

Dear Editor,
In eukaryotic cells, the endoplasmic reticulum (ER) is a continuous membrane system involved in many critical cellular processes, including protein synthesis, lipid synthesis, and calcium storage. Morphologically, the ER is composed of cistern-like sheet structures and a reticular network of tubules. Classes of integral membrane proteins that shape the ER have been identified: the reticulons and DP1/Yop1p proteins generate ER tubules by inducing high curvature in the membrane (Voeltz et al., 2006), the atlastin GTPases and Sey1p/RHD3 proteins mediate fusion of ER membranes, forming a tubular network (Hu et al., 2009; Orso et al., 2009), and Climp63, Kinectin, and p180 play a role in stabilizing ER sheets (Shibata et al., 2010). Mutations in the determinants of ER tubules cause growth defects, short root hairs, and a neurodegenerative disease called hereditary spastic paraplegia (HSP) (Hu et al., 2009), and sheet-formation proteins are strongly upregulated in professional secretory cells when ER sheet expansion is needed, suggesting that ER morphology is tightly associated with its physiological functions (Shibata et al., 2010).
ER morphology has rarely been studied in protozoan parasites, likely due to difficulties posed by the lack of genetic manipulation and relatively small scale of these cells. In Entamoeba histolytica, the intestinal protozoan that causes invasive amebiasis, the ER was first thought to be composed of vesicles of varying size but recently shown to be a continuous network (Teixeira and Huston, 2008). In Toxoplasma gondii, the infectious agent resulting in toxoplasmosis, expansion and partitioning of the ER has been followed during the cell cycle (Nishi et al., 2008). The ER in Plasmodium parasites, the causative agents of malaria, has been visualized during the erythrocytic cycle (van Dooren et al., 2005); it transforms from a perinuclear structure with no distinctive morphological characters into a reticular network throughout the cytoplasm. Plasmodium develops first in the Anopheles mosquito, and then invades the liver upon injection into mammalian hosts. Whether the ER adopts its characteristic shapes in these stages is yet to be determined, and the molecular determinants of the ER in Plasmodium parasites remain unclear.
To identify Plasmodium falciparum orthologs of ER-shaping proteins, we conducted Blast searches of the P. falciparum genomic database (PlasmoDB, www.plasmoDB.org, v. 6.3, released December 22, 2009) using Saccharomyces cerevisiae Yop1p (ScYop1p) and human DP1 protein sequences as queries. Both searches revealed 35%–42% identity (55%–60% homology) with PFC0730w (Gene ID PF3D7_0316700 in PlasmoDB v.26, released October 15, 2015). The PFC0730w sequence was further used to query P. berghei proteins using the tblastn tool. This search returned PBANKA_0414500 and PBANKA_1135000 as the closest P. berghei homologs (Fig. S1). PBANKA_0414500 was recently annotated as a putative HVA22/TB2/DP1 family protein and PBANKA_1135000 as a HVA22-like protein. Because HVA22 is the Yop1p homolog in plants and TB2 is a previously used alias for DP1, our homologous searches confirm the annotation of the genomic database. We renamed PBANKA_0414500 as PbYOP1 and PBANKA_1135000 as PbYOP1L (Fig. 1A and 1B). When searching for the reticulon homolog in the P. berghei genome, we found PBANKA_1139900 and renamed it PbRTN1 (Fig. 1B). According to PlasmoDB database, the three potential Plasmodium tubule-forming proteins have distinct expression profiles during the asexual cycle: the expression of PbYOP1 gradually increases and peaks in trophozoites and schizonts, the levels of PbYOP1L peak in rings and trophozoites but decrease in schizonts, and levels of PbRTN1 are relatively constant. These data suggest non-redundant functions of these proteins.
Figure 1.
