68)90073-1. [DOI] [PubMed] [Google Scholar]
- 46.Glover V., Halket J.M., Watkins P.J., Clow A., Goodwin B.L., Sandler M. Isatin: Identity with the purified endogenous monoamine oxidase inhibitor tribulin. J. Neurochem. 1988;51:656–659. doi: 10.1111/j.1471-4159.1988.tb01089.x. [DOI] [PubMed] [Google Scholar]
- 47.Kalinovskaya N.I., Ivanova E.P., Alexeeva Y.V., Gorshkova N.M., Kuznetsova T.A., Dmitrenok A.S., Nicolau D.V. Low-molecular-weight, biologically active compounds from marine Pseudoalteromonas species. Curr. Microbiol. 2004;48:441–446. doi: 10.1007/s00284-003-4240-0. [DOI] [PubMed] [Google Scholar]
- 48.Moree W.J., McConnell O.J., Nguyen D.D., Sanchez L.M., Yang Y.-L., Zhao X., Liu W.-T., Boudreau P.D., Srinivasan J., Atencio L., et al. Microbiota of Healthy Corals Are Active against Fungi in a Light-Dependent Manner. ACS Chem. Biol. 2014;9:2300–2308. doi: 10.1021/cb500432j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Andersen R.J., Wolfe M.S., Faulkner D.J. Autotoxic antibiotic production by a marine Chromobacterium. Mar. Biol. 1974;27:281–285. doi: 10.1007/BF00394363. [DOI] [Google Scholar]
- 50.Agarwal V., El Gamal A.A., Yamanaka K., Poth D., Kersten R.D., Schorn M., Allen E.E., Moore B.S. Biosynthesis of polybrominated aromatic organic compounds by marine bacteria. Nat. Chem. Biol. 2014;10:640–647. doi: 10.1038/nchembio.1564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Liu M., Hansen P.E., Lin X. Bromophenols in Marine Algae and Their Bioactivities. Mar. Drugs. 2011;9:1273–1292. doi: 10.3390/md9071273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zsolnai T. Versuche zur entdeckung neuer fungistatika-I: Phenol-derivate. Biochem. Pharmacol. 1960;5:1–19. doi: 10.1016/0006-2952(60)90002-2. [DOI] [Google Scholar]
- 53.Burkholder P.R., Pfister R.M., Leitz F.H. Production of a pyrrole antibiotic by a marine bacterium. Appl. Microbiol. 1966;14:649–653. doi: 10.1128/am.14.4.649-653.1966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lovell F.M. The Structure of a Bromine-Rich Marine Antibiotic. J. Am. Chem. Soc. 1966;88:4510–4511. doi: 10.1021/ja00971a040. [DOI] [Google Scholar]
- 55.Fehér D., Barlow R., McAtee J., Hemscheidt T.K. Highly Brominated Antimicrobial Metabolites from a Marine Pseudoalteromonas sp. J. Nat. Prod. 2010;73:1963–1966. doi: 10.1021/np100506z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ridley C.P., Lee H.Y., Khosla C. Evolution of polyketide synthases in bacteria. Proc. Natl. Acad. Sci. USA. 2008;105:4595–4600. doi: 10.1073/pnas.0710107105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Shen B., Cheng Y.-Q., Christenson S.D., Jiang H., Ju J., Kwon H.-J., Lim S.-K., Liu W., Nonaka K., Seo J.-W., et al. Polyketides. Volume 955. American Chemical Society; Washington, DC, USA: 2007. Polyketide Biosynthesis beyond the Type I, II, and III Polyketide Synthase Paradigms: A Progress Report; pp. 154–166. [Google Scholar]
- 58.Katsuyama Y., Ohnishi Y. Type III polyketide synthases in microorganisms. Methods Enzymol. 2012;515:359–377. doi: 10.1016/B978-0-12-394290-6.00017-3. [DOI] [PubMed] [Google Scholar]
- 59.Zhang H., Wang Y., Wu J., Skalina K., Pfeifer B.A. Complete Biosynthesis of Erythromycin A and Designed Analogs Using E. coli as a Heterologous Host. Chem. Biol. 2010;17:1232–1240. doi: 10.1016/j.chembiol.2010.09.013. [DOI] [PubMed] [Google Scholar]
- 60.Pickens L.B., Tang Y. Decoding and Engineering Tetracycline Biosynthesis. Metab. Eng. 2009;11:69–75. doi: 10.1016/j.ymben.2008.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Murphy A.C., Fukuda D., Song Z., Hothersall J., Cox R.J., Willis C.L., Thomas C.M., Simpson T.J. Engineered Thiomarinol Antibiotics Active against MRSA Are Generated by Mutagenesis and Mutasynthesis of Pseudoalteromonas SANK73390. Angew. Chem. Int. Ed. 2011;50:3271–3274. doi: 10.1002/anie.201007029. [DOI] [PubMed] [Google Scholar]
