

Abstract
Children from different socioeconomic backgrounds have differing abilities to delay gratification, and impoverished children have the greatest difficulties in doing so. In the present study, we examined the role of vagal tone in predicting the ability to delay gratification in both resource-rich and resource-poor environments. We derived hypotheses from evolutionary models of children’s conditional adaptation to proximal rearing contexts. In Study 1, we tested whether elevated vagal tone was associated with shorter delay of gratification in impoverished children. In Study 2, we compared the relative role of vagal tone across two groups of children, one that had experienced greater impoverishment and one that was relatively middle-class. Results indicated that in resource-rich environments, higher vagal tone was associated with longer delay of gratification. In contrast, high vagal tone in children living in resource-poor environments was associated with reduced delay of gratification. We interpret the results with an eye to evolutionary-developmental models of the function of children’s stress-response system and adaptive behavior across varying contexts of economic risk.
Keywords: delay of gratification, stress reactions, poverty, evolutionary psychology
To wait, or not to wait: That is the question. For more than 50 years, psychologists have put this query to young children in laboratory tasks in which they are asked to sit patiently and are promised a larger reward if they can resist the immediate temptation of a smaller reward (Mischel, Shoda, & Rodriguez, 1989). And this has been a fruitful line of research as studies have documented that children’s ability to suppress the dominant response to eat the treat sitting in front of them is associated with a wide range of positive outcomes, including socioemotional competence, prosocial behavior, and reduced psychopathology (e.g., Eisenberg et al., 2004; Mischel et al., 1989; Razza & Raymond, 2013). Thus, normative psychological models propose that longer delay of gratification reflects greater regulation and that this ability to exercise self-control is the optimal outcome given its association with socioemotional adjustment (e.g., Calkins, 2009). However, research has documented the presence of wide socioeconomic disparities in children’s ability to delay gratification; impoverished children display greater difficulties compared with their more affluent counterparts (e.g., Evans & English, 2002; Raver et al., 2011). This raises a provocative question: Why would children living in heightened economic uncertainty show a greater propensity toward taking the immediate reward if that decision is maladaptive with respect to adjustment?
Perhaps part of the answer lies in the tendency for researchers to interpret behavior using theories and models of developmental psychology that are based on benchmarks derived largely from children living within relatively secure environments. Thus, the meaning and function of behavior across broad contexts is missed (e.g., Cicchetti & Lynch, 1993; Henrich, Heine, & Norenzayan, 2010). Evolutionary-developmental models offer an alternative conceptual framework, positing that natural selection has shaped people’s ability to modify regulatory behaviors so that they better match local environmental conditions (e.g., Daly & Wilson, 2005; Ellis, Figueredo, Brumbach, & Schlomer, 2009). This framework suggests that
when people encounter stressful environments, this does not so much disturb their development as direct or regulate it toward strategies that are adaptive under stressful conditions, even if those strategies are currently harmful in terms of the long-term welfare of the individual or society as a whole. (Ellis, Boyce, Belsky, Bakermans-Kranenburg, & Van IJzendoorn, 2011, p. 8)
Consequently, different environments should elicit different strategies depending on the perception of present-future trade-offs (Belsky, Steinberg, & Draper, 1991; Chisholm, 1999). In environments in which resources and support are plentiful, the ability to control immediate impulses and desires in the service of regulation is more than likely to result in greater rewards and benefits. In contrast, early rearing contexts consisting of heightened impoverishment, diminished access to resources, and harsher living conditions reduce the likelihood of a future payoff. Under these circumstances, evolutionary frameworks suggest that children should lawfully shift preferences toward immediate rewards (Fawcett, McNamara, & Houston, 2012).
However, this line of reasoning does not explain how such a shift occurs, and attention has turned toward understanding the antecedents of children’s ability to delay gratification, particularly in contexts of risk (e.g., Lengua, Zalewski, Fisher, & Moran, 2013). Cardiac vagal tone is an early-life biological marker that has been implicated in models of children’s self-regulation. Vagal tone is typically assessed by measuring resting levels of respiratory sinus arrhythmia (RSA), which captures parasympathetic dominance over cardiac functioning in the autonomic nervous system (e.g., Beauchaine, 2001). In particular, the polyvagal theory (Porges & Furman, 2011) proposes that vagal tone represents a physiological index of an individual’s capacity to respond quickly and flexibly to environmental demands as well as to regulate more effectively from heightened arousal. Higher levels of basal vagal tone are thought to reflect heightened sensitivity to environmental cues and the enactment of behaviors that are tailored to local conditions. In contrast, lower levels of basal vagal tone are purported to indicate difficulties in attending to or reacting to environmental contexts (or both), resulting in more ineffective and less malleable behavioral coping strategies (Brosschot & Thayer, 1998). Thus, developmental models of self-regulation consider vagal tone an important factor in understanding children’s effortful control.
