

Abstract
BACKGROUND
A decline in fertility is evident in human females past their middle thirties. This ‘reproductive senescence’, marked by a sharp decline in pregnancy rates, may be attributed to reductions in numbers of available oocytes and their quality. Because Old World primates exhibit ovarian morphology and physiological control and timing of menstrual cycles closely resembling those of humans, the current study investigated the rhesus macaque as a potential model for human reproductive senescence.
METHODS
Ovaries collected from females aged 1–25 years and divided into five age groups were analysed histologically.
RESULTS
General ovarian morphology demonstrated significant changes as the females approached menopause. The proportions of primordial and primary follicles all demonstrated significant differences across age groups (primordial: 77.1, 79.9, 69.7, 62.9, 55.1%; primary: 21.5, 18.8, 28.5, 35.2, 43.1% for age groups 1 to 5 respectively; P < 0.0001 for both). Samples from females approaching or undergoing the menopausal transition (aged 20–25 years) demonstrated evidence of ovarian senescence, having scattered and atretic follicles, low numbers of primordial follicles and reduced stromal tissue.
CONCLUSION
This study supports the value of the rhesus monkey as a model for reproductive ageing because its ovary undergoes follicular reservoir depletion similiar to that seen in humans.
Keywords: follicle, oocytes, ovary, primates, reproductive ageing
Introduction
The primordial follicle population within an ovary represents the reservoir of potential viable oocytes available to the reproducing female. This population suffers considerable decreases over the postnatal lifespan partly due to follicular recruitment processes, but mostly due to follicular atresia within the ovary. As the female ages, the ovary undergoes senescence culminating in the menopause when the ovary has exhausted the reserve of viable follicles and oocytes (Ginsberg, 1991; Faddy and Gosden, 1996). At this time, ovulatory activity ceases and estrogen levels have decreased. Women approaching or undergoing the menopausal transition or climacteric may experience difficulty in conceiving and often resort to assisted reproductive techniques to achieve fertilization. In many of these cases, the woman will undergo a stimulation regimen to maximize follicle recruitment for IVF procedures. However, by this point, the ovaries may have lost the capacity to respond to such treatment due to diminished ovarian reserve and oocyte quality (Volarcik et al., 1998; Hansen et al., 2003).
The development of a non-human primate model is of particular interest for human reproductive ageing studies (Lane, 2000). Old World non-human primates, in particular macaque species, are likely to be the most appropriate model for reproductive ageing studies because their menstrual cycling, hormonal secretion patterns, and morphological characteristics of the reproductive organs are similar to those of the human female during the reproductive cycle. The rhesus macaque (Macaca mulatta) is perhaps the ideal model for studies of this nature because it is a highly studied and well-defined species. Female rhesus macaques are reproductively active over a span of ~20 years (from 5 to 25 years of age) and undergo the pathological and hormonal changes characteristic of the human female climacteric as she approaches the end of her reproductive lifespan (Walker, 1995; Gilardi et al., 1997; Shideler et al., 2001). These changes include a decrease in estrogen levels, the cessation of ovulatory events and an increase in circulating FSH levels (Walker, 1995).
Early observations on total oocyte numbers within the rhesus ovary by Green and Zuckerman (1951) did not find a significant decrease in total oocyte numbers with age. However, these studies utilized a small number of individuals (n = 12) from the beginning of the reproductive spectrum (aged 3–9 years) with age being estimated by dentition and body weight. Presumably, at these earlier ages, major decreases in oocyte numbers would not be evident. The same group added additional observations on total oocyte numbers in the rhesus ovary (Green and Zuckerman, 1954) and, based solely on the assumption that heavier individuals were older, determined that oocyte numbers did decrease with age. Recent studies in pigtailed macaques (Macaca nemestrina) demonstrate some of the pathological characteristics of approaching menopause (a significant decrease in primordial follicle numbers), although older females (aged 15–25 years) were not examined (Miller et al., 1997, 1999). It must be noted that ageing studies utilizing older, peri- to post-menopausal monkeys are sparse and that certain details of human menopause, e.g. the subtle and gradual onset of menopausal-related changes occurring before irregular cycling begins (Santoro, 2002), are not shared by macaque females. Macaques instead appear to undergo abrupt changes at the onset of irregular cycles (Shideler et al., 2001).
