

Abstract
Cortical networks are composed of glutamatergic excitatory projection neurons and local GABAergic inhibitory interneurons which gate signal flow and sculpt network dynamics. Although they represent a minority of the total neocortical neuronal population, GABAergic interneurons are highly heterogeneous, forming functional classes based on their morphological, electrophysiological and molecular features as well as connectivity and in vivo patterns of activity. Here we review our current understanding of neocortical interneuron diversity and the properties that distinguish among cell types. We then discuss how the involvement of multiple cell types, each with a specific set of cellular properties, plays a crucial role in diversifying and increasing the computational power of a relatively small number of simple circuit motifs forming cortical networks. We illustrate how recent advances in the field have shed light onto the mechanisms by which GABAergic inhibition contributes to network operations.
I. Introduction
The computations taking place in the cerebral cortex depend on highly interconnected and dynamic microcircuits composed of two broad types of neurons: (1) glutamatergic excitatory neurons or principal cells (PCs), which propagate signals within and among various stations and (2) GABAergic interneurons (INs), which gate signal flow and sculpt network dynamics. The activity patterns of these interneurons thus play a critical role in information processing in cortex. To maximize flexibility, the cortex relies on the existence of a large diversity of GABAergic INs, which, as discussed in this review, differ over a large array of parameters (Ascoli et al., 2008). Anatomically, cortical GABAergic INs show a variety of somatic, dendritic and axonal morphologies, including the specific subcellular domain of pyramidal cells (and INs) targeted by their axons (Kawaguchi and Kubota, 1997; Kubota, 2014; Markram et al., 2004; Somogyi et al., 1998). IN subtypes also differ in their input and output connectivity with different cell types (both PCs and INs), which determines their circuit affiliation (Beierlein et al., 2003; Gibson et al., 1999; Jiang et al., 2015; Pfeffer et al., 2013). Electrophysiologically, a plethora of firing patterns have been observed, a consequence of the interplay of membrane cable properties and ion channel composition defining the passive and active membrane biophysical properties among IN subtypes (Kawaguchi and Kubota, 1997; Markram et al., 2004). In addition, the efficacy, kinetics and short-term dynamics of synaptic inputs and outputs have been shown to differ among INs (Beierlein et al., 2003; Gupta et al., 2000). There is also evidence that the synapses of specific IN types are associated with GABA receptors differing in subunit composition, which can affect the kinetics of the GABAergic response (Ali and Thomson, 2008; Freund, 2003). All these properties affect IN responses to excitatory inputs and their postsynaptic impact onto target cells. Reflecting differential receptor expression, GABAergic interneuron subtypes also vary in their response to neuromodulators such as acetylcholine (Ach), serotonin (5-HT), noradrenaline and dopamine, which profoundly affect the function of neocortical circuits and are responsible for dynamic changes associated with different brain states and behavioral contexts (Kawaguchi and Shindou, 1998; Munoz and Rudy, 2014). Finally, IN subtypes differ in their expression of molecules such as calcium-binding proteins and neuropeptides (Ascoli et al., 2008; Kawaguchi and Kubota, 1997; Kepecs and Fishell, 2014; Kubota, 2014; Markram et al., 2004). All these features highlight a large diversity within the GABAergic interneuronal population and most can have tremendous consequences on cellular and network computations. Although they represent a minority of all cortical neurons (10–15% in rodents; (Meyer et al., 2011) their local axons ramify extensively. While all GABAergic INs release GABA on their postsynaptic targets, the differences in subcellular targeting domain, connectivity, synaptic kinetics and intrinsic membrane properties result in highly specific and precise spatio-temporal inhibitory control of the activity of principal neurons and local networks.
The importance of INs has been appreciated since these cells were first described. Based on the observation that the abundance of “short-axon cells” increased during evolution, Santiago Ramon y Cajal concluded that the “functional superiority of the human brain is intimately bound up with the prodigious abundance and the unusual wealth of forms of the so called neurons with short axons” (Defelipe and Jones, 1988). Over the years evidence has accumulated suggesting that GABAergic interneurons have important roles in many cortical functions including gain control and dynamic range modulation of cortical circuits, sensory feature selectivity, plasticity, temporal precision of pyramidal cell firing, regulation of firing rates and bursting, synchronization and generation of cortical rhythms, and in the maintenance of the excitatory and inhibitory balance necessary for the transfer of information while preventing runaway excitation. Consistent with this role, malfunction of inhibitory INs has been associated with the generation of several types of epilepsy (Goldberg and Coulter, 2013). Moreover, INs have also been implicated in other diseases including schizophrenia, anxiety disorders and autism (Lewis, 2014; Marin, 2012).
Yet, it has been difficult to study cortical INs in the neocortex due to their large diversity and small representation of the total neuronal population. Recently, largely as a result of advances in molecular genetic methods to label, monitor and manipulate specific IN populations, there has been an explosion in the study of cortical INs. Application of these methods has facilitated the study of the circuits involving INs and the role they play in cortical function in behaving animals. Several reviews have been published in recent years highlighting these new discoveries (Kepecs and Fishell, 2014; Roux and Buzsaki, 2015) (see volume on Inhibition: synapses, neurons and circuits. Curr. Opin Neurobiol. Vol. 26, 2014). There have also been important advances in our understanding of the diversity of GABAergic interneurons in the neocortex. Although there is still no clear consensus on how many inhibitory cell types there are and how to differentiate them, clear patterns are emerging. In this review we discuss how the properties of different IN subtypes impact the function of the circuits in which they are embedded, aspects that recent reviews have for the most part not addressed. We first provide an updated understanding of the diversity of interneurons in the neocortex and cover the anatomical, molecular and functional properties defining different IN subtypes (Figure 1, Table I). We then discuss how distinct IN subtypes with unique properties integrate into circuits with specific computational impacts. We aim to show how recent research combining in vitro and in vivo preparations with genetic targeting and manipulations is helping shed light onto the division of labor among INs subtypes in neocortex.
Figure 1. Diversity, classification and properties of neocortical GABAergic interneurons.

Nearly all the INs in neocortex express one of the main three, non-overlapping, markers: Parvalbumin (PV, blue), somatostatin (Sst, red) and the ionotropic serotonin receptor 5HT3a (5HT3aR, green-yellow). Further subdivisions within each molecular group are revealed by morphological features, cellular and subcellular targeting biases, the expression of other markers, as well as some known anatomical, electrophysiological and synaptic properties.
Table I.