Empirical work examining associations between RSA and children’s regulation ability supports the suggestion that RSA may operate as an antecedent of delay-control ability in affective tasks (Beauchaine, 2001; Calkins, 2009). In accordance with RSA’s proposed function of supporting appropriate engagement with the environment, research has shown that high basal RSA is associated with sustained attention (Suess, Porges, & Plude, 1994), effortful control (Taylor, Eisenberg, & Spinrad, 2015), and executive function (Marcovitch et al., 2010). However, much of the previous work in this area has been conducted in samples of children who hail primarily from middle-class and higher socioeconomic backgrounds. A small group of studies using primarily low-income and impoverished children has found associations between high vagal tone and poor regulation (e.g., Blair & Peters, 2003; Davies, Sturge-Apple, Cicchetti, Manning, & Zale, 2009; Kidwell & Barnett, 2007). If the physiological function of vagal tone is to facilitate the prioritization of behaviors in response to environmental cues in a manner that benefits the individual, these results may not necessarily be contradictory. In conditions characterized by elevated resources and support, high vagal tone may result in delayed gratification with an eye toward a larger future reward. However, under conditions of reduced resources, high vagal tone may support the adoption of quick reward-oriented strategies. Thus, both of these strategies may be adaptive within their given contexts.
In summary, we investigated the role of basal vagal tone, assessed during the toddler years, in association with children’s decision to delay gratification across resource-rich and resource-poor environmental contexts. In the first study, we used a sample of children living in highly impoverished conditions to test the association of basal vagal tone with delay of gratification. We hypothesized that higher vagal tone would predict shorter delay of gratification in the context of poverty, consistent with an evolutionary-developmental framework. In the second study, we sought both to replicate findings obtained in Study 1 and to replicate findings within the larger literature by testing how environmental contexts moderated the role of vagal tone in predicting children’s delay ability. To maximize comparability, we made certain that the procedures were similar across studies and were approved by the institutional review board at the research site.
Study 1
Participants
Two hundred one 2-year-old children (44% female) and their mothers from a moderately sized metropolitan area in the northeastern United States were the participants. They were recruited through agencies that serve disadvantaged children and families, including Women, Infants, and Children and rosters from Temporary Assistance to Needy Families, administered through the local Department of Human Services. We wanted to obtain a sample of 200 mothers, and recruitment ended when we achieved this goal. Participants’ median annual household income was $18,300 (U.S.), and a substantial minority of mothers (30%) had not completed high school. Most families were receiving public assistance (95%) and were living below the U.S. federal poverty level (99.5%). Fifty-six percent of participants were Black, 23% were White, 11% were Latino, 7% were multiracial, and 3% were “other.” The cumulative retention rate across the two measurement occasions was 87%. To test for selective attrition, we conducted statistical comparisons between the mother-child dyads that participated in each wave and the dyads that dropped out during the longitudinal component of the study along the primary, covariate, and demographic variables at the first assessment (e.g., family income, maternal education). No significant differences between these groups were identified in the analyses.
Procedure
Data were collected when mothers and toddlers visited the laboratory; visits occurred when the children were 2 and 4 years old.
Heart-monitor procedure
Basal heart rate data were collected on each child at the first measurement wave using a monitor (MiniLogger 2010 Series; Mini Mitter, Inc., Bend, OR). The MiniLogger detects each electrocardiographic (ECG) R-wave and records every interval between successive R-waves to the nearest millisecond. Mothers held their children on their laps while a trained experimenter placed two electrodes on the child’s chest (one medially on the right collar bone and one on the child’s left side below the rib cage). Consistent with prior research, children were given low-arousal activities during recording to keep them calm and facilitate baseline data collection (Hastings et al., 2008).
Delay of gratification
At the second wave of data collection (i.e., when the children were 4 years old), each child was placed at a small table furnished with two plates and a bell. The experimenter placed two M&Ms on one plate and five M&Ms on the other plate. Children were told how to ring the bell. Then the experimenter pointed out the difference in the number of M&Ms on each plate and told the child that if he or she could wait until the experimenter returned, they would receive the five pieces of candy. If they couldn’t wait, they were to ring the bell to signal the experimenter to return, and then they could eat the two pieces of candy. The experimenter then left the room for a 10-min waiting period.
