

ABSTRACT
Several signaling pathways, including the JAK/STAT and Toll pathways, are known to activate blood cells (hemocytes) in Drosophila melanogaster larvae. They are believed to regulate the immune response against infections by parasitoid wasps, such as Leptopilina boulardi, but how these pathways control the hemocytes is not well understood. Here, we discuss the recent discovery that both muscles and fat body take an active part in this response. Parasitoid wasp infection induces Upd2 and Upd3 secretion from hemocytes, leading to JAK/STAT activation mainly in hemocytes and in skeletal muscles. JAK/STAT activation in muscles, but not in hemocytes, is required for an efficient encapsulation of wasp eggs. This suggests that Upd2 and Upd3 are important cytokines, coordinating different tissues for the cellular immune response in Drosophila. In the fat body, Toll signaling initiates a systemic response in which hemocytes are mobilized and activated hemocytes (lamellocytes) are generated. However, the contribution of Toll signaling to the defense against wasps is limited, probably because the wasps inject inhibitors that prevent the activation of the Toll pathway. In conclusion, parasite infection induces a systemic response in Drosophila larvae involving major organ systems and probably the physiology of the entire organism.
KEYWORDS: Drosophila, fat body, JAK/STAT, muscles, tissue communication, Toll
Introduction
In contrast to vertebrates, which utilize both innate and adaptive immune responses to battle infection, flies are thought to rely entirely on their well-developed innate immune system. Depending on the type of infection, different functions of this immune system are mobilized. For example, microbes induce a humoral immune response, which is mediated by antimicrobial peptides that are secreted by the fat body, hemocytes, or epithelia,1,2 and controlled by 2 conserved signaling pathways, the Toll and IMD pathways.3,4 Eggs laid by parasitoid wasps induce a different type of response, triggering hemocytes (blood cells) to attack and enclose the parasite in a melanized capsule. This cellular immune response involves 3 morphologically distinguishable types of hemocytes: plasmatocytes, lamellocytes, and crystal cells. Lamellocytes are specialized cells that are only seen in the context of an infection and they participate in the encapsulation of the parasite. Several signaling pathways are thought to be involved in this encapsulation response. For instance, loss-of-function mutations in the JAK/STAT (Janus kinase/signal transducers and activators of transcription) or Toll pathways reduce the encapsulation response against wasp infection,5 while artificial activation of the same pathways triggers a spontaneous immune response, causing hemocytes to aggregate in melanized masses, in a way that is reminiscent of the encapsulation response.6-11 Altogether, these findings suggested that the JAK/STAT and Toll pathways play a positive role in the cellular defense against wasp infection. Recently, the role of these pathways has turned out to be unexpectedly complex. The cellular response is orchestrated in a systemic interaction between hemocytes, fat body and, surprisingly, the somatic musculature, and this interplay is controlled by JAK/STAT and Toll signaling in these tissues.12,13
JAK/STAT activation in muscles is important for cellular immunity
In Drosophila, JAK/STAT signaling involves at least 3 cytokines, Unpaired 1, 2 and 3 (Upd1-3), the receptor Domeless (Dome), the tyrosine kinase Hopscotch (Hop), and the transcription factor Stat92E.14-16 In order to study the role of this pathway in the cellular immune response we used the 10xStat-GFP reporter17 to visualize JAK/STAT activity in vivo in wasp-infected Drosophila larvae. Surprisingly we observed JAK/STAT activation not only in hemocytes, but also in muscles.12 This effect, illustrated in Figure 1, was unexpected. So far, research on the cellular immune response in Drosophila has focused on what happens in circulating hemocytes and in the haematopoietic tissues of “lymph glands” and sub-epithelial clusters of sessile cells.15,18,19 Sessile hemocytes have been shown to interact with peripheral neurons,20 and circulating hemocytes are known to communicate with the fat body as well as with wounds and tumors,13,21,22 but it was not obvious why the body musculature should also respond to the presence of a parasite egg.
Figure 1.
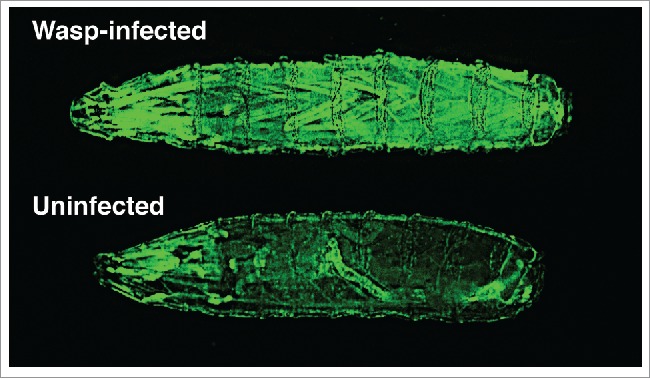
Example of JAK/STAT activation in a Drosophila larva, infected by the wasp Leptopilina boulardi. Stat92E-dependent transcription is visualized with the 10xStat92E-GFP reporter.17 The figure is a montage of 2 photographs, identically exposed and enhanced.
