

Abstract
The process of endospore germination in Clostridium difficile, and other Clostridia, increasingly is being found to differ from the model spore-forming bacterium, Bacillus subtilis. Germination is triggered by small molecule germinants and occurs without the need for macromolecular synthesis. Though differences exist between the mechanisms of spore germination in species of Bacillus and Clostridium, a common requirement is the hydrolysis of the peptidoglycan-like cortex which allows the spore core to swell and rehydrate. After rehydration, metabolism can begin and this, eventually, leads to outgrowth of a vegetative cell. The detection of hydrolyzed cortex fragments during spore germination can be difficult and the modifications to the previously described assays can be confusing or difficult to reproduce. Thus, based on our recent report using this assay, we detail a step-by-step protocol for the colorimetric detection of cortex fragments during bacterial spore germination.
Keywords: Infection, Issue 112, bacteria, spore, cortex, germination, microbiology, Ehrlich's reagent, N-acetylglucosamine, reducing sugar
Introduction
Endospores are metabolically dormant forms of bacteria that allow bacteria to persist in unfavorable environments. In many spore-forming bacteria, spore formation is induced by nutrient deprivation but this process can be controlled by changes to pH, exposure to oxygen or other stresses1. While in their metabolically dormant spore form, bacteria resist UV radiation, desiccation, higher temperatures, and freezing2. Most of the knowledge on the sporulation process comes from studies in the model organism, Bacillus subtilis. The sporulation process begins with DNA replication and the formation of an axial filament3,4. An asymmetric septum then divides the cell into two unequally sized compartments. The larger compartment, the mother cell, engulfs the smaller compartment, the forespore. Both the mother cell and forespore coordinate gene expression to mature the spore and the mother cell eventually lyses, releasing the dormant spore into the surrounding environment1.
The spore structure and composition is conserved across many bacterial species. The spore core has a lower water content than the vegetative cell and is rich with dipicolinic acid (DPA)1,2. During spore formation, DPA is pumped into the spore core in exchange for water. Surrounding the core is an inner spore membrane where, in most spore forming bacteria, the receptors which recognize the small molecules that stimulate germination (germinants) are located2. Located just outside the spore's inner membrane is a layer of cell wall peptidoglycan. A specialized peptidoglycan layer (cortex) surrounds the cell wall peptidoglycan and is composed of many of the same components as cell wall peptidoglycan [alternating N-acetylglucosamine (NAG) and N-acetylmuramic acid (NAM)]. However, approximately 50% of the NAM residues in the cortex have been converted to muramic-δ-lactam5,6. During spore germination, these muramic-δ-lactam residues are recognized by the spore cortex lytic enzymes (SCLEs), thus allowing the cortex to be degraded (a process essential to complete germination) but not the cell wall. Surrounding the cortex is an outer membrane and several layers of coat proteins 2.
Controlling the process of germination is critically important for spore-forming bacteria. Germination is initiated when germinant receptors interact with their respective germinants2. In many spore-forming bacteria, these germinants are amino acids, sugars, nucleotides, ions or combinations thereof2. C. difficile spore germination is initiated by combinations of certain bile acids, [e.g., taurocholic acid (TA)] and amino acids (e.g., glycine)7-10. Though there are differences between the C. difficile germination pathway and the pathways studied in other spore-forming bacteria, such as B. subtilis, common to all is the absolute requirement to degrade the spore cortex to allow a vegetative cell to grow from the germinated spore2,8. Cortex degradation can be accomplished by the SCLEs CwlJ/SleB (as found in B. subtilis) or SleC (as found in many Clostridia). Cortex hydrolysis reduces the constraint on the cell wall and the spore. This allows for full core rehydration, a necessary step in reactivating many of the proteins necessary for cellular metabolism 2.
When spores germinate, the dormant spore changes from a phase-bright state to a phase-dark state and this process can be measured by a change in optical density (OD) at 600nm11. A previous report suggests that much of this change in OD is due to the release of DPA from the spore12. During our recent study, we sought to compare the timing of C. difficile spore germination and monitored the release of DPA and cortex fragments9. For this study it was critical to monitor the release of cortex fragments as they began to be released by germinating spores.