Measures
Respiratory sinus arrhythmia
RSA is the fluctuation in heart rate associated with respiration. Interbeat-interval data were extracted from the ECG assessment using Mini-Log 2000W software (Mini Mitter, Inc.), edited for artifacts, and analyzed using CMet software (Allen, Chambers, & Towers, 2007). Specifically, the CMetX program converted the interbeat-interval series into a time series via linear interpolation at a sampling rate of 10 Hz. Then, the variance in IBI within the frequency band of 0.24 to 1.04 Hz (i.e., the band of spontaneous respiration for toddlers; Porges, Doussard-Roosevelt, Portales, & Greenspan, 1996) was extracted via a band-pass filter. The natural log of this variance was used as an estimate of children’s average RSA for the baseline session.
Delay of gratification
Children’s ability to delay gratification was operationalized as the length of time children waited to eat the M&M during the delay-of-gratification task (Mischel & Ebbesen, 1970). If a child touched the M&Ms or rang the bell, his or her score was from the number of minutes and seconds before that point. If a child waited the entire time, his or her score was 10 min. In the present sample, only 16% of children waited the entire 10 min.
Results
For descriptive purposes, Table 1 provides the means, standard deviations, and intercorrelations among the measures of main constructs. A path model examining the association between children’s RSA at 2 years of age and delay of gratification at 4 years of age was conducted using Mplus software (Version 7.0; Muthén & Muthén, 2012). To maximize our sample size, we used the full-information, maximum-likelihood (FIML) estimation procedure available in Mplus (e.g., Enders, 2001). This method is appropriate when no identifiable pattern exists in the missing data and the amount of missing data is as high as 50% (Schlomer, Bauman, & Card, 2010). An analysis of missing data showed that the data were missing completely at random (MCAR), χ2(6, N = 175) = 0.850, n.s. We included two demographic covariates, maternal education and income, to maximize our ability to compare results in our two studies, given the socioeconomic constructs used to create the two groups in Study 2. Child age at the time of delay of gratification was also covaried. We examined the data for the presence of outliers and found four participants whose family income was more than 3 standard deviations from the mean. We removed these participants and reran the analyses to determine whether their data were influential; results were identical to those with the participants’ data included. Given the similarity in findings, we elected to retain these cases in our analyses. Finally, delay of gratification evidenced moderate skew. We transformed the variable using a logarithmic function to reduce skew, and analyses were run with the transformed variable.
Table 1.
Means, Standard Deviations, and Intercorrelations Among the Main Variables in Studies 1 and 2
| Variable | M | SD | Correlations |
|||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||
| Study 1 | ||||||
| 1. Respiratory sinus arrhythmia (log Hz) | 6.62 | 1.36 | — | |||
| 2. Delay-of-gratification time (seconds) | 140 | 225 | −.27* | — | ||
| 3. Child’s age (years) | 2.14 | 0.14 | .24* | .10 | — | |
| 4. Maternal educationa | 2.84 | 1.68 | −.21* | .32* | −.08 | — |
| 5. Family income (thousands of dollars) | 20.82 | 12.27 | −.17* | .24* | −.07 | .36* |
| Study 2 | ||||||
| 1. Respiratory sinus arrhythmia (log Hz) | 5.91 | 2.18 | — | |||
| 2. Delay-of-gratification time (seconds) | 463 | 210 | .03 | — | ||
| 3. Child’s age (years) | 5.29 | 0.24 | −.12 | .14 | — | |
Maternal education was coded as follows: 0 = none, 1 = elementary/junior high school, 2 = high-school-equivalency test, 3 = high school diploma, 4 = vocational/tech diploma or 1 year of college, 5 = associate’s degree, 6 = diploma in nursing, 7 = bachelor’s degree, 8 = master’s degree, 9 = doctoral degree.
p < .05.
The path model used to test associations (Fig. 1) between basal RSA activity and delay of gratification was fully identified, χ2(0, N = 201). There was a significant effect of basal RSA at age 2 on delay of gratification at age 4, β = −0.23, SE = 0.02, z = −2.82, p < .01, in the context of other proximal demographic factors entered as simultaneous predictors in the model. Our findings suggest that early-occurring elevation in basal RSA activity in the context of poverty was associated with shorter delay of gratification at the age of 4.
Fig. 1.

