

ABSTRACT
Recent work has strengthened Drosophila imaginal discs as a model system for regeneration studies. Evidence is accumulating that oxidative stress drives the cellular responses for repair and regeneration. Drosophila imaginal discs generate a burst of reactive oxygen species (ROS) upon damage that is necessary for the activation of the Jun N-terminal kinase (JNK) and p38 MAP kinase signaling pathways. Moreover, these pathways are pivotal in the activation of regenerative growth. A hypothetical mechanism of how the ROS are initiated, and how repair and regeneration is activated is discussed here.
KEYWORDS: Drosophila, JNK, oxidative stress, p38, regeneration, ROS
Introduction
There is increasing interest in Drosophila imaginal discs as a model to decipher the molecular basis of cell renewal, tissue repair and regeneration. Imaginal discs follow a rigid prescribed development program, which could be highly sensitive to any disturbances. However, accumulating evidence reveals their capacity for cell renewal and the recovery of the developmental balance after severe experimental interference. This developmental homeostasis allows animals to develop quite normally, despite defective genes and hostile environments.1 Taking advantage of this capacity, imaginal discs are being used to examine the molecular basis of epithelial regeneration.
Several unsolved questions in regenerative biology are currently being addressed using Drosophila imaginal discs in combination with genetically engineered tools; for example, clarification of the nature of the overall physiological response to damage. A paradigm within which to study this issue is the delay of pupariation that occurs after damage. This delay is known to facilitate the recovery of lost tissue. Damaged discs communicate with the brain to mediate the delay through a neural network activated by the Drosophila insulin-like peptide 8 (Dilp8) that controls the balance between growth in the regenerating organ (imaginal disc) and growth in the rest of the body.2-5 Recent analysis of the transcriptome of regenerating discs has revealed the cytokine IL-6 family member unpaired (upd) to be highly elevated transcripts in damaged discs.6 While upd triggers the JAK/STAT signaling pathway and thereby promotes regenerative proliferation, dilp8 could retard pupariation thus allowing recovery of the damaged zone. Interestingly, loss of function of hop, the fly tyrosine kinase JAK, abolishes dilp8 up-regulation. In contrast, loss of function of dilp8 does not abolish regenerative proliferation. Thus, 2 roles of JAK/STAT have been reported: one controlling local cell proliferation and the other allowing regeneration by delaying development.6 An implication of these findings is that a local or a systemic response would have to be coordinated to control regenerative growth.
Another classic issue in regenerative biology is determination of whether genetic programs exist for regeneration or whether regeneration is a recapitulation of development. Regeneration implies regrowth and therefore re-specification of cell fates.7-11 However, tissues must be protected from anomalous cell fate divergence. This issue has been recently considered after isolating the gene taranis (tara) from a genetic screen; the authors found that tara protects cell fate from deleterious side effects of damage only in regeneration context.12 Another recent evidence supporting regeneration programs in imaginal discs is the discovery of a regeneration-specific wingless (wg) enhancer that functions only when regeneration is tolerated, but is epigenetically silenced when the capacity to regenerate is abolished.13,14
In addition to those findings, another key question in regenerative biology is to unveil the nature of the earliest signals that initiate repair and regeneration. Recent evidence emphasizes the role of oxidative stress in regeneration. For example, the production of reactive oxygen species (ROS) upstream in signaling pathways required for regeneration has been reported in Xenopus and zebrafish.15,16 In flies, ROS have been shown to function as a wound chemoattractant for macrophages.17,18 To understand the origin of ROS production after injury, it has been shown that the NADPH oxidase DUOX, which produces H2O2, is activated by calcium.19 However, little is known of the beneficial roles of ROS for tissue repair. For example, the JNK 8,9,11,20-27 and p38 28,29 MAPK pathways that respond to physical damage, cell death or stressors are sensitive to oxidation. Oxidative stress is powered by the production of ROS: highly reactive molecules derived from oxygen. Moderate levels of ROS can sensitize the fly haematopoietic progenitors to enter differentiation through the JNK pathway.30 Thus, the production of ROS could create a microenvironment in which the JNK and p38 pathways can be activated, leading to the onset of regeneration. A possible mechanism that would explain the ROS-dependent onset of regeneration is discussed below.
