

Abstract
Key points
Hepatic insulin resistance in patients with obesity or type 2 diabetes has been suggested to result from hepatic mitochondrial dysfunction.
High‐resolution respirometry (HRR) can be used to assess oxidative phosphorylation by measuring the mitochondrial oxygen consumption rate in the individual complexes of the mitochondria.
By using HRR, the present study demonstrates no difference in hepatic mitochondrial oxidative phosphorylation among subjects with obesity with or without type 2 diabetes and non‐obese controls.
Furthermore, the amount of mitochondria, assessed by the citrate synthase activity, is not different between the three groups.
Together the present findings indicate that hepatic mitochondrial oxidative phosphorylation capacity is not impaired in patients with obesity or type 2 diabetes.
Abstract
Obese patients with type 2 diabetes (T2DM) and without type 2 diabetes (OB) are characterized by high hepatic lipid content and hepatic insulin resistance. This may be linked to impaired hepatic mitochondrial oxidative phosphorylation (OXPHOS) capacity. The aim of the present study was to investigate and compare hepatic mitochondrial OXPHOS capacity in T2DM, OB and non‐obese controls (CON). Seventeen obese patients (nine OB and eight T2DM) and six CON patients had perioperative liver biopsies taken. Samples were divided into three parts to measure (1) complex I, II and IV linked respiration, (2) citrate synthase (CS) activity and (3) lipid droplet (LD) size and area (% of total tissue area filled by LDs). State 3 respiration of complex I, II and IV and the CS activity did not differ in OB, T2DM and CON. LD size was significantly higher in T2DM compared with CON, and LD area tended (P = 0.10) to be higher in T2DM and OB compared with CON. The present findings indicate that hepatic OXPHOS capacity is not different in patients with markedly different weight and glycaemic control. Furthermore, the results do not support impaired hepatic mitochondrial respiratory capacity playing a major role in the development of obesity‐induced type 2 diabetes.
Key points
Hepatic insulin resistance in patients with obesity or type 2 diabetes has been suggested to result from hepatic mitochondrial dysfunction.
High‐resolution respirometry (HRR) can be used to assess oxidative phosphorylation by measuring the mitochondrial oxygen consumption rate in the individual complexes of the mitochondria.
By using HRR, the present study demonstrates no difference in hepatic mitochondrial oxidative phosphorylation among subjects with obesity with or without type 2 diabetes and non‐obese controls.
Furthermore, the amount of mitochondria, assessed by the citrate synthase activity, is not different between the three groups.
Together the present findings indicate that hepatic mitochondrial oxidative phosphorylation capacity is not impaired in patients with obesity or type 2 diabetes.
Abbreviations
- ALAT
alanine transaminase
- ASAT
Aspartate transaminase
- BMI
body mass index
- CI–V
mitochondrial complexes I–V
- CI + IIE
electron transfer system capacity
- CIL
state 2 leak respiration
- CIP
state 3 complex I OXPHOS capacity
- CIIP
state 3 complex II OXPHOS capacity
- CIVP
state 3 complex IV OXPHOS capacity
- CON
control subjects
- CS
citrate synthase
- CytC
cytochrome C
- CIP + ETFP
state 3 lipid OXPHOS capacity
- ETS
electron transport system
- HbA1c
glycated heamoglobin A1c
- HRR
high‐resolution respirometry
- LD
lipid droplet
- MRS
magnetic resonance spectroscopy
- NAFLD
non‐alcoholic fatty liver disease
- NASH
non‐alchoholic steatohepatitis
- OB
obese subjects without type 2 diabetes
- OXPHOS
mitochondrial oxidative phosphorylation
- PAS
periodic acid–Schiff
- RYGB
Roux‐en‐Y gastric bypass
- SDH
succinate dehydrogenase
- T2DM
subjects with type 2 diabetes
Introduction
The majority of individuals with obesity are characterized by ectopic lipid accumulation in non‐adipose tissues such as skeletal muscle and the liver (Lionetti et al. 2009). In the liver, a high lipid content, which is also characteristic of patients with type 2 diabetes, has been linked to an impaired ability of insulin to inhibit hepatic glucose production, hepatic insulin resistance, in most (Bonadonna et al. 1990; Kotronen et al. 2008; Seppala‐Lindroos et al. 2002) but not all studies (Amaro et al. 2010; Visser et al. 2011). Thus, patients with obesity and type 2 diabetes require a higher plasma insulin concentration to maintain a normal hepatic glucose production. As type 2 diabetes progresses, the insulin‐induced inhibition of hepatic glucose production gradually fails and hyperglycaemia worsens.