- 62.Isnansetyo A., Kamei Y. MC21-A, a Bactericidal Antibiotic Produced by a New Marine Bacterium, Pseudoalteromonas phenolica sp. nov. O-BC30T, against Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2003;47:480–488. doi: 10.1128/AAC.47.2.480-488.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Jiang Z., Boyd K.G., Mearns-spragg A., Adams D.R., Wright P.C., Burgess J.G. Two Diketopiperazines and One Halogenated Phenol from Cultures of the Marine Bacterium, Pseudoalteromonas luteoviolacea. Nat. Prod. Lett. 2006;14:435–440. doi: 10.1080/10575630008043781. [DOI] [Google Scholar]
- 64.Shiozawa H., Kagasaki T., Kinoshita T., Haruyama H., Domon H., Utsui Y., Kodama K., Takahashi S. Thiomarinol, a new hybrid antimicrobial antibiotic produced by a marine bacterium. Fermentation, isolation, structure, and antimicrobial activity. J. Antibiot. 1993;46:1834–1842. doi: 10.7164/antibiotics.46.1834. [DOI] [PubMed] [Google Scholar]
- 65.Shiozawa H., Shimada A., Takahashi S. Thiomarinols D, E, F and G, new hybrid antimicrobial antibiotics produced by a marine bacterium; isolation, structure, and antimicrobial activity. J. Antibiot. 1997;50:449–452. doi: 10.7164/antibiotics.50.449. [DOI] [PubMed] [Google Scholar]
- 66.Shiozawa H., Kagasaki T., Torikata A., Tanaka N., Fujimoto K., Hata T., Furukawa Y., Takahashi S. Thiomarinols B and C, new antimicrobial antibiotics produced by a marine bacterium. J. Antibiot. 1995;48:907–909. doi: 10.7164/antibiotics.48.907. [DOI] [PubMed] [Google Scholar]
- 67.Qin Z., Huang S., Yu Y., Deng H. Dithiolopyrrolone Natural Products: Isolation, Synthesis and Biosynthesis. Mar. Drugs. 2013;11:3970–3997. doi: 10.3390/md11103970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Strieker M., Tanović A., Marahiel M.A. Nonribosomal peptide synthetases: Structures and dynamics. Curr. Opin. Struct. Biol. 2010;20:234–240. doi: 10.1016/j.sbi.2010.01.009. [DOI] [PubMed] [Google Scholar]
- 69.Challis G.L., Naismith J.H. Structural aspects of non-ribosomal peptide biosynthesis. Curr. Opin. Struct. Biol. 2004;14:748–756. doi: 10.1016/j.sbi.2004.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Koglin A., Walsh C.T. Structural insights into nonribosomal peptide enzymatic assembly lines. Nat. Prod. Rep. 2009;26:987–1000. doi: 10.1039/b904543k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Desriac F., Jégou C., Balnois E., Brillet B., Chevalier P.L., Fleury Y. Antimicrobial Peptides from Marine Proteobacteria. Mar. Drugs. 2013;11:3632–3660. doi: 10.3390/md11103632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Nguyen D.D., Wu C.-H., Moree W.J., Lamsa A., Medema M.H., Zhao X., Gavilan R.G., Aparicio M., Atencio L., Jackson C., et al. MS/MS networking guided analysis of molecule and gene cluster families. Proc. Natl. Acad. Sci. USA. 2013;110:2611–2620. doi: 10.1073/pnas.1303471110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Speitling M., Smetanina O.F., Kuznetsova T.A., Laatsch H. Bromoalterochromides A and A’, unprecedented chromopeptides from a marine Pseudoalteromonas maricaloris strain KMM 636T. J. Antibiot. 2007;60:36–42. doi: 10.1038/ja.2007.5. [DOI] [PubMed] [Google Scholar]
- 74.Sobolevskaya M.P., Smetanina O.F., Speitling M., Shevchenko L.S., Dmitrenok P.S., Laatsch H., Kuznetsova T.A., Ivanova E.P., Elyakov G.B. Controlling production of brominated cyclic depsipeptides by Pseudoalteromonas maricaloris KMM 636T. Lett. Appl. Microbiol. 2005;40:243–248. doi: 10.1111/j.1472-765X.2005.01635.x. [DOI] [PubMed] [Google Scholar]
- 75.Rungprom W., Siwu E.R.O., Lambert L.K., Dechsakulwatana C., Barden M.C., Kokpol U., Blanchfield J.T., Kita M., Garson M.J. Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron. 2008;64:3147–3152. doi: 10.1016/j.tet.2008.01.089. [DOI] [Google Scholar]