We further tested the role of the JAK/STAT pathway in the immune response by suppressing it separately in those tissues where it was most strongly activated, in plasmatocytes and muscles. It turned out that suppression of JAK/STAT signaling in muscles, but not in hemocytes, significantly reduced the encapsulation of parasite eggs, suggesting that muscles take an active part in the immune response.12 Occasionally, wasp infection also triggered JAK/STAT activity in a small number of fat body cells but, as shown in Figure 2C, the encapsulation response was not significantly affected when we suppressed JAK/STAT pathway in fat body by expressing the domeDN dominant-negative construct with the fat body-specific FB-Gal4 driver. In conclusion, JAK/STAT-dependent events in the somatic muscles are of particular importance for a successful defense against a parasitoid wasp. Thus, Drosophila larval muscles, rather than acting as bystanders, actively participate in the cellular immune response against wasp infection.
Figure 2.
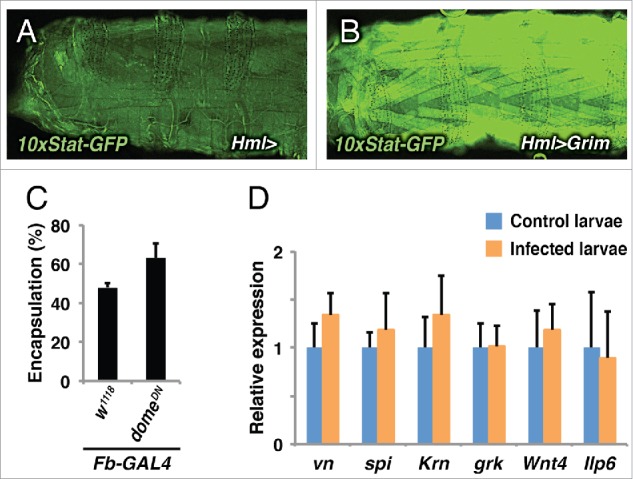
Role of JAK/STAT pathway during cellular immune response (A-B). Compared to the control (A), the JAK/STAT pathway (10xStat92E-GFP reporter) is activated in muscles when apoptosis is induced in hemocytes (B). C. The encapsulation of wasp eggs is not affected when JAK/STAT signaling is suppressed in the fat body (FB-Gal4>domeDN). Encapsulation rates were scored after 26 hours of wasp infection. Histograms represent average ± standard deviation of at least 3 independent experiments (unpaired t-test). D. The relative expression of the genes for the indicated candidate signaling in skeletal muscles were assayed by quantitative PCR in control larvae (blue bars) and wasp infected larvae (orange bars). Histograms represent average ± standard deviation of at least 4 independent experiments (unpaired t-test).
Hemocyte signaling to the muscles: The cytokines Upd2 and Upd3
It remains to find out how infection can activate the muscles in the first place and how muscles can affect the cellular immune defense. A partial answer to the first question was found when we traced the origin of the cytokines that stimulate the JAK/STAT pathway. Using reporter strains and quantitative RT-PCR, we found that wasp infection induces the cytokine genes upd2 and upd3 in hemocytes.12 The upd1 gene is not induced under these conditions, and we saw no induction of upd2 or upd3 in other tissues. Furthermore, upd2 and upd3 single or double mutants have an impaired cellular immune response against wasp infection. Conversely, artificial overexpression of upd2 or upd3 in hemocytes triggers JAK/STAT activity in muscles, causing hemocytes to differentiate into activated lamellocytes.12 In conclusion, the hemocytes themselves send a signal to the muscles, via the cytokines Upd2 and Upd3, and the response of the muscles is in turn required for hemocytes to efficiently attack the parasite.
The Unpaired cytokines have also been implicated as stress signals under several other circumstances, such as bacterial infection, tumor response, wounding, and virus infection. For instance, bacteremia induces Upd3 production in hemocytes of the adult fly, which activates JAK/STAT signaling in the fat body.23 This induces the Turandot A gene, which encodes a secreted stress peptide of uncertain function.24 In this case, the response depends on the simultaneous activation of the transcription factor Relish in the fat body.23 Furthermore, bacterial infection in the gut induces the unpaired genes, particularly upd3, in intestinal cells. This promotes epithelial regeneration in the gut, which is essential for defense against bacterial infection.25 As discussed below, this effect is mediated via JAK/STAT activation in the nearby visceral muscles, which respond by secreting the growth factor Vein.26 A third example is the response to tumors or wounds, which triggers JNK activity and induces Upd1, Upd2, and Upd3 production in local tissues. These cytokines activate the JAK/STAT pathway both in hemocytes and in fat body, which in turn help to restrict tumor growth or defend against infection following wounding.21 Finally, Drosophila C virus or cricket paralysis virus infection induce Upd2 and Upd3 production, and the JAK/STAT pathway contributes to the protection against these viruses.27
These examples suggest that Upd2 and Upd3 may be specialized coordinators of a systemic response to infection. Depending on the circumstances, different tissues are activated in a joint effort to fight infection. Otherwise, the JAK/STAT pathway plays an important role during development, and strong upd1 mutant alleles are lethal during embryonic stage.28 However, upd2 and upd3 single or double mutants are viable and do not show any obvious developmental defects25 but they encode cytokines that are essential for inter-tissue communication during infection and wounding.