Morphological and electrophysiological properties of IN subtypes in neocortex
| Anatomical properties | Connectivity | Electrophysiological properties | Diversity | |
|---|---|---|---|---|
|
PV Basket cells (FS cells) (PV BCs) |
L2-6 Mostly multipolar dendritic arbor, with highly branching axonal arbor1–3. |
Form perisomatic chemical synapses onto soma and proximal dendrites of PCs and INs, especially other PV cells1,2,4–7. Electrically connected to each other5–8 and ChCs9, but apparently not to other INs5–8. |
Fast spiking firing properties. Brief spikes (300 us at~30°C), large fAHP. Can sustain high frequency firing with little or no adaptation. Low Rin, low Vrest1,10–12a. Subthreshold oscillations in the gamma range1,10–12a. Receive strongly depressing synaptic inputs, except from cortico-thalamic PCs5–8. Strongly depressing synaptic outputs5–8. |
Morphological: Layer-specific dendritic and axonal arbor. Intra- columnar (L4); trans- columnar (L2/3; L5/6) Intralaminar, translaminar types12b,12c. Electrophysiological: Diverse firing patterns (delayed, non- delayed, stuttering)3, 12a, 13,14 Molecular: Co-expression of molecular markers in PV basket cell subpopulations (e.g. calbindin); neuromodulators and receptors (e.g. cortistatin, tachykinin (TAC-1), substance P)15–17 |
| Chandelier cells (ChCs) or axo-axonic cells | Mainly L2, L5-6. Enriched in L1-L2 border and L618. More abundant in frontal than sensory cortices18. INs with most stereotyped morphology. Axonal branches with dense“cartridges” of vertically oriented strings of synaptic boutons1–3,9,18. Dendrites of L2 ChCs show bias for L19. |
Particularly specialized regarding postsynaptic target. All boutons of ChCs target exclusively the AIS of PCs1–3 (but see ref. 4). A single ChC innervates a few hundred PCs in a clustered manner19. May produce depolarizing and excitatory synaptic responses20. Electrically interconnected with each other and PV BCs9. |
Fast spiking with significant differences with PV BCs. In L2 higher Rin; slightly broader spike; smaller fAHP; earlier depolarization block. In L2 most PV BCs show long delays to first spike and spiking pauses; both are absent in ChCs9,18,19. | Molecular: PV+ and PV−18. |
| Multipolar bursting neurons (MPB)22 | Upper L2/3. Local, multipolar dendrites, wider axonal spread than PV BCs reaching other layers. Express PV and Calb. | Target dendritic shafts, sometimes spines, rarely perisomatic. High connection probability to L2/3 PCs with PPF. Excitatory inputs depressing. Unidirectional connectivity to FS cells. Electrically and chemically (with facilitation) connected to each other. | Initial burst followed by adapting, regular spiking. Wider spikes than FS. Carbachol-induced theta frequency oscillations in MPB network. | |
|
Sst Martinotti cells |
L2/3 and L5/6. Local axon arbor and long ascending axon that spreads horizontally and arborizes significantly in L1. Bipolar or multipolar dendrites1,2,23–25. | Dendritic targeting1–3,26. Target tuft dendrites of PCs in L126. Also target basal dendrites of PCs with local arborization26a. Some L5/6 Martinotti cells have significant axon arbor in L2/3; targets in L2/3 unknown4,23. Connect to other INs (PV, VIP) but not to each other 4–6, 27,28. Electrically connected to each other28a,29 |
Strongly facilitating excitatory inputs. Moderately depressing outputs. Powerful muscarinic depolarization 5,23,24,29–31. Higher Rin, Vrest, slower time constant than PV BCs. Bursting, RS or LTS (mainly in L5/6) firing patterns1,2,5–7,23,24,32. |
Electrophysiological: Different firing patterns: Bursting, RS or LTS 1,2,23,24,32. Molecular expression: CR, NPY, Kv3.2, reelin, calbindin, preprodynorphin, oxytocin receptora,16,17,17a, 17b. |
| Non-Martinotti cells | In L4 and L5. L4 non- Martinotti cells: Axon branches extensively in L4 with some branching in L2/323,24. Qualitatively similar to L4 PV BCs. L5/6 non-Martinotti cells: Local axon and ascending plexus mostly targeting L423. |
L4 non-Martinotti: Dendritic targeting. Innervate L4 PV cells more thanL4 PCs, but not other Sst INs. Electrically connected to each other. Weak TC innervation7,23,24,32,32a L5 non-Martinotti cells: Connectivity unknown |
Strongly facilitating excitatory inputs. Moderately depressing outputs. Powerful muscarinic depolarization. Faster action potential than Martinotti and slightly slower than PV BCs. Spike frequency adaptation7,23,24,32. | L4 vs L5/6 |
|
5HT3aR VIP bipolar34–36, 44 |
L2-6. Enriched in L2/3 (60% of VIP cells in L2/3). Rare in L1. Very small, ovoid soma. | IN targeting, mainly Sst INs | Irregular, regular, bursting and strongly adapting firing patterns Very high Rin44 |
Morphology: Bipolar, bitufted or tripolar with dendritic arbor largely vertically oriented. L4-6 bipolar cells have longer vertical dendrites than L2/3 VIP bipolar cells. Molecular: CR, ChAT, |
| VIP multipolar34–36 | L1-L2 border & deep layers | Not known | Regular adapting spiking; Very high Rin | Includes VIP+ CCK small basket cells. |
| NGFCs | L1-6. Major fraction of L1 INs. Multipolar cells with a small soma from which multiple short dendrites that spread radially and a wider, very dense, axonal plexus composed of fine branches1–3. | Outputs: High connection probability to most local PCs and INs. May mediate volume GABA transmission38. Make synaptic contacts with dendritic shafts and spines with little synapses onto cell bodies1,41,58. L1 NGFCs inhibit L5 PCs and L2/3 INs4,39,40. Produces slow GABAA and GABAB responses41. Inputs: In L1, conflicting data on whether they receive input from L2/3 PCs4,39,40 Callosal inputs42. Input from thalamic matrix in mPFC37. |
Late spiking firing pattern near threshold: a slow ramp depolarization preceding firing. Non-adapting spike trains and spiking pauses near threshold. Sometimes spike frequency acceleration, resembling what is observed in delayed FS INs. Small to moderate adaptation during suprathreshold depolarizations. Large AHP, slow ADP, very fast voltage sag1,2,37,39,40. |
Morphology: Trans columnar or “elongated” axonal arbor (in L1) and intra columnar or spherical axonal arbor in other layers1,2,39,40,43. Molecular: Most express reelin44,45; NPY, but less in L1. nNOS?1,2,37 |
| “Classical- accommodating;” “Cells with descending axons”; “Wide arbor cells”: “Single bouquet cells” (SBCs); Non LS (NLS)37,39,40,46 | Second ill characterized major component of L1; mainly inner part of L1; distribution in other layers unknown. Axon descends to L2 or deeper; sparser local axonal arborization than NGFCs. |
Does not produce GABAB responses; Receives input from L2/3 PCs. Innervates L2/3 INs. NLS cells in the mPFC responded primarily to cortico-cortical input terminating in the inner part of L1. Weak responses from thalamic matrix37. |
Compared to NGFCs: Broader AP, Smaller, slower AHP, no ADP. Spike trains with stronger adaptation than NGFCs and spike broadening. Large slow sag. | |
| CCK basket cellsc | Mainly layers 2/3 Includes large basket cells (CCK only), small basket cells, descending basket cells (also express VIP); and some double bouquet cells, a subtype of bipolar VIP cell2,47,48. | Perisomatic synapses on PCs and INs2,47,48. | Regular or burst spiking2,47,48. Asynchronous GABA release49,50. Contain CB1 receptors that inhibit GABA release and produce DSI (depolarization- induced synaptic inhibition)50–53. |
Express VIP or vglut354 |
| Long Range projecting33 | Mainly in L6 and white matter. | Mostly Sst (coexpressing nNOS)33,55, some PV33,56,57 and Vip57; corticocortical and corticofugal |
Not known if these molecular differences apply to Martinotti and/or non-Martinotti cells.
Not clear if this applies to both bipolar and multipolar VIP INs.
In the hippocampus there are basket cells and dendritic targeting CCK expressing interneurons (Cope et al., 2002); it is not clear if there are dendritic targeting CCK cells in the neocortex.