Hepatic lipid accumulation may be the result of decreased mitochondrial lipid oxidation due to mitochondrial dysfunction (Lowell & Shulman, 2005), and lower hepatic ATP content and production have been found in patients with type 2 diabetes compared with matched controls by 31P magnetic resonance spectroscopy (MRS) (Szendroedi et al. 2009; Schmid et al. 2011).
ATP production is the final step of oxidative phosphorylation (OXPHOS). It is preceded by a series of energy releasing redox reactions in the mitochondrial complexes I–V (CI–V) of the electron transport system (ETS). The energy created drives proton efflux in CI, CIII and CIV building an electrochemical membrane potential across the inner mitochondrial membrane, which drives the formation of ATP. Obviously OXPHOS is a complex process and therefore 31P MRS is methodologically limited by the inability to assess individual parts of the ETS. Furthermore, 31P MRS is not mitochondria specific and assesses all ATP generating pathways in the cell.
High‐resolution respirometry (HRR) can be used to assess OXPHOS by measuring mitochondrial oxygen consumption rate in the individual complexes of the mitochondria (Pesta & Gnaiger, 2012). Currently, hepatic mitochondrial OXPHOS has not been investigated in patients with type 2 diabetes. Thus, there is lack of detailed information regarding hepatic mitochondrial OXPHOS capacity and its role in type 2 diabetes.
With the present study we aimed to investigate hepatic OXPHOS capacity in obese patients with and without type 2 diabetes and in a non‐obese control group. We hypothesized that hepatic OXPHOS capacity would be impaired in obese patients compared with controls. Secondly, in line with the prior 31P MRS results, we expected that hepatic OXPHOS capacity would be impaired in patients with type 2 diabetes compared with weight‐matched controls.
Methods
Seventeen patients (eight with type 2 diabetes (T2DM) and nine without type 2 diabetes (OB)) scheduled for a laparoscopic Roux‐en‐Y gastric bypass (RYGB) operation were recruited. Inclusion criteria were as follows: age, 18–60 years; and body mass index (BMI) > 40 kg m−2 or > 35 kg m−2 with obesity‐related co‐morbidities (e.g. type 2 diabetes). In addition, six patients scheduled for laparoscopic cholecystectomy were recruited as a non‐obese control group (CON). Patient self‐reported alcohol consumption was < 20 g day−1. Patients suffering from cardiovascular disease, polycystic ovary syndrome, thyroid disease and other diseases demanding medication that could affect the test results were excluded. All patients gave informed signed consent. The study was approved by the Copenhagen Science Ethics Committee (journal no. H‐C‐2009‐050) and performed according to the Declaration of Helsinki. The present study is part of a larger project (GASMITO) investigating the metabolic and psycho‐social effects of gastric bypass surgery (Wimmelmann et al. 2014, 2015). Anthropometric measures, HbA1c and fasting glucose concentrations from some of the subjects have previously been published (Lund et al. 2015; Hansen et al. 2016).
Fasting plasma glucose, alanine transaminase (ALAT), aspartate transaminase (ASAT), bilirubin and HbA1c were assessed prior to surgery. During the scheduled laparoscopic RYGB (OB and T2DM) and during the scheduled laparoscopic cholecystectomy (CON) subjects had a wedge liver biopsy from lobus dexter margo inferior taken. The biopsy was immediately divided in two equal‐sized tissue blocks. The first part was transferred to a chilled mitochondrial preservation buffer (described in detail in Boushel et al. 2007) for later analysis of mitochondrial OXPHOS capacity. The second part was fixed by immersion into 2% paraformaldehyde supplemented with 0.15% picric acid and embedded in paraffin. After arrival at the lab, a portion of the first part of the biopsy was frozen in liquid nitrogen and stored at −80°C for later analysis of citrate synthase (CS) activity. The remaining part of the biopsy was used for analysis of mitochondrial OXPHOS capacity. The thin outer fascia was removed with two thin forceps and the tissue was carefully dissected in 2 × 2 mm tissue blocks while in chilled mitochondrial preservation buffer. The tissue was washed in respiration medium for 10 min, and 2–3 mg tissue was transferred to the respirometer (Oxygraph‐2k, Oroboros Instruments, Innsbruck, Austria; described in detail in Boushel et al. 2007), and two respiratory protocols were applied. The liver tissue was not permeabilized prior to doing HRR since pilot experiments using either saponin or digitonin as permeabilization agent showed no change in flux compared with no permeabilization (data not shown).