In an attempt to further test the conclusion that hemocytes are the main contributors of Upd2 and Upd3 production upon wasp infection, we tried to ablate circulating hemocytes by expressing an apoptosis gene, grim, with the hemocyte-specific Hml-GAL4 driver. Surprisingly, we found that the apoptotic hemocytes induced JAK/STAT activation in muscles (Fig. 2A-B). However, this is in line with the observation that apoptosis in hemocytes also induces lamellocyte formation, a hallmark of an activated cellular immune response.29 Similarly, when we artificially activated the JNK or p38 signaling pathways, or even JAK/STAT itself, in hemocytes, we saw a JAK/STAT response in the muscles.12 As a result, hemocytes are activated and lamellocytes are generated.11 This may appear paradoxical, since the cellular immune response is essentially normal even when the JNK, p38 or JAK/STAT signaling pathways are blocked in these cells.12 Apparently, cytokine production can be triggered by several different perturbations of the signaling systems in hemocytes. The cytokines, Upd2 and Upd3, will then transmit a danger signal to other tissues, where the effect on the somatic musculature is particularly important for the cellular immune response.
Local cross-talk between intestine and visceral muscles
Although it is still not clear how somatic muscles of the Drosophila larva affect the cellular immune response, it is better understood how muscles control local responses in the Drosophila intestine, where visceral muscles are tightly connected with intestinal trachea, neurons and the gut itself. These tissues cross-talk in an intimate local interaction during gut regeneration and immune response. Visceral muscles secrete several proteins that non-autonomously regulate regeneration in neighboring tissues. For instance, Wingless (Wg), a ligand for the Wnt pathway, is secreted by the visceral muscles, regulating intestinal stem cell maintenance and proliferation under damage or stress.30,31 Furthermore, during enteric infection or damage, gut epithelial cells (enterocytes) and their precursors (the enteroblasts) secrete Upd3, which activates JAK/STAT signaling in the visceral muscles and in the enteroblasts themselves. This triggers the visceral muscles to secrete Vein (Vn), one of the EGFR pathway ligands. The stimulated enteroblasts secrete a second EGFR ligand, Spitz (Spi). Together, Vein and Spitz remotely activate EGFR signaling in the intestinal stem cells, stimulating stem cell proliferation.26,32–35
To test if similar mechanisms are involved when skeletal muscles activate hemocytes, we assayed the expression of the genes for 4 known Drosophila EGFR ligands, Vein, Spitz, Keren and Gurken, in the skeletal muscles, but none of them was significantly induced in wasp-infected larvae (Fig. 2D). To test the role of Wnt signaling we also assayed Wnt4, the only Wnt-like gene identified as a JAK/STAT target in Drosophila tissue culture cells,36 with a negative result. Nor could we detect a significant effect on the Ilp6 gene, encoding an insulin-like peptide (Fig. 2D). Thus, the question remains how JAK/STAT signaling in the skeletal muscles promote the cellular immune response during wasp infection.
Systemic effects of somatic muscles on general physiology
Several studies during the past decade have demonstrated a major role of skeletal muscles in systemic responses to exercise, stress and aging, in mammals as well as in the model organisms Drosophila melanogaster and Caenorhabditis elegans.37-39 Muscles secrete myokines and metabolites with major systemic effects on metabolism, controlling physiological homeostasis, aging and disease progression. The role of these factors in immunity is less well understood.
In Drosophila, suppression of the insulin pathway in skeletal muscles leads to reduced muscle size and decreased feeding behavior, which finally affects the growth of other tissues.40 Overexpression of FOXO and its target 4E-BP in skeletal muscles not only prevents aging in muscles but also in other tissues, by decreased accumulation of protein aggregates and increased autophagy.41,42 Recently Demontis et al.43 also reported that Myoglianin, a myokine, plays an important role in tissue communication between Drosophila skeletal muscles and adipose tissue during aging.
In mammals, interleukin-6 (IL-6) is perhaps the most thoroughly studied myokine.37 IL-6 was found to be secreted in vast amounts by human skeletal contracting muscles during exercise, causing plasma levels to increase by almost 2 orders of magnitude.44,45 Not only exercise but also other stimuli, such as bacterial lipopolysaccharides, reactive oxygen species, and inflammatory cytokines, induce IL-6 production from skeletal muscles.46-49 Secreted IL-6 exerts a positive or negative effect on metabolism and insulin sensitivity, depending on the physiological or disease context.39 Other myokines, including insulin-like growh factor 1(IGF-1), IL-8, IL15, and meteorin-like (Metrnl), with different effects on physiology, were shown to be secreted by muscles.37,50 Interestingly, Metrnl was reported to affect the immune system. Metrnl, induced by exercise training, does not only regulate energy homeostasis, but this factor also stimulates several immune cell subtypes to enter the adipose tissues and secrete anti-inflammatory factors.50
By analogy to these observations it is possible that the somatic muscles in Drosophila may control immunity by secreting still unknown myokines that act directly on the immune cells. Alternatively, they may affect immunity indirectly, by acting as a master switch of metabolism. The skeletal musculature would then be a central player in a systemic response, regulating nutrient availability according to the needs of the cellular immune defense. A similar mechanism may also be involved in the wasting of muscles and adipose tissue seen in human chronic disease.