- 76.Nakano M., Danowski T.S., Weitzel W. The technical assistance of D. R. Crystalline Mammalian l-Amino Acid Oxidase from Rat Kidney Mitochondria. J. Biol. Chem. 1966;241:2075–2083. [PubMed] [Google Scholar]
- 77.Yu Z., Wang J., Lin J., Zhao M., Qiu J. Exploring regulation genes involved in the expression of l-amino acid oxidase in Pseudoalteromonas sp. Rf-1. PLoS ONE. 2015;10:129. doi: 10.1371/journal.pone.0122741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gómez D., Espinosa E., Bertazzo M., Lucas-Elío P., Solano F., Sanchez-Amat A. The macromolecule with antimicrobial activity synthesized by Pseudoalteromonas luteoviolacea strains is an l-amino acid oxidase. Appl. Microbiol. Biotechnol. 2008;79:925–930. doi: 10.1007/s00253-008-1499-x. [DOI] [PubMed] [Google Scholar]
- 79.James S.G., Holmström C., Kjelleberg S. Purification and characterization of a novel antibacterial protein from the marine bacterium D2. Appl. Environ. Microbiol. 1996;62:2783–2788. doi: 10.1128/aem.62.8.2783-2788.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Chen W.M., Lin C.Y., Chen C.A., Wang J.T., Sheu S.Y. Involvement of an l-amino acid oxidase in the activity of the marine bacterium Pseudoalteromonas flavipulchra against methicillin-resistant Staphylococcus aureus. Enzyme Microb. Technol. 2010;47:52–58. doi: 10.1016/j.enzmictec.2010.03.008. [DOI] [Google Scholar]
- 81.Yu M., Tang K., Shi X., Zhang X.-H. Genome Sequence of Pseudoalteromonas flavipulchra JG1, a Marine Antagonistic Bacterium with Abundant Antimicrobial Metabolites. J. Bacteriol. 2012;194:3735. doi: 10.1128/JB.00598-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.McCarthy S.A., Johnson R.M., Kakimoto D. Characterization of an antibiotic produced by Alteromonas luteoviolacea Gauthier 1982, 85 isolated from Kinko Bay, Japan. J. Appl. Bacteriol. 1994;77:426–432. doi: 10.1111/j.1365-2672.1994.tb03445.x. [DOI] [PubMed] [Google Scholar]
- 83.Defer D., Bourgougnon N., Fleury Y. Screening for antibacterial and antiviral activities in three bivalve and two gastropod marine molluscs. Aquaculture. 2009;293:1–7. doi: 10.1016/j.aquaculture.2009.03.047. [DOI] [Google Scholar]
- 84.Aranda C.P., Valenzuela C., Barrientos J., Paredes J., Leal P., Maldonado M., Godoy F.A., Osorio C.G. Bacteriostatic anti-Vibrio parahaemolyticus activity of Pseudoalteromonas sp. strains DIT09, DIT44 and DIT46 isolated from Southern Chilean intertidal Perumytilus purpuratus. World J. Microbiol. Biotechnol. 2012;28:2365–2374. doi: 10.1007/s11274-012-1044-z. [DOI] [PubMed] [Google Scholar]
- 85.Barja J.L., Lemos M.L., Toranzo A.E. Purification and characterization of an antibacterial substance produced by a marine Alteromonas species. Antimicrob. Agents Chemother. 1989;33:1674–1679. doi: 10.1128/AAC.33.10.1674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Chelossi E., Milanese M., Milano A., Pronzato R., Riccardi G. Characterisation and antimicrobial activity of epibiotic bacteria from Petrosia ficiformis (Porifera, Demospongiae) J. Exp. Mar. Biol. Ecol. 2004;309:21–33. doi: 10.1016/j.jembe.2004.03.006. [DOI] [Google Scholar]
- 87.Hentschel U., Schmid M., Wagner M., Fieseler L., Gernert C., Hacker J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001;35:305–312. doi: 10.1111/j.1574-6941.2001.tb00816.x. [DOI] [PubMed] [Google Scholar]
- 88.Holmström C., Rittschof D., Kjelleberg S. Inhibition of Settlement by Larvae of Balanus amphitrite and Ciona intestinalis by a Surface-Colonizing Marine Bacterium. Appl. Environ. Microbiol. 1992;58:2111–2115. doi: 10.1128/aem.58.7.2111-2115.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Egan S., James S., Holmström C., Kjelleberg S. Inhibition of algal spore germination by the marine bacterium Pseudoalteromonas tunicata. FEMS Microbiol. Ecol. 2001;35:67–73. doi: 10.1111/j.1574-6941.2001.tb00789.x. [DOI] [PubMed] [Google Scholar]