In view of the limited number of studies on older non-human primates and the existence of variant menopausal details, the current project examined ovarian senescence within the rhesus macaque across age groups from pre-pubertal to presumptively post-menopausal females (ages ~ 1–25 years), thereby covering the entire reproductive lifespan of this species. Results from histological examination provide evidence of follicular depletion over time, with a marked reduction at later ages.
Materials and methods
Female rhesus macaques of various ages (0.93–25.44 years) were systematically euthanized for medical reasons over a 5 year period at the Tulane National Primate Research Center (TNPRC, USA). Ovaries were collected and archived for later examination. These samples were divided into several age groups (<5 years, n = 12; 5–10 years, n = 14; 10–15 years, n = 14; 15–20 years, n = 11; >20 years, n = 13) in order to determine the possibility of age-related trends in ovarian morphology. Serum samples from each euthanized female were also obtained at the time of necropsy and archived for subsequent analysis of hormone profiles.
Histology
Ovaries were sectioned into two equal-sized halves, and preserved in 10% formalin. Fixation was followed by a water rinse and subsequent storage in 70% ethanol. The tissues were subsequently embedded in paraffin wax (Fisher Scientific, USA) following immersion in a graded series of alcohols (70–100%) and limonene-based clearing agent Xylene (Fisher Scientific). Embedded tissue was sectioned (5 μm) using a rotary microtome. The sections were adhered to microscope slides coated with gelatin (Staon; Surgipath, USA) and dried at 56°C for 24 h.
Slides from each specimen’s ovary sample were stained using Harris’s haematoxylin and eosin (HHE) to visualize general cellular structure throughout the tissue (Wheater et al., 1987). Briefly, slides were passed through a series of the clearing agent Xylene and rehydrated in a graded series of ethanols (100, 100, 100, 95, 80%). Following a distilled water bath, the slides were incubated in Harris’s haematoxylin as the primary staining agent. The samples were then rinsed through a running tap water bath and Clarifier 1 (Richard–Allan Scientific, USA) to remove excess haematoxylin. Following another water bath, a strong basic solution of 0.3% ammonia water was used to shift the colour of the haematoxylin in the sections to blue. This step was followed by another water bath, incubation in the eosin counterstain and subsequent dehydration in a graded series of ethanol (80, 95, 95, 95, 100, 100, 100%), and immersion in Xylene, then the slides were coverslipped with Cytoseal-60 (Stephens Scientific, USA).
Ovarian follicles were examined using a Nikon Optiphot microscope (Nikon Instruments Inc., USA). Follicles containing an oocyte whose nucleolus could be clearly defined were scored for follicle classification (primordial, primary and antral) and count. One ovary per individual was used based on previous findings by Miller et al. (1999) that demonstrated symmetry of follicle numbers between ovaries in macaques. Because the aim of the study was to demonstrate ovarian senescence occurring over time, a representative single section was taken through the middle of each ovary and follicles were counted over the entire area. Although this approach precludes consideration of any possible heterogeneous distribution within the ovary, it was impractical to section the entire ovary of each animal. But as the size of each age group was sufficiently large and the sampling method was kept consistent throughout the study, it was thought that this method would provide an accurate reflection of any morphological changes. Characterization of follicles as primordial, primary or antral was based on the criteria previously described by Bloom and Fawcett (1970). Primordial follicles contain oocytes with a large nucleus and pale dispersed chromatin (Figure 1A). These oocytes are surrounded by a single layer of flattened granulosa or follicular cells. Primary follicles (Figure 1B) possess either single or multiple layers of granulosa cells that are cuboidal or low columnar. The zona pellucida forms during the primary follicle stage and can be seen as a deeply staining layer between the oocyte and surrounding granulosa cells. Antral follicles (Figure 1C) contain a large fluid-filled space or antrum. At the antral stage, follicular cells continue to proliferate; however, the oocyte itself has reached full size.