Toll signaling and cellular immunity
The fat body is a bulky and dominating tissue in the growing larva, with important functions for fat and protein storage and for general metabolism.51 It is also important in the humoral immune response, being the main site of synthesis of antimicrobial peptides. The role of the Toll pathway in this response is well documented.1,2,4,52–54 More recently it has become clear that the fat body can also activate a hemocyte response in Drosophila larvae, and that this effect also depends on activation of Toll signaling in the fat body.13 It had previously been shown that constitutively active Toll gain-of-function mutants such as Toll10b show signs of an activated cellular immune response, for instance, loss of sessile hemocytes, lamellocyte formation and formation of melanotic masses of activated hemocytes, while loss-of function Toll mutants have a reduced capacity to encapsulate parasitoid wasp eggs.5,8–10 It is sufficient to express the Toll10b mutant in hemocytes to generate a Toll gain-of-function phenotype,11 but recently it was found that artificial Toll activation in other tissues, notably in the fat body, gave an even stronger hemocyte activation phenotype.13 The hemocyte phenotype of the Toll10b mutant, expressed under its endogenous Toll promoter, was suppressed when downstream components in the Toll pathway were knocked down in fat body, but not in hemocytes.13 These results suggest that Toll activation in the fat body is a more important factor for the cellular immune response than Toll activity in the hemocytes themselves.
The involvement of muscles and fat body in the cellular immune response is schematically depicted in Figure 3. While JAK/STAT activity in the somatic muscles is necessary and probably sufficient for a strong cellular immune response, Toll signaling in the fat body is sufficient to trigger hemocyte activation, but it is not absolutely necessary for the defense against wasp eggs. Only a minority of wasp-infected larvae shows a strong Toll response, but a positive correlation was seen between the Toll response and the ability to fight the infection.13 Interestingly, a more consistent Toll response was seen in larvae that the wasp had stung, without injecting an egg. This suggests that together with the egg, the wasp injects an inhibitor of Toll activation.
Figure 3.
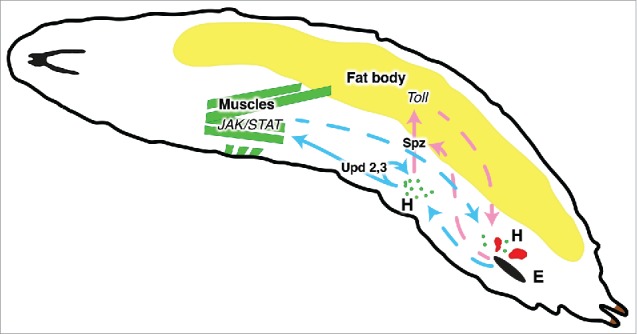
Schematic representation of interactions between a wasp egg (E), hemocytes (H), muscles and fat body. Plasmatocytes are shown in green, lamellocytes in red. Interactions via identified cytokines are shown as solid arrows. Other interactions (which may be indirect) are shown as dashed arrows.
The ligand of the receptor Toll is the TGFβ-related cytokine Spätzle, which is constitutively secreted by hemocytes. It can be further induced, for instance by bacterial infection,55 but it must be processed by proteolytic cleavage before it is active.56 In the Drosophila embryo, Spätzle is processed by the serine protease Easter while a related protein, the Spätzle-Processing Enzyme (SPE), does the same job in the hemolymph. Both are in turn activated by complement-like serine protease cascades,4,54 but it is not clear how, or if, Spätzle is processed in the context of a wasp infection. Nor is it clear how Toll signaling in the fat body leads to a mobilization of the cellular immune response. One interesting candidate signaling molecule is Edin, which is secreted by the fat body in response to wasp infection and required for mobilization of sessile hemocytes, but not for lamellocyte formation.57,58
Other tissues involved in the immune response
Further examples of tissue interactions in the cellular immune response of Drosophila have been reported recently, for instance between neurons and sessile hemocytes and between fat body and brain.57 During larval development, specific neurons in the peripheral nervous system supply a microenvironment in “pockets” between the epidermis and the somatic muscles. These subepidermal sites attract colonizing hemocytes and provide inductive factors for hemocyte expansion. These resident hemocytes co-localize with particular peripheral neurons and the ablation of these neurons leads to loss of resident hemocytes.59 Apparently, communication between peripheral neurons and subepidermal hemocytes is critical for the maintenance of this sessile hemocyte population.