In protocol I, state 2 leak respiration (CIL) was assessed in the presence of malate (2 mm) and glutamate (10 mm). With no ADP present, this respiration primarily represents compensation for mitochondrial proton leak. State 3 complex I (CIP) OXPHOS capacity was assessed after addition of ADP (1 mm). State 3 lipid OXPHOS capacity (CIP + ETFP) was measured after addition of octanoyl carnitine (1.5 mm), which stimulates complex I and through β‐oxidation feed of electrons to complex II via the electron‐transferring flavoprotein. After addition of succinate (10 mm), cytochrome C (CytC) (10 μm) was then used to test outer mitochondrial membrane integrity. Finally, electron transfer system capacity (CI + IIE) in the presence of the substrates used in protocol I was tested with stepwise (0.4 μm) addition of carbonylcyanide p‐trifluoromethoxyphenylhydrazone (FCCP).
In Protocol II state 3 succinate dehydrogenase (SDH)/complex II (SDH/CIIP) OXPHOS capacity was assessed by addition of malate, glutamate, ADP, rotenone (0.5 μm) and succinate. In addition, state 3 complex IV (CIVP) OXPHOS capacity was analysed after addition of CytC, antimycin A (5 μm), ascorbate (2 mm) and N,N,N′,N′‐tetramethyl‐p‐phenylenediamine (TMPD) (0.5 mm).
A CytC response ≤ 5 pmol s−1 mg−1 was used as an indicator for preserved outer mitochondrial membrane structure. Respiratory assessments were carried out in duplicate and after hyperoxygenation (400 nmol ml−1) to avoid oxygen limiting the respiration. Chamber temperature during the experiments was 37°C. The substrate concentrations used in protocol I and II were chosen from previous unpublished pilot studies.
CS activity was used as a marker for mitochondrial content. It was assessed as described before (Larsen et al. 2015) with the exception that 20 mg of liver tissue initially was homogenized. CS activity is expressed as micromole substrate per minute per gram dry weight of liver tissue.
Paraffin embedding and histological staining
The liver tissue was fixed by immersion in 2% depolymerized paraformaldehyde supplemented with 0.15% picric acid for 24 h, transferred to 70% ethanol until embedded in paraffin on a Shandon Excelsior EStissue processor (Thermo Scientific, MA, USA). Paraffin embedded liver samples were cut in 4 μm‐thick sections and mounted on a microscope glass slide. For glycogen visualization, sections were stained with periodic acid–Schiff (PAS). Sections were de‐paraffinated in xylene and rehydrated by descending ethanol from 99% to 55% to water. After 5 min of incubation in periodic acid and 5 min wash in running tap water, sections were incubated in Schiff's reagent for 40 min. Then, sections were immersed in haematoxylin for 5 min and washed in running tap water for 10 min. Finally, sections were dehydrated in ascending ethanol to 99% and mounted in Pertex mounting medium (Histolab, Gothenburg, Sweden). Histopathological classification of non‐alchoholic steatohepatitis (NASH) according to Brunt's classification was performed blinded by an expert liver pathologist (Brunt & Tiniakos, 2010).
Image acquisition and processing
Image acquisition was performed with a Zeiss Axio Scan.Z1 slide scanner through a 20×/0.8 plan apochromat objective. Images were processed semi‐automatically with Zen Blue image processing software (Carl Zeiss, Germany). During tissue processing, lipids were washed out; hence, lipid droplets (LDs) appeared as perfectly rounded non‐stained (white) areas dispersed in the tissue. In the analysis program, white areas were designated as LDs based on threshold (R: 170–255; G: 115–255; B: 181–255), shape (circularity: 0.85–1) and size (area: 10–2000 μm2). The areas of the tissue that were stained by PAS (glycogen staining) were segmented by thresholds (R: 41–94; G: 0–88; B: 91–203), as was the total tissue area (R: 0–195; G: 0–120; B: 112–234). Results are expressed as average LD size (μm2), LD density (counts per total tissue area), percentage of total tissue area filled by LDs and percentage of total tissue area filled with glycogen. Representative images of the PAS staining and lipid droplet and glycogen segmentation are shown in Fig. 1 A–C.
Figure 1. Hepatic PAS staining and lipid droplet and glycogen segmentation .