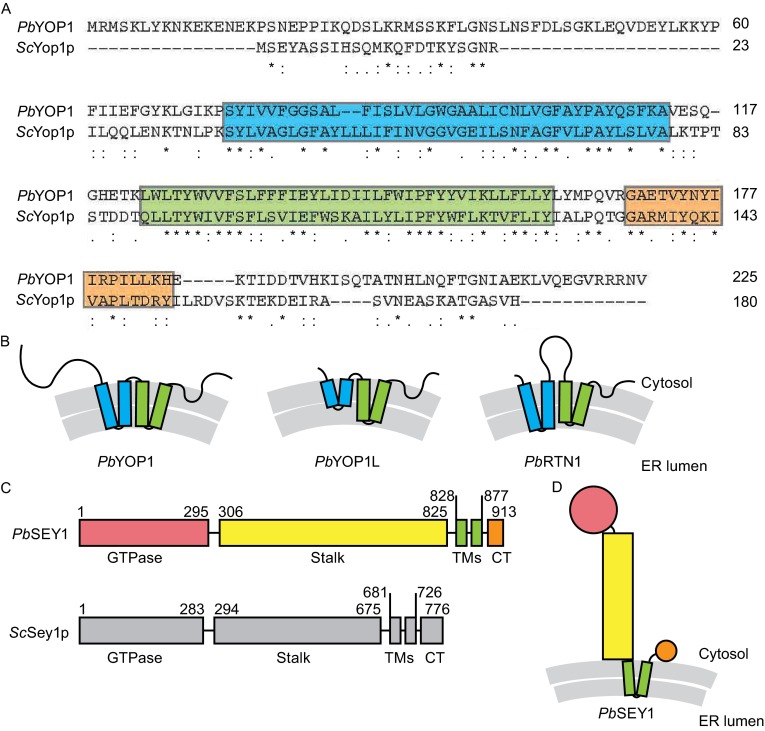
Identification of Plasmodium ER-shaping proteins. (A) Sequence alignment of P. berghei YOP1 (PbYOP1) and S. cerevisiae Yop1p (ScYop1p). Residues are numbered; identical residues are indicated by asterisks and similar residues by dots. The first transmembrane segment is highlighted in cyan, the second in green, and the predicted C-terminal amphipathic helix in orange. (B) Topology diagrams of three ER tubule-forming proteins identified in the P. berghei genome. Lipid bilayers are shown in gray, the first transmembrane segment in cyan, and the second one in green. The length of each transmembrane segment and loop is shown in proportion to the predicted length. (C) Domain diagrams of P. berghei SEY1 (PbSEY1) and S. cerevisiae Sey1p (ScSey1p). Domain boundaries are labeled. TM, transmembrane segment; CT, C-terminal tail. (D) Topology diagram of PbSEY1. The domains are colored as in (C)
Thus, three potential ER tubule-forming proteins were identified in P. berghei. A common feature of ER tubule-forming proteins is a reticulon-homology domain (RHD) consisting of two tandem transmembrane hairpins (TMH) (Fig. 1B). The sequences of all three candidates for ER tubule formation in P. berghei exhibited characteristics of the RHD domain. Notably, the intervening loop between the two TMHs in PbRTN1 is much longer than that of Yop1 family proteins, including PbYOP1 and PbYOP1L (Fig. 1B). In contrast, PbYOP1 bears a much longer N-terminus than PbYOP1L and PbRTN1. The length of the TMHs is also variable, with the first TMH of PbYOP1L and the second TMH of PbRTN1 being shorter than other TMHs (Figs. 1B and S2), suggesting that it partially traverses the lipid bilayer. Importantly, the primary structure of PbYOP1 is highly conserved among Plasmodium species (Fig. S2), implying a fundamental role of the protein.
We performed a similar search for the homolog of Sey1p in the P. berghei genome and identified PBANKA_1026600 as PbSEY1 (Fig. 1C and 1D). Four conserved motifs were identified in the GTPase domains of the dynamin superfamily: the P-loop (G1, β1–α1), the switch 1 region (G2, α1–β2), the switch 2 region (G3, β3–α2), and the G4 motif (after β5). Most residues in these signature motifs are identical between ScSey1p and PbSEY1 (Fig. S3). The predicted stalk domain of PbSEY1, which is between the GTPase domain and the transmembrane domains, is ~150 amino acids longer than that of ScSey1p and ~400 amino acids longer than those found in ATLs. The length of the stalk domain of ScSey1p has been shown to be essential for fusion (Yan et al., 2015) and to possibly contribute to the unique GTP cycle of ScSey1p when compared to ATL. If this region forms a helical bundle as predicted, it would be taller than that of ScSey1p, implying likely different dynamics during membrane fusion reactions. Taken together, the ER-shaping proteins identified in P. berghei may function similarly to those of yeast and mammalian orthologs and may possess unique features.