Furthermore, high dietary fat and sugar can induce Upd2 secretion from the fat body.60 Secreted Upd2 indirectly stimulates insulin-producing cells in the brain to release insulin, which systemically affects growth and metabolism.60 It has also been reported that a lipid-rich diet stimulates Drosophila hemocytes to secrete Upd3, which systemically impairs glucose homeostasis and reduces lifespan.61
Conclusions
It is clear that the response to an infection in Drosophila larvae, for instance by a parasitic wasp, is not a local phenomenon that is restricted to a limited number of specialized cells of the immune system. On the contrary, it is a major event that affects the physiology of the entire organism and it involves all major organ systems in a concerted systemic immune response. The findings we have discussed here may also have relevance for the relationship between physical activity, nutrition, life span and immunity, in flies as well as in man.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We apologize that space limitations have prevented us from citing many relevant articles.
Funding
The authors were supported by grants from the Swedish Research Council, the Swedish Cancer Society, the Kempe Foundation, the Academy of Finland and the Sigrid Juselius Foundation.
References
- [1].Hultmark D. Drosophila immunity: paths and patterns. Curr Opin Immunol 2003; 15:12-9; PMID:12495727; http://dx.doi.org/ 10.1016/S0952-7915(02)00005-5 [DOI] [PubMed] [Google Scholar]
- [2].Lemaitre B, Hoffmann J. The host defense of Drosophila melanogaster. Annu Rev Immunol 2007; 25:697-743; PMID:17201680; http://dx.doi.org/ 10.1146/annurev.immunol.25.022106.141615 [DOI] [PubMed] [Google Scholar]
- [3].Kleino A, Silverman N. The Drosophila IMD pathway in the activation of the humoral immune response. Dev Comp Immunol 2014; 42:25-35; PMID:23721820; http://dx.doi.org/ 10.1016/j.dci.2013.05.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Lindsay SA, Wasserman SA. Conventional and non-conventional Drosophila Toll signaling. Dev Comp Immunol 2014; 42:16-24; PMID:23632253; http://dx.doi.org/ 10.1016/j.dci.2013.04.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Sorrentino RP, Melk JP, Govind S. Genetic analysis of contributions of dorsal group and JAK-Stat92E pathway genes to larval hemocyte concentration and the egg encapsulation response in Drosophila. Genetics 2004; 166:1343-56; PMID:15082553; http://dx.doi.org/ 10.1534/genetics.166.3.1343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Luo H, Hanratty WP, Dearolf CR. An amino acid substitution in the Drosophila hopTum-l Jak kinase causes leukemia-like hematopoietic defects. EMBO J 1995; 14:1412-20; PMID:7729418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Harrison DA, Binari R, Nahreini TS, Gilman M, Perrimon N. Activation of a Drosophila Janus kinase (JAK) causes hematopoietic neoplasia and developmental defects. EMBO J 1995; 14:2857-65; PMID:7796812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Gerttula S, Jin Y, Anderson KV. Zygotic expression and activity of the Drosophila Toll gene, a gene required maternally for embryonic dorsal-ventral pattern formation. Genetics 1988; 119:123-33; PMID:2456252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Lemaitre B, Meister M, Govind S, Georgel P, Steward R, Reichhart J-M, Hoffmann JA. Functional analysis and regulation of nuclear import of dorsal during the immune response in Drosophila. EMBO J 1995; 14:536-45; PMID:7859742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Qiu P, Pan PC, Govind S. A role for the Drosophila Toll/Cactus pathway in larval hematopoiesis. Development 1998; 125:1909-20; PMID:9550723 [DOI] [PubMed] [Google Scholar]
- [11].Zettervall CJ, Anderl I, Williams MJ, Palmer R, Kurucz E, Ando I, Hultmark D. A directed screen for genes involved in Drosophila blood cell activation. Proc Natl Acad Sci USA 2004; 101:14192-7; PMID:15381778; http://dx.doi.org/ 10.1073/pnas.0403789101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Yang H, Kronhamn J, Ekström J-O, Korkut GG, Hultmark D. JAK/STAT signaling in Drosophila muscles controls the immune response against parasitoid infection. EMBO Rep 2015; 16:1664-72; PMID:26412855; http://dx.doi.org/ 10.15252/embr.201540277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Schmid MR, Anderl I, Vesala L, Vanha-aho L-M, Deng X-J, Rämet M, Hultmark D. Control of Drosophila blood cell activation via Toll signaling in the fat body. PLoS One 2014; 9:e102568; PMID:25102059; http://dx.doi.org/ 10.1371/journal.pone.0102568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Arbouzova NI, Zeidler MP. JAK/STAT signalling in Drosophila: insights into conserved regulatory and cellular functions. Development 2006; 133:2605-16; PMID:16794031; http://dx.doi.org/ 10.1242/dev.02411 [DOI] [PubMed] [Google Scholar]