(Kubota, 2014);
(Thomson and Deuchars, 1997);
(Uematsu et al., 2008);
(Li and Huntsman, 2014);
(de Lecea et al., 1997);
(Blazquez-Llorca et al., 2015);
(Marlin and Carter, 2014);
(Cruikshank et al., 2010);
(Miyoshi et al., 2010);
(Kawaguchi and Kubota, 1998);
(Eggan et al., 2010);
(Neu et al., 2007);
II. Interneuron diversity in the neocortex
It is unquestionable that a mechanistic understanding of cortical circuits requires a complete knowledge of the neuronal elements that contribute to these circuits. Due to their diversity, this has been particularly difficult for GABAergic neurons. In fact, an effort to come to a consensus on IN classification in the cerebral cortex that took place a few years ago in Petilla de Aragon, the birthplace of Ramon y Cajal, did not succeed in accomplishing this goal. Instead, we produced a document discussing the features that can be a basis for distinguishing among IN subtypes (Ascoli et al., 2008).
In the hippocampal CA1 region, Somogyi and his colleagues have been successful at implementing an interneuron classification that starts with morphological features, i.e. somatic location and dendritic and axonal innervation fields (Klausberger and Somogyi, 2008; Somogyi and Klausberger, 2005). Then, the functional and molecular diversity can be mapped onto the IN classes proposed based on these morphological criteria. The success of this classification scheme depends largely on the simplified laminar architecture of the hippocampus, where axonal location largely defines possible postsynaptic targets of a given IN subtype. However, in the neocortex the intermingling of cell types in most cortical layers and cytoarchitectural differences among neocortical areas have made a parallel effort impractical. In fact, an attempt to produce a systematic classification of neocortical INs based on morphological features alone by a large number of investigators showed that there are too many ambiguities for “experts” to agree on the identity of most IN subtypes when strictly relying on morphological data (DeFelipe et al., 2013). In addition, morphological data has been for the most part obtained from brain slices and therefore only partial information is generally available (Stepanyants et al., 2009).
In the years since the Petilla meeting, the expression of certain molecular markers has emerged as a good starting point for IN classification in the neocortex. The evidence suggests that expression of these markers can provide an initial platform to start characterizing specific IN subtypes. Furthermore, advances in molecular genetics have allowed the generation of transgenic rodents expressing fluorescent proteins or the enzymes Cre or Flp recombinase under the control of the promoters for these markers, allowing the identification and manipulation of specific groups of INs (Taniguchi et al., 2011). Morphological and electrophysiological analysis of the INs expressing these markers is beginning to reveal clear patterns, as well as segregation of classes based on developmental origin and connectivity, all of which together are leading to a better understanding of IN subtypes in neocortex. These markers include the calcium-binding protein parvalbumin (PV), the neuropeptide somatostatin (Sst or Som), and the ionotropic serotonin receptor 5HT3a (5HT3aR)1. These three markers are expressed in largely non-overlapping interneuron populations in neocortex (Lee et al., 2010; Tasic et al., 2016; Zeisel et al., 2015) and together the three populations account for nearly 100% of GAD-67 mRNA-expressing neurons in somatosensory cortex (Lee et al., 2010). Based on these observations, neocortical INs can be divided in three major groups: PV, Sst and 5HT3aR INs (Figure 1). Importantly, INs from each of these groups show strong biases in functionally relevant properties that are either exclusive or not as prominent in other groups (Table I) as well as in gene expression patterns (Tasic et al., 2016; Zeisel et al., 2015), suggesting that this first level of separation is a good starting point for a hierarchical classification scheme.
The observation that PV, Sst and 5HT3aR account for nearly 100 % of the GABAergic neurons in primary somatosensory cortex (S1) suggests that if additional molecular groups that do not overlap with any of these three markers are discovered, they must represent minor populations, which does not imply that they are unimportant. It should be stressed however, that it is not clear to what extent a given molecular marker is a critical determinant of an IN group, since in most cases there is no clear causal relationship between the expression of a marker and most known functional features of an IN group. Thus, it is possible that INs not expressing any of the three markers could be lumped into one of the groups by the fact that they share other properties with that group. For example, Taniguchi et al. (Taniguchi et al., 2013) found that a significant fraction of chandelier cells, interneurons typically considered part of the PV group (see below), express PV protein weakly or perhaps not all. Weakly or non-PV-expressing chandelier cells have the same developmental origin as those that express the protein and it remains to be seen whether they are otherwise different.
The PV and the 5HT3aR IN groups can be subdivided each in at least two major, and clearly distinct, subgroups (Figure 1). The PV group includes the chandelier or axo-axonic cells, which target the axon initial segment of pyramidal cells and the fast-spiking (FS) basket cells, INs that make perisomatic “basket” terminals on PCs and INs, the latter being numerically dominant. Overall, PV cells are the largest IN population in the neocortex. The 5HT3aR group can be divided in two subgroups based on whether or not a 5HT3aR IN expresses vasointestinal pepetide (Vip), which is found in about 40 % of all 5HT3aR INs and is not expressed in PV or Sst neurons (Lee et al., 2010). The IN types shown in Figure 1 are still heterogeneous and include subtypes differing in morphological, electrophysiological and molecular properties (Table I). When such differences are revealed to be physiologically important and thus might define functional subtypes, they can be incorporated into an iterative classification scheme to achieve a more refined and comprehensive picture of IN diversity in the neocortex.
We discuss in the following sections how differences in morphological and electrophysiological features can be used to unravel the diversity present in each group shown in Figure 1. As we illustrate below the utility of these parameters depends on IN type. Morphological features, and specifically the organization and anatomical distribution of axonal and dendritic arbors, have the advantage that they are clearly physiologically relevant since they determine the potential inputs and output targets a given IN subtype might have and should be in principle the best parameter to consider next in the iterative classification scheme. On the other hand while basic electrophysiological features will impact a cell’s excitability, it is often unclear which of the various properties extracted by experimentalists are functionally relevant enough to segregate cells as different types within an IN group, in addition to the various experimental conditions used by experimentalists that can affect these parameters (e.g. temperature, solution content, animal age, etc.).
In addition to PV, Sst, 5HT3aR and Vip, other molecular markers are often used to label and study cortical interneurons. These include reelin, the Ca2+-binding proteins calbindin (here CB but also Calb1) and calretinin (here CR but also Calb2), and the neuropeptides cholecystokinin (CCK) and neuropeptide Y (NPY). In contrast to the first four, these markers are not expressed in non-overlapping populations of interneurons. For example, CR is expressed in subsets of both Sst and Vip INs. Therefore, by themselves these markers do not describe discrete IN populations (Figure 1; Table I). However, if any of them, or other gene products, turn out to be expressed in a functionally interesting subpopulation of any of the major groups of INs they could become a useful tool to label and manipulate specific subgroups using intersectional genetics (Fenno et al., 2014; Huang, 2014)(He et al., 2016). In addition, recent technical advances in single-cell mRNA-sequencing methods are now enabling unbiased, high-resolution transcriptomic analysis of individual neurons (Cadwell et al., 2016; Fuzik et al., 2016; Tasic et al., 2016; Zeisel et al., 2015). While traditional methods are limited by the need to average across a large population, the single-cell mRNA profiling provides a powerful approach to characterize and classify neurons at the single cell level. This approach opens up the possibility to identify new subpopulations of cells. One of the challenges in the single-cell RNA sequencing is the high level of variation due to the low efficiency of capturing each cell’s transcript complement. Thus, it remains to be addressed whether this noise limitation in the single-cell approach will obscure the identification of subpopulations. More importantly, it remains to be seen how the transcriptomic data from a single cell can be combined with and compared to other information that contributes to identify the cell (Cadwell et al., 2016; Fuzik et al., 2016). Furthermore, molecular subpopulations could represent different states or developmental histories of the same cell type, and their subdivision may not be of interest from the functional point of view. Nevertheless, the approach provides a great opportunity to identify new genes to better target subpopulations of INs. However, given the relatively young age of this field and that the functional significance of the newly proposed subtypes needs to be established, here we will not cover this new ground in detail.