- 90.Buck J.D., Meyers S.P. In vitro inhibition of Rhodotorula minuta by a variant of the marine bacterium, Pseudomonas piscicida. Helgoländer Wiss. Meeresunters. 1966;13:171–180. doi: 10.1007/BF01612662. [DOI] [Google Scholar]
- 91.Longeon A., Peduzzi J., Barthelemy M., Corre S., Nicolas J.-L., Guyot M. Purification and Partial Identification of Novel Antimicrobial Protein from Marine Bacterium Pseudoalteromonas Species Strain X153. Mar. Biotechnol. 2004;6:633–641. doi: 10.1007/s10126-004-3009-1. [DOI] [PubMed] [Google Scholar]
- 92.Gauthier M.J. Alteromonas citrea, a New Gram-Negative, Yellow-Pigmented Species from Seawater. Int. J. Syst. Bacteriol. 1977;27:349–354. doi: 10.1099/00207713-27-4-349. [DOI] [Google Scholar]
- 93.Gauthier M.J., Breittmayer V.A. A New Antibiotic-Producing Bacterium from Seawater: Alteromonas aurantia sp. nov. Int. J. Syst. Bacteriol. 1979;29:366–372. doi: 10.1099/00207713-29-4-366. [DOI] [Google Scholar]
- 94.Scheffler R.J., Colmer S., Tynan H., Demain A.L., Gullo V.P. Antimicrobials, drug discovery, and genome mining. Appl. Microbiol. Biotechnol. 2013;97:969–978. doi: 10.1007/s00253-012-4609-8. [DOI] [PubMed] [Google Scholar]
- 95.Katz L., Baltz R.H. Natural product discovery: past, present, and future. J. Ind. Microbiol. Biotechnol. 2016;43:155–176. doi: 10.1007/s10295-015-1723-5. [DOI] [PubMed] [Google Scholar]
- 96.Weber T., Blin K., Duddela S., Krug D., Kim H.U., Bruccoleri R., Lee S.Y., Fischbach M.A., Müller R., Wohlleben W., et al. antiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015;43:237–243. doi: 10.1093/nar/gkv437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ziemert N., Podell S., Penn K., Badger J.H., Allen E., Jensen P.R. The Natural Product Domain Seeker NaPDoS: A Phylogeny Based Bioinformatic Tool to Classify Secondary Metabolite Gene Diversity. PLoS ONE. 2012;7:129. doi: 10.1371/journal.pone.0034064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Anand S., Prasad M.V.R., Yadav G., Kumar N., Shehara J., Ansari M.Z., Mohanty D. SBSPKS: Structure based sequence analysis of polyketide synthases. Nucleic Acids Res. 2010 doi: 10.1093/nar/gkq340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Li M.H., Ung P.M., Zajkowski J., Garneau-Tsodikova S., Sherman D.H. Automated genome mining for natural products. BMC Bioinform. 2009;10:129. doi: 10.1186/1471-2105-10-185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Stachelhaus T., Mootz H.D., Marahiel M.A. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 1999;6:493–505. doi: 10.1016/S1074-5521(99)80082-9. [DOI] [PubMed] [Google Scholar]
- 101.Röttig M., Medema M.H., Blin K., Weber T., Rausch C., Kohlbacher O. NRPSpredictor2—A web server for predicting NRPS adenylation domain specificity. Nucleic Acids Res. 2011 doi: 10.1093/nar/gkr323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Khayatt B.I., Overmars L., Siezen R.J., Francke C. Classification of the Adenylation and Acyl-Transferase Activity of NRPS and PKS Systems Using Ensembles of Substrate Specific Hidden Markov Models. PLoS ONE. 2013;8:129. doi: 10.1371/journal.pone.0062136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Van Heel A.J., de Jong A., Montalbán-López M., Kok J., Kuipers O.P. BAGEL3: Automated identification of genes encoding bacteriocins and (non-)bactericidal posttranslationally modified peptides. Nucleic Acids Res. 2013;41:W448–W453. doi: 10.1093/nar/gkt391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Amoutzias G.D., Chaliotis A., Mossialos D. Discovery Strategies of Bioactive Compounds Synthesized by Nonribosomal Peptide Synthetases and Type-I Polyketide Synthases Derived from Marine Microbiomes. Mar. Drugs. 2016;14:80. doi: 10.3390/md14040080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chávez R., Fierro F., García-Rico R.O., Vaca I. Filamentous fungi from extreme environments as a promising source of novel bioactive secondary metabolites. Front. Microbiol. 2015;6 doi: 10.3389/fmicb.2015.00903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Tietz J., Mitchell D. Using Genomics for Natural Product Structure Elucidation. Curr. Top. Med. Chem. 2015;16:1645–1694. doi: 10.2174/1568026616666151012111439. [DOI] [PubMed] [Google Scholar]