- [15].Morin-Poulard I, Vincent A, Crozatier M. The Drosophila JAK-STAT pathway in blood cell formation and immunity. JAKSTAT 2013; 2:e25700; PMID:24069567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Myllymäki H, Rämet M. JAK/STAT pathway in Drosophila immunity. Scand J Immunol 2014; 79:377-85; http://dx.doi.org/ 10.1111/sji.12170 [DOI] [PubMed] [Google Scholar]
- [17].Bach EA, Ekas LA, Ayala-Camargo A, Flaherty MS, Lee H, Perrimon N, Baeg G-H. GFP reporters detect the activation of the Drosophila JAK/STAT pathway in vivo. Gene Expr Patterns 2007; 7:323-31; PMID:17008134; http://dx.doi.org/ 10.1016/j.modgep.2006.08.003 [DOI] [PubMed] [Google Scholar]
- [18].Williams MJ. Drosophila hemopoiesis and cellular immunity. J Immunol 2007; 178:4711-6; PMID:17404248; http://dx.doi.org/ 10.4049/jimmunol.178.8.4711 [DOI] [PubMed] [Google Scholar]
- [19].Honti V, Csordás G, Kurucz E, Márkus R, Andó I. The cell-mediated immunity of Drosophila melanogaster: Hemocyte lineages, immune compartments, microanatomy and regulation. Dev Comp Immunol 2014; 42:47-56; PMID:23800719; http://dx.doi.org/ 10.1016/j.dci.2013.06.005 [DOI] [PubMed] [Google Scholar]
- [20].Makhijani K, Bruckner K. Of blood cells and the nervous system: Hematopoiesis in the Drosophila larva. Fly (Austin) 2012; 6:254-60; PMID:23022764; http://dx.doi.org/ 10.4161/fly.22267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Pastor-Pareja JC, Wu M, Xu T. An innate response of blood cells to tumors and tissue damage in Drosophila. Dis Models Mech 2008; 1:144-54; discussion 53; http://dx.doi.org/ 10.1242/dmm.000950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Parisi F, Stefanatos RK, Strathdee K, Yu Y, Vidal M. Transformed epithelia trigger non-tissue-autonomous tumor suppressor response by adipocytes via activation of Toll and Eiger/TNF signaling. Cell Rep 2014; 6:855-67; PMID:24582964; http://dx.doi.org/ 10.1016/j.celrep.2014.01.039 [DOI] [PubMed] [Google Scholar]
- [23].Agaisse H, Petersen U-M, Boutros M, Mathey-Prevot B, Perrimon N. Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev Cell 2003; 5:441-50; PMID:12967563; http://dx.doi.org/ 10.1016/S1534-5807(03)00244-2 [DOI] [PubMed] [Google Scholar]
- [24].Ekengren S, Tryselius Y, Dushay MS, Liu G, Steiner H, Hultmark D. A humoral stress response in Drosophila. Curr Biol 2001; 11:714-8 (Addendum on p. 1479); PMID:11369236; http://dx.doi.org/ 10.1016/S0960-9822(01)00203-2 [DOI] [PubMed] [Google Scholar]
- [25].Osman D, Buchon N, Chakrabarti S, Huang YT, Su WC, Poidevin M, Tsai YC, Lemaitre B. Autocrine and paracrine unpaired signalling regulate intestinal stem cell maintenance and division. J Cell Sci 2012; 125:5944-9; PMID:23038775; http://dx.doi.org/ 10.1242/jcs.113100 [DOI] [PubMed] [Google Scholar]
- [26].Buchon N, Broderick NA, Kuraishi T, Lemaitre B. Drosophila EGFR pathway coordinates stem cell proliferation and gut remodeling following infection. BMC Biol 2010; 8:152; PMID:21176204; http://dx.doi.org/ 10.1186/1741-7007-8-152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Kemp C, Mueller S, Goto A, Barbier V, Paro S, Bonnay F, Dostert C, Troxler L, Hetru C, Meignin C, et al.. Broad RNA interference-mediated antiviral immunity and virus-specific inducible responses in Drosophila. J Immunol 2013; 190:650-8; PMID:23255357; http://dx.doi.org/ 10.4049/jimmunol.1102486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Harrison DA, McCoon PE, Binari R, Gilman M, Perrimon N. Drosophila unpaired encodes a secreted protein that activates the JAK signaling pathway. Genes Dev 1998; 12:3252-63; PMID:9784499; http://dx.doi.org/ 10.1101/gad.12.20.3252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Arefin B, Kucerova L, Krautz R, Kranenburg H, Parvin F, Theopold U. Apoptosis in hemocytes induces a shift in effector mechanisms in the Drosophila immune system and leads to a pro-inflammatory state. PLoS One 2015; 10:e0136593; PMID:26322507; http://dx.doi.org/ 10.1371/journal.pone.0136593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Lin G, Xu N, Xi R. Paracrine Wingless signalling controls self-renewal of Drosophila intestinal stem cells. Nature 2008; 455:1119-23; PMID:18806781; http://dx.doi.org/ 10.1038/nature07329 [DOI] [PubMed] [Google Scholar]