The colorimetric assay used here was based upon a method for detecting sugars with reducing ends developed Ghuysen et al.13. Because others have described protocols for the detection of reducing sugars14 or have modified those protocols15, the literature on this subject can be confusing. Here, we detail a step-by-step method for the colorimetric detection of reducing sugars liberated from germinating C. difficile spores. Though this study uses C. difficile spores, the reducing sugars released during the germination of spores from other spore-forming bacteria are able to be detected with this protocol9,16,17.
Protocol
1. Generating Samples
Heat activate C. difficile spores at 65 °C for 30 min and store on ice. Note: C. difficile spores can be produced and purified as described previously7,9,10.
Prepare the germination solution at X + 1 ml where X is the number of samples to be taken during the assay. Note: The germination solution is specific to the species of bacteria spore being studied. For assaying C. difficile spore germination, we used a solution of 10 mM Tris (pH 7.5), 150 mM NaCl, 100 mM glycine and 10 mM taurocholic acid in deionized water.
Before beginning the assay, draw a 1.0 ml sample of the germination solution without spores to serve as a blank for cortex fragment detection.
Add spores to an OD600 of ~3.0 to the germination solution and vortex quickly. Note: For our germination studies in B. subtilis, we used the same OD600 spores as for C. difficile. Note: This amount of spores worked well for monitoring cortex fragment release during C. difficile spore germination. The amount of spores used in this assay should be determined experimentally for each bacterium or strain.
Remove a 1.2 ml sample immediately after adding the spores and centrifuge at room temperature for 1 min at 14,000 × g. This is the zero time point.
Transfer 1.0 ml of the supernatant from the centrifuged sample to a fresh tube for subsequent cortex fragment analysis. Note: Should DPA need to be monitored, remove a separate 100 μl sample for the detection of DPA in the solution using an assay based on terbium fluorescence9,18,19.
Freeze the collected sample at -80 °C.
Repeat steps 1.4 and 1.5 at regular intervals until all time points completed. Note: Because of time required for centrifugation and sampling, time points less than 2 min apart were not possible during our procedure, so our sampling intervals were once every 2 min for 14 min.
As a positive control for reducing sugar detection and quantification, include the following amounts of N-acetylglucosamine (nmol): 0, 12.5, 25, 50, 100, 250, 500, and 5,000. Prepare these samples at the same time as cortex samples and run along with the cortex samples so that the standard curve is relevant to the data generated by germinating spores.
After all time point samples are collected, lyophilize the samples. Note: We lyophilized our samples in 2 ml screw-cap tubes.
2. Measuring Cortex Fragments
Resuspend lyophilized samples in 120 µl of 3 N HCl supplemented with 1% phenol and 0.5% β-mercaptoethanol.
Transfer the resuspended samples to 2-ml screw-cap tubes. Note: For reproducibility, ensure all the lyophilized sample is resuspended and transferred.
Incubate the samples in a recirculating water bath at 95 °C for 4 hr.
After incubation, place the samples on ice until they are cool.
Neutralize the samples with 120 µl of 3 M NaOH.
Add 80 µl of a saturated sodium bicarbonate solution and 80 µl of a 5% acetic anhydride solution to each sample and vortex.
Incubate the samples at room temperature for 10 min.
Transfer the samples back to the 95 °C water bath for EXACTLY 3 min. Note: This step is time sensitive and longer times can affect downstream signal development.
Remove samples from the water bath and cool on ice.
Add 400 µl of 6.54% K2B4O7·4H2O to each tube and vortex.
Incubate the samples in a recirculating water bath at 95 °C for EXACTLY 7 min. Note: This step is time sensitive and longer times can affect downstream signal development.
Remove the samples and place on ice for 5 min.
3. Color Reagent
While samples are on ice (Step 2.12), prepare the color reagent. Dissolve 0.320 g of p-dimethylaminobenzaldehyde (DMAB; Ehrlich's reagent) in 1.9 ml of glacial acetic acid. Note: The reagent is time, temperature and light sensitive and should not be made in advance.
After the DMAB completely dissolves, add 100 µl of 10 N HCl and vortex.
Add 5 ml of glacial acetic acid and vortex.
4. Colorimetric Reaction
Transfer 100 µl of each cooled cortex sample to a new 1.5 ml microcentrifuge tube.
Add 700 µl of the freshly-prepared color solution to each sample.
Immediately transfer the samples to a 37 °C water bath and incubate for EXACTLY 20 min.