Purified ScYop1p, when reconstituted into proteoliposomes, leads to the generation of membrane tubules in vitro, indicating that Yop1p is sufficient to induce high curvature in membranes (Hu et al., 2008). To test whether PbYOP1 is capable of bending membranes as demonstrated for ScYop1p, we performed in vitro reconstitution assays. Recombinant PbYOP1 (residues 39–186) lacks the predicted flexible region in the N- and C-termini for optimized expression and stability. PbYOP1 was expressed and purified from Escherichia coli (Fig. 2A), mixed with preformed liposomes, and detergents were removed by the addition of bio-beads. When the reconstituted products were visualized by transmission electron microscopy (TEM), tubular structures similar to those of ScYop1p and ScRtn1p were observed (Fig. 2B). The diameters of these tubules are relatively uniform (15–20 nm), which is consistent with the ScYop1p tubules. In the absence of PbYOP1, liposomes exhibited a characteristic round shape (Fig. 2B). These results suggest that PbYOP1 alone is adequate to induce high curvature in membranes and cause tubule formation upon reconstitution with lipids.
Figure 2.
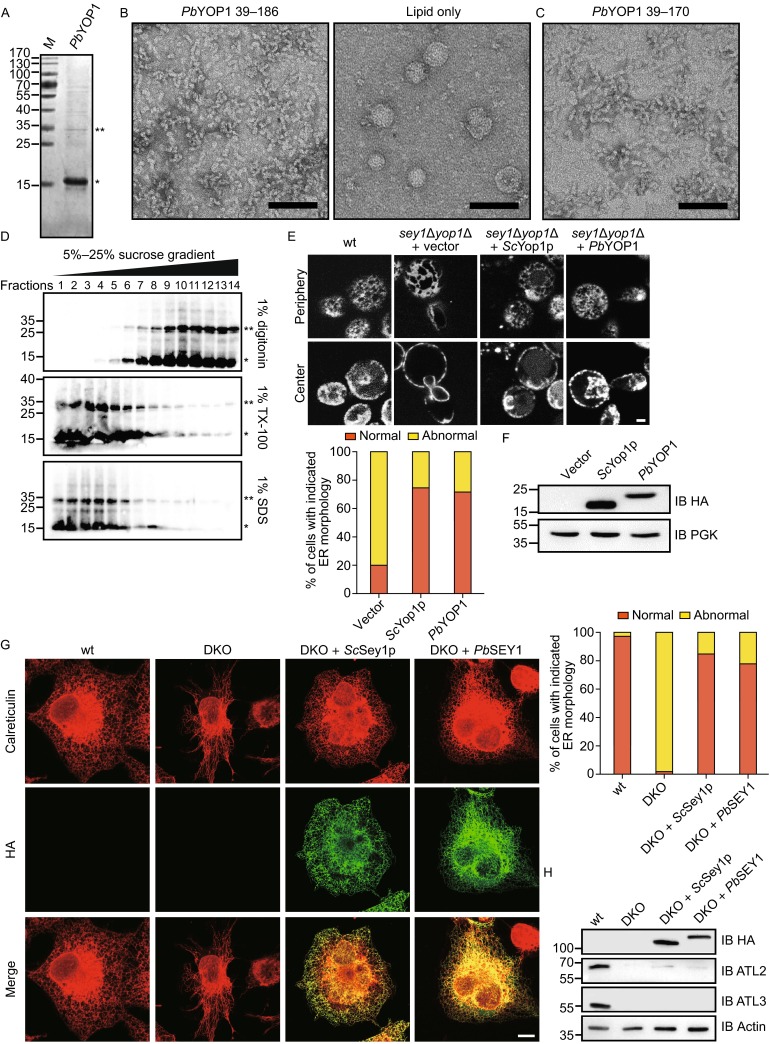
Functional tests of PbYOP1 and PbSEY1. (A) Purified PbYOP1-HA (residues 39–186) was analyzed by SDS-PAGE and coomassie staining. SDS-resistant dimers are indicated by asterisks. Molecular marker is shown in kDa. (B) PbYOP1-HA was mixed with E. coli polar lipids in Foscholine-12 and the detergent removed with Bio-beads over 4 h. The proteoliposomes were analyzed by negative stain EM. The right panel shows the sample with lipid only. Scale bar = 200 nm. (C) As in (B), but with PbYOP1 lacking the C-terminal amphipathic helix (residues 39–170). (D) Sucrose gradient centrifugation of reconstituted PbYOP1 treated with the indicated detergents. PbYOP1-HA in 1% digitonin (top panel), 1% Triton X-100 (middle panel), or 1% SDS (bottom panel) was loaded onto a sucrose step gradient. After centrifugation for 2 h at 25°C, fractions were analyzed by SDS-PAGE and immunoblotting with HA antibodies. Detergent-resistant dimers are indicated by asterisks. (E) A RFP targeted to the ER lumen (ss-RFP-HDEL) was expressed in wild type (wt) or sey1Δ yop1Δ yeast cells. The localization of the protein was determined by fluorescence microscopy focusing at the center or periphery of the cells. Empty vector or indicated SEY1s were also expressed. The ER morphology was determined by counting at least 100 cells for each sample. The results are representative of at least three repetitions. Scale bar = 2 μm. (F) Indicated SEY1s expressed in sey1Δ yop1Δ cells were determined by anti-HA immunoblotting. PGK was used as a loading control. (G) ATLs-deleted COS-7 cells (DKO) were transfected with HA-tagged SEY1s. The ER morphology of indicated cells was visualized using calreticulin, an endogenous luminal ER protein, with indirect immunofluorescence and confocal microscope, and categorized as “normal” or “abnormal”. A total of 80–100 cells were counted for each sample. All graphs are representative of three repetitions. Scale bar = 10 μm. (H) ATL2, ATL3, and SEY1s levels were determined by immunoblotting. Actin was used as a loading control