Origin of ROS
Most ROS are generated as by-products during mitochondrial electron transport or oxidation reactions. ROS result from the sequential reduction of oxygen through the addition of electrons. For example, one-electron reduction of oxygen leads to the formation of the free radical superoxide (O2−); while two-electron reduction leads to hydrogen peroxide (H2O2), which is not a free radical since all its electrons are paired. Both products are very common ROS in cellular environments.
A widely accepted rule in tissue homeostasis is that an excess of ROS is generally counteracted by ubiquitously expressed antioxidants. If the generation of ROS exceeds the capacity of the antioxidants to neutralize them, cells will undergo severe dysfunction ending in cell death. However, low concentrations of ROS may enhance cell survival and proliferation. Many molecules involved in ROS-induced cellular responses are known to be regulated by the intracellular reduction–oxidation state, which depends on the balance between the levels of oxidizing and reducing equivalents.
Wing imaginal discs damaged by physical injury or genetic activation of apoptosis share rapid production of ROS.31 Impairment of ROS production by chemical means (vitamin C, N-acetyl cysteine or trolox) or enzymatic antioxidants (Sod or Cat transgenes) can block regeneration.31 The activation of the pro-apoptotic gene reaper (rpr) results in high levels of ROS accumulation in dying cells. This has been demonstrated through the use of ROS indicators (CellROX and H2DCFDA) that are sensitive to general oxidative stress. It is important, however, to discover how and where ROS are generated. Rpr localizes in mitochondrial membranes and promotes mitochondrial fragmentation by binding to the pro-fusion Marf protein.32-34 In insects, as in mammals, alterations in mitochondria trigger the generation of ROS which oxidize apoptotic cells. Thus, it is very likely that in rpr-induced cell death, mitochondrial ROS accumulate in dying cells. In addition, apoptotic cells activate JNK and in turn JNK activates rpr,35 generating an amplification loop that not only ensures cell death but that could be responsible for the burst of mitochondrial ROS in dying cells.
Propagation of ROS to regenerating cells
ROS have also been found in living cells near dying cells. We recently found that in surviving cells, ROS are only detected with general indicators.31 This could be due to the ROS being produced by oxidases that are not mitochondrial. In Drosophila embryos, a calcium wave induced upon wounding activates DUOX, which contains a calcium binding EF-hand domain.19 Upon DUOX activation, H2O2 is produced and the activity of DUOX is propagated through the cells near the wound by the intercellular spreading of calcium through gap junctions.19 Thus, it is possible that ROS found in living cells are provided by calcium-dependent oxidases after a flash of calcium spreads from the wound to the neighboring cells. Therefore, a possible model for early damage response in discs implies that mitochondrial ROS will originate in cells in which rpr is expressed (dying cells in Fig. 1), and the intracellular environment of oxidative stress can be spread directly to nearby living cells, or indirectly by calcium flashes produced by the mechanical stress generated between the dying and healthy cells (Fig. 1). The stress response could further propagate via aquaporins 36 or gap junctions.37
Figure 1.
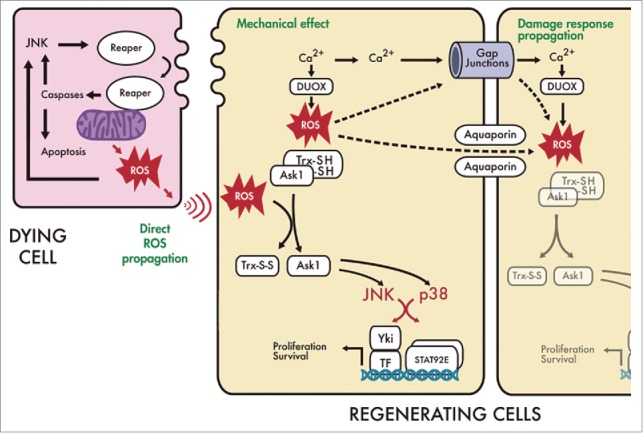
Model for ROS stimulation of regeneration. Turning on cell death machinery can induce regenerative growth in the nearby healthy cells in a ROS-dependent manner. In the dying cell Reaper inhibits the Inhibitor of Apoptosis (DIAP) which results in caspase-dependent apoptosis. Reaper/caspase act near the mitochondria,34 where ROS are released. High levels of JNK (and p53) function upstream the pro-apoptotic genes (e.g. reaper). Caspases, such as DRONC, can also activate JNK (and p53) which in turn results in reaper expression.35 This can create an amplification loop that ensures cell death. ROS in dying cells can also promote JNK activity. Three non-exclusive mechanistic scenarios can interplay to promote an early damage response in the regenerating cells: (1) Direct ROS propagation. ROS could propagate directly from dying