- 107.Zerikly M., Challis G.L. Strategies for the Discovery of New Natural Products by Genome Mining. ChemBioChem. 2009;10:625–633. doi: 10.1002/cbic.200800389. [DOI] [PubMed] [Google Scholar]
- 108.Challis G.L. Genome Mining for Novel Natural Product Discovery. J. Med. Chem. 2008;51:2618–2628. doi: 10.1021/jm700948z. [DOI] [PubMed] [Google Scholar]
- 109.Zarins-Tutt J.S. Ph.D. Thesis. University of St Andrews; Scotland, UK: 2015. Gene Mining of Biosynthesis Genes and Biosynthetic Manipulation of Marine Bacteria for the Production of New Antibiotic Candidates. [Google Scholar]
- 110.Machado H., Sonnenschein E.C., Melchiorsen J., Gram L. Genome mining reveals unlocked bioactive potential of marine Gram-negative bacteria. BMC Genom. 2015;16:129. doi: 10.1186/s12864-015-1365-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Papaleo M.C., Fondi M., Maida I., Perrin E., Lo Giudice A., Michaud L., Mangano S., Bartolucci G., Romoli R., Fani R. Sponge-associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012;30:272–293. doi: 10.1016/j.biotechadv.2011.06.011. [DOI] [PubMed] [Google Scholar]
- 112.Maansson M., Vynne N.G., Klitgaard A., Nybo J.L., Melchiorsen J., Nguyen D.D., Sanchez L.M., Ziemert N., Dorrestein P.C., Andersen M.R., et al. An Integrated Metabolomic and Genomic Mining Workflow to Uncover the Biosynthetic Potential of Bacteria. mSystems. 2016;1:e00028–15. doi: 10.1128/mSystems.00028-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wang P., Yu Z., Li B., Cai X., Zeng Z., Chen X., Wang X. Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas. Microb. Cell Fact. 2015;14:1–11. doi: 10.1186/s12934-015-0194-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Duilio A., Tutino M.L., Marino G. Recombinant protein production in Antarctic Gram-negative bacteria. Recomb. Gene Expr. Rev. Protoc. 2004 doi: 10.1385/1-59259-774-2:225. [DOI] [PubMed] [Google Scholar]
- 115.Giuliani M., Parrilli E., Pezzella C., Rippa V., Duilio A., Marino G., Tutino M.L. A Novel Strategy for the Construction of Genomic Mutants of the Antarctic Bacterium Pseudoalteromonas haloplanktis TAC125. In: Lorence A., editor. Recombinant Gene Expression. Volume 824. Humana Press; Totowa, NJ, USA: 2012. pp. 219–233. [DOI] [PubMed] [Google Scholar]
- 116.Yu Z.-C., Zhao D.-L., Ran L.-Y., Mi Z.-H., Wu Z.-Y., Pang X., Zhang X.-Y., Su H.-N., Shi M., Song X.-Y., et al. Development of a genetic system for the deep-sea psychrophilic bacterium Pseudoalteromonas sp. SM9913. Microb. Cell Fact. 2014;13 doi: 10.1186/1475-2859-13-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Miranda C.D., Rojas R., Geisse J., Romero J., González-Rocha G. Scallop larvae hatcheries as source of bacteria carrying genes encoding for non-enzymatic phenicol resistance. Mar. Pollut. Bull. 2015;95:173–182. doi: 10.1016/j.marpolbul.2015.04.026. [DOI] [PubMed] [Google Scholar]
- 118.Bakunina I.Y., Nedashkovskaya O.I., Alekseeva S.A., Ivanova E.P., Romanenko L.A., Gorshkova N.M., Isakov V.V., Zvyagintseva T.N., Mikhailov V.V. Degradation of Fucoidan by the Marine Proteobacterium Pseudoalteromonas citrea. Microbiology. 2002;71:41–47. doi: 10.1023/A:1017994131769. [DOI] [PubMed] [Google Scholar]
- 119.Ivanova E.P., Kurilenko V.V., Kurilenko A.V., Gorshkova N.M., Shubin F.N., Nicolau D.V., Chelomin V.P. Tolerance to Cadmium of Free-Living and Associated with Marine Animals and Eelgrass Marine Gamma-Proteobacteria. Curr. Microbiol. 2002;44:357–362. doi: 10.1007/s00284-001-0017-5. [DOI] [PubMed] [Google Scholar]