The two TMHs of the RHD domain are thought to occupy more space in the outer leaflet than the inner leaflet of the lipid bilayer, deforming membranes as a wedge insertion. A conserved amphipathic helix C-terminal to the second TMH of ScYop1p was recently identified to provide additional wedging (Brady et al., 2015). Deletion of the helix in ScYop1p abolished its tubule-forming ability in vitro. Through secondary structure prediction and helical wheel analysis, we found two potential helices that exhibit an amphipathic nature, one on each side of the RHD domain of PbYOP1. To test the role of these helices, we synthesized a peptide corresponding to the sequences of the C-terminal helix (Fig. S4A, residues 171–184) and determined its membrane association as expected for other membrane-deforming amphipathic helices. Circular dichroism spectrum analysis revealed that the peptide only adopted an α-helical configuration when liposomes were present (Fig. S4B), indicating stabilization of the peptide in a membrane environment. To our surprise, mutant PbYop1 (residues 39–170) missing the entire C-terminus, including the predicted amphipathic helix, could still form tubules in vitro (Fig. 2C). Therefore, we investigated the role of the N-terminal helix. We found that deletion of the N-terminal region of PbYOP1 did not yield any usable recombinant protein (Fig. S4C). Instead, we obtained a major contaminant after attempting to purify truncated PbYOP1 (residues 73–186). When the contaminant underwent the same reconstitution process, no tubular structures were formed (Fig. S4D), further confirming that the tubules we observed could only be specifically generated by functional PbYOP1 proteins. As an alternative, we generated point mutations in the predicted hydrophobic face of the helix. However, mutant PbYOP1 (residues 39–186, L45D/L49D/V52D) formed tubules like the wild-type protein when reconstituted with lipids (Fig. S4D). Taken together, these results suggest that the C-terminal region of the RHD domain is dispensable for generating membrane tubules and the N-terminus is important for the stability of the protein but may not function as an amphipathic helix to augment wedge insertion.
In addition to the “wedge” mechanism, ScYop1p and ScRnt1p can form homo- and hetero-oligomers through which these proteins may act as arc-like scaffolds to shape the membrane tubules (Shibata et al., 2009). We tested the oligomerization tendency of PbYOP1 using a sucrose-density gradient. When reconstituted PbYOP1 was solubilized by digitonin, a relatively mild detergent, it migrated in the gradient at a position that corresponds to a much higher molecular weight than that of a monomer (Fig. 2D). The oligomerization was largely disrupted when Triton X-100 or SDS was used instead (Fig. 2D). Notably, SDS-resistant dimers were often observed when PbYOP1 was separated by SDS-PAGE and analyzed by immunoblotting (Fig. 2D). These results confirm that PbYOP1 forms oligomers.