cells to nearby cells which will induce a non-deleterious oxidative stress necessary for the damage response.31 In the regenerating cell, oxidation of the sulfhydryl groups of Trx, result in the liberation of ASK1.45 Free ASK1 acts upstream JNK and p38.45 Both p38 and JNK signals promote transcription of upd, which signals JAK/STAT to the phosphorylated dimerization of the transcription factor STAT92E.31 Yki entrance into de nucleus is JNK dependent 41 and could promote upd transcription.44 It is not known whether p38 control of upd/STAT92E is Yki dependent. Both, Yki and STAT92E promote survival and proliferation during regeneration. (2) Mechanical effect upstream of ROS. Dying cells disrupt cell membranes, adhesion and junctions, and as consequence cell membranes of the nearby healthy cells undergo mechanical stress that can originate Ca2+ flashes, as found in wounds 19 . The release of Ca2+ is necessary for DUOX to convert water and oxygen in H2O2. These ROS could release ASK1 which in turn triggers p38 and JNK. (3) Damage response propagation. Ca2+ or ROS can spread through gap junctions 37 to other healthy cells to recruit them for the regeneration response. Also, aquaporins or water channels can transfer ROS from cell-to-cell.36 This mechanism can provide a fast propagation of signals necessary to activate damage response an ultimately regenerative growth.
The role of ROS in MAPK activation
ROS are beneficial for the regeneration process and stimulate kinases that are activated by stress such as p38 and JNK.31 A mechanism that would explain this activation could be that the intracellular oxidative environment fosters the liberation of intracellular kinases upstream of the JNK and p38.38 For example, inactive MAP3K apoptosis signal-regulating kinase-1 (ASK1) tends to form a complex with thioredoxin (Trx); but under oxidative stress conditions, the sulfhydryl groups of Trx become oxidized, ASK1 is released, and p38 and JNK activity is stimulated.39
Another mechanism could be the inactivation of puckered, a MAP kinase phosphatase (MKP). There are examples of oxidation of the catalytic cysteine by ROS accumulation that lead to phosphatase inactivation, and the release of JNK.38 This mechanism, based on capacity to regulate MKP, seems to respond to ROS by protecting cells. It causes apoptosis when the levels of ROS are high, which results in blocked MKP and prolonged JNK activation. However, it protects the cell from dying when the levels of ROS are low, which results in active MKP and transient JNK activation.40
The MAP kinases JNK and p38 in regenerative growth
There is evidence of MAP kinases being involved in cytoskeletal organization, cell shape changes, cell death and proliferation. One possible role for JNK and p38 is the expression of cytokines, which in turn stimulate proliferation. As mentioned above, evidence indicates that upd cytokines are transcriptionally activated in regenerating cells and that inhibition of the JAK/STAT signaling blocks regeneration.6,25,31 Also, mutations or chemical inhibitors of JNK or p38 interfere with upd transcription and regeneration. In addition, the anomalies that result from neutralizing ROS with antioxidants or with p38 inhibitors can be reverted after ectopic expression of upd.31 Another effector required for regeneration is the Hippo pathway transcriptional co-regulator yorkie (yki), which is known to be downstream of JNK.23,41-43 Evidence points upd and yki as key genes in regenerative growth, yet ectopic expression of yki can result in upd transcription in wing discs,44 which positions yki between JNK and upd.
Thus, these findings consolidate Drosophila imaginal discs as a model for exploring epithelial plasticity, homeostasis and regeneration. Moreover, advanced fly genetic tools and the flow of information will allow current topics in regeneration biology such as epigenetic control, long non-coding RNA, micro RNA, metabolism, and systemic responses to damage, to be tested in vivo.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
I wish to thank P. Santabárbara, E. Vizcaya-Molina, I. Pardo, A. Santos, I. Martínez and M. Corominas for the discussions. I also thank J. Villarroya for helping with the artwork.
Funding
This work was supported by the Spanish Ministerio de Economía y Competitividad grant BFU2015-67623-P.
