

ABSTRACT
Based on their sequence homology to Arabidopsis NDR1 and tobacco (Nicotiana tabacum) HIN1, 45 NHL (NDR1/HIN1-like) family genes are found in Arabidopsis genome. Recently, we reported that overexpression of NHL6, a member of NHL family, modulated seed germination under abiotic stresses through affecting ABA biosynthesis and signaling. We also carried out qPCR and investigated the expression of the other 8 member genes (NHL7a, 16, 17, 21, 25, 26, 41, 43) whose transcriptional data are publicly unavailable, and found that expression of NHL17 was induced more than 2 folds in ABA treated seedlings. Furthermore, in addition to the plasma membrane localization, YFP-NHL6 fusion protein was also observed in the cytosol (as dots) or on the membrane of small vacuoles or vesicles. As a member of the pathogen infection related genes, expression of NHL6 was significantly induced by salicylic acid and NHL6s are evolutionarily conserved among different plant species. A working model of NHL6 in ABA response was proposed.
KEYWORDS: ABA, abiotic stress, gene expression, NHL family, NHL6
Abbreviations
- ABA
abscisic acid
- SA
salicylic acid
- NDR1
nonrace-specific disease resistance 1
- HIN1
harpin-induced 1
- NHL
NDR1/HIN1-like
- LEA
late embryogenesis abundant
- PYR/PYL
pyrabactin resistance/PYR like
- RCAR
regulatory component of ABA receptors
- qPCR
quantitative polymerase chain reaction
- PR1
pathogenesis-related protein 1
- SnRK2
SNF1-related protein kinase 2
- PP2C
protein phosphatase 2C
- ABRE
ABA-reponsive elements
- AREB
ABRE-binding
- ABF
ABRE-binding factor
- NCED
9-cis-epoxycarotenoid dioxygenase
- AAO
ABA aldehyde oxidase
During their whole life cycles, plants are threatened by different kinds of biotic or abiotic stresses. Transcriptional and post transcriptional regulations including gene expression and physiological dynamics are utilized to counteract these unfavorable environmental factors.1-3 The importance of plant hormone abscisic acid (ABA) in stress response and seed germination has been broadly studied,4-6 as advanced by the discovery of its receptors PYR/PRL or RCARs.7,8 Recently, we showed that a conventional pathogen infection related NHL (NDR1/HIN1-like) family gene NHL6 modified Arabidopsis seed germination and early seedling growth under abiotic stress conditions, through altering ABA biosynthesis and sensitivity in transgenic plants.9 Interestingly, when we examined the transcriptional expression of the 45 NHL family genes in senescent leaves, a good tissue for mimicking dehydration, NHL6 was found to be the most highly expressed and induced by ABA at the same time. Characteristics of NHL6 in dehydration and ABA response convinced us to study further the function of NHL6, and its roles in ABA-mediated seed germination and early seedling growth.
Based on the data published by Goda and co-workers,10 4 of the 45 Arabidopsis NHL genes, NHL1, NHL4, NHL6 and NHL39 were induced to more than 2 folds by ABA. However, no data are available for the rest 8 NHL genes: NHL7a, NHL16, NHL17, NHL21, NHL25, NHL26, NHL41 and NHL43 in regard to ABA induced response. To better and comprehensively understand their roles in response to ABA, qPCR was performed using 7-day-old seedlings.11 NHL17 was found to be significantly induced by ABA (Table 1). Bioinformatic analyses indicated that a conserved LEA-14 like domain exists in most of the ABA-responsive NHL members,12 with transmembrane domains of each protein except NHL29 (Table 1).13,14 Furthermore, LEA proteins are closely correlated with land plants' ability to acquire dehydration tolerance.15 Our results as well as the published microarray data of NHL family genes will add more evidence of this family in ABA and abiotic stress responses.
Table 1.