PV INs
The PV group includes the fast-spiking (FS) basket and chandelier cells (Figure 1, Table I). Chandelier INs, also known as axo-axonic neurons, are the most recognizable INs in terms of morphology (DeFelipe et al., 2013) due to the unique candlestick-like synaptic terminal arrays they form to specifically target the axon initial segment of pyramidal cells. In contrast, the much more numerous basket cells make perisomatic “basket” terminals on the soma and proximal dendrites of PCs and INs. By producing hyperpolarizing and/or shunting inhibition (Fishell and Rudy, 2011) close to the site of action potential generation, both types powerfully influence the output of their target neurons. A third type of PV cells not shown in Figure 1 are the “multipolar bursting cells” (Blatow et al., 2003; Caputi et al., 2009). These neurons, found mainly in upper L2, differ from FS multipolar basket cells in their morphological and electrophysiological properties. This IN type has not been studied extensively, and no further information on these cells beyond the initial description has appeared.
PV basket cell subtypes have been shown to be associated with diverse dendritic and axonal arborization territories (Freund et al., 1983; Kisvarday, 1992; Kisvarday et al., 1985; Markram et al., 2004; Martin et al., 1983; Munoz et al., 2014; Somogyi et al., 1983; Thomson and Lamy, 2007; Wang et al., 2002)(Freund et al., 1983; Kisvarday, 1992; Kisvarday et al., 1985; Markram et al., 2004; Martin et al., 1983; Munoz et al., 2014; Somogyi et al., 1983; Thomson and Lamy, 2007; Wang et al., 2002)(Freund et al., 1983; Kisvarday, 1992; Kisvarday et al., 1985; Markram et al., 2004; Martin et al., 1983; Munoz et al., 2014; Somogyi et al., 1983; Thomson and Lamy, 2007; Wang et al., 2002)(Table I). Furthermore, laminar and columnar biases in axonal and dendritic arbors and connectivity have been observed to correlate with somatic laminar location (Bortone et al., 2014; Buchanan et al., 2012; Jiang et al., 2015; Kisvarday, 1992; Markram et al., 2004; Packer and Yuste, 2011; Thomson and Lamy, 2007). Some cortical layers (L4 in S1) contain only PV basket cells with largely local axon, while others (e.g. L5) in addition to PV basket cells with largely local axon have PV basket cells with local and translaminar axons. Since basket cells make perisomatic synapses, local cells will inhibit mainly local populations, while translaminar cells provide a means for interlaminar interactions via inhibition. In addition, some PV basket cells (particularly in supragranular and infragranular layers, but not in L4) have axons that span several columns, suggesting that in addition to providing inhibition to the column where they are located, they can influence neighboring columns. This is also true for some Sst-expressing Martinotti cells, and elongated neurogliaform cells as described below.
In some cases, association between morphological subtypes, synaptic properties and in vivo activity has been reported. For example, the complexity and extent of the dendritic arbor of supragranular PV INs in visual cortex correlate with their selectivity to visual stimuli (Runyan and Sur, 2013). In infragranular layers, it has been shown that PV cells with translaminar axons had excitatory inputs exhibiting less depression than locally projecting PV basket cells. This was due to expression of presynaptic NMDA receptors of glutamatergic axons contacting translaminar, but not locally projecting PV INs (Buchanan et al., 2012), which likely participates in making them more responsive to local pyramidal cell inputs (Bortone et al., 2014). This implies that PCs differentiate these PV basket cells as different subtypes. In the hippocampus, it has recently been shown that somatic laminar position and/or dendritic fields of PV expressing bistratified, axo-axonic and basket cells further segregate each class functionally during ripple events (Varga et al., 2014). Altogether, these studies illustrate that morphologically distinct PV basket INs, even from the same cortical layer, can exhibit functional differences.
As a whole, PV FS basket cells are the largest population of INs in the neocortex (Figure 2), and until recently they were the most studied IN population due to their number and very stereotypical fast and non-adapting firing pattern. Collectively, studies of their intrinsic properties have shown that PV basket cells have a remarkable array of molecular and cellular specializations to ensure that they produce a fast, reliable, strong and temporally precise inhibition on their target cells (reviewed in (Hu et al., 2014). The speed and precision of FS basket cell signaling are impressive. The delay between the peak of an action potential in a FS basket cell soma and the start of the uIPSC in a postsynaptic pyramidal cell is on the average 0.7 ms (at ~31 °C) and the jitter between different responses 0.19 ms (Rossignol et al., 2013). On the other hand the latency of disynaptic inhibition, which will include the latency in exciting the PV cell is less than 2 ms (Miles, 1990; Pouille and Scanziani, 2001). These specializations allow FS basket cells to function as coincidence detectors and impose this function onto their postsynaptic targets (see Box 1).
Figure 2. Laminar distribution of IN groups.

GABAergic interneurons are unevenly distributed within the cortical mantle. The PV group is a major component throughout the cortex, except in L1, where it is virtually absent. Sst neurons are found in all layers, most prominently in infragranular layers. 5HT3aR INs dominate in supragranular layers, however there is a clear laminar separation between VIP-expressing INs, the largest population in L2/3 and non-VIP interneurons, which represent ~ 90% of L1 INs. These laminar distributions are from mouse somatosensory cortex, but very similar distributions have been found in frontal and visual cortices (see (Xu et al., 2010) for comparison).
Box 1. PV FS basket cells are specialized for speed, efficiency and temporal precision.
The biophysical and molecular specializations responsible for the speed and precision of FS cell function cover the entire cell; from the excitatory synapses a FS basket cell receives in its dendrites, all the way through its axon and presynaptic terminals.
-
Specializations promoting fast excitation of PV FS cells. Fast EPSPs ensure rapid and reliable excitation of FS cells and limit EPSP summation to near synchronous inputs, promoting coincidence detection and facilitating compartmentalization of distinct input streams.
Fast Ca2+ permeable AMPA receptors containing GluR1flip subunits and lacking GluR2 subunits1,2
Low input resistance, and very fast time constant perhaps due in part to the developmentally-regulated expression of TASK-3 leak K+ channels3,4,. FS cells compensate for the low membrane resistance by receiving large excitatory synaptic inputs.
Membrane resistance lowest at the soma and proximal dendrites as compared to distal dendrites, helping the generation of brief EPSPs while facilitating fast propagation of EPSPs to the soma5.
Active dendritic properties (Kv3 channels and low density of voltage gated Na+ channels) facilitate generation of fast, temporally independent EPSPs6, enhancing FS cell’s ability to detect temporally coincident inputs and promote fast, reliable and temporally precise EPSP-action potential conversion6,7.
-
Specializations responsible for the generation of spikes with short latency and the ability to discharge very brief action potentials repetitively at very high frequencies with little adaptation during sustained stimulation,
Sub threshold-operating Kv1 channels at the axon initial segment of FS basket cells filter slowly rising depolarizing events, which activate these channels and suppress spike initiation8. Therefore, FS cells are preferentially driven by large quickly rising inputs that initiate spikes with short latency.
Expression of high levels of voltage-gated K+ channels with a depolarized activation voltage and fast deactivation rates of the Kv3 subfamily; channels specialized for rapid repolarization with little interference during the inter-spike interval. Kv3 channels also contribute to the generation of a large and fast AHP that facilitates the recovery of Na+ channels from inactivation9–11.