References
- [1].Alcock J. Animal Behavior: An Evolutionary Approach. 9th ed. Sunderland, MA: Sinauer Associates, Inc; 2013. 89p [Google Scholar]
- [2].Halme A, Cheng M, Hariharan IK. Retinoids regulate a developmental checkpoint for tissue regeneration in Drosophila. Curr Biol 2010; 20:458-63; PMID:20189388; http://dx.doi.org/ 10.1016/j.cub.2010.01.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Garelli A, Gontijo AM, Miguela V, Caparros E, Dominguez M. Imaginal Discs Secrete Insulin-Like Peptide 8 to Mediate Plasticity of Growth and Maturation. Science (80-) 2012; 336:579-82; http://dx.doi.org/ 10.1126/science.1216735 [DOI] [PubMed] [Google Scholar]
- [4].Vallejo DM, Juarez-Carreno S, Bolivar J, Morante J, Dominguez M. A brain circuit that synchronizes growth and maturation revealed through Dilp8 binding to Lgr3. Science (80-) 2015; 350:aac6767; http://dx.doi.org/ 10.1126/science.aac6767 [DOI] [PubMed] [Google Scholar]
- [5].Colombani J, Andersen DS, Leopold P. Secreted Peptide Dilp8 Coordinates Drosophila Tissue Growth with Developmental Timing. Science (80-) 2012; 336:582-5; http://dx.doi.org/ 10.1126/science.1216689 [DOI] [PubMed] [Google Scholar]
- [6].Katsuyama T, Comoglio F, Seimiya M, Cabuy E, Paro R. During Drosophila disc regeneration, JAK/STAT coordinates cell proliferation with Dilp8-mediated developmental delay. Proc Natl Acad Sci U S A 2015; 112:E2327-36; PMID:25902518; http://dx.doi.org/ 10.1073/pnas.1423074112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Repiso A, Bergantinos C, Corominas M, Serras F. Tissue repair and regeneration in Drosophila imaginal discs. Dev Growth Differ 2011; 53:177-85; PMID:21338344; http://dx.doi.org/ 10.1111/j.1440-169X.2010.01247.x [DOI] [PubMed] [Google Scholar]
- [8].Lee N, Maurange C, Ringrose L, Paro R. Suppression of Polycomb group proteins by JNK signalling induces transdetermination in Drosophila imaginal discs. Nature 2005; 438:234-7; PMID:16281037; http://dx.doi.org/ 10.1038/nature04120 [DOI] [PubMed] [Google Scholar]
- [9].Herrera SC, Martin R, Morata G. Tissue homeostasis in the wing disc of Drosophila melanogaster: immediate response to massive damage during development. PLoS Genet 2013; 9:e1003446; PMID:23633961; http://dx.doi.org/ 10.1371/journal.pgen.1003446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Herrera SC, Morata G. Transgressions of compartment boundaries and cell reprogramming during regeneration in Drosophila. Elife 2014; 3:e01831; PMID:24755288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Smith-Bolton RK, Worley MI, Kanda H, Hariharan IK. Regenerative growth in Drosophila imaginal discs is regulated by Wingless and Myc. Dev Cell 2009; 16:797-809; PMID:19531351; http://dx.doi.org/ 10.1016/j.devcel.2009.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Schuster KJ, Smith-Bolton RK. Taranis Protects Regenerating Tissue from Fate Changes Induced by the Wound Response in Drosophila. Dev Cell 2015; 34:119-28; PMID:26096735; http://dx.doi.org/ 10.1016/j.devcel.2015.04.017 [DOI] [PubMed] [Google Scholar]
- [13].Harris RE, Setiawan L, Saul J, Hariharan IK. Localized epigenetic silencing of a damage-activated WNT enhancer limits regeneration in mature Drosophila imaginal discs. Elife 2016; 5:pii: e11588; http://dx.doi.org/ 10.7554/eLife.11588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Schubiger M, Sustar A, Schubiger G. Regeneration and transdetermination: the role of wingless and its regulation. Dev Biol 2010; 347:315-24; PMID:20816798; http://dx.doi.org/ 10.1016/j.ydbio.2010.08.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Love NR, Chen Y, Ishibashi S, Kritsiligkou P, Lea R, Koh Y, Gallop JL, Dorey K, Amaya E. Amputation-induced reactive oxygen species are required for successful Xenopus tadpole tail regeneration. Nat Cell Biol 2013; 15:222-8; PMID:23314862; http://dx.doi.org/ 10.1038/ncb2659 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Niethammer P, Grabher C, Look AT, Mitchison TJ. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009; 459:996-9; PMID:19494811; http://dx.doi.org/ 10.1038/nature08119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Moreira S, Stramer B, Evans I, Wood W, Martin P. Prioritization of competing damage and developmental signals by migrating macrophages in the Drosophila embryo. Curr Biol 2010; 20:464-70; PMID:20188558; http://dx.doi.org/ 10.1016/j.cub.2010.01.047 [DOI] [PubMed] [Google Scholar]