To further confirm the ER-shaping function of PbYOP1, we performed ER morphology rescue assays in yeast cells. When Sey1p, the ER fusogen in yeast, and Yop1p or Rtn1p are deleted, the peripheral ER (also termed cortical ER in yeast because it localizes underneath the plasma membrane) displays abnormal morphology. Large areas of the cortex are void of ER structures and most of the tubular ER network becomes sheet-like; the re-introduction of either Yop1p or Rtn1p in these cells restores the ER morphology (Hu et al., 2009). To ensure the expression of PbYOP1 in yeast, we utilized a 2μ vector with high copy numbers (pESC-URA) and drove protein expression using an inducible GAL promoter. The coding region of PbYOP1 was optimized for yeast codon usage to achieve detectable expression. As expected, most sey1Δyop1Δ cells expressing PbYOP1 exhibited normal ER morphology (Fig. 2E). The PbYOP1 levels were similar to ScYop1p expressed under control of the endogenous promoter (Fig. 2F). These results suggest that PbYOP1 can replace ScYop1p in maintaining proper ER morphology.
Next, we performed ER morphology rescue assays in mammalian cells and tested the function of PbSEY1. ER tubules become long and unbranched when ATL2 and ATL3 are deleted using CAS9/CRISPR system in COS-7 cells (ATL1 is not detectable in COS-7 cells) (Hu et al., 2015; Wu et al., 2015), indicative of the lack of fusion between ER tubules (Fig. 2G and 2H). As previously reported (Yan et al., 2015), when ScSey1p were expressed in these cells, the defects in ER morphology were largely restored (Fig. 2H). PbSEY1 do not express in yeast or mammalian cells, likely due to different codon usage. When codon-optimized PbSEY1 was expressed in ATL-deleted COS-7 cells, a majority of the mutant cells exhibited normal tubular ER network (Fig. 2H). These results confirm that PbSEY1 is functionally analogous to its yeast or mammalian orthologs.
Our findings reveal four ER-shaping proteins in the P. berghei genome. PbYOP1, PbYOP1L, and PbSEY1 have not been characterized previously. We showed that, similar to ScYop1p, PbYOP1 generated membrane tubules when purified and reconstituted in vitro, even though a conserved amphipathic helix next to the transmembrane domain is less important than in ScYop1p. We also confirmed the role of PbYOP1 and PbSEY1 in maintaining the tubular ER network in cells. Although the ER shaping activities of these proteins need to be further tested in Plasmodium, our results imply that the ER of Plasmodium parasites likely forms a tubular network via common mechanisms as described for yeast and mammalian cells. In addition, the ER-shaping proteins of Plasmodium parasites possess unique features that may meet specialized demands on the parasitic ER. The homolog of PbRTN1 in Plasmodium yoelii nigeriensis has been termed Pyn_chl091 due to its transcriptional up-regulation in mosquito vector when treated with chloroquine, an anti-malarial drug (Silveira et al., 2007). These ER-shaping proteins, including PbRNT1, likely adopt specialized roles during stressed conditions.
Very little is known about the morphology and function of the Plasmodium ER, especially during the pathogenic path of the parasites. Identification and preliminary characterization of Plasmodium ER-shaping proteins lays the foundation for further investigation of the Plasmodium ER. A lack of individual ER-shaping proteins in unicellular organisms, such as yeast, causes no drastic defects. However, in Candida albicans, an infectious fungus, deletion of Sey1p decreases its virulence (Yamada-Okabe and Yamada-Okabe, 2002), leading to the possibility that ER morphology determinants play a role in the infection of protozoan parasites, including malaria.
Electronic supplementary material
Below is the link to the electronic supplementary material.
FOOTNOTES
We thank S. Guo and D. An for assistance with electron microscopy. Q. Wang is supported by startup funds from Tianjin Medical University. Sha Sun, Li Lv, Zhi Yao, Purnima Bhanot, Junjie Hu and Qian Wang declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.
Footnotes
Sha Sun and Li Lv have contributed equally to this work.