Voltage-gated Na+ channels with slower inactivation and faster recovery likely enable constant Na+ channel availability during spike trains and prevent spike frequency adaptation and spike threshold accommodation16.
-
Specializations producing fast and reliable action potential propagation in FS cell axons and terminals, including the ability of transmitting the high frequency firing generated in the proximal axon and contributing to fast and synchronous transmitter release.
FS cell axons contain an excessively high density of Na+ channels and prominent expression of Kv3 channels12. The high Na+ channel density compensates for the unfavorable morphological properties of PV basket cell axons (small diameter, extensive branching, and high bouton density) and increase conduction velocity reducing the delay between action potential initiation and uIPSC onset in postsynaptic cells.
Kv3 channels are also present in the synaptic terminals of FS cells13. Brief spikes in the axon and terminals contribute to fast and synchronous transmitter release12.
-
Specializations that produce fast, efficient, reliable and temporally precise transmitter release
Axo-axonic or chandelier cells are also considered fast spiking, although some differences in intrinsic electrophysiological properties with PV basket cells have been reported (Woodruff et al., 2009). However, much less is known about chandelier cells and it is not clear to what extent the features of speed and precision of FS basket cells described in Box 1 also apply to chandelier cells. In addition, some reports have found different excitatory input sources and in vivo responses between PV FS basket and chandelier INs (Massi et al., 2012; Xu and Callaway, 2009; Zhu et al., 2004).
The lack until now of a specific marker for chandelier cells has hampered a systematic analysis. However, alternative genetic strategies have improved the targeting of this cell type (Taniguchi et al., 2013; Woodruff et al., 2009). Chandelier cells are particularly specialized regarding their postsynaptic target. All postsynaptic boutons of chandelier INs have been reported to target exclusively the axon initial segment of pyramidal cells (Howard et al., 2005) an observation confirmed by many authors (however, this notion has recently been challenged based on paired recordings, see (Jiang et al., 2015)). This is by far the highest level of target specificity to be ever reported concerning IN type and connectivity. Chandelier cells have recently generated additional attention as a result of the discovery that in neocortex, GABAergic synapses in the axon initial segment have a depolarized reversal potential compared to those innervating the somatic domain due to a higher intracellular chloride concentration at the axon initial segment (Szabadics et al., 2006). Consequently, axo-axonic cells may excite rather than inhibit their postsynaptic pyramidal cells (Szabadics et al., 2006). However, this remains controversial (Glickfeld et al., 2009; Wang et al., 2014). It is not clear if depolarizing with a reversal potential still below threshold has predominantly an excitatory or shunting effect and will require further investigation, but it has been recently suggested that this depends on the excitatory state of the postsynaptic cell (Woodruff et al., 2011).
Sst INs
In contrast to PV INs, Sst INs are dendritic targeting (Dennison-Cavanagh et al., 1993) (de Lima and Morrison, 1989; Kawaguchi and Kubota, 1996, 1997; Wang et al., 2004), a feature that has important functional consequences (discussed in the Interneuron Circuits section). Sst INs also differ drastically from other INs in the dynamics of their excitatory inputs (Figure 1, Table I). Most INs have strongly or moderately depressing excitatory synapses. In stark contrast, excitatory inputs onto Sst INs, apparently regardless of subtypes, are strongly facilitating (Beierlein et al., 2003; Kapfer et al., 2007; Pouille and Scanziani, 2004; Silberberg and Markram, 2007; Thomson, 2003; Xu et al., 2013). This is a property determined by the postsynaptic cell, since the same excitatory axon has depressing synapses on a PV cell and facilitating onto a Sst IN (Buchanan et al., 2012; Reyes et al., 1998; Scanziani et al., 1998). Experiments in the hippocampus have shown that this unusual behavior is the result of the expression on Sst INs of the extracellular leucine-rich repeat fibronectin containing 1 (Elfn1) protein which regulates the release probability of the presynaptic terminal (Sylwestrak and Ghosh, 2012). It remains to be investigated whether the same or related proteins are responsible for the low release probability of excitatory synapses on SST cells in the neocortex.
As a result of the facilitating dynamics of their excitatory synapses, and other membrane properties of Sst INs that allow EPSP summation (Table I) excitatory inputs onto these cells produce supralinear responses. While PV INs require the synchronous firing of many presynaptic cells to fire due to their strongly depressing excitatory synapses and fast membrane time constant, the facilitation onto Sst INs enable even a single high frequency burst from one presynaptic cell to recruit Sst INs and produce feedback inhibition (Kapfer et al., 2007; Silberberg and Markram, 2007) (see Interneuron Circuits). Another feature of Sst INs is a muscarinic mediated depolarization. In response to bath applied agonists, the depolarization is strong enough that it is capable of producing prolonged spiking (Beierlein et al., 2000; Fanselow et al., 2008; Kawaguchi, 1997; Xu et al., 2013).
There is increasing evidence that Sst INs constitute a diverse group including cells that differ in morphological, electrophysiological and molecular properties (Table I). Despite these differences, all SST INs seem to have facilitating excitatory inputs and a muscarinic-mediated depolarization (Beierlein et al., 2003; Kapfer et al., 2007; Silberberg and Markram, 2007; Xu et al., 2013). Based on morphology, we can segregate Sst INs into two broad subgroups, Martinotti and non-Martinotti cells. In this review, we broadly define Martinotti cells as Sst expressing INs with a plexus of axon in L1, where it is known to target the tuft dendrites of pyramidal cells, including making synapses on spines (Chiu et al., 2013; Kawaguchi and Kubota, 1996; Wang et al., 2004). Non-Martinotti cells are here referred to as Sst INs lacking a significant axonal plexus in L1, despite sharing many of the properties of Martinotti cells (see Table I). We argue that this distinction is important since the cells we define as non-Martinotti (or non-L1-targeting) will clearly synapse onto different subcellular compartments or cell types and therefore will have a different impact on cellular and network computations than Martinotti cells (see below).
Martinotti cells are mainly present in L2/3 and L5/6 (Figure 1 and 2). In addition to arborizing in L1, a significant proportion of their axonal arbor, presumably contacting the basal dendrites of other neurons, is present in the layer where the soma is located. It seems that the vast majority of the Sst INs in supragranular layers and a significant fraction of those in infragranular layers are Martinotti cells. In contrast, the axon of most Sst INs in L4 of S1 largely remains within this layer, some of them additionally project to L2/3 (Ma et al., 2006; Xu et al., 2013). Moreover, L4 non-Martinotti Sst INs have several intrinsic electrophysological properties that differ from those of the Martinotti cells in supra and infragranular layers (see Figure 1, Table 1, (Ma et al., 2006; Xu et al., 2013)). Interestingly, L4 non-Martinotti INs also differ from Martinotti INs in terms of connectivity. While L2/3 Sst cells predominantly inhibit pyramidal neurons, L4 Sst INs predominantly target local PV INs and thus may produce disinhibition of L4 principal cells (Xu et al., 2013) (see Section on Disinhibition). However, it remains unclear whether the granular layer of other cortical areas contains Sst INs resembling those in S1. The morphological features of L4 non-Martinotti Sst INs are qualitatively very similar to L4 PV basket cells. However, synaptic dynamics, connectivity and electrophysiological properties, in addition to marker expression, clearly show that they are a functionally distinct IN subtype.