- [18].Fogarty CE, Diwanji N, Lindblad JL, Tare M, Amcheslavsky A, Makhijani K, Brückner K, Fan Y, Bergmann A. Extracellular Reactive Oxygen Species Drive Apoptosis-Induced Proliferation via Drosophila Macrophages. Curr Biol 2016; 26(5):575-84; PMID:26898463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Razzell W, Evans IR, Martin P, Wood W. Calcium Flashes Orchestrate the Wound Inflammatory Response through DUOX Activation and Hydrogen Peroxide Release. Curr Biol 2013; 23:424-9; PMID:23394834; http://dx.doi.org/ 10.1016/j.cub.2013.01.058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Ryoo HD, Gorenc T, Steller H. Apoptotic cells can induce compensatory cell proliferation through the JNK and the Wingless signaling pathways. Dev Cell 2004; 7:491-501; PMID:15469838; http://dx.doi.org/ 10.1016/j.devcel.2004.08.019 [DOI] [PubMed] [Google Scholar]
- [21].Bosch M, Serras F, Martin-Blanco E, Baguna J. JNK signaling pathway required for wound healing in regenerating Drosophila wing imaginal discs. Dev Biol 2005; 280:73-86; PMID:15766749; http://dx.doi.org/ 10.1016/j.ydbio.2005.01.002 [DOI] [PubMed] [Google Scholar]
- [22].Bergantiños C, Corominas M, Serras F. Cell death-induced regeneration in wing imaginal discs requires JNK signalling. Development 2010; 137:1169-79; http://dx.doi.org/ 10.1242/dev.045559 [DOI] [PubMed] [Google Scholar]
- [23].Repiso A, Bergantinos C, Serras F. Cell fate respecification and cell division orientation drive intercalary regeneration in Drosophila wing discs. Development 2013; 140:3541-51; PMID:23903186; http://dx.doi.org/ 10.1242/dev.095760 [DOI] [PubMed] [Google Scholar]
- [24].Mattila J, Omelyanchuk L, Kyttala S, Turunen H, Nokkala S. Role of Jun N-terminal Kinase (JNK) signaling in the wound healing and regeneration of a Drosophila melanogaster wing imaginal disc. Int J Dev Biol 2005; 49:391-9; PMID:15968584; http://dx.doi.org/ 10.1387/ijdb.052006jm [DOI] [PubMed] [Google Scholar]
- [25].Pastor-Pareja JC, Wu M, Xu T. An innate immune response of blood cells to tumors and tissue damage in Drosophila. Dis Model Mech 2008; 1:144-54; PMID:19048077; http://dx.doi.org/ 10.1242/dmm.000950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Fan Y, Wang S, Hernandez J, Yenigun VB, Hertlein G, Fogarty CE, Lindblad JL, Bergmann A. Genetic models of apoptosis-induced proliferation decipher activation of JNK and identify a requirement of EGFR signaling for tissue regenerative responses in Drosophila. PLoS Genet 2014; 10:e1004131; PMID:24497843; http://dx.doi.org/ 10.1371/journal.pgen.1004131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Galko MJ, Krasnow MA. Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biol 2004; 2:E239; PMID:15269788; http://dx.doi.org/ 10.1371/journal.pbio.0020239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Craig CR, Fink JL, Yagi Y, Ip YT, Cagan RL. A Drosophila p38 orthologue is required for environmental stress responses. EMBO Rep 2004; 5:1058-63; PMID:15514678; http://dx.doi.org/ 10.1038/sj.embor.7400282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Seisenbacher G, Hafen E, Stocker H. MK2-dependent p38b signalling protects Drosophila hindgut enterocytes against JNK-induced apoptosis under chronic stress. PLoS Genet 2011; 7:e1002168; PMID:21829386; http://dx.doi.org/ 10.1371/journal.pgen.1002168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Owusu-Ansah E, Banerjee U. Reactive oxygen species prime Drosophila haematopoietic progenitors for differentiation. Nature 2009; 461:537-41; PMID:19727075; http://dx.doi.org/ 10.1038/nature08313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Santabárbara-Ruiz