Transfer 200 µl of each sample to a clear 96-well plate.
Quantify the samples at 585 nm on a plate reader. Samples should start at a yellow color and a positive reaction will shift to purple. The more cortex present, the deeper the purple color.
Representative Results
Table 1 shows typical results using NAG standards. Data are used to generate a standard curve. Table 2 shows typical results from a germination assay in germination buffer supplemented with 100 mM glycine and 10 mM TA (germination-promoting conditions) or 100 mM glycine only (non-germinating conditions). In the absence of TA, C. difficile spores do not germinate and there is little change in the presence of cortex fragments in the solution. However, in the presence of both TA and glycine, C. difficile spores germinate and release cortex fragments into the solution. This is observed as an increase in the OD585 over time of the reacted samples. The total amount of cortex fragments (nmol) released during this assay was calculated using a standard curve similar to Figure 1B. In this assay, most of the cortex is hydrolyzed (or released) by 15 minutes post-germinant exposure, suggesting that most of the spores have germinated in this time frame (Figure 2; consistent with a previous publication7).
After developing the samples in the color reagent, most samples may not show the purple color to the naked eye. Figure 1 shows a representative sample generated from N-acetylglucosamine. Many of the samples do not show a visible purple color. However, the 250 nmol, 500 nmol and 5,000 nmol samples appear visually different from the negative control (0 nmol). When measured at OD585 nm, the signal generated best fits a linear regression (R2 = 0.999).
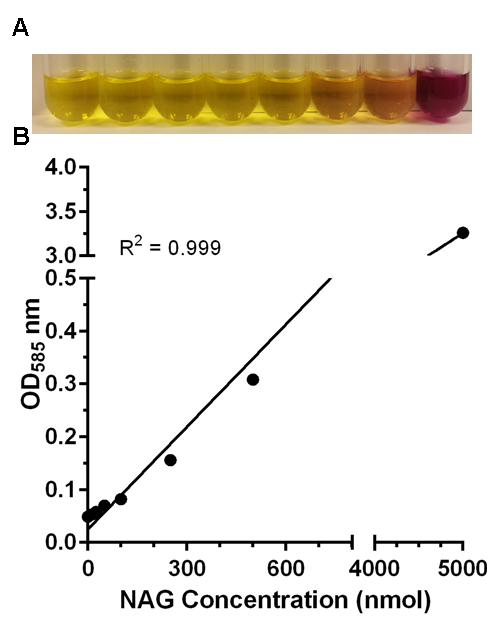 Figure 1:Detecting N-acetylglucosamine. (A) N-acetylglucosamine samples (0 nmol, 12.5 nmol, 25 nmol, 50 nmol, 100 nmol, 250 nmol, 500 nmol, 5,000 nmol) were lyophilized and reacted according to the described procedure. (B) Samples were added to a 96-well plate and the amount of reacted material detected at 585 nm. The signal was used to generate a standard curve.
Figure 1:Detecting N-acetylglucosamine. (A) N-acetylglucosamine samples (0 nmol, 12.5 nmol, 25 nmol, 50 nmol, 100 nmol, 250 nmol, 500 nmol, 5,000 nmol) were lyophilized and reacted according to the described procedure. (B) Samples were added to a 96-well plate and the amount of reacted material detected at 585 nm. The signal was used to generate a standard curve. 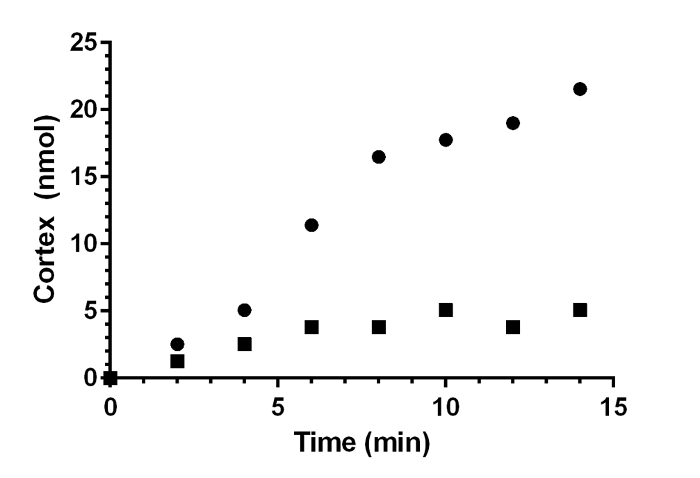 Figure 2:
Graphical Representation of Germination Results. The calculated nmol cortex in Table 2 was plotted vs. time. (●) 100 mM glycine + 10 mM TA; (■) 100 mM glycine. Please click here to view a larger version of this figure.