A, representative image of hepatic PAS staining with the image analysis module in ZEN Blue software (Carl Zeiss, Germany). B and C, segmented lipid droplets are shown in red (B) and segmented glycogen stain is shown in orange (C). Software settings for lipid droplet and glycogen quantification are described in detail in Methods.
Biochemistry
Blood samples for ALAT and ASAT assessment were collected in BD (Becton, Dickinson and Company, NJ, USA) aprotinin‐containing vacutainers. Samples for biliubin were transferred to EDTA‐containing tubes. All samples were immediately cooled to 4°C and centrifuged at 2000 g for 10 min whereafter plasma was collected and stored at −80°C until time of analysis. All samples were analysed in duplicates on a Cobas 6000, C 501 analyser (Roche Diagnostics, Mannheim, Germany). Fasting glucose was analysed on an ABL 800 flex blood gas analyser (Radiometer Medical ApS, Brønshøj, Denmark). HbA1c was analysed on a DCA Vantage Analyser (Siemens Healthcare Diagnostics ApS, Ballerup, Denmark).
Statistical analysis
Patient characteristics, LD and glycogen percentage area were analysed by Student's unpaired t test and one‐way analysis of variance for comparison of OB and T2DM or all three groups, respectively. Kruskal–Wallis one‐way analysis of variance on ranks followed by Dunn's post hoc test was used to compare LD size and LD area. Respiratory protocols were analysed by two‐way analysis of variance for repeated measures followed by a Student–Newman–Keul's post hoc test. Data were log‐transformed before statistical analyses when appropriate. A P value < 0.05 was considered significant. Statistical analysis was performed in Sigmaplot 12.3 (Systat Software, San Jose, CA, USA). Data are presented as the mean ± SEM.
Results
There was no difference in age, weight, BMI and liver‐specific biochemical parameters between OB and T2DM (Table 1). Fasting glucose and HbA1c were lower and fasting insulin was similar in OB compared with T2DM. Body weight and BMI were lower in CON compared with OB and T2DM, while age‐ and liver‐specific biochemical parameters were similar (Table 1).
Table 1.
Patient characteristics in obese patients with (T2DM) and without (OB) type 2 diabetes and controls (CON)
| Patient characteristics | OB | T2DM | CON |
|---|---|---|---|
| n | 9 | 8 | 6 |
| Sex (M/F) | 2/7 | 4/4 | 1/5 |
| Age (years) | 36 ± 3 | 39 ± 4 | 45 ± 7 |
| Weight (kg) | 128 ± 9 | 119 ± 8 | 80 ± 10* |
| BMI (kg m−2) | 42 ± 1 | 39 ± 1 | 26 ± 2* |
| Fasting glucose (mmol l−1) | 5.4 ± 0.1 | 7.3 ± 0.5† | — |
| HbA1c (%) | 5.2 ± 0.1 | 6.4 ± 0.3† | — |
| HbA1c (mmol mol−1) | 33 ± 1 | 46 ± 3† | — |
| Fasting insulin (pmol l−1) | 88 ± 14 | 91 ± 16 | — |
| ALAT (U/L) | 24 ± 2 | 33 ± 10 | 34 ± 10 |
| ASAT (U/L) | 21 ± 1 | 25 ± 6 | 24 ± 3 |
| Bilirubin (μmol l−1) | 5 ± 1 | 6 ± 1 | 8 ± 1 |
| Duration of diabetes (years) | — | 5 ± 3 | — |
| Brunt classification score | 3.0 ± 0.4 | 3.5 ± 0.4 | 2.3 ± 0.2 |
| Medication (no. of patients) | |||
| Metformin | 4 | ||
| GLP‐1 analogue | 2 | ||
| Insulin | 1 |
Data are the mean ± SEM. Results were compared by unpaired t test or one‐way ANOVA when data from two or three groups were present, respectively. *Significantly different from OB and T2DM. †Significantly different from OB. Abbreviations: ALAT, alanine transaminase; ASAT, aspartate transaminase.
Results of the respiratory analyses are shown in Fig. 2. There was no difference in state 3 complex I (Fig. 2 A), II (Fig. 2 B) and IV (Fig. 2 D) OXPHOS capacity, state 3 lipid OXPHOS capacity (Fig. 2 C), leak (Fig. 2 E) or ETS capacity (Fig. 2 F) among OB, T2DM and CON. A single sample in the CON group (from protocol I) was excluded from the statistical analysis due to a high CytC response. CS activity did not differ in OB, T2DM and CON (12.4 ± 0.2, 13.6 ± 0.4 and 12.2 ± 0.8 μmol g−1 min−1, respectively). As a result, OXPHOS capacities corrected for the CS activity did not differ among groups (data not shown).