Characteristics of selected NHL members and their ABA induced expression.
| Locus | Name | ABAa | CDS (bp) | Length (aa) | MW (kD) | TMsb | Interproc |
|---|---|---|---|---|---|---|---|
| At1g54540 | NHL4 | 14.26 | 720 | 239 | 26.38 | 1 | LEA-14 |
| At1g17550 | NHL29 | 5.82 | 1536 | 511 | 56.06 | 0 | PP2C-like |
| At1g65690 | NHL6 | 3.61 | 759 | 252 | 28.57 | 1 | LEA-14 |
| AT3G11660 | NHL1 | 2.72 | 630 | 209 | 23.66 | 1 | LEA-14 |
| At3g54200 | NHL39 | 2.68 | 708 | 235 | 25.78 | 1 | LEA-14 |
| At3g44220 | NHL17 | 2.5 | 621 | 206 | 23.25 | 1 | LEA-14 |
| At5g56050 | NHL43 | 1.18 | 852 | 283 | 32.26 | 1 | Low complexity |
| At5g36970 | NHL25 | 1.18 | 747 | 248 | 27.96 | 1 | LEA-14 |
| At3g24600 | NHL41 | 0.91 | 1251 | 506 | 55.18 | 3 | LEA-14 |
| At3g20610 | NHL16 | 0.9 | 669 | 222 | 25.73 | 2 | Low complexity |
| At5g53730 | NHL26 | 0.51 | 642 | 213 | 23.91 | 1 | LEA-14 |
| At1g08140 | NHL7a | 0.16 | 2457 | 818 | 93.39 | 10 | Cation/H+ exchanger |
| At4g05220 | NHL21 | 0.07 | 681 | 226 | 26.04 | 1 | LEA-14 |
Dates are acquired from TAIR. (a) Expression of NHL family genes in ABA treated seedlings by Goda et al., 2008. Rows in gray background were generated from our qPCR result. (b) Number of TMs (transmembrane domains) predicted by TMHMM. (c) Conserved domains in NHL proteins are predicted by Inetrpro (https://www.ebi.ac.uk/interpro/). Primers used for qPCR: NHL7a (forward: 5′-AGCTGCCTACACCCTCTTGA-3′; reverse: 5′-GGGAATCTTGTGATGGGCTA-3′);
NHL16 (forward: 5′-GACACGAGAAGAAGGCCAAG-3′; reverse: 5′-CCCATCCTCTTTGTCCTTGA-3′);
NHL17 (forward: 5′-ATCTTACCGCAACCAACAGG-3′; reverse: 5′-CACGTTCCCACTTTCCATCT-3′);
NHL21 (forward: 5′-CTAAACCCGTTTTTCCAGCA-3′; reverse: 5′-GATCAACCCGTCTCTTCCAA-3′);
NHL25 (forward: 5′-TCCGAGGGAGAAGAAAGGAT-3′; reverse: 5′-TTTCTCGTTGGGATTTTTCG-3′);
NHL26 (forward: 5′-GTTGCCCAGTCCTTTGGTTA-3′; reverse: 5′-TGTTCATTCCAAAAGCCACA-3′);
NHL41 (forward: 5′-TATGTATTGGCCGGAGAAGC-3′; reverse: 5′-AACGCTCTTAACGGAGACGA-3′);
NHL43 (forward: 5′-GAAACACCACCATGGGAAAC-3′; reverse: 5′-TCCGGCTTGTTCTTAGCTGT-3′).
Many ABA signal components such as receptor-like kinase GHR1 (guard cell hydrogen peroxide-resistant 1), calcium sensor CBL9 (calcineurin B-Like 9) and GPCR (G-protein coupled receptor) type G-protein are plasma membrane localized.17-19 In addition to its co-localization with the plasma membrane tethered AtTLP family protein AtTLP11,9,16 YFP-NHL6 fusion proteins were also observed in the cytosol and on the membrane of small vacuole or vesicles, when transiently expressed in tobacco leaf epidermal cells (Fig. 1A). This suggests that NHL6 may modulate ABA biosynthesis and signaling on the plasma membrane, or probably through protein trafficking and sorting.20 The self interaction of NHL6 also implies that NHL6 might hetero-dimerize with other NHL family proteins, as supported by the observation that NHLs are highly conserved.21,22 Therefore, it would be interesting to study the relationship between NHL6 and other NHL family members in Arabidopsis seed germination or ABA response. Biochemical and genetic assays will help us to figure out whether they are working counteractively or collaboratively.
Figure 1.