- 120.Ivanova E.P., Gorshkova N.M., Kurilenko V.V. Tolerance of Marine Proteobacteria of the Genera Pseudoalteromonas and Alteromonas to Heavy Metals. Microbiology. 2001;70:239–241. doi: 10.1023/A:1010498017807. [DOI] [PubMed] [Google Scholar]
- 121.Ivanova E.P., Mikhailov V.V. A New Family, Alteromonadaceae fam. nov., Including Marine Proteobacteria of the Genera Alteromonas, Pseudoalteromonas, Idiomarina, and Colwellia. Microbiology. 2001;70:10–17. doi: 10.1023/A:1004876301036. [DOI] [PubMed] [Google Scholar]
- 122.Dieckmann R., Graeber I., Kaesler I., Szewzyk U., von Döhren H. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by Intact-Cell-MALDI-TOF mass spectrometry (ICM-MS) Appl. Microbiol. Biotechnol. 2004;67:539–548. doi: 10.1007/s00253-004-1812-2. [DOI] [PubMed] [Google Scholar]
- 123.Romanenko L.A., Uchino M., Kalinovskaya N.I., Mikhailov V.V. Isolation, phylogenetic analysis and screening of marine mollusc-associated bacteria for antimicrobial, hemolytic and surface activities. Microbiol. Res. 2008;163:633–644. doi: 10.1016/j.micres.2006.10.001. [DOI] [PubMed] [Google Scholar]
- 124.Jin G., Wang S., Yu M., Yan S., Zhang X.-H. Identification of a marine antagonistic strain JG1 and establishment of a polymerase chain reaction detection technique based on the gyrB gene. Aquac. Res. 2010;41:1867–1874. doi: 10.1111/j.1365-2109.2010.02591.x. [DOI] [Google Scholar]
- 125.Hayashida-Soiza G., Uchida A., Mori N., Kuwahara Y., Ishida Y. Purification and characterization of antibacterial substances produced by a marine bacterium Pseudoalteromonas haloplanktis strain. J. Appl. Microbiol. 2008;105:1672–1677. doi: 10.1111/j.1365-2672.2008.03878.x. [DOI] [PubMed] [Google Scholar]
- 126.Ivanova E.P., Sawabe T., Alexeeva Y.V., Lysenko A.M., Gorshkova N.M., Hayashi K., Zukova N.V., Christen R., Mikhailov V.V. Pseudoalteromonas issachenkonii sp. nov., a bacterium that degrades the thallus of the brown alga Fucus evanescens. Int. J. Syst. Evol. Microbiol. 2002;52:229–234. doi: 10.1099/00207713-52-1-229. [DOI] [PubMed] [Google Scholar]
- 127.Pujalte M.J., Ortigosa M., Macián M.C., Garay E. Aerobic and facultative anaerobic heterotrophic bacteria associated to Mediterranean oysters and seawater. Int. Microbiol. 2010;2:259–266. [PubMed] [Google Scholar]
- 128.Huggett M.J., Williamson J.E., de Nys R., Kjelleberg S., Steinberg P.D. Larval settlement of the common Australian sea urchin Heliocidaris erythrogramma in response to bacteria from the surface of coralline algae. Oecologia. 2006;149:604–619. doi: 10.1007/s00442-006-0470-8. [DOI] [PubMed] [Google Scholar]
- 129.Ivanova E.P., Shevchenko L.S., Sawabe T., Lysenko A.M., Svetashev V.I., Gorshkova N.M., Satomi M., Christen R., Mikhailov V.V. Pseudoalteromonas maricaloris sp. nov., isolated from an Australian sponge, and reclassification of [Pseudoalteromonas aurantia] NCIMB 2033 as Pseudoalteromonas flavipulchra sp. nov. Int. J. Syst. Evol. Microbiol. 2002;52:263–271. doi: 10.1099/00207713-52-1-263. [DOI] [PubMed] [Google Scholar]
- 130.El-Tarabily K.A., Youssef T. Improved growth performance of the mangrove Avicennia marina seedlings using a 1-aminocyclopropane-1-carboxylic acid deaminase-producing isolate of Pseudoalteromonas maricaloris. Plant Growth Regul. 2011;65:473–483. doi: 10.1007/s10725-011-9618-6. [DOI] [Google Scholar]
- 131.Microbial Community of Black Band Disease on Infection, Healthy, and Dead Part of Scleractinian Montipora sp. Colony at Seribu Islands, Indonesia. [(accessed on 15 January 2016)]. Available online: http://www.academia.edu/11707009/MICROBIAL_COMMUNITY_OF_BLACK_BAND_DISEASE_ON_INFECTION_HEALTHY_AND_DEAD_PART_OF_SCLERACTINIAN_Montipora_sp._COLONY_AT_SERIBU_ISLANDS_INDONESIA.