Figure 2. Hepatic mitochondrial OXPHOS capacity in obese patients with (T2DM) or without (OB) type 2 diabetes and controls (CON) .
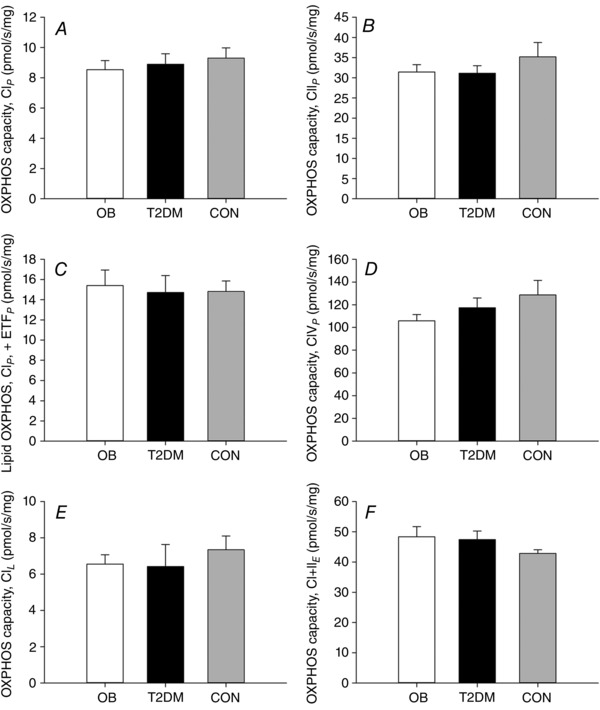
A, complex I linked state 3 OXPHOS capacity (CIP); B, complex II linked state 3 OXPHOS capacity (CIIP); C, state 3 lipid OXPHOS capacity (ETFP); D, complex IV linked state 3 OXPHOS capacity (CIVP); E, state 2 leak respiration (CIL); F, electron transfer system capacity when supported by CI + II. Open bars, OB; black bars, T2DM; grey bars, CON. Data are the mean ± SEM.
LD content and size and glycogen content results are shown in Fig. 3. LD size did not differ between OB and T2DM, but was lower in CON compared with T2DM (P < 0.05). LD area tended to be lower in CON compared with OB and T2DM (P = 0.10). The amount of LDs related to the total tissue area (LD density) and the glycogen area did not differ among the three groups. Brunt classification did not differ among OB, T2DM and CON (Table 1). No patients had NASH according to the Brunt classification. Representative images of PAS‐stained liver biopsies from the different groups are shown in Fig. 4.
Figure 3. Individual quantitative data on lipid droplets (A–C) and glycogen content (D) in liver biopsies from obese patients with (T2DM) or without (OB) type 2 diabetes and controls (CON) .
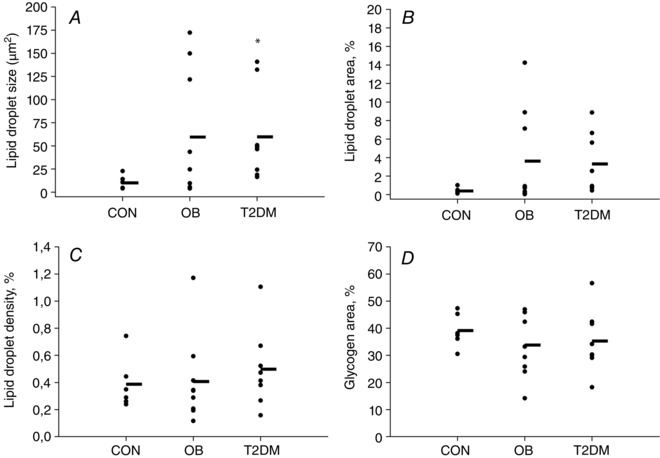
LD and glycogen area are defined as the percentage of total tissue area filled by LDs or glycogen, respectively. The horizontal line represents the average value. Results were compared by the Kruskal–Wallis test (lipid droplet size and area) and one‐way ANOVA (lipid droplet density and glycogen area). *Significantly different from CON. Data are the mean ± SEM.
Figure 4. Variation in hepatic lipid content .