L5 of S1 seems to contain a yet undetermined but significant proportion of non-L1-targeting Sst cells (Ma et al., 2006; Munoz et al., 2014; Tan et al., 2008). The axons of infragranular non-Martinotti INs target mainly L4, but it is not known whether they also preferentially innervate PV cells, like L4 Sst INs do. In addition to these types, a few Sst cells in deep layers express nNOS (neuronal nitric oxide synthase) and are thought to have long-range projecting axons (Tamamaki and Tomioka, 2010) (see below).
It is not well established at present how other reported differences among Sst INs fit into this morphological classification. For example, L5 Sst INs have often been found to have an LTS (low-threshold spiking) firing pattern (Kawaguchi and Kubota, 1997; Ma et al., 2006). Although the term LTS has often been applied more broadly to Sst INs in general, the presence of rebound spikes from hyperpolarized potentials appears to be present more specifically in a subset of infragranular Martinotti cells, some of which are labeled with GFP in the transgenic X98 mouse line (Ma et al., 2006). Non-LTS Sst INs in L2/3 and L5 show regular spiking or burst spiking discharge patterns. On the other hand L4 and L5 non-Martinotti Sst INs typically have lower input resistance, brief spikes and fire at higher frequencies, resembling FS basket cells, but show much stronger firing frequency adaptation than these cells (Fanselow et al., 2008; Ma et al., 2006; Xu et al., 2013). The physiological significance of these differences in firing pattern is not yet clear, but they suggest a correlation between morphological subtype and electrophysiological properties.
Sst INs also display molecular heterogeneity. About 15–30% of the Sst INs in mouse neocortex express CR. Although so far only minor electrophysiological or morphological differences between CR+ and CR- Sst INs have been identified, distinct excitatory input patterns have been found (Xu and Callaway, 2009; Xu et al., 2006). Several other molecules are also expressed in subpopulations of Sst INs (see Table I). The relationship between molecular expression and morphological or electrophysiological diversity is not yet clear. However, recent transcriptional analysis of single cells suggests that Sst INs comprise genetically discreet subtypes (Tasic et al., 2016). As we discussed for PV cells, some of the molecules expressed by subpopulations of Sst INs are clearly important physiologically. For instance, a small subpopulation of Sst neurons in prefrontal cortex express oxytocin receptors, and this expression is critical to the modulation of sociosexual behavior by this hormone (Nakajima et al., 2014).
5HT3aR INs
5HT3aR INs represent ~30% of all neocortical INs and are thought to be more heterogeneous than the PV and Sst groups. However, all 5HT3aR INs express functional 5HT3a and nicotinic receptors (Lee et al., 2010). They are enriched in supragranular layers, where they represent the largest IN population (Figure 2). As mentioned previously, the 5HT3aR group can be divided in two subgroups based on the expression of the neuropeptide VIP (Figure 1). All neurons in L1 are GABAergic INs and most belong to the 5HT3aR group and are largely non-VIP-expressing. This layer contains the distal dendritic tufts of pyramidal cells, as well as intracortical axons from local PCs, long range inputs from other areas and corticopetal axons from high order thalamic nuclei and neuromodulatory centers. There is a great interest in this layer because of its presumed associative role and in top-down regulation of cortical processing as a result of the presence of projections from high order structures (Larkum, 2013). Based on their supragranular location, it has been suggested that 5HT3aR INs might be important mediators of such operations, a hypothesis supported by recent observations, as we will discuss further in this review.
Vip INs
Vip neurons represent about 40% of 5HT3aR INs in barrel cortex. They are present mainly in L2/3, but can be found in all layers (Figure 2). The large majority of Vip INs have a vertically oriented, bipolar-like dendritic morphology, the remaining being multipolar (Bayraktar et al., 2000; Pronneke et al., 2015). Dendritic trees of most bipolar Vip INs tend to be narrow and cross several layers in either direction and thus can sample translaminar inputs in several layers restricted to one column. Although the number of Vip INs in L1 is low, the dendrites of Vip INs in L2/3 extend fully through L1 reaching close to the pial surface, where they can be targeted by the many intracortical and subcortical projections to this layer. Consistent with their translaminar dendrites, L2/3 bipolar INs have been shown to receive inputs from several layers, which was less common for most other cell types studied (Xu and Callaway, 2009). Subtle differences among Vip INs with vertically oriented dendrites have been described. Some are bitufted, while others are single tufted, bipolar or tripolar (Bayraktar et al., 2000; Cauli et al., 2014). However, it is not clear that these differences in dendritic morphology are physiologically significant since they tend to sample similar intracolumnar and translaminar sectors. Here, we will use the term “bipolar” to denote all Vip INs with vertically oriented dentritic arbor. The axon of L2/3 Vip INs is also directed vertically, in a narrow columnar fashion, where their axonal projections often reach L4 and L5/6, in addition to their local axonal arbor (Bayraktar et al., 2000; Porter et al., 1998; Pronneke et al., 2015). These axonal features are reminiscent of what has been described as “horsetail” and double bouquet cells in primates (DeFelipe et al., 2006). Therefore, the direct influence of L2/3 bipolar Vip INs is likely to be vertically broad and laterally restricted. Interestingly, it has been observed that bipolar Vip INs in deeper layers follow different trends in their dendritic and axonal fields. L2/3 Vip INs had their dendrites largely restricted to supragranular layers and their axon extending to both supra- and infragranular layers. In contrast, Vip INs in deeper layers had dendrites spanning both supra- and infragranular layers, but had their axons restricted to L5/6 (Kawaguchi and Kubota, 1996; Pronneke et al., 2015). Multipolar VIP INs include VIP INs expressing CCK (see below) and a group of L6 multipolar VIP cells with an intralaminar axon spanning laterally (Bayraktar et al., 2000; Pronneke et al., 2015).
Subpopulations of Vip neurons express molecular markers that may help in subdividing this group of INs (see table I) (Cauli et al., 2014; Ferezou et al., 2007; Taki et al., 2000). About 10–30 % of Vip INs express Cck and about 50–70% of Vip INs express CR. In addition, CR and Cck are largely non-overlapping on Vip cells. Vip CR+ INs behave differently than Vip CR- neurons in terms of the role of activity in their migration and maturation during development (De Marco Garcia et al., 2011), suggesting that CR could be a useful marker to differentiate between Vip IN subpopulations. In fact, using intersectional Vip flp x CR cre mice, He et al., (He et al., 2016) found that VIP/CR cells were significantly enriched in irregular spiking bipolar neurons. Some Vip INs also express the ACh synthetizing enzyme choline acetyl transferase (ChAT). ChAT-expressing INs appear to be irregular spiking, CR+, and have “bipolar” dendritic morphology (Cauli et al., 2014; Porter et al., 1998). However, the cholinergic nature of these neurons is not clear. In fact in mouse and humans the vesicular acetylcholine transporter is not expressed in ChAT INs (Cauli et al., 2014). It is therefore unclear whether ChAT expression in VIP neurons is of functional significance. In contrast to Vip CR INs, Cck-expressing Vip INs tend to exhibit multipolar or bitufted dendrites, although bipolar cells can also be found (Freund et al., 1986; Kubota and Kawaguchi, 1997)(He et al., 2016). These neurons have small soma and are largely found in L2, although they are also present in other layers. These INs likely correspond to what has been referred to as small CCK basket cells, which, in contrast to the typical vertically oriented translaminar axonal arbor of the Vip bipolar cells, have a rather local axonal arbor (Freund et al., 1986; Kawaguchi and Kubota, 1996; Kubota, 2014; Wang et al., 2002).. At least a portion of Vip INs seems to form perisomatic basket terminals on their postsynaptic targets. Vip containing boutons have been found on both PCs and INs (David et al., 2007; Freund et al., 1986; Hioki et al., 2013; Kawaguchi and Kubota, 1996; Peters, 1990; Staiger et al., 2004). However, it is not clear if small basket cells expressing both Vip and Cck are the only source of these boutons. As we will discuss in the disinhibition section, Vip neurons, as a population, preferentially form synapses onto Sst neurons. This seems to be particularly true for the Vip bipolar (CR+) cells (Caputi et al., 2009; Jiang et al., 2015), which are the majority of the Vip cells. It is doubtful that Cck-expressing Vip INs exhibit the same connectivity pattern.