Figure 1.
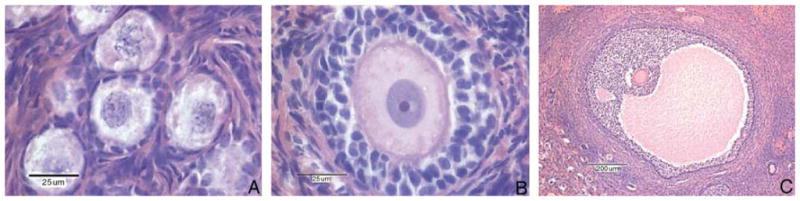
(A–C) Various stages of ovarian follicle development. Primordial follicles (A) are characterized by a flattened granulosa cell border. The primary follicle (B) exhibits several layers of granulosa cells and a zona pellucida surrounding the oocyte. The antral follicle (C) contains a well-developed antrum.
Photographs of ovarian sections were obtained using a Photometrics Coolsnap cf digital camera (Roper Scientific, USA) under Metamorph Imaging System software (Universal Imaging Corp., USA) on a Nikon Eclipse E800 microscope.
Radioimmunoassay
Radioimmunoassays of sex hormones were conducted using Coat-A-Count kits purchased from Diagnostic Products Corporation (USA). These are no-extraction, solid phase radioimmunoassays designed for measurement of the specific hormone in serum. Intra-assay and inter-assay precision averaged 5.3 and 5.65% respectively.
Serum FSH concentrations were determined by radioimmunoassay in all samples at the completion of the study using protocols and reagents specifically developed for macaques obtained from the National Institute of Health (NIH) National Hormone and Peptide Program directed by A.F. Parlow (Harbor-UCLA Medical Center, USA). The intra-assay coefficient of variation for FSH was 6.1%.
Statistics
Data were analysed by least squares analysis of variance using the JMP software (SAS Institute, 2000). Individual means were compared by orthogonal contrast (SAS Institute Statistics and Graphics Guide, 2000). For the determination of number of primordial, primary and antral follicles, as well as for the results of the hormone assays, the age of female and number of parturitions of each female were entered into the model as effects.
Results
Follicles within ovary sections were examined, assigned to one of the three categories (Figure 1A–C) and counted. Most sections revealed obvious and distinct changes in follicle populations with increasing age (Figure 2). Analysis of the proportion of the three follicle classes revealed that while the percentage of antral follicles remained nearly unchanged, the percentage of primary and primordial follicles changed significantly with age (Figure 3, P < 0.05). Analysis of the total number of primordial, primary and antral follicles within each section likewise showed a significant decrease with increasing age of the female (Figure 4A–C, P < 0.05).
Figure 2.

Ovarian sections illustrating changing follicle populations with increasing age.
Figure 3.