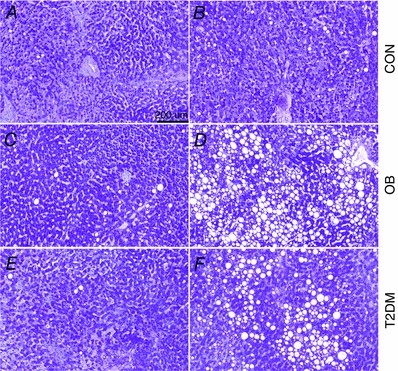
Representative images of hepatic PAS staining showing subjects with low (A, C and E) and high (B, D and F) hepatic lipid content in CON (A and B), OB (C and D) and T2DM (E and F).
Discussion
The primary finding in the present study is the similar hepatic mitochondrial OXPHOS capacity in obese patients with and without type 2 diabetes and controls (Fig. 2). Furthermore, the CS activity, a measure of mitochondrial content, is not different among the three groups. Thus, we are unable to detect any impairment in hepatic mitochondrial respiration in the obese hyperglycaemic state.
Hepatic lipid content varied between individuals within OB and T2DM (Figs 3 and 4) and a 6‐fold difference in LD size and a 10‐fold difference in LD area (percentage of total tissue area filled by LDs) between the two obese groups and CON were seen (Figs 3 A and B).
This is the first study to report human hepatic mitochondrial OXPHOS capacity in patients with type 2 diabetes using HRR. At first glance, it would seem our results are in contrast to previous 31P MRS results comparing ATP production in patients with type 2 diabetes and matched controls (Szendroedi et al. 2009; Schmid et al. 2011). However, there are important methodological differences using 31P MRS and HRR. First, the 31P MRS methodology is limited by the inability to distinguish between ATP produced by cytosolic creatine kinases and the mitochondria (From & Ugurbil, 2011). These cytosolic enzymes are washed out during the preparation of tissue for respiratory analysis. Second, 31P MRS provides a measure of the present hepatic ATP production and content, and the HRR provides the maximal hepatic mitochondrial OXPHOS capacity with saturating substrate concentrations. Thus, the present results indicate that the maximal capacity of the individual complexes of the ETS are not impaired in patients with obesity with and without type 2 diabetes. To confirm our findings, more detailed HRR investigations such as titration analysis stimulating the individual complexes are required to further elucidate hepatic mitochondrial respiratory function in patients with obesity with and without type 2 diabetes.
Hepatic lipid and glycogen content in the obese groups varied (Figs 3 and 4). Whereas glycogen area (percentage of total tissue area filled by glycogen; Fig. 3 D) did not differ among groups, LD size and area (percentage of total tissue area filled by LDs) tended to be higher in OB and T2DM compared with CON (Fig. 3 A and B). LD density, defined as the number of LDs relative to the area of analysed tissue, did not differ among groups (Fig. 3 C). Thus, in the present study the increased amount of lipids was stored in existing hepatocyte LDs leading to macrovesicular steatosis. This way of storing lipids is characteristic for patients with ectopic lipid accumulation and with non‐alcoholic fatty liver disease (NAFLD) (Brunt, 2007).
A recently published study by Koliaki et al. found higher hepatic OXPHOS capacity, corrected for CS activity, in individuals with obesity with or without NAFLD compared with lean subjects (Koliaki et al. 2015). In the same publication, the authors showed a decreased hepatic OXPHOS capacity in patients with NASH (Koliaki et al. 2015). This last notion is supported by Perez‐Carreras et al. who found markedly decreased activity in all five complexes of the ETS by spectrophotometry in patients with NASH compared with controls (Perez‐Carreras et al. 2003). In the present study no patients were classified as having NASH. In addition, there was no difference in NAFLD score (Brunt classification; Table 1) between the groups. Nevertheless, we were not able to confirm the findings by Koliaki et al., that subjects with obesity but without NAFLD have increased OXPHOS capacity per mitochondria compared with non‐obese subjects. The OXPHOS capacity per mitochondria is calculated by dividing the OXPHOS capacity with the CS activity. Therefore, the most likely explanation for the discrepancy between the two studies is the similar CS activity among OB, CON and T2DM in the present study compared with the (non‐significant) ∼25% lower CS activity in subjects with obesity compared with lean subjects reported by Koliaki et al. This difference between the two studies is puzzling and not easy to explain, but the relationship between CS activity and mitochondrial content has not been investigated in the liver as it has in skeletal muscle (Larsen et al. 2012). Therefore, it is not known whether the CS activity is a suitable marker for mitochondrial content in the liver. This is a limitation to this study as well as the study by Koliaki et al. In the present study there are several other limitations: most importantly, a relatively low number of participants were included in our study, and therefore cautious interpretation of our results is warranted and the results need to be confirmed in a larger cohort of patients. A wedge biopsy was used to obtain the liver tissue and whether this tissue is representative of the general hepatic lipid content cannot be confirmed in the present study. For methodological reasons, we used octanoyl carnitine, a medium‐chained fatty acid, and not the more physiologically relevant long‐chain palmitoyl carnitine fatty acid in our mitochondrial testing. This allowed us to evaluate lipid induced OXPHOS independent of the carnitine palmitoyltransferase II. Finally, since assessment of hepatic insulin sensitivity was not performed in the present study, no causal link between mitochondrial function and hepatic insulin sensitivity can be made based on the present findings.