- [31].Cordero JB, Stefanatos RK, Scopelliti A, Vidal M, Sansom OJ. Inducible progenitor-derived Wingless regulates adult midgut regeneration in Drosophila. EMBO J 2012; 31:3901-17; PMID:22948071; http://dx.doi.org/ 10.1038/emboj.2012.248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Jiang H, Grenley MO, Bravo MJ, Blumhagen RZ, Edgar BA. EGFR/Ras/MAPK signaling mediates adult midgut epithelial homeostasis and regeneration in Drosophila. Cell Stem Cell 2011; 8:84-95; PMID:21167805; http://dx.doi.org/ 10.1016/j.stem.2010.11.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Biteau B, Jasper H. EGF signaling regulates the proliferation of intestinal stem cells in Drosophila. Development 2011; 138:1045-55; PMID:21307097; http://dx.doi.org/ 10.1242/dev.056671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Xu N, Wang SQ, Tan D, Gao Y, Lin G, Xi R. EGFR, Wingless and JAK/STAT signaling cooperatively maintain Drosophila intestinal stem cells. Dev Biol 2011; 354:31-43; PMID:21440535; http://dx.doi.org/ 10.1016/j.ydbio.2011.03.018 [DOI] [PubMed] [Google Scholar]
- [35].Zhou F, Rasmussen A, Lee S, Agaisse H. The UPD3 cytokine couples environmental challenge and intestinal stem cell division through modulation of JAK/STAT signaling in the stem cell microenvironment. Dev Biol 2013; 373:383-93; PMID:23110761; http://dx.doi.org/ 10.1016/j.ydbio.2012.10.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Bina S, Wright VM, Fisher KH, Milo M, Zeidler MP. Transcriptional targets of Drosophila JAK/STAT pathway signalling as effectors of haematopoietic tumour formation. EMBO Rep 2010; 11:201-7; PMID:20168330; http://dx.doi.org/ 10.1038/embor.2010.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Pedersen BK, Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat Rev Endocrinol 2012; 8:457-65; PMID:22473333; http://dx.doi.org/ 10.1038/nrendo.2012.49 [DOI] [PubMed] [Google Scholar]
- [38].Droujinine IA, Perrimon N. Defining the interorgan communication network: systemic coordination of organismal cellular processes under homeostasis and localized stress. Front Cell Infect Microbiol 2013; 3:82; PMID:24312902; http://dx.doi.org/ 10.3389/fcimb.2013.00082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Rai M, Demontis F. Systemic nutrient and stress signaling via myokines and myometabolites. Annu Rev Physiol 2016; 78:85-107; PMID:26527185; http://dx.doi.org/ 10.1146/annurev-physiol-021115-105305 [DOI] [PubMed] [Google Scholar]
- [40].Demontis F, Perrimon N. Integration of Insulin receptor/Foxo signaling and dMyc activity during muscle growth regulates body size in Drosophila. Development 2009; 136:983-93; PMID:19211682; http://dx.doi.org/ 10.1242/dev.027466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Demontis F, Perrimon N. FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging. Cell 2010; 143:813-25; PMID:21111239; http://dx.doi.org/ 10.1016/j.cell.2010.10.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Bai H, Kang P, Hernandez AM, Tatar M. Activin signaling targeted by insulin/dFOXO regulates aging and muscle proteostasis in Drosophila. PLoS Genet 2013; 9:e1003941; PMID:24244197; http://dx.doi.org/ 10.1371/journal.pgen.1003941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Demontis F, Patel VK, Swindell WR, Perrimon N. Intertissue control of the nucleolus via a myokine-dependent longevity pathway. Cell Rep 2014; 7:1481-94; PMID:24882005; http://dx.doi.org/ 10.1016/j.celrep.2014.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Steensberg A, van Hall G, Osada T, Sacchetti M, Saltin B, Klarlund Pedersen B. Production of interleukin-6 in contracting human skeletal muscles can account for the exercise-induced increase in plasma interleukin-6. J Physiol 2000; 529 (Pt 1):237-42; PMID:11080265; http://dx.doi.org/ 10.1111/j.1469-7793.2000.00237.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Ostrowski K, Rohde T, Zacho M, Asp S, Pedersen BK. Evidence that interleukin-6 is produced in human skeletal muscle during prolonged running. J Physiol 1998; 508 (Pt 3):949-53; PMID:9518745; http://dx.doi.org/ 10.1111/j.1469-7793.1998.949bp.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Frost RA, Nystrom GJ, Lang CH. Lipopolysaccharide regulates proinflammatory cytokine expression in mouse myoblasts and skeletal muscle. Am J Physiol Regul Integr Comp Physiol 2002; 283:R698-709; PMID:12185005; http://dx.doi.org/ 10.1152/ajpregu.00039.2002 [DOI] [PubMed] [Google Scholar]