- 132.Zheng L., Yan X., Han X., Chen H., Lin W., Lee F.S.C., Wang X. Identification of norharman as the cytotoxic compound produced by the sponge (Hymeniacidon perleve)-associated marine bacterium Pseudoalteromonas piscicida and its apoptotic effect on cancer cells. Biotechnol. Appl. Biochem. 2006;44:135–142. doi: 10.1042/BA20050176. [DOI] [PubMed] [Google Scholar]
- 133.Nelson E.J., Ghiorse W.C. Isolation and identification of Pseudoalteromonas piscicida strain Cura-d associated with diseased damselfish (Pomacentridae) eggs. J. Fish Dis. 1999;22:253–260. doi: 10.1046/j.1365-2761.1999.00168.x. [DOI] [Google Scholar]
- 134.Zhang X., Nakahara T., Miyazaki M., Nogi Y., Taniyama S., Arakawa O., Inoue T., Kudo T. Diversity and function of aerobic culturable bacteria in the intestine of the sea cucumber Holothuria leucospilota. J. Gen. Appl. Microbiol. 2012;58:447–456. doi: 10.2323/jgam.58.447. [DOI] [PubMed] [Google Scholar]
- 135.Singh R.P., Kumari P., Reddy C.R.K. Antimicrobial compounds from seaweeds-associated bacteria and fungi. Appl. Microbiol. Biotechnol. 2015;99:1571–1586. doi: 10.1007/s00253-014-6334-y. [DOI] [PubMed] [Google Scholar]
- 136.Ivanova E.P., Gorshkova N.M., Sawabe T. Pseudoalteromonas ruthenica sp. nov., isolated from marine invertebrates. Int. J. Syst. Evol. Microbiol. 2002;52:235–240. doi: 10.1099/00207713-52-1-235. [DOI] [PubMed] [Google Scholar]
- 137.Li J., Tan B., Mai K. Isolation and identification of a bacterium from marine shrimp digestive tract: A new degrader of starch and protein. J. Ocean Univ. China. 2011;10:287–292. doi: 10.1007/s11802-011-1849-7. [DOI] [Google Scholar]
- 138.Egan S., Thomas T., Holmström C., Kjelleberg S. Phylogenetic relationship and antifouling activity of bacterial epiphytes from the marine alga Ulva lactuca. Environ. Microbiol. 2000;2:343–347. doi: 10.1046/j.1462-2920.2000.00107.x. [DOI] [PubMed] [Google Scholar]
- 139.Wilson G.S., Raftos D.A., Corrigan S.L., Nair S.V. Diversity and antimicrobial activities of surface-attached marine bacteria from Sydney Harbour, Australia. Microbiol. Res. 2010;165:300–311. doi: 10.1016/j.micres.2009.05.007. [DOI] [PubMed] [Google Scholar]
- 140.Pujalte M.J., Sitjà-Bobadilla A., Macián M.C., Álvarez-Pellitero P., Garay E. Occurrence and virulence of Pseudoalteromonas spp. in cultured gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Molecular and phenotypic characterisation of P. undina strain U58. Aquaculture. 2007;271:47–53. doi: 10.1016/j.aquaculture.2007.06.015. [DOI] [Google Scholar]
- 141.Radjasa O.K., Kencana D.S., Sabdono A., Hutagalung R.A., Lestari E.S. Antibacterial Activity of Marine Bacteria Associated with sponge Aaptos sp. against Multi Drugs Resistant (MDR) strains. J. Mat. Sains. 2009;12:147–152. [Google Scholar]
- 142.Flemer B., Kennedy J., Margassery L.M., Morrissey J.P., O’Gara F., Dobson A.D.W. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol. 2012;112:289–301. doi: 10.1111/j.1365-2672.2011.05211.x. [DOI] [PubMed] [Google Scholar]
- 143.Shnit-Orland M., Sivan A., Kushmaro A. Antibacterial Activity of Pseudoalteromonas in the Coral Holobiont. Microb. Ecol. 2012;64:851–859. doi: 10.1007/s00248-012-0086-y. [DOI] [PubMed] [Google Scholar]