Path model from Study 1. This model shows the degree to which children’s basal vagal tone at age 2 predicted their ability to delay gratification at age 4. Solid lines and values with asterisks indicate significant effects (*p < .05). Dashed lines indicate nonsignificant effects. All possible correlational pathways between exogenous predictor variables were included when estimating the model, but these pathways are not shown in the figure. RSA = respiratory sinus arrhythmia.
Study 2
Participants
Data were drawn from a longitudinal study of mother-child relationships. The sample consisted of 140 mother-child pairs (48.4% female) recruited when the children were 18 months old. Participants were recruited from the metropolitan area of a midsized city in the northeastern United States. Mother-toddler dyads fell into one of two groups on the basis of socioeconomic status (SES): low SES (n = 69) and high SES (n = 71). For study enrollment, mothers in the high-SES group were required to have completed a bachelor’s degree at an accredited 4-year college or university and to have an income not supplemented through government assistance (e.g., food stamps). Mothers in the low-SES group were required to have not completed a bachelor’s degree and to be receiving some form of public assistance (verified through Department of Human Services records). The goal was to obtain two groups of 70 mothers each, and recruitment was ended when we had obtained these samples. Median family income of all participants was $46,093 per year; however, one third of the sample reported earnings below $10,200 per year. Mothers were between 18 and 42 years of age (mean age = 24 years). Participants identified themselves as White (55% of mothers and 49% of children), Black (26% of mothers and 25% of children), Latina/Latino (11% of mothers and 12% of children), biracial (5% of mothers and 14% of children), Asian (< 1% of mothers, 0% of children), and Native American/Alaskan (< 1% of mothers, 0% of children). Retention rate from Wave 1 to Wave 2 was 74%.
Procedure
Mothers and their children visited the lab when the children were 18 months old (Wave 1) and again when the children were 5 years old (Wave 2). The children completed the delay-of-gratification task and the mothers filled out surveys at each visit.
Heart-monitor procedure
At Wave 1, heart monitors from Alive Technologies (Ashmore, Queensland, Australia) were used to record and store the children’s ECG signals during a silent baseline session. The portable monitors received signals through a precordial, two-pole ECG lead that was placed on children’s chests, and data were stored on a flash memory card inside the monitor, which was worn by the child. The ECG signal was sampled at 300 Hz and had a voltage range of −2.5 to 2.5 V.
Delay of gratification
At Wave 2, children’s delay of gratification was assessed using a delay task similar to that used in Study 1. However, children chose between 3 and 8 M&Ms.
Measures
Socioeconomic diversity
At recruitment, participants for the current study were identified as either low SES or high SES. Group membership was used as a measure of socioeconomic diversity. Mothers in the low-SES group reported an average family income of $23,518 per year (range = $20,625–$26,410) and reported an average of 2.5 children in the household (range = 1–11). In contrast, mothers in the high-SES group reported an average income of $85,727 (range = $77,709–$287,000) and reported an average of 1.8 children in the household (range = 1–4). We also used the mothers’ addresses to examine neighborhood statistics. Compared with mother-child dyads in the high-SES group, mother-child dyads in the low-SES group resided in areas demarcated by higher population density and higher rates of violent crime (http://www.city-data.com/city/Rochester-New-York.html). Thus, mothers and children in our low-SES group were also characterized by their experience of heightened adversity within their immediate residential environment (Table 2).
Table 2.
Ecological Conditions of Participants in Study 2: Means and Standard Deviations
| Variable | Full sample (n = 140) |
Low-SES group (n = 71) |
High-SES group (n = 69) |
|||
|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |
| Population density (1,000s of people per square mile) |
1.87 | 1.08 | 2.56 | 0.64 | 1.08 | 0.94 |
| Violent crimes (per 100,000 people) |
6.27 | 3.25 | 8.31 | 1.79 | 3.90 | 2.95 |
Note: SES = socioeconomic status.
Respiratory sinus arrhythmia
As in Study 1, RSA was calculated using CMet software (Allen et al., 2007).
Delay of gratification
Children’s ability to delay gratification was scored as in Study 1; higher scores indicated longer delay.
Results
Table 1 shows means, standard deviations, and intercorrelations for the main variables in Study 2. There were significant differences in delay ability between the two groups, χ2(1, N = 104) = 3.89, p < .05. In results consistent with the larger literature, children in the low-SES group were less likely to delay gratification (58%) than were the children in the high-SES group (41%). Data were checked for outliers, and none were found. As in Study 1, delay of gratification evidenced moderate skew. We transformed the variable using a logarithmic function and used the transformed variable in analyses.