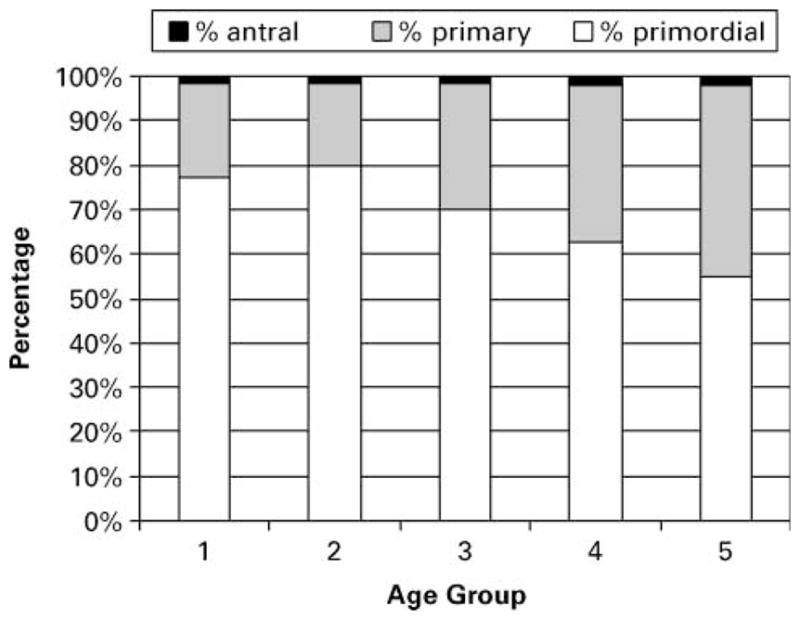
Effects of age on the proportion of primordial, primary and antral follicles.
Figure 4.
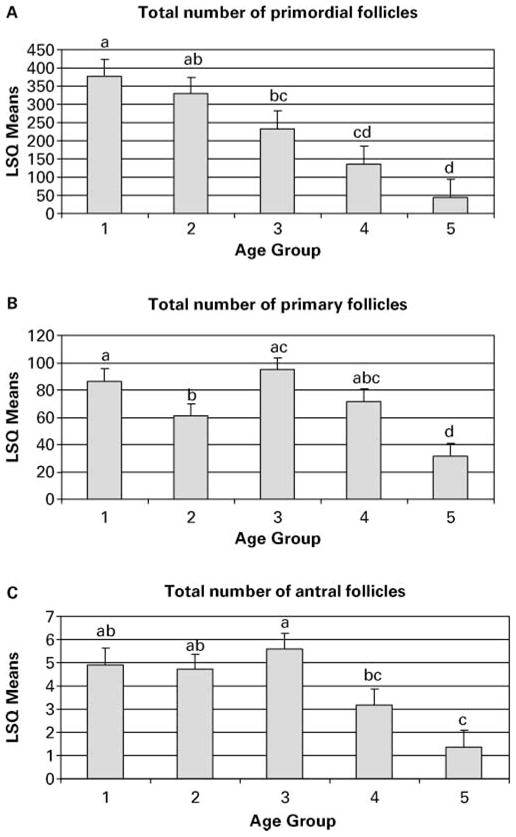
(A–C) Follicle counts across age groups. Age group 1, <5 years; 2, 5–10 years; 3, 10–15 years; 4, 15–20 years; 5, >20 years. Different letters above error bars indicate significant differences in follicle number among age groups (P < 0.05).
Females undergoing the peri-menopausal and menopausal transition (age group 20–25 years) provided the most obvious visual evidence of ovarian reserve attrition, having scattered and atretic follicles, occasional primordial follicles and reduced amounts of stromal tissue. Statistical analysis also demonstrated that the number of parturitions of each female proved to have a significant effect on the proportion of primordial and primary follicles (Figure 5, P < 0.01) that was independent of age, with a decrease in the percentage of primordial follicles and a concomitant increase in mean percentage of primary follicles.
Figure 5.
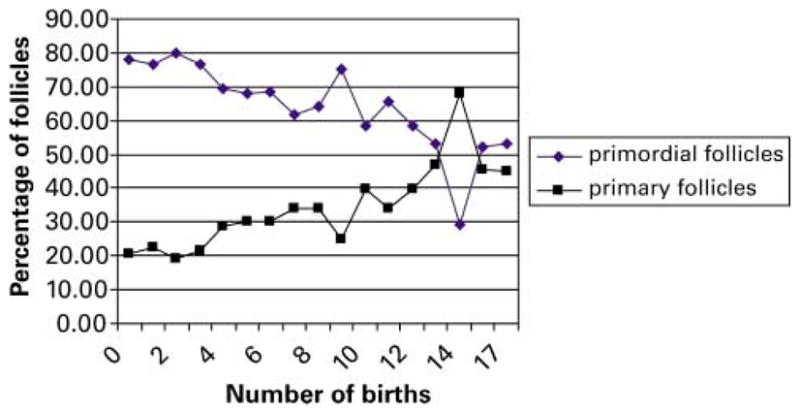
Effect of the number of parturitions on the proportion of primordial and primary follicles.