- 144.Chen Y.-H., Kuo J., Sung P.-J., Chang Y.-C., Lu M.-C., Wong T.-Y., Liu J.-K., Weng C.-F., Twan W.-H., Kuo F.-W. Isolation of marine bacteria with antimicrobial activities from cultured and field-collected soft corals. World J. Microbiol. Biotechnol. 2012;28:3269–3279. doi: 10.1007/s11274-012-1138-7. [DOI] [PubMed] [Google Scholar]
- 145.Defer D., Desriac F., Henry J., Bourgougnon N., Baudy-Floc’h M., Brillet B., Le Chevalier P., Fleury Y. Antimicrobial peptides in oyster hemolymph: The bacterial connection. Fish Shellfish Immunol. 2013;34:1439–1447. doi: 10.1016/j.fsi.2013.03.357. [DOI] [PubMed] [Google Scholar]
- 146.Desriac F., Chevalier P., Brillet B., Leguerinel I., Thuillier B., Paillard C., Fleury Y. Exploring the hologenome concept in marine bivalvia: Haemolymph microbiota as a pertinent source of probiotics for aquaculture. FEMS Microbiol. Lett. 2014;350:107–116. doi: 10.1111/1574-6968.12308. [DOI] [PubMed] [Google Scholar]
- 147.Goulden E.F., Hall M.R., Pereg L.L., Høj L. Identification of an Antagonistic Probiotic Combination Protecting Ornate Spiny Lobster (Panulirus ornatus) Larvae against Vibrio owensii Infection. PLoS ONE. 2012;7:129. doi: 10.1371/journal.pone.0039667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Lokmer A., Wegner K.M. Hemolymph microbiome of Pacific oysters in response to temperature, temperature stress and infection. ISME J. 2014;9:670–682. doi: 10.1038/ismej.2014.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Carillo S., Pieretti G., Parrilli E., Tutino M.L., Gemma S., Molteni M., Lanzetta R., Parrilli M., Corsaro M.M. Structural Investigation and Biological Activity of the Lipooligosaccharide from the Psychrophilic Bacterium Pseudoalteromonas haloplanktis TAB 23. Chem. Eur. J. 2011;17:7053–7060. doi: 10.1002/chem.201100579. [DOI] [PubMed] [Google Scholar]
- 150.Maaetoft-Udsen K., Vynne N., Heegaard P.M., Gram L., Frøkiær H. Pseudoalteromonas strains are potent immunomodulators owing to low-stimulatory LPS. Innate Immun. 2013;19:160–173. doi: 10.1177/1753425912455208. [DOI] [PubMed] [Google Scholar]
- 151.Corsaro M.M., Piaz F.D., Lanzetta R., Parrilli M. Lipid A structure of Pseudoalteromonas haloplanktis TAC 125: Use of electrospray ionization tandem mass spectrometry for the determination of fatty acid distribution. J. Mass Spectrom. 2002;37:481–488. doi: 10.1002/jms.304. [DOI] [PubMed] [Google Scholar]
- 152.Krasikova I.N., Kapustina N.V., Isakov V.V., Gorshkova N.M., Solov’eva T.F. Elucidation of Structure of Lipid A from the Marine Gram-Negative Bacterium Pseudoalteromonas haloplanktis ATCC 14393T. Russ. J. Bioorg. Chem. 2004;30:367–373. doi: 10.1023/B:RUBI.0000037264.53294.da. [DOI] [PubMed] [Google Scholar]
- 153.Gardères J., Bedoux G., Koutsouveli V., Crequer S., Desriac F., Pennec G.L. Lipopolysaccharides from Commensal and Opportunistic Bacteria: Characterization and Response of the Immune System of the Host Sponge Suberites domuncula. Mar. Drugs. 2015;13:4985–5006. doi: 10.3390/md13084985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Silipo A., Molinaro A., Nazarenko E.L., Gorshkova R.P., Ivanova E.P., Lanzetta R., Parrilli M. The O-chain structure from the LPS of marine halophilic bacterium Pseudoalteromonas carrageenovora-type strain IAM 12662T. Carbohydr. Res. 2005;340:2693–2697. doi: 10.1016/j.carres.2005.09.007. [DOI] [PubMed] [Google Scholar]