References
- Brady JP, Claridge JK, Smith PG, Schnell JR. A conserved amphipathic helix is required for membrane tubule formation by Yop1p. Proc Natl Acad Sci USA. 2015;112:E639–648. doi: 10.1073/pnas.1415882112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu J, Shibata Y, Voss C, Shemesh T, Li Z, Coughlin M, Kozlov MM, Rapoport TA, Prinz WA. Membrane proteins of the endoplasmic reticulum induce high-curvature tubules. Science. 2008;319:1247–1250. doi: 10.1126/science.1153634. [DOI] [PubMed] [Google Scholar]
- Hu J, Shibata Y, Zhu PP, Voss C, Rismanchi N, Prinz WA, Rapoport TA, Blackstone C. A class of dynamin-like GTPases involved in the generation of the tubular ER network. Cell. 2009;138:549–561. doi: 10.1016/j.cell.2009.05.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu X, Wu F, Sun S, Yu W, Hu J. Human atlastin GTPases mediate differentiated fusion of endoplasmic reticulum membranes. Protein Cell. 2015;6:307–311. doi: 10.1007/s13238-015-0139-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishi M, Hu K, Murray JM, Roos DS. Organellar dynamics during the cell cycle of Toxoplasma gondii. J Cell Sci. 2008;121:1559–1568. doi: 10.1242/jcs.021089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orso G, Pendin D, Liu S, Tosetto J, Moss TJ, Faust JE, Micaroni M, Egorova A, Martinuzzi A, McNew JA, et al. Homotypic fusion of ER membranes requires the dynamin-like GTPase atlastin. Nature. 2009;460:978–983. doi: 10.1038/nature08280. [DOI] [PubMed] [Google Scholar]
- Shibata Y, Hu J, Kozlov MM, Rapoport TA. Mechanisms shaping the membranes of cellular organelles. Annu Rev Cell Dev Biol. 2009;25:329–354. doi: 10.1146/annurev.cellbio.042308.113324. [DOI] [PubMed] [Google Scholar]
- Shibata Y, Shemesh T, Prinz WA, Palazzo AF, Kozlov MM, Rapoport TA. Mechanisms determining the morphology of the peripheral ER. Cell. 2010;143:774–788. doi: 10.1016/j.cell.2010.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silveira H, Ramos S, Abrantes P, Lopes LF, do Rosario VE, Abrahamsen MS. Effect of chloroquine on gene expression of Plasmodium yoelii nigeriensis during its sporogonic development in the mosquito vector. Malar J. 2007;6:84. doi: 10.1186/1475-2875-6-84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teixeira JE, Huston CD. Evidence of a continuous endoplasmic reticulum in the protozoan parasite Entamoeba histolytica. Eukaryot Cell. 2008;7:1222–1226. doi: 10.1128/EC.00007-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Dooren GG, Marti M, Tonkin CJ, Stimmler LM, Cowman AF, McFadden GI. Development of the endoplasmic reticulum, mitochondrion and apicoplast during the asexual life cycle of Plasmodium falciparum. Mol Microbiol. 2005;57:405–419. doi: 10.1111/j.1365-2958.2005.04699.x. [DOI] [PubMed] [Google Scholar]
- Voeltz GK, Prinz WA, Shibata Y, Rist JM, Rapoport TA. A class of membrane proteins shaping the tubular endoplasmic reticulum. Cell. 2006;124:573–586. doi: 10.1016/j.cell.2005.11.047. [DOI] [PubMed] [Google Scholar]
- Wu F, Hu X, Bian X, Liu X, Hu J. Comparison of human and Drosophila atlastin GTPases. Protein Cell. 2015;6:139–146. doi: 10.1007/s13238-014-0118-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamada-Okabe T, Yamada-Okabe H. Characterization of the CaNAG3, CaNAG4, and CaNAG6 genes of the pathogenic fungus Candida albicans: possible involvement of these genes in the susceptibilities of cytotoxic agents. FEMS Microbiol Lett. 2002;212:15–21. doi: 10.1111/j.1574-6968.2002.tb11238.x. [DOI] [PubMed] [Google Scholar]
- Yan L, Sun S, Wang W, Shi J, Hu X, Wang S, Su D, Rao Z, Hu J, Lou Z. Structures of the yeast dynamin-like GTPase Sey1p provide insight into homotypic ER fusion. J Cell Biol. 2015;210:961–972. doi: 10.1083/jcb.201502078. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.