Analyses of blood samples taken at the time of euthanasia for concentration of estrogen, progesterone, and FSH did not show any difference between females of various ages.
Discussion
This study describes ovarian follicle morphology and depletion in rhesus macaques from pre-pubescence until menopausal stages. It expands the observations of Gilardi et al. (1997) in describing ovarian senescence in ageing laboratory rhesus macaques by including individuals from the entire reproductive lifespan. Ovarian senescence in aged macaques supports the value of the rhesus monkey as a model for reproductive ageing in humans because similar follicular depletion patterns occur (Richardson et al., 1987; Faddy et al., 1992). Samples from female macaques approaching or undergoing the menopausal transition (age group 20–25 years) provided evidence of ovarian attrition, having scattered and atretic follicles, occasional primordial follicles and reduced amounts of stromal tissue.
Ovaries of young female rhesus monkeys contained large populations of primordial follicles, which decreased significantly with age. Decreases in primordial follicle populations can be, in part, attributed to follicle recruitment during the follicular phase of the menstrual cycle. The factors determining which follicles are recruited to grow and develop during a reproductive cycle remain undefined, but growth factors and gonadotrophins are believed to be necessary to activate initial follicular growth phases. The majority of the follicle population decrease, however, is due to natural attrition. Follicular atresia or degeneration is a prominent feature in the life of the mammalian ovary, beginning in utero, occurring primarily at birth and before puberty (Baker, 1963; Forabosco et al., 1991; Faddy, 2000) and continuing at a slower rate throughout reproductive life. It is intriguing that the recent study by Johnson et al. (2004) describing the persistence of germline stem cells in the mouse ovary appears to challenge the long-held view that females have finite gamete reserves. In the mouse, ‘ovoid’ bodies within the ovary seem to continue with oogenesis, thereby slowing down the rate of follicular attrition. This may have significant implication for human reproduction, although it remains to be determined whether this phenomenon also occurs in other species.
The present study showed that the rhesus macaque ovary exhibits an age-related pattern of follicle depletion similar to that described in the human ovary (Faddy, 2000). As the rhesus female approaches the third decade of life, morphological characteristics of the menopausal transition or climacteric become evident. These characteristics include loss of stromal tissue and extremely low populations of primordial, primary and antral follicles, many of which have become atretic.
Although total numbers of primary follicles within representative sections observed decreased with age, the percentage of the total follicle population represented by this stage increased over time. The decrease in number of primary follicles reflects the degenerative processes occurring within the ovary. The increase in population percentage of the primary follicles may be explained through the natural process of primordial follicle recruitment during the follicular phase, in which selected follicles are initiated into growth and begin to proliferate granulosa cell layers surrounding the oocyte. This process is under the control of growth hormones and results in a fraction of the primordial follicles being converted into primary follicles (Webb et al., 2003). Further growth is gonadotrophin dependent (Macklon and Fauser, 1999) and many of the newly formed primary follicles will cease development possibly due to lack of FSH receptors on the surrounding follicular cells, and so become arrested at the primary stage or become atretic. When number of births was considered, there was a significant increase in the percentage of primary follicles with an increase in parturitions. The cause of this intriguing observation is not clear, but presumably a female who gives birth to a larger number of offspring is pregnant for much of her reproductive lifespan. During pregnancy, the menstrual cycle ceases as a result of the continued production of progesterone from the corpus luteum and later from the placenta, which blocks the pituitary from producing FSH (Pohl et al., 1982). Consequently, primary follicle recruitment and growth is halted, while recruitment and loss of primordial follicles would continue throughout life.