Before running analyses, predictor variables were centered to avoid problems with multicollinearity (Aiken & West, 1991). Interaction terms were created by multiplying the centered predictor variables. We tested our hypothesized model using path analysis as in Study 1. To evaluate whether data were missing completely at random, we examined the patterns of missing data using Little’s MCAR test (Little, 1988). Results showed that the data were missing completely at random, χ2(1, N = 140) = 29.19, n.s.
Model analyses were run in accordance with our study aims. All paths were estimated, and thus the model was fully saturated. As in previous research, there was a significant effect of SES group on children’s delay of gratification, β = 0.31, SE = 0.09, p < .05, such that children in the high-SES group had longer delay time. There was no significant main effect of RSA, β = 0.07, SE = 0.02, p = .56; however, the interaction between RSA and SES had a significant effect on children’s delay of gratification, β = 0.46, SE = 0.04, p < .001. The interaction term explained 18% of the variance in children’s delay of gratification. Given the difficulties inherent in detecting moderators in field research (r2 values are typically less than .03; McClelland & Judd, 1993), the magnitude of the effect in the current study is noteworthy.
To probe this effect, we conducted simple-slopes analyses with SES group as the moderator variable using an online utilities program (Preacher, Curran, & Bauer, 2006; http://www.quantpsy.org/interact/mlr2.htm). Results showed that the simple slope for children in the low-SES group was negative and significantly different from zero, β = −0.08, t(135) = 2.31, p < .05, whereas the simple slope for children in the high-SES group was positive and significantly different from zero, β = 0.10, t(135) = 3.02, p < .05 (Fig. 2). Children’s basal RSA was plotted at 1 standard deviation above and below the mean (Fig. 2). Our findings indicate that higher basal RSA in the context of poverty is associated with shorter delay of gratification, whereas higher basal RSA in the context of more resource-rich rearing environments is associated with longer delay of gratification.
Fig. 2.
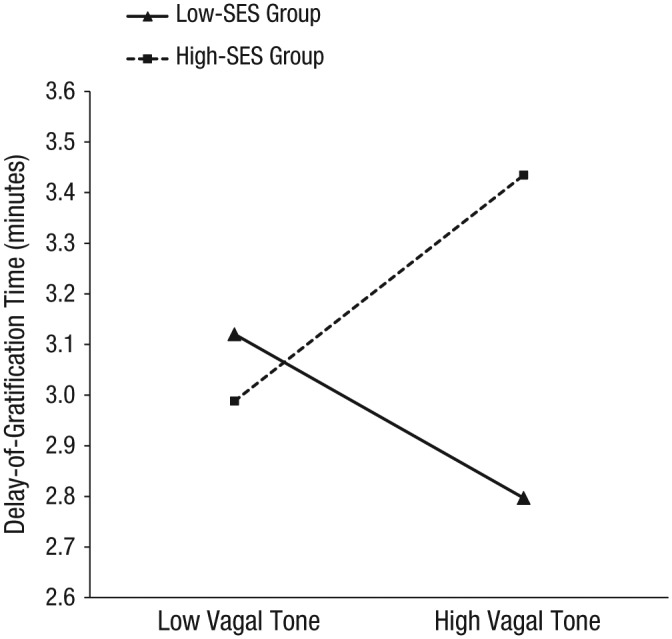
Results from the simple-slopes analysis in Study 2. Delay-of-gratification time at age 5 is plotted as a function of basal vagal tone at the age of 18 months. High and low basal vagal tone refer to values 1 SD above and below the mean, respectively. SES = socioeconomic status.
Discussion
Psychological models of self-regulation highlight the importance of resisting impulses to facilitate health and well-being (Calkins, 2009). Empirical research has identified vagal tone as a potential early physiological precursor of children’s ability to self-regulate; however, findings have been equivocal to date. In the current study, a first foray into testing the predictive value of vagal tone for children’s delay control across environmental conditions, our results across two studies indicate that context matters. Specifically, we found that for children in high-resource environments, higher levels of vagal tone were associated with delay of gratification in a predictable fashion according to normative models of regulation. However, in resource-poor environments, higher basal vagal tone predicted a lower propensity to delay gratification.