In summary, the present study demonstrates for the first time that hepatic OXPHOS capacity is not impaired in patients with type 2 diabetes. The amount of mitochondria, assessed by the CS activity, is also not different the three groups with markedly different BMI, hepatic lipid content and glycaemic control. Furthermore, hepatic OXPHOS capacity in complex I, II and IV together with state 3 lipid OXPHOS capacity are unrelated to BMI and hepatic lipid content. While our results do not rule out differences in hepatic mitochondrial ATP production in situ, they indicate that the maximal capacity of the mitochondrial ETS is intact in patients with obesity and type 2 diabetes.
Additional information
Competing interests
The authors declare that they have no conflict of interest associated with this article.
Author contributions
M.L. performed the respiratory measurements and wrote manuscript. M.H. collected and analysed anthropometric and biochemical data and contributed to writing the manuscript. M.D.K., C.P., B.V. and S.S.P. processed and analysed liver histology data and revised the manuscript. M.S., A.K.F. and L.T. recruited subjects, performed the surgeries, acquired the liver biopsies, researched data and revised the manuscript. J.W.H. and F.D. designed the study, interpreted data and critically revised the manuscript. F.D. is guarantor of the manuscript. All authors approved the final version of the manuscript, all persons designated as authors qualify for authorship, and all those who qualify for authorship are listed in the manuscript. The experimental procedures in this manuscript were performed at Xlab, Centre for Healthy Aging, Department of Biomedical Sciences, University of Copenhagen. The surgeries were performed at Køge Hospital, Department of Surgery, Koege, Denmark.
Funding
Financial support from the Innovation Fund Denmark (0603‐00381B) and the Nordea Foundation is gratefully acknowledged.
Acknowledgements
We thank laboratory technicians Jeppe Bach, Regitze Kraunsøe and Katrine Qvist for technical assistance during the study.
Linked articles This article is highlighted by a Perspective by Perry & Wright. To read this Perspective, visit http://dx.doi.org/10.1113/JP272573.
References
- Amaro A, Fabbrini E, Kars M, Yue P, Schechtman K, Schonfeld G & Klein S (2010). Dissociation between intrahepatic triglyceride content and insulin resistance in familial hypobetalipoproteinemia. Gastroenterology 139, 149–153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonadonna RC, Groop L, Kraemer N, Ferrannini E, Del PS & DeFronzo RA (1990). Obesity and insulin resistance in humans: a dose‐response study. Metabolism 39, 452–459. [DOI] [PubMed] [Google Scholar]
- Boushel R, Gnaiger E, Schjerling P, Skovbro M, Kraunsoe R & Dela F (2007). Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia 50, 790–796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunt EM (2007). Pathology of fatty liver disease. Mod Pathol 20 Suppl 1, S40–S48. [DOI] [PubMed] [Google Scholar]
- Brunt EM & Tiniakos DG (2010). Histopathology of nonalcoholic fatty liver disease. World J Gastroenterol 16, 5286–5296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- From AH & Ugurbil K (2011). Standard magnetic resonance‐based measurements of the Pi→ATP rate do not index the rate of oxidative phosphorylation in cardiac and skeletal muscles. Am J Physiol Cell Physiol 301, C1–C11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen M, Lund MT, Jorgensen AL, Palsoe MK, van HG, Helge JW & Dela F (2016). The effects of diet‐ and RYGB‐induced weight loss on insulin sensitivity in obese patients with and without type 2 diabetes. Acta Diabetol 53, 423–432. [DOI] [PubMed] [Google Scholar]