Perhaps, the most salient intrinsic electrophysiological feature of Vip INs is their relatively high input resistance, higher than most cortical neurons (Table I), a property that makes Vip neurons particularly sensitive to excitatory inputs. For instance, although thalamic stimulation in thalamocortical slices produces weak excitatory synaptic currents on Vip INs in L4 and deep L3, these can produce substantial depolarization due to their high input resistance (Lee et al., 2010).
Vip INs have often been described as having an irregular spiking (IS) firing pattern in response to depolarizing steps (Cauli et al., 2000; Lee et al., 2010; Porter et al., 1998). IS INs are characterized by an initial burst of action potentials followed by intermittent action potentials at an irregular frequency. The IS property is seen mainly at near threshold depolarizations, and is replaced by a regular adapting firing pattern during larger depolarizations. Porter et al., 1998, found that low concentrations of 4-AP, as well as DTX-I and DTX-K convert the IS firing pattern to a more regular discharge pattern, suggesting that a Kv1-mediated ID-like K+ current contributes to irregular spiking. Irregular spiking might be seen often in Vip cells as a result of their high input resistance, which increases the possibility that noise, an intrinsic subthreshold oscillation, or a small synaptic input will produce sufficient depolarization to reach spike threshold. The intermittent spikes observed during the train may represent spikes that escape the adaptation produced by the ID-like K+ current. In addition to irregular spiking, bursting and strongly adapting Vip cells have been reported (Cauli et al., 2000; Kawaguchi and Kubota, 1996; Lee et al., 2010; Porter et al., 1998; Pronneke et al., 2015). It is not clear to what extent these differences in firing pattern are reflective of distinct Vip IN subpopulations and what their functional significance might be. Excitatory inputs to VIP cells are depressing, as is the case for the inputs from most INs (Porter et al., 1998; Rozov et al., 2001). However, Caputi et al. (Caputi et al., 2009) suggested that the output synapses of CR+ Vip cells on pyramidal cells, as well as on PV and somatostatin INs are slightly facilitating.
Like all 5HT3aR INs, Vip INs are strongly depolarized by 5HT3aR agonists (Ferezou et al., 2002; Lee et al., 2010), in addition to showing nicotinic ACh responses, suggesting that activity in neuromodulatory centers, such as raphe and basal forebrain neurons, could rapidly activate these interneurons. Anatomical, pharmacological and optogenetic evidence support this view (Acsady et al., 1993; Arroyo et al., 2012; Choi and Callaway, 2011; Ferezou et al., 2002; Lee et al., 2010). It has been reported that Vip-expressing bipolar neurons are also depolarized by muscarinic agonists (Kawaguchi, 1997), however this has not been extensively studied.
non-Vip 5HT3aR INs
Non-Vip 5HT3aR INs represent about 60% of 5HT3aR INs and about 90% of all L1 INs (Figure 2). They include the neurogliaform cells (NGFC), Cck-expressing INs (presumably non-VIP Cck basket cells) and other less clearly defined types (Table I).
Neurogliaform cells
NGFCs, called spiderweb cells by Cajal, have a characteristic multipolar morphology consisting of a small, round soma from which multiple, very short dendrites spread radially in all directions and have a wider, spherical, very dense axonal plexus composed of fine branches (Kawaguchi and Kubota, 1997; Kubota, 2014; Olah et al., 2007). NGFCs have been described in all layers, but might be more prevalent in supragranular layers and are a major component of L1. The neurogliaform morphology has often been associated with a late-spiking (LS) firing pattern, characterized by a slow ramp depolarization preceding firing and non-adapting spike trains near threshold (Hestrin and Armstrong, 1996; Kawaguchi, 1995; Kubota et al., 2011a; Olah et al., 2009; Tamas et al., 2003). During larger suprathreshold depolarizations, the cells fire adapting spike trains (Kawaguchi and Kubota, 1997; Tamas et al., 2003). In fact, although the neurogliaform definition is an anatomical one, often reports of NGFCs properties have not relied on anatomical identification, likely because of limitations related to morphological recovery of the thin axon of these cells. Instead, many studies have simply assumed the late-spiking discharge pattern as a bona fide indicator of a neurogliaform cell. However, although a strong trend, this has been shown to not always be the case (Jiang et al., 2015). NGFC morphology and/or LS firing pattern have been associated with some markers, although none are though to exclusively target this cell type specifically (Table I).
In L1, a large proportion of INs with neurogliaform-type morphology have been reported to have an horizontal axonal arbor that extends for longer horizontal distances than classical NGFCs, perhaps spanning several columns and largely remaining within the same layer (Hestrin and Armstrong, 1996; Jiang et al., 2015; Jiang et al., 2013; Kubota et al., 2011b; Zhou and Hablitz, 1996). Sometimes called “elongated neurogliaform cells” (Jiang et al., 2015; Jiang et al., 2013) this subtype of NGFC resembles typical “spherical” NGFCs not only in having a very dense axonal arbor surrounding a short multipolar dendritic tree, but in having seemingly similar intrinsic and functional properties, including a frequent late spiking firing pattern, an apparent lack of spike frequency adaptation at threshold and in mediating GABAB responses in connected postsynaptic targets (Cruikshank et al., 2012; Hestrin and Armstrong, 1996; Jiang et al., 2013; Wozny and Williams, 2011).
The properties of the output synapses of NGFCs are very distinct from those of other INs. NGFCs elicit slow, long-lasting IPSPs on PCs and other INs through a combined activation of both GABAA and GABAB receptors (Olah et al., 2007; Tamas et al., 2003). GABAA responses from NGFCs exhibit unusually slow kinetics as compared to other INs, with a decay time constant in the order of tens of milliseconds (Price et al., 2008; Szabadics et al., 2007; Tamas et al., 2003). In addition, NGFCs are the only interneurons that have been shown so far to elicit unitary GABAB responses in response to a single action potential (Price et al., 2005; Price et al., 2008; Tamas et al., 2003). GABAB receptors display high affinity for GABA, slow G protein-coupled mediated signaling and a predominantly extrasynaptic localization (Gonzalez-Burgos, 2010). Repetitive firing of individual cells or concerted action of several INs is thought to produce sufficient extracellular accumulation of GABA to activate extrasynaptic receptors (Kim et al., 1997; Mody et al., 1994; Scanziani, 2000; Thomson and Destexhe, 1999; Thomson et al., 1996). For example, a single FS IN in auditory cortex can produce GABAB responses on connected PCs when stimulated at 80 Hz (Oswald et al., 2009). Repetitive firing of Sst INs has also been shown to produce GABAB responses (Urban-Ciecko et al., 2015). However, the NGFCs synapses are unable to sustain repetitive firing of the interneuron (Price et al., 2005; Tamas et al., 2003), which is inconsistent with the observation that repetitive or strong presynaptic activation is often necessary to recruit GABAB responses. The structure of NGFC axon and synapses is thought to explain their unusual output responses. NGFC synapses show small junctional area and are believed to have a relatively large cleft distance (Szabadics et al., 2007), which could increase the diffusion of GABA to reach extrasynaptic receptors. In addition, NGFC axons show high release-site density, many of which are apparently not associated with synapses and may thus mediate volume transmission of GABA (Olah et al., 2009). These factors are thought to produce increased GABA spillover at NGFC synapses, resulting in extrasynaptic activation of GABAB receptors as well as δ-subunit-containing GABAA receptors responsible for tonic inhibition (Szabadics et al., 2007). The slow GABAA response is thought to be the result of the GABA transient produced by NGFC cells as a result of the structural specializations of their synapse. NGFC synapses thus seem specialized for sparse temporal operation and tuned for long lasting ionotropic and metabotropic effects.