Taken together, the present findings do not readily support conceptualizations drawn from normative psychological models proposing that high vagal tone is a marker of context-independent developmentally appropriate regulation. Instead, the current findings suggest that the function of high vagal tone may operate in a more curvilinear fashion toward facilitating behavioral fit within specific environmental contexts (Boyce & Ellis, 2005; West-Eberhard, 2003). In other words, high vagal tone may function differently within specific contexts, rather than having a context-independent function. This dichotomy naturally prompts the question: How does context inform our understanding of what is “adaptive” with respect to how vagal tone is associated with the regulation of behavior? Although our conclusions are speculative, we believe that these inconsistencies dovetail nicely with emerging evolutionary-developmental theories that stress placing development within its proximal ecology when interpreting children’s developmental outcomes (Belsky & Pluess, 2013). These frameworks assert that humans adapt their behavior in ways that attempt to increase fitness in a specific local environmental condition. In a local context in which obtaining future rewards is stochastic and uncertain, taking the immediate treat may make perfect sense for children, even though this is viewed as maladaptive by society as a whole. In support of this assertion, experimental research showing a positive association between heightened mortality cues and reward-oriented decision making has attributed this link to greater orientation to the present in the context of heightened adversity (Griskevicius, Tybur, Delton, & Robertson, 2011).
By extension, our results also raise the possibility that the way in which the function of vagal tone is interpreted with respect to children’s regulatory behaviors, such as delay of gratification, may depend on the proximal conditions. In impoverished contexts, in which access to resources is unstable, higher basal vagal tone, by virtue of its sensitivity to contextual cues, may function to support development through directing behavioral strategies that value immediate reward as opposed to long-term gain. In contrast, long-term gains may be more likely in environments with higher resources and greater predictability, and thus high vagal tone may support deferring the immediate sweets because of the context-driven (or informed) expectancy that a larger cache is waiting. Thus, across groups, higher vagal tone functions to promote the behavioral strategy that is more adaptive to the specific context.
Several limitations must be acknowledged in interpreting our results. First, the current study used a single assessment of children’s vagal tone and delay of gratification. Thus, we were unable to model potential transactions across these two domains of regulation as well as developmental change in these constructs over time; to test this, it would be important for future work to include a broader range of assessments of these constructs. Second, although the focus was to examine basal vagal tone, our sampling precludes examination of vagal reactivity and regulation in response to stress. It is clear from previous work that both basal vagal tone and reactive vagal tone are important to consider in relation to behavioral outcomes. Third, given the homogeneity with respect to SES in Study 1, we were unable to test how different contextual conditions within the context of poverty (e.g., differences in maternal education or income) may further explain the findings presented. Finally, the results of the current study are developmentally specific to the early childhood period and may not generalize to older children and adults.
Against a backdrop of previous empirical research examining the regulatory role of vagal tone in shaping human development, our interpretation of vagal tone as a potential sensitivity factor in determining self-regulation requires confirmatory evidence from future studies. In addition, delineating proximal psychological processes that operate as critical mediating mechanisms underlying this association should be an important focus of future research. Although we were unable to do so here, it would be interesting to test for potential switch points (i.e., points at which the association between vagal tone and regulation reverses) in the gradient of SES. This would be particularly informative for interventions targeting chronically marginalized and impoverished children.
These limitations notwithstanding, the present study highlights the potential explanatory utility of integrating evolutionary-developmental models within traditional developmental research (Davies, Sturge-Apple, & Cicchetti, 2011; Sturge-Apple, Davies, Martin, Cicchetti, & Hentges, 2012) and within clinical interventions (Thibodeau, August, Cicchetti, & Symons, 2016). We contend that viewing development within an evolutionary framework naturally shifts the nature of empirical questions from a value-laden approach focused primarily on the form of a behavior (inherently good or inherently bad) toward understanding the proximate function of children’s behavior when considered within their rearing context. We believe this is particularly true for interpreting behavior in highly stressful and impoverished conditions. In such contexts, children’s development may actually be elegantly honed to match their environmental constraints; thus, of necessity, their behavior may not cohere to normative models of child development.
Footnotes
Action Editor: Ian H. Gotlib served as action editor for this article.
Declaration of Conflicting Interests: The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding: This research was supported by National Institute of Nursing Research Grant R21-NR010857 (to F. A. Rogosch and M. L. Sturge-Apple) and National Institute of Mental Health Grant R01-MH071256 (to P. T. Davies and D. Cicchetti).