- [47].Frost RA, Nystrom GJ, Lang CH. Lipopolysaccharide and proinflammatory cytokines stimulate interleukin-6 expression in C2C12 myoblasts: role of the Jun NH2-terminal kinase. Am J Physiol Regul Integr Comp Physiol 2003; 285:R1153-64; PMID:12842862; http://dx.doi.org/ 10.1152/ajpregu.00164.2003 [DOI] [PubMed] [Google Scholar]
- [48].Kosmidou I, Vassilakopoulos T, Xagorari A, Zakynthinos S, Papapetropoulos A, Roussos C. Production of interleukin-6 by skeletal myotubes: Role of reactive oxygen species. Am J Respir Cell Mol Biol 2002; 26:587-93; PMID:11970911; http://dx.doi.org/ 10.1165/ajrcmb.26.5.4598 [DOI] [PubMed] [Google Scholar]
- [49].Luo G, Hershko DD, Robb BW, Wray CJ, Hasselgren PO. IL-1beta stimulates IL-6 production in cultured skeletal muscle cells through activation of MAP kinase signaling pathway and NF-κB. Am J Physiol Regul Integr Comp Physiol 2003; 284:R1249-54; PMID:12676746; http://dx.doi.org/ 10.1152/ajpregu.00490.2002 [DOI] [PubMed] [Google Scholar]
- [50].Rao RR, Long JZ, White JP, Svensson KJ, Lou J, Lokurkar I, Jedrychowski MP, Ruas JL, Wrann CD, Lo JC, et al.. Meteorin-like is a hormone that regulates immune-adipose interactions to increase beige fat thermogenesis. Cell 2014; 157:1279-91; PMID:24906147; http://dx.doi.org/ 10.1016/j.cell.2014.03.065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Rizki TM. Fat body In: Ashburner M, Wright TRF, eds. The genetics and biology of Drosophila. London: Academic Press, 1978:561-601. [Google Scholar]
- [52].Rosetto M, Engström Y, Baldari CT, Telford JL, Hultmark D. Signals from the IL-1 receptor homolog, Toll, can activate an immune response in a Drosophila hemocyte cell line. Biochem Biophys Res Commun 1995; 209:111-6; PMID:7726823; http://dx.doi.org/ 10.1006/bbrc.1995.1477 [DOI] [PubMed] [Google Scholar]
- [53].Lemaitre B, Nicolas E, Michaut L, Reichhart J-M, Hoffmann JA. The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996; 86:973-83; PMID:8808632; http://dx.doi.org/ 10.1016/S0092-8674(00)80172-5 [DOI] [PubMed] [Google Scholar]
- [54].Valanne S, Wang JH, Rämet M. The Drosophila Toll signaling pathway. J Immunol 2011; 186:649-56; PMID:21209287; http://dx.doi.org/ 10.4049/jimmunol.1002302 [DOI] [PubMed] [Google Scholar]
- [55].Shia AK, Glittenberg M, Thompson G, Weber AN, Reichhart J-M, Ligoxygakis P. Toll-dependent antimicrobial responses in Drosophila larval fat body require Spätzle secreted by haemocytes. J Cell Sci 2009; 122:4505-15; PMID:19934223; http://dx.doi.org/ 10.1242/jcs.049155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Weber AN, Tauszig-Delamasure S, Hoffmann JA, Lelievre E, Gascan H, Ray KP, Morse MA, Imler JL, Gay NJ. Binding of the Drosophila cytokine Spätzle to Toll is direct and establishes signaling. Nat Immunol 2003; 4:794-800; PMID:12872120; http://dx.doi.org/ 10.1038/ni955 [DOI] [PubMed] [Google Scholar]
- [57].Vanha-aho L-M, Valanne S, Rämet M. Cytokines in Drosophila immunity. Immunol Lett 2016; 170:42-51; PMID:26730849; http://dx.doi.org/ 10.1016/j.imlet.2015.12.005 [DOI] [PubMed] [Google Scholar]
- [58].Vanha-aho L-M, Anderl I, Vesala L, Hultmark D, Valanne S, Rämet M. Edin expression in the fat body is required in the defense against parasitic wasps in Drosophila melanogaster. PLoS Pathog 2015; 11:e1004895; PMID:25965263; http://dx.doi.org/ 10.1371/journal.ppat.1004895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Makhijani K, Alexander B, Tanaka T, Rulifson E, Brückner K. The peripheral nervous system supports blood cell homing and survival in the Drosophila larva. Development 2011; 138:5379-91; PMID:22071105; http://dx.doi.org/ 10.1242/dev.067322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Rajan A, Perrimon N. Drosophila cytokine unpaired 2 regulates physiological homeostasis by remotely controlling insulin secretion. Cell 2012; 151:123-37; PMID:23021220; http://dx.doi.org/ 10.1016/j.cell.2012.08.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Woodcock Katie J, Kierdorf K, Pouchelon Clara A, Vivancos V, Dionne Marc S, Geissmann F. Macrophage-derived upd3 cytokine causes impaired glucose homeostasis and reduced lifespan in Drosophila fed a lipid-rich diet. Immunity 2015; 42:133-44; PMID:25601202; http://dx.doi.org/ 10.1016/j.immuni.2014.12.023 [DOI] [PMC free article] [PubMed] [Google Scholar]