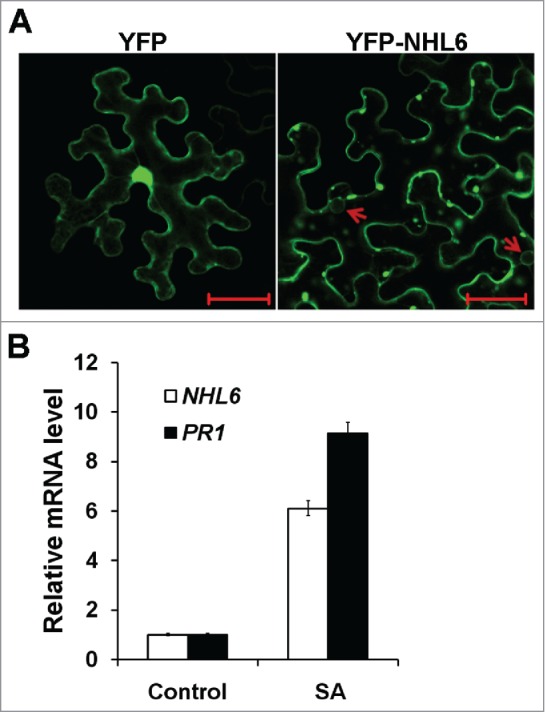
Subcellular localization of YFP-NHL6 and salicylic acid induced expression of NHL6. (A) Subcellular localization of YFP-NHL6 in tobacco leaf epidermal cells. Two small vesicles are indicated by red arrows. Empty YFP vector is used as a control. Bar = 50 μm. (B) cDNA derived from the rosette leaves of 4-week-old Arabidopsis plants treated with 100 µM SA for 12 hours was used as template. qPCR primers for NHL6: forward primer-5′ AACCGTACAATTGTCCACACTCAGG 3′; reverse primer-5′ TTCAGCTCCTTGTCGGTGGTGTT 3′. PR1 (Forward primer-5′AGGCACGAGGAGCGGTAGG3′; reverse primer-5′CTTTGGCACATCCGAGTCTCAC3′) and ACTIN2 (Forward primer-5′GGAAGGATCTGTACGGTAAC3′; reverse primer-5′GGACCTGCCTCATCATACT3′) were employed as positive and native controls, respectively.
Besides the crucial role in abiotic stress response, ABA is essential for plant resistance to pathogens.23-26 As a member of the pathogen related NHL family genes,21,27,28 NHL6 might function in disease response, too. We examined the expression level of NHL6 in the rosette leaves of wild type Arabidopsis plants and found that, same as PR1, NHL6 was also highly induced by SA (salicylic acid) (Fig. 1B). Since NHL6 could trigger ABA biosynthesis under abiotic stress condition,9 it is tempting to speculate that NHL6 may also play a role in disease response, probably through regulating ABA biosynthesis and/or signaling. Conserved motifs in homologous proteins of Arabidopsis NHL6 are discovered among different plant species (Fig. 2), implying the functional conservation of NHL6 in ABA and SA response.
Figure 2.

Protein sequence alignment of PtNHL6 (poplar, XP_002318799.1), VvNHL6 (Vitis, XP_002263732.1), MtNHL6 (Medicago, XP_003616105.1), AtNHL6 (Arabidopsis, NP_003022.3), ZmNHL6 (maize, DAA49515.1) and OsNHL6 (rice, NP_001054263.1). The conserved NHL motifs such as motif 1 (LILWLILRPXKPKFXVQDATV), motif2 (NPNKRIGIYYD) and motif 3 (PFYQGHKN) are indicated in black lines above the sequences.
A proposed working model of NHL6 in ABA and stress response is drawn based on our current research (Fig. 3). Further study on the possible target(s) that interact(s) with NHL6 will offer more information for the understanding of its molecular function(s) in biotic and abiotic stresses.
Figure 3.

A proposed working model of NHL6. ABA biosynthesis is induced by salt and mannitol. Upon the binding of ABA to its co-receptors PYR/PRL/RCARs-PP2Cs, the phospatase activity of PP2Cs is inhibited. When the kinase activity of SnRK2s is released from PP2Cs, they phosphorylate and activate AREBs and ABF3. Then, activated AREBs and ABF3 bind to the cis-element ABRE in the promoter of NHL6 and induce its transcription. The internalization of homo-dimerized NHL6 from plasma membrane to the cytosol may also be triggered by salt, mannitol and ABA, to induce endogenous ABA accumulation through up-regulating the expression of ABA biosynthesis genes NCED3, ABA3, AAO2 and AAO3.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This work has been supported by the following grants: the National Natural Science Foundation of China (31371228, 31370670); the National Mega Project of GMO Crops (2014ZX0800942B); and the Strategic Priority Research Program of the Chinese Academy of Sciences XDA08030108.