- Koliaki C, Szendroedi J, Kaul K, Jelenik T, Nowotny P, Jankowiak F, Herder C, Carstensen M, Krausch M, Knoefel WT, Schlensak M & Roden M (2015). Adaptation of hepatic mitochondrial function in humans with non‐alcoholic fatty liver is lost in steatohepatitis. Cell Metab 21, 739–746. [DOI] [PubMed] [Google Scholar]
- Kotronen A, Juurinen L, Tiikkainen M, Vehkavaara S & Yki‐Jarvinen H (2008). Increased liver fat, impaired insulin clearance, and hepatic and adipose tissue insulin resistance in type 2 diabetes. Gastroenterology 135, 122–130. [DOI] [PubMed] [Google Scholar]
- Larsen S, Danielsen JH, Sondergard SD, Sogaard D, Vigelsoe A, Dybboe R, Skaaby S, Dela F & Helge JW (2015). The effect of high‐intensity training on mitochondrial fat oxidation in skeletal muscle and subcutaneous adipose tissue. Scand J Med Sci Sports 25, e59–e69. [DOI] [PubMed] [Google Scholar]
- Larsen S, Nielsen J, Hansen CN, Nielsen LB, Wibrand F, Stride N, Schroder HD, Boushel R, Helge JW, Dela F & Hey‐Mogensen M (2012). Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. J Physiol 590, 3349–3360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionetti L, Mollica MP, Lombardi A, Cavaliere G, Gifuni G & Barletta A (2009). From chronic overnutrition to insulin resistance: the role of fat‐storing capacity and inflammation. Nutr Metab Cardiovasc Dis 19, 146–152. [DOI] [PubMed] [Google Scholar]
- Lowell BB & Shulman GI (2005). Mitochondrial dysfunction and type 2 diabetes. Science 307, 384–387. [DOI] [PubMed] [Google Scholar]
- Lund MT, Hansen M, Skaaby S, Dalby S, Stockel M, Floyd AK, Bech K, Helge JW, Holst JJ & Dela F (2015). Preoperative beta‐cell function in patients with type 2 diabetes is important for the outcome of Roux‐en‐Y gastric bypass surgery. J Physiol 593, 3123–3133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez‐Carreras M, Del HP, Martin MA, Rubio JC, Martin A, Castellano G, Colina F, Arenas J & Solis‐Herruzo JA (2003). Defective hepatic mitochondrial respiratory chain in patients with nonalcoholic steatohepatitis. Hepatology 38, 999–1007. [DOI] [PubMed] [Google Scholar]
- Pesta D & Gnaiger E (2012). High‐resolution respirometry: OXPHOS protocols for human cells and permeabilized fibres from small biopsies of human muscle. Methods Mol Biol 810, 25–58. [DOI] [PubMed] [Google Scholar]
- Schmid AI, Szendroedi J, Chmelik M, Krssak M, Moser E & Roden M (2011). Liver ATP synthesis is lower and relates to insulin sensitivity in patients with type 2 diabetes. Diabetes Care 34, 448–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seppala‐Lindroos A, Vehkavaara S, Hakkinen AM, Goto T, Westerbacka J, Sovijarvi A, Halavaara J & Yki‐Jarvinen H (2002). Fat accumulation in the liver is associated with defects in insulin suppression of glucose production and serum free fatty acids independent of obesity in normal men. J Clin Endocrinol Metab 87, 3023–3028. [DOI] [PubMed] [Google Scholar]
- Szendroedi J, Chmelik M, Schmid AI, Nowotny P, Brehm A, Krssak M, Moser E & Roden M (2009). Abnormal hepatic energy homeostasis in type 2 diabetes. Hepatology 50, 1079–1086. [DOI] [PubMed] [Google Scholar]
- Visser ME, Lammers NM, Nederveen AJ, van der Graaf M, Heerschap A, Ackermans MT, Sauerwein HP, Stroes ES & Serlie MJ (2011). Hepatic steatosis does not cause insulin resistance in people with familial hypobetalipoproteinaemia. Diabetologia 54, 2113–2121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmelmann CL, Dela F & Mortensen EL (2014). Psychological predictors of mental health and health‐related quality of life after bariatric surgery: a review of the recent research. Obes Res Clin Pract 8, e314–e324. [DOI] [PubMed] [Google Scholar]
- Wimmelmann CL, Smith E, Lund MT, Hansen M, Dela F & Mortensen EL (2015). The psychological profile of bariatric patients with and without type 2 diabetes: baseline results of the longitudinal GASMITO‐PSYC study. Surg Obes Relat Dis 11, 412–418. [DOI] [PubMed] [Google Scholar]