During the menopausal transition of rhesus females, there is a marked variability in levels of reproductive hormones (Gilardi et al., 1997; Shideler et al., 2001). Reproductive hormone variability may be a result of follicular depletion as proposed for the human female (O’Connor et al., 2001). As the female ages, there is a shift and decrease in overall follicular hormonal signalling, partly due to decreases in total numbers of follicles and possibly also due to the quality of remaining oocytes. In this study, because there were no available comparative data for the female macaques nor information regarding stage of menstrual cycle at the time of their deaths, the single time-point measurements did not provide useful information and, not surprisingly, no age-related differences were observed in sex hormone or gonadotrophin levels. Further studies are currently underway to examine long-term changes in hormone profiles associated with the end of reproductive life through controlled sequential samples taken over prolonged periods of time before and after menopause.
In conclusion, the current study demonstrates striking similarities between the rhesus monkey and human species in changing follicular population distributions related to reproductive senescence. Future studies with the rhesus macaque model could be useful for understanding the mechanisms underlying reproductive ageing so that assisted reproductive technologies can be devised to address this phenomenon.
Acknowledgments
The authors gratefully acknowledge Carol Coyne for her assistance in the preparations of the tissue sections. This study was supported in part by NIH grant no. R24 RR15395 and by the Tulane National Primate Research Center base grant NIH 5P51 RR00164-41.
References
- Baker TG. A quantitative and cytological study of germ cells in human ovaries. Proc R Soc Lond B. 1963;158:417–433. doi: 10.1098/rspb.1963.0055. [DOI] [PubMed] [Google Scholar]
- Bloom W, Fawcett DW. A Textbook of Histology. 9. WB Saunders Company; Philadelphia, PA: 1970. pp. 728–742. [Google Scholar]
- Faddy MJ. Follicle dynamics during ovarian ageing. Mol Cell Endocrinol. 2000;163:43–48. doi: 10.1016/s0303-7207(99)00238-5. [DOI] [PubMed] [Google Scholar]
- Faddy MJ, Gosden RG. A model confirming the decline in follicle numbers to the age of menopause in women. Hum Reprod. 1996;11:1484–1486. doi: 10.1093/oxfordjournals.humrep.a019422. [DOI] [PubMed] [Google Scholar]
- Faddy MJ, Gosden RG, Gougeon A, Richardson SJ, Nelson JF. Accelerated disappearance of ovarian follicles in mid-life: implications for forecasting menopause. Hum Reprod. 1992;7:1342–1346. doi: 10.1093/oxfordjournals.humrep.a137570. [DOI] [PubMed] [Google Scholar]
- Forabosco A, Sforza C, DePol A, Vizzotto L, Marzona L, Ferrario VF. Morphometric study of the human neonatal ovary. Anat Rec. 1991;231:201–208. doi: 10.1002/ar.1092310208. [DOI] [PubMed] [Google Scholar]
- Gilardi KVK, Shideler SE, Valverde CR, Roberts JA, Lasley BL. Characterization of the onset of menopause in the rhesus macaque. Biol Reprod. 1997;57:335–340. doi: 10.1095/biolreprod57.2.335. [DOI] [PubMed] [Google Scholar]
- Ginsberg J. What determines the age at menopause? Br Med J. 1991;302:1288–1289. doi: 10.1136/bmj.302.6788.1288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green SH, Zuckerman S. The number of oocytes in the mature rhesus monkey (Macaca mulatto) J Endocrinol. 1951;7:194–202. doi: 10.1677/joe.0.0070194. [DOI] [PubMed] [Google Scholar]