P, López-Santillán M, Martínez-Rodríguez I, Binagui-Casas A, Pérez L, Milán M, Corominas M, Serras F. ROS-Induced JNK and p38 Signaling Is Required for Unpaired Cytokine Activation during Drosophila Regeneration. PLoS Genet 2015; 11:e1005595; http://dx.doi.org/ 10.1371/journal.pgen.1005595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Varkey J, Chen P, Jemmerson R, Abrams JM. Altered cytochrome c display precedes apoptotic cell death in Drosophila. J Cell Biol 1999; 144:701-10; PMID:10037791; http://dx.doi.org/ 10.1083/jcb.144.4.701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Thomenius M, Freel CD, Horn S, Krieser R, Abdelwahid E, Cannon R, Balasundaram S, White K, Kornbluth S. Mitochondrial fusion is regulated by Reaper to modulate Drosophila programmed cell death. Cell Death Differ 2011; 18:1640-50; PMID:21475305; http://dx.doi.org/ 10.1038/cdd.2011.26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Clavier A, Rincheval-Arnold A, Colin J, Mignotte B, Guénal I. Apoptosis in Drosophila: which role for mitochondria? Apoptosis 2015; 21(3):239-51 [DOI] [PubMed] [Google Scholar]
- [35].Shlevkov E, Morata G. A dp53/JNK-dependant feedback amplification loop is essential for the apoptotic response to stress in Drosophila. Cell Death Differ 2012; 19:451-60; PMID:21886179; http://dx.doi.org/ 10.1038/cdd.2011.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Bienert GP, Møller ALB, Kristiansen KA, Schulz A, Møller IM, Schjoerring JK, Jahn TP. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J Biol Chem 2007; 282:1183-92; PMID:17105724; http://dx.doi.org/ 10.1074/jbc.M603761200 [DOI] [PubMed] [Google Scholar]
- [37].Feine I, Pinkas I, Salomon Y, Scherz A. Local oxidative stress expansion through endothelial cells–a key role for gap junction intercellular communication. PLoS One 2012; 7:e41633; PMID:22911831; http://dx.doi.org/ 10.1371/journal.pone.0041633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Son Y, Kim S, Chung H-T, Pae H-O. Reactive oxygen species in the activation of MAP kinases. Methods Enzymol 2013; 528:27-48; PMID:23849857; http://dx.doi.org/ 10.1016/B978-0-12-405881-1.00002-1 [DOI] [PubMed] [Google Scholar]
- [39].Tobiume K, Matsuzawa A, Takahashi T, Nishitoh H, Morita K, Takeda K, Minowa O, Miyazono K, Noda T, Ichijo H. ASK1 is required for sustained activations of JNK/p38 MAP kinases and apoptosis. EMBO Rep 2001; 2:222-8; PMID:11266364; http://dx.doi.org/ 10.1093/embo-reports/kve046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Ilmarinen P, Moilanen E, Kankaanranta H. Mitochondria in the center of human eosinophil apoptosis and survival. Int J Mol Sci 2014; 15:3952-69; PMID:24603536; http://dx.doi.org/ 10.3390/ijms15033952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Sun G, Irvine KD. Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev Biol 2011; 350:139-51; PMID:21145886; http://dx.doi.org/ 10.1016/j.ydbio.2010.11.036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Grusche FA, Degoutin JL, Richardson HE, Harvey KF. The Salvador/Warts/Hippo pathway controls regenerative tissue growth in Drosophila melanogaster. Dev Biol 2011; 350:255-66; PMID:21111727; http://dx.doi.org/ 10.1016/j.ydbio.2010.11.020 [DOI] [PubMed] [Google Scholar]
- [43].Meserve JH, Duronio RJ. Scalloped and Yorkie are required for cell cycle re-entry of quiescent cells after tissue damage. Development 2015; 142(16):2740-51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Shaw RL, Kohlmaier A, Polesello C, Veelken C, Edgar BA, Tapon N. The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 2010; 137:4147-58; PMID:21068063; http://dx.doi.org/ 10.1242/dev.052506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Shiizaki S, Naguro I, Ichijo H. Activation mechanisms of ASK1 in response to various stresses and its significance in intracellular signaling. Adv Biol Regul 2013; 53:135-44; PMID:23031789; http://dx.doi.org/ 10.1016/j.jbior.2012.09.006 [DOI] [PubMed] [Google Scholar]