References
- Aiken L. S., West S. G. (1991). Multiple regression: Testing and interpreting interactions. Thousand Oaks, CA: Sage. [Google Scholar]
- Allen J. J., Chambers A. S., Towers D. N. (2007). The many metrics of cardiac chronotropy: A pragmatic primer and a brief comparison of metrics. Biological Psychology, 74, 243–262. [DOI] [PubMed] [Google Scholar]
- Beauchaine T. (2001). Vagal tone, development, and Gray’s motivational theory: Toward an integrated model of autonomic nervous system functioning in psychopathology. Development and Psychopathology, 13, 183–214. [DOI] [PubMed] [Google Scholar]
- Belsky J., Pluess M. (2013). Beyond risk, resilience, and dysregulation: Phenotypic plasticity and human development. Development and Psychopathology, 25, 1243–1261. [DOI] [PubMed] [Google Scholar]
- Belsky J., Steinberg L., Draper P. (1991). Childhood experience, interpersonal development, and reproductive strategy: An evolutionary theory of socialization. Child Development, 62, 647–670. [DOI] [PubMed] [Google Scholar]
- Blair C., Peters R. (2003). Physiological and neurocognitive correlates of adaptive behavior in preschool among children in Head Start. Developmental Neuropsychology, 24, 479–497. [DOI] [PubMed] [Google Scholar]
- Boyce W. T., Ellis B. J. (2005). Biological sensitivity to context: I. An evolutionary–developmental theory of the origins and functions of stress reactivity. Development and Psychopathology, 17, 271–301. [DOI] [PubMed] [Google Scholar]
- Brosschot J. F., Thayer J. F. (1998). Anger inhibition, cardiovascular recovery, and vagal function: A model of the link between hostility and cardiovascular disease. Annals of Behavioral Medicine, 20, 326–332. [DOI] [PubMed] [Google Scholar]
- Calkins S. D. (2009). Regulatory competence and early disruptive behavior problems: The role of physiological regulation. In Olson S. L., Sameroff A. J. (Eds.), Biopsychosocial regulatory processes in the development of childhood behavioral problems (pp. 86–115). New York, NY: Cambridge University Press. [Google Scholar]
- Chisholm J. S. (1999). Death, hope and sex: Steps to an evolutionary ecology of mind and morality. New York, NY: Cambridge University Press. [Google Scholar]
- Cicchetti D., Lynch M. (1993). Toward an ecological/transactional model of community violence and child maltreatment: Consequences for children’s development. Psychiatry, 56, 96–118. [DOI] [PubMed] [Google Scholar]
- Daly M., Wilson M. (2005). Carpe diem: Adaptation and devaluing the future. The Quarterly Review of Biology, 80, 55–60. [DOI] [PubMed] [Google Scholar]
- Davies P. T., Sturge-Apple M. L., Cicchetti D. (2011). Interparental aggression and children’s adrenocortical reactivity: Testing an evolutionary model of allostatic load. Development and Psychopathology, 23, 801–814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies P. T., Sturge-Apple M. L., Cicchetti D., Manning L. G., Zale E. (2009). Children’s patterns of emotional reactivity to conflict as explanatory mechanisms in links between interpartner aggression and child physiological functioning. Journal of Child Psychology and Psychiatry, 50, 1384–1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberg N., Spinrad T. L., Fabes R. A., Reiser M., Cumberland A., Shepard S. A., . . . Thompson M. (2004). The relations of effortful control and impulsivity to children’s resiliency and adjustment. Child Development, 75, 25–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis B. J., Boyce W. T., Belsky J., Bakermans-Kranenburg M. J., Van IJzendoorn M. H. (2011). Differential susceptibility to the environment: An evolutionary–neurodevelopmental theory. Development and Psychopathology, 23, 7–28. [DOI] [PubMed] [Google Scholar]
- Ellis B. J., Figueredo A. J., Brumbach B. H., Schlomer G. L. (2009). Fundamental dimensions of environmental risk. Human Nature, 20, 204–268. [DOI] [PubMed] [Google Scholar]
- Enders C. K. (2001). The impact of nonnormality on full information maximum-likelihood estimation for structural equation models with missing data. Psychological Methods, 6, 352–370. [PubMed] [Google Scholar]
- Evans G. W., English K. (2002). The environment of poverty: Multiple stressor exposure, psychophysiological stress, and socioemotional adjustment. Child Development, 73, 1238–1248. [DOI] [PubMed] [Google Scholar]
- Fawcett T. W., McNamara J. M., Houston A. I. (2012). When is it adaptive to be patient? A general framework for evaluating delayed rewards. Behavioural Processes, 89, 128–136. [DOI] [PubMed] [Google Scholar]