References
- 1.Zhu JK. Salt and drought stress signal transduction in plants. Annu Rev Plant Physiol 2002; 53:247-73; http://dx.doi.org/ 10.1146/annurev.arplant.53.091401.143329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jones JD, Dangl JL. The plant immune system. Nature 2006; 444:323-9; PMID:17108957; http://dx.doi.org/ 10.1038/nature05286 [DOI] [PubMed] [Google Scholar]
- 3.Yamaguchi-Shinozaki K, Shinozaki K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Physiol 2006; 57:781-803; http://dx.doi.org/ 10.1146/annurev.arplant.57.032905.105444 [DOI] [PubMed] [Google Scholar]
- 4.Finkelstein RR, Gampala SSL, Rock CD. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002; 14:S15-S45; PMID:12045268; http;//dx.doi.org/http://dx.doi.org/ 10.1105/tpc.010441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nambara E, Marion-Poll A. Abscisic acid biosynthesis and catabolism. Annu Rev Plant Physiol 2005; 56:165-85; http://dx.doi.org/ 10.1146/annurev.arplant.56.032604.144046 [DOI] [PubMed] [Google Scholar]
- 6.Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR. Abscisic acid: emergence of a core signaling network. Annu Rev Plant Physiol 2010; 61:651-79; http;//dx.doi.org/ 10.1146/annurev-arplant-042809-112122 [DOI] [PubMed] [Google Scholar]
- 7.Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009; 324:1064-8; PMID:19407143; http://dx.doi.org/ 10.1126/science.1172408 [DOI] [PubMed] [Google Scholar]
- 8.Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, et al.. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009; 324:1068-71; PMID:19407142; http;//dx.doi.org/ 10.1126/science.1173041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bao Y, Song WM, Pan J, Jiang CM, Srivastava R, Li B, Zhu LY, Su HY, Gao XS, Liu H, et al.. Overexpression of the NDR1/HIN1-like gene NHL6 modifies seed germination in response to abscisic acid and abiotic stresses in Arabidopsis. PLoS One 2016; 11:e0148572; http://dx.doi.org/ 10.1371/journal.pone.0148572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Goda H, Sasaki E, Akiyama K, Maruyama-Nakashita A, Nakabayashi K, Li W, Ogawa M, Yamauchi Y, Preston J, Aoki K, et al.. The AtGenExpress hormone and chemical treatment data set: experimental design, data evaluation, model data analysis and data access. Plant J 2008; 55:526-42; PMID:18419781; http://dx.doi.org/ 10.1111/j.1365-313X.2008.03510.x [DOI] [PubMed] [Google Scholar]
- 11.Bao Y, Wang CT, Jiang CM, Pan J, Zhang GB, Liu H, Zhang HX. The tumor necrosis factor receptor-associated factor (TRAF)-like family protein SEVEN IN ABSENTIA 2 (SINA2) promotes drought tolerance in an ABA-dependent manner in Arabidopsis. New Phytol 2014; 202:174-87; PMID:24350984; http://dx.doi.org/ 10.1111/nph.12644 [DOI] [PubMed] [Google Scholar]
- 12.Mitchell A, Chang HY, Daugherty L, Fraser M, Hunter S, Lopez R, McAnulla C, McMenamin C, Nuka G, Pesseat S, et al.. The InterPro protein families database: the classification resource after 15 years. Nucleic Acids Res 2015; 43:D213-D221; http://dx.doi.org/ 10.1093/nar/gku1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol 2001; 305:567-80; PMID:11152613; http://dx.doi.org/ 10.1006/jmbi.2000.4315 [DOI] [PubMed] [Google Scholar]
- 14.Hand SC, Menze MA, Toner M, Boswell L, Moore D. LEA proteins during water stress: not just for plants anymore. Annu Rev Physiol 2011; 73:115-34; PMID:21034219; http://dx.doi.org/ 10.1146/annurev-physiol-012110-142203 [DOI] [PubMed] [Google Scholar]
- 15.Sonnhammer EL, von Heijne G, Krogh A. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc Int Conf Intell Syst Mol Biol 1998; 6:175-82; PMID:9783223 [PubMed] [Google Scholar]