- Green SH, Zuckerman S. Further observations on oocyte numbers in mature rhesus monkeys (Macaca mulatto) J Endocrinol. 1954;10:284–290. doi: 10.1677/joe.0.0100284. [DOI] [PubMed] [Google Scholar]
- Hansen KR, Morris JL, Thyer AC, Soules MR. Reproductive aging and variability in the ovarian antral follicle count: application in the clinical setting. Fertil Steril. 2003;80:577–583. doi: 10.1016/s0015-0282(03)00741-6. [DOI] [PubMed] [Google Scholar]
- Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature. 2004;428:145–150. doi: 10.1038/nature02316. [DOI] [PubMed] [Google Scholar]
- Lane MA. Nonhuman primate models in biogerontology. Exp Geron. 2000;35:533–541. doi: 10.1016/s0531-5565(00)00102-9. [DOI] [PubMed] [Google Scholar]
- Macklon NS, Fauser BC. Aspects of ovarian follicle development throughout life. Horm Res. 1999;52:161–170. doi: 10.1159/000023456. [DOI] [PubMed] [Google Scholar]
- Miller PB, Charleston JS, Battaglia DE, Klein NA, Soules MR. An accurate, simple method for unbiased determination of primordial follicle number in the primate ovary. Biol Reprod. 1997;56:909–915. doi: 10.1095/biolreprod56.4.909. [DOI] [PubMed] [Google Scholar]
- Miller PB, Charleston JS, Battaglia DE, Klein NA, Soules MR. Morphometric analysis of primordial follicle number in pigtailed monkey ovaries: symmetry and relationship with age. Biol Reprod. 1999;61:553–556. doi: 10.1095/biolreprod61.2.553. [DOI] [PubMed] [Google Scholar]
- O’Connor KA, Holman DJ, Wood JW. Menstrual cycle variability and the perimenopause. Am J Hum Biol. 2001;13:465–478. doi: 10.1002/ajhb.1078. [DOI] [PubMed] [Google Scholar]
- Pohl CR, Richardson DW, Marshall G, Knobil E. Mode of action of progesterone in the blockade of gonadotropin surges in the rhesus monkey. Endocrinology. 1982;110:1454–1455. doi: 10.1210/endo-110-4-1454. [DOI] [PubMed] [Google Scholar]
- Richardson SJ, Senikas V, Nelson JF. Follicular depletion during the menopausal transition: evidence for accelerated loss and ultimate exhaustion. J Clin Endocrinol Metab. 1987;65:1231. doi: 10.1210/jcem-65-6-1231. [DOI] [PubMed] [Google Scholar]
- SAS Institute. JMP Version 4. SAS Institute; Cary, NC: 2000. Statistics and Graphics Guide. [Google Scholar]
- Santoro N. The menopause transition: an update. Hum Reprod Update. 2002;8:155–160. doi: 10.1093/humupd/8.2.155. [DOI] [PubMed] [Google Scholar]
- Shideler SE, Gee NA, Chen J, Lasley BL. Estrogen and progesterone metabolites and follicle-stimulating hormone in the aged macaque female. Biol Reprod. 2001;65:1718–1725. doi: 10.1095/biolreprod65.6.1718. [DOI] [PubMed] [Google Scholar]
- Volarcik K, Sheean L, Goldfarb J, Woods L, Abdul-Karim FW, Hunt P. The meiotic competence of in-vitro matured human oocytes is influenced by donor age: evidence that folliculogenesis is compromised in the reproductively aged ovary. Hum Reprod. 1998;13:154–160. doi: 10.1093/humrep/13.1.154. [DOI] [PubMed] [Google Scholar]
- Walker ML. Menopause in female rhesus monkeys. Am J Primatol. 1995;35:59–71. doi: 10.1002/ajp.1350350106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb R, Nicholas B, Gong JG, Campbell BK, Gutierrez CG, Garverick HA, Armstrong DG. Mechanisms regulating follicular development and selection of the dominant follicle. Reprod Suppl. 2003;61:71–90. [PubMed] [Google Scholar]
- Wheater PR, Burkitt HG, Daniels VG. Functional Histology: A Text and Colour Atlas. Churchill Livingstone; New York: 1987. [Google Scholar]