Figure 2:
Graphical Representation of Germination Results. The calculated nmol cortex in Table 2 was plotted vs. time. (●) 100 mM glycine + 10 mM TA; (■) 100 mM glycine. Please click here to view a larger version of this figure.
| NAG Concentration (nmol) | OD585 |
| 0 | 0.047 |
| 12.5 | 0.054 |
| 25 | 0.066 |
| 50 | 0.084 |
| 100 | 0.128 |
| 250 | 0.285 |
| 500 | 0.556 |
| 5,000 | 4.0 |
| blank = | 0.046 |
Table 1: NAG Standard.
| Clostridium difficile Spore Germination | ||||
| Germination Solution (10 mM Tris (pH 7.5), 150 mM NaCl) | ||||
| 100 mM Glycine + 10 mM TA | 100 mM Glycine | |||
| Time (minutes) | OD585 | Calculated (nmol) | OD585 | Calculated (nmol) |
| 0 | 0.046 | 0.00 | 0.047 | 0.00 |
| 2 | 0.048 | 2.54 | 0.048 | 1.27 |
| 4 | 0.05 | 5.07 | 0.049 | 2.54 |
| 6 | 0.055 | 11.41 | 0.05 | 3.80 |
| 8 | 0.059 | 16.49 | 0.05 | 3.80 |
| 10 | 0.06 | 17.76 | 0.051 | 5.07 |
| 12 | 0.061 | 19.02 | 0.05 | 3.80 |
| 14 | 0.063 | 21.56 | 0.051 | 5.07 |
Table 2: Sample Spore Germination Results.
Discussion
Upon stimulation, spores undergoing the process of germination lose their resistance properties without the need for macromolecular synthesis. When spore germination is triggered, the spore core releases DPA in exchange for water2. Due to the high DPA content of the dormant spore, the spore core is under intense osmotic pressure and the specialized peptidoglycan, cortex, helps prevent the core from swelling by acting, presumably, as a barrier to expansion2.
The method described above uses Ehrlich's reagent (a yellow solution) to detect the presence of reducing sugars as a measurable shift to a purple tint. During cortex hydrolysis, NAG and NAM sugars are liberated (depending on the extent of hydrolyzing enzymes, the sugars can be liberated as disaccharides or polysaccharides). Hydrolysis of the glycosidic linkages by HCl (Step 2.1) generates monosaccharides with free reducing ends7. After neutralizing the acid with an equal amount of sodium hydroxide (Step 2.5), the alcohols surrounding the NAG and NAM monosaccharides are acylated by acetic anhydride (Step 2.6)13. Subsequently, unreacted acetic anhydride is converted to acetic acid by heating the sample (Step 2.8). As hypothesized by Morgan and Elson20, the terminal aldehyde spontaneously converts to the enol form. Under alkaline conditions (Step 2.10), this enol form can react with the adjacent acetyl to generate an oxazole derivative which reacts with Ehrlich's reagent (Step 4.3) to generate a purple color15,20.
This method described above yields consistent, reliable results for the detection of cortex fragments from germinating spores, both B. subtilis and C. difficile9,16,17. The assay requires special attention to ensure the reaction times, temperatures and procedures are performed with precision. Without proper attention to detail, it is easy for the reaction conditions to fail or not reproduce between experiments13. However, the inclusion of a NAG standard aids in the application of the assay between laboratory environments. The NAG positive control standard included in every experiment controls for any inter-experimental variability (e.g., slight variations in reaction times or effectiveness of the color reagent). Reactions using only buffer conditions and reactions with spores incapable of hydrolyzing cortex both yielded no change in color.
The most common problem with this method is inter-experimental variability or complete lack of signal. To control for the variability, the spore preparations must be free of any vegetative cell debris. This cell debris will contain peptidoglycan fragments (NAG and NAM) and will result in a high background signal for the germinating spores. To ensure a pure spore preparation (>99.9% spores), refer to the literature for methods to prepare pure spore suspensions in the organism to be studied7,9,21-23.