- 16.Bao Y, Song WM, Jin YL, Jiang CM, Yang Y, Li B, Huang WJ, Liu H, Zhang HX. Characterization of Arabidopsis Tubby-like proteins and redundant function of AtTLP3 and AtTLP9 in plant response to ABA and osmotic stress. Plant Mol Biol 2014; 86:471-83; PMID:25168737; http://dx.doi.org/ 10.1007/s11103-014-0241-6 [DOI] [PubMed] [Google Scholar]
- 17.Hua DP, Wang C, He JN, Liao H, Duan Y, Zhu ZQ, Guo Y, Chen ZZ, Gong ZZ. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell 2012; 24:2546-61; PMID:22730405; http://dx.doi.org/ 10.1105/tpc.112.100107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pandey GK, Cheong YH, Kim KN, Grant JJ, Li LG, Hung W, D′Angelo C, Weinl S, Kudla J, Luan S. The calcium sensor calcineurin B-Like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004; 16:1912-24; PMID:15208400; http://dx.doi.org/ 10.1105/tpc.021311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pandey S, Nelson DC, Assmann SM. Two novel GPCR-type G proteins are abscisic acid receptors in Arabidopsis. Cell 2009; 136:136-48; PMID:19135895; http://dx.doi.org/ 10.1016/j.cell.2008.12.026 [DOI] [PubMed] [Google Scholar]
- 20.Vilaine F, Kerchev P, Clement G, Batailler B, Cayla T, Bill L, Gissot L, Dinant S. Increased expression of a phloem membrane protein encoded by NHL26 alters phloem export and sugar partitioning in Arabidopsis. Plant Cell 2013; 25:1689-708; PMID:23715470; http://dx.doi.org/ 10.1105/tpc.113.111849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dörmanna P, Gopalan S, He SY, Benning C. A gene family in Arabidopsis thaliana with sequence similarity to NDR1 and HIN1. Plant Physiol Biochem 2000; 38:789-96; http://dx.doi.org/ 10.1016/S0981-9428(00)01186-4 [DOI] [Google Scholar]
- 22.Zheng MS, Takahashi H, Miyazaki A, Hamamoto H, Shah J, Yamaguchi I, Kusano T. Up-regulation of Arabidopsis thaliana NHL10 in the hypersensitive response to Cucumber mosaic virus infection and in senescing leaves is controlled by signalling pathways that differ in salicylate involvement. Planta 2004; 218:740-50; PMID:14666423; http://dx.doi.org/ 10.1007/s00425-003-1169-2 [DOI] [PubMed] [Google Scholar]
- 23.Mauch-Mani B, Mauch F. The role of abscisic acid in plant-pathogen interactions. Curr Opin Plant Biol 2005; 8:409-14; PMID:15939661; http://dx.doi.org/ 10.1016/j.pbi.2005.05.015 [DOI] [PubMed] [Google Scholar]
- 24.Adie BAT, Perez-Perez J, Perez-Perez MM, Godoy M, Sanchez-Serrano JJ, Schmelz EA, Solano R. ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 2007; 19:1665-81; PMID:17513501; http://dx.doi.org/ 10.1105/tpc.106.048041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fan J, Hill L, Crooks C, Doerner P, Lamb C. Abscisic acid has a key role in modulating diverse plant-pathogen interactions. Plant Physiol 2009; 150:1750-61; PMID:19571312; http://dx.doi.org/ 10.1104/pp.109.137943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ton J, Flors V, Mauch-Mani B. The multifaceted role of ABA in disease resistance. Trends Plant Sci 2009; 14:310-7; PMID:19443266; http://dx.doi.org/ 10.1016/j.tplants.2009.03.006 [DOI] [PubMed] [Google Scholar]
- 27.Varet A, Parker J, Tornero P, Nass N, Nurnberger T, Dangl JL, Scheel D, Lee J. NHL25 and NHL3, two NDR1/HIN1-Like genes in Arabidopsis thaliana with potential role(s) in plant defense. Mol Plant Microbe Interact 2002; 15:608-16; PMID:12059109; http://dx.doi.org/ 10.1094/MPMI.2002.15.6.608 [DOI] [PubMed] [Google Scholar]
- 28.Varet A, Hause B, Hause G, Scheel D, Lee J. The Arabidopsis NHL3 gene encodes a plasma membrane protein and its overexpression correlates with increased resistance to Pseudomonas syringae pv. tomato DC3000. Plant Physiol 2003; 132:2023-33; PMID:12913158; http://dx.doi.org/ 10.1104/pp.103.020438 [DOI] [PMC free article] [PubMed] [Google Scholar]