- Griskevicius V., Tybur J. M., Delton A. W., Robertson T. E. (2011). The influence of mortality and socioeconomic status on risk and delayed rewards: A life history theory approach. Journal of Personality and Social Psychology, 100, 1015–1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hastings P. D., Nuselovici J. N., Utendale W. T., Coutya J., McShane K. E., Sullivan C. (2008). Applying the polyvagal theory to children’s emotion regulation: Social context, socialization, and adjustment. Biological Psychology, 79, 299–306. [DOI] [PubMed] [Google Scholar]
- Henrich J., Heine S. J., Norenzayan A. (2010). Most people are not WEIRD. Nature, 466, 29. [DOI] [PubMed] [Google Scholar]
- Kidwell S. L., Barnett D. (2007). Adaptive emotion regulation among low-income African American children. Merrill-Palmer Quarterly, 53, 155–183. [Google Scholar]
- Lengua L. J., Zalewski M., Fisher P., Moran L. (2013). Does HPA-axis dysregulation account for the effects of income on effortful control and adjustment in preschool children? Infant and Child Development, 22, 439–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Little R. J. A. (1988). A test of missing completely at random for multivariate data with missing values. Journal of the American Statistical Association, 83, 1198–1202. [Google Scholar]
- Marcovitch S., Leigh J., Calkins S. D., Leerks E. M., O’Brien M., Blankson A. N. (2010). Moderate vagal withdrawal in 3.5-year-old children is associated with optimal performance on executive function tasks. Developmental Psychobiology, 52, 603–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClelland G. H., Judd C. M. (1993). Statistical difficulties of detecting interactions and moderator effects. Psychological Bulletin, 114, 376–390. [DOI] [PubMed] [Google Scholar]
- Mischel W., Ebbesen E. B. (1970). Attention in delay of gratification. Journal of Personality and Social Psychology, 16, 329–337. [DOI] [PubMed] [Google Scholar]
- Mischel W., Shoda Y., Rodriguez M. I. (1989). Delay of gratification in children. Science, 244, 933–938. [DOI] [PubMed] [Google Scholar]
- Muthén L. K., Muthén B. O. (2012). Mplus user’s guide (7th ed.). Los Angeles, CA: Author. [Google Scholar]
- Porges S. W., Doussard-Roosevelt J. A., Portales A. L., Greenspan S. I. (1996). Infant regulation of the vagal “brake” predicts child behavior problems: A psychobiological model of social behavior. Developmental Psychobiology, 29, 697–712. [DOI] [PubMed] [Google Scholar]
- Porges S. W., Furman S. A. (2011). The early development of the autonomic nervous system provides a neural platform for social behaviour: A polyvagal perspective. Infant and Child Development, 20, 106–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preacher K. J., Curran P. J., Bauer D. J. (2006). Computational tools for probing interactions in multiple linear regression, multilevel modeling, and latent curve analysis. Journal of Educational and Behavioral Statistics, 31, 437–448. [Google Scholar]
- Raver C. C., Jones S. M., Li-Grining C., Zhai F., Bub K., Pressler E. (2011). CSRP’s impact on low-income preschoolers’ preacademic skills: Self-regulation as a mediating mechanism. Child Development, 82, 362–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Razza R. A., Raymond K. (2013). Associations among maternal behavior, delay of gratification, and school readiness across the early childhood years. Social Development, 22, 180–196. [Google Scholar]
- Schlomer G. L., Bauman S., Card N. A. (2010). Best practices for missing data management in counseling psychology. Journal of Counseling Psychology, 57, 1–10. [DOI] [PubMed] [Google Scholar]
- Sturge-Apple M. L., Davies P. T., Martin M. J., Cicchetti D., Hentges R. F. (2012). An examination of the impact of harsh parenting contexts on children’s adaptation within an evolutionary framework. Developmental Psychology, 48, 791–805. [DOI] [PubMed] [Google Scholar]
- Suess P. E., Porges S. W., Plude D. J. (1994). Cardiac vagal tone and sustained attention in school-age children. Psychophysiology, 31, 17–22. [DOI] [PubMed] [Google Scholar]
- Taylor Z. E., Eisenberg N., Spinrad T. L. (2015). Respiratory sinus arrhythmia, effortful control, and parenting as predictors of children’s sympathy across early childhood. Developmental Psychology, 51, 17–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thibodeau E. L., August G. J., Cicchetti D., Symons F. J. (2016). Application of environmental sensitivity theories in personalized prevention for youth substance abuse: A transdisciplinary translational perspective. Translational Behavioral Medicine, 6, 81–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West-Eberhard M. J. (2003). Developmental plasticity and evolution. New York, NY: Oxford University Press. [Google Scholar]