If the assay is not yielding any detectable signal, we observed that the lyophilizing step can introduce variability. Due to the force with which the vacuum is released, we observed that the dried sample powder can be blown out of the sample tube. To reduce/prevent this from occurring, use a small syringe needle to poke holes in the tops of sample tubes. The small area of the introduced holes will reduce the likelihood of the lyophilized sample (now in powder form) from being disturbed. Finally, as noted in the methods above, the several incubation steps are very time sensitive. Ensure that all timed steps are monitored carefully. An important consideration for this assay is that the cortex fragments must be released from the germinating spore in order to be detected. If the cortex is hydrolyzed into large fragments, these fragments may be too large to be released through the spore coat. Though an important consideration when troubleshooting an experiment that does not yield a signal, we recommend to confirm that the conditions used to germinate most of the spores in the solution and the other troubleshooting options listed here prior to considering cortex fragment size as a potential problem in the assay.
Not all germination buffers/conditions are suitable for this assay. Because this assay detects the presence of reducing sugars liberated during the germination process13, any germination buffer that contains reducing sugars or any germinant that is, itself, a reducing sugar (e.g., glucose) will result in a strong signal in this assay. If this is the case, high pressure liquid chromatography (HPLC) can be used to detect the presence of cortex fragments24-26. However, because not all laboratories are equipped with such instrumentation, the method described herein can be used by most laboratories and offers simple and easily quantifiable results.
Germination by bacterial spores has largely been studied in Bacillus and a few Clostridium species2,8. The process is generally conserved, however differences exist. Importantly, not all organisms germinate using mechanisms of spore germination as observed in the model spore-formers9,27. Because germination is becoming studied in more diverse organisms, the assay described above can be applied to these organisms to detect cortex fragments during spore germination. By applying this method to these newly studied organisms, researchers can determine the role various germination proteins play in cortex hydrolysis.
Disclosures
The authors have nothing to disclose.
Acknowledgments
The Project described is supported by Award Number 5R01AI116895 from the National Institute of Allergy and Infectious Diseases. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases or the National Institutes of Health.
References
- Al-Hinai MA, Jones SW, Papoutsakis ET. The Clostridium sporulation programs: diversity and preservation of endospore differentiation. Microbiol Mol Biol Rev. 2015;79:19–37. doi: 10.1128/MMBR.00025-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Setlow P. Germination of spores of Bacillus species: what we know and do not know. J Bacteriol. 2014;196:1297–1305. doi: 10.1128/JB.01455-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan IS, Ramamurthi KS. Spore formation in Bacillus subtilis. Environ Microbiol Rep. 2014;6:212–225. doi: 10.1111/1758-2229.12130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fimlaid KA, Shen A. Diverse mechanisms regulate sporulation sigma factor activity in the Firmicutes. Curr Opin Microbiol. 2015;24:88–95. doi: 10.1016/j.mib.2015.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popham DL, Helin J, Costello CE, Setlow P. Analysis of the peptidoglycan structure of Bacillus subtilis endospores. J Bacteriol. 1996;178:6451–6458. doi: 10.1128/jb.178.22.6451-6458.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atrih A, Zollner P, Allmaier G, Foster SJ. Structural analysis of Bacillus subtilis 168 endospore peptidoglycan and its role during differentiation. J Bacteriol. 1996;178:6173–6183. doi: 10.1128/jb.178.21.6173-6183.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorg JA, Sonenshein AL. Bile salts and glycine as cogerminants for Clostridium difficile spores. J. Bacteriol. 2008;190:2505–2512. doi: 10.1128/JB.01765-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paredes-Sabja D, Shen A, Sorg JA. Clostridium difficile spore biology: sporulation, germination, and spore structural proteins. Trends Microbiol. 2014;22(7) doi: 10.1016/j.tim.2014.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis MB, Allen CA, Sorg JA. Spore cortex hydrolysis precedes DPA release during Clostridium difficile spore germination. J Bacteriol. 2015;197(14) doi: 10.1128/JB.02575-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis MB, Allen CA, Shrestha R, Sorg JA. Bile acid recognition by the Clostridium difficile Germinant Receptor, CspC, is important for establishing infection. PLoS Pathog. 2013;9:e1003356. doi: 10.1371/journal.ppat.1003356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moir A, Smith DA. The Genetics of bacterial spore germination. Annu. Rev. Microbiol. 1990;44:531–553. doi: 10.1146/annurev.mi.44.100190.002531. [DOI] [PubMed] [Google Scholar]
- Zhang P, Kong L, Wang G, Setlow P, Li YQ. Combination of Raman tweezers and quantitative differential interference contrast microscopy for measurement of dynamics and heterogeneity during the germination of individual bacterial spores. Journal of biomedical optics. 2010;15:056010. doi: 10.1117/1.3494567. [DOI] [PubMed] [Google Scholar]
- Ghuysen J-M, Tipper DJ, Strominger JL. Enzymes that degrade bacterial cell walls. Methods Enzymol. 1966;8:685–699. [Google Scholar]
- Park JT, Johnson MJ. A submicrodetermination of glucose. J Biol Chem. 1949;181:149–151. [PubMed] [Google Scholar]
- Thompson JS, Shockman GD. A modification of the Park and Johnson reducing sugar determination suitable for the assay of insoluble materials: its application to bacterial cell walls. Anal Biochem. 1968;22:260–268. doi: 10.1016/0003-2697(68)90315-1. [DOI] [PubMed] [Google Scholar]
- Paredes-Sabja D, Setlow P, Sarker MR. SleC is essential for cortex peptidoglycan hydrolysis during germination of spores of the pathogenic bacterium Clostridium perfringens. J Bacteriol. 2009;191:2711–2720. doi: 10.1128/JB.01832-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paredes-Sabja D, Sarker MR. Effect of the cortex-lytic enzyme SleC from non-food-borne Clostridium perfringens on the germination properties of SleC-lacking spores of a food poisoning isolate. Can J Microbiol. 2010;56:952–958. doi: 10.1139/w10-083. [DOI] [PubMed] [Google Scholar]
- Hindle AA, Hall EA. Dipicolinic acid (DPA) assay revisited and appraised for spore detection. The Analyst. 1999;124:1599–1604. doi: 10.1039/a906846e. [DOI] [PubMed] [Google Scholar]
- Kort R, et al. Assessment of heat resistance of bacterial spores from food product isolates by fluorescence monitoring of dipicolinic acid release. Appl Environ Microbiol. 2005;71:3556–3564. doi: 10.1128/AEM.71.7.3556-3564.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elson LA, Morgan WT. A colorimetric method for the determination of glucosamine and chondrosamine. The Biochemical journal. 1933;27:1824–1828. doi: 10.1042/bj0271824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramirez N, Abel-Santos E. Requirements for germination of Clostridium sordellii spores in vitro. J. Bacteriol. 2010;192:418–425. doi: 10.1128/JB.01226-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akoachere M, et al. Indentification of an in vivo inhibitor of Bacillus anthracis spore germination. J. Biol. Chem. 2007;282:12112–12118. doi: 10.1074/jbc.M611432200. [DOI] [PubMed] [Google Scholar]
- Duncan CL, Strong DH. Improved medium for sporulation of Clostridium perfringens. Appl Microbiol. 1968;16:82–89. doi: 10.1128/am.16.1.82-89.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heffron JD, Lambert EA, Sherry N, Popham DL. Contributions of four cortex lytic enzymes to germination of Bacillus anthracis spores. J Bacteriol. 2010;192:763–770. doi: 10.1128/JB.01380-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meador-Parton J, Popham DL. Structural analysis of Bacillus subtilis spore peptidoglycan during sporulation. J Bacteriol. 2000;182:4491–4499. doi: 10.1128/jb.182.16.4491-4499.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert EA, Popham DL. The Bacillus anthracis SleL (YaaH) protein is an N-acetylglucosaminidase involved in spore cortex depolymerization. J Bacteriol. 2008;190:7601–7607. doi: 10.1128/JB.01054-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Shen A, Setlow P, Li YQ. Characterization of the Dynamic Germination of Individual Clostridium difficile Spores Using Raman Spectroscopy and Differential Interference Contrast Microscopy. J Bacteriol. 2015;197:2361–2373. doi: 10.1128/JB.00200-15. [DOI] [PMC free article] [PubMed] [Google Scholar]