NGFCs have a high probability of connection to all neighboring neurons (Jiang et al., 2015; Olah et al., 2009). These observations are consistent with the idea that NGFCs mediate volume transmission of GABA (Olah et al., 2009). On the other hand, electron microscopy data showed synaptic contacts from NGFCs on dendrites and spines and only very few on soma (Kawaguchi and Kubota, 1997; Kisvarday et al., 1990; Tamas et al., 2003), suggesting that NGFCs also produce synaptic GABA release, and that this produces dendritic inhibition. Furthermore, the GABA released to the extracellular fluid from local NGFCs could bind to GABA receptors in dendrites and also produce dendritic inhibition. L1 NGFCs are connected to L2/3 and L5 PCs (Jiang et al., 2015; Wozny and Williams, 2011). Since their axon is largely restricted to layer 1, the “connection” must occur on the tuft dendrites of the pyramidal cells. Together these data suggest that along with SST INs, NGFCs are a major group of dendritic-targeting INs.
Cck basket cells
Other 5HT3aR INs include the non-Vip-expressing, Cck-expressing basket cells. Neocortical Cck non-Vip basket cells differ from those expressing Vip in a number of parameters. First, they have been mostly associated with the large basket cell morphology, having larger somata, dendritic and axonal span than Vip and Cck-expressing small basket INs (Galarreta et al., 2004; Karube et al., 2004; Kawaguchi and Kubota, 1997; Kubota, 2014; Kubota and Kawaguchi, 1997; Wang et al., 2002). In addition, INs expressing Cck having large somata also express CB1 cannabinoid receptors and sometimes Vglut3. On the other hand it is still unclear whether Vip INs expressing Cck express CB1 receptors (Bodor et al., 2005; Galarreta et al., 2004; Somogyi et al., 2004; Tasic et al., 2016). Cck large basket cells are INs with interesting functional properties and responses to modulators that are likely to contribute to their influence in neocortical networks. Like PV basket cells these INs provide perisomatic inhibition to principal cells. They have been studied in the hippocampus and neocortex, where their properties have been contrasted to those of PV FS basket cells (Freund and Katona, 2007; Glickfeld and Scanziani, 2006; Klausberger et al., 2005). These studies have suggested that while PV basket cells, which are efficiently and faithfully driven by local PCs, operate as “clockworks” controlling spike timing and the precision of cortical network oscillations, the activity of Cck INs depends on subcortical inputs that carry information about “mood” and the autonomic state of the animal (Freund and Katona, 2007). This hypothesis is based on the fact that in contrast to PV cells, Cck INs express 5-HT3 serotonin receptors and α7 and α4 nicotinic receptors postsynaptically, and CB1 cannabinoid receptors presynaptically. Activation of the CB1 receptors by cannabinoids inhibits GABA release, and mediates a phenomenon known as depolarization-induced synaptic inhibition (DSI) in which endocannabinoids generated in postsynaptic cells in response to depolarization suppress the inhibition mediated by the presynaptic cells. In addition, Cck basket cells release Cck, which has anxiogenic effects, and influence their targets via GABAA receptors enriched in α2 subunits that are known to mediate the anxiolytic effects of benzodiazepines (Freund and Katona, 2007). Furthermore, in contrast to PV basket cells, Cck basket cells release GABA asynchronously and can thus produce long-lasting inhibition (Hefft and Jonas, 2005). In the hippocampus there are dendritic-targeting CCK-expressing interneurons in addition to CCK basket cells (Cope et al., 2002). It is not known if this is the case also in the neocortex.
Electrical connectivity in interneurons
Electrical synapses mediated by gap junctions have been used as a defining feature of different interneuron subtypes and are a major component of the connectivity between interneurons. While pyramidal cells do not show electrical coupling in mature animals, the electrical connection probability among related interneurons remains high, but apparently mainly among interneurons of the same class. For instance, as initial studies showed, PV-expressing FS cells within a distance of about 100–150 um are densely interconnected, as is the case among “LTS” SST-expressing INs, but FS cells and SST cells are not electrically connected to each other (Amitai et al., 2002; Galarreta and Hestrin, 1999; Gibson et al., 1999; Hestrin and Galarreta, 2005). Homotypic coupling has also been reported for multipolar bursting PV INs and irregular-spiking INs expressing cannabinoid receptors; presumably CCK basket cells (Blatow et al., 2003; Hestrin and Galarreta, 2005) as well as for VIP interneurons (Karnani et al., 2016). Electrical coupling between cells has therefore been interpreted as a strong indicator of INs belonging to the same subtype. However, conflicting data exists regarding the electrical connectivity of neurogliaform cells. Chu et al., (Chu et al., 2003) reported that LS INs in layer 1 are interconnected by gap-junctions but that these INs are not connected to L1 non-LS INs. On the other hand Simon et al., (Simon et al., 2005) found that NGFCs in layer 2/3 are not only densely electrically connected among themselves, but also to many other IN types including FS basket cells, suggesting that electrical coupling of NGFCs is promiscuous (Simon et al., 2005). This apparent discrepancy needs to be clarified; more generally, the rules of electrical connectivity among the different types of 5HT3aR INs need to be investigated in more detail. Furthermore, now that it is clear that the PV and SST IN groups are heterogeneous it would be of interest to determine whether there is selectivity of electrical connectivity between IN subtypes that are members of the same major group (i.e. are Martinotti and not Martinotti cells in L5 interconnected?). There are already indications that PV basket cells and chandelier cells are electrically connected (Hestrin and Galarreta, 2005; Woodruff et al., 2011).
Gap junctions are symmetrical bidirectional synapses that pass both depolarizing and hyperpolarizing signals resulting in both excitatory and inhibitory PSPs. This depends on the speed of the AP and the speed and size of the AHP and the low-pass filtering properties of electrical synapses, which result in the transmission of slow membrane potential variations but the attenuation of fast changes such as fast action potentials (Galarreta and Hestrin, 2001; Gibson et al., 2005; Hestrin and Galarreta, 2005). As extensively shown by modeling and pair recordings in slices, electrical connectivity results in the generation of IN networks that fire synchronously, however small to no effects of the knockout of connexin 36, the main connexin isoform in interneurons, have been observed on oscillations and rhythms in spite of the loss of electrical connectivity (Buhl et al., 2003), an observation that could be explained by compensatory changes.
Long-range projecting GABAergic interneurons
As reflected in the term ‘interneurons’, the majority of cortical GABAergic neurons strictly target nearby cells and control the local network activity. However, it has been known that some cortical GABAergic neurons also project to other brain areas (Alonso and Kohler, 1982). Long-range inhibitory cells have been found to reciprocally connect hippocampus and septum, hippocampus and entorihnal cortex and different neocortical areas (reviewed in (Caputi et al., 2013)) as well as corticofugal neocortical GABAergic neurons projecting to the amygdala (Lee et al., 2014b) and basal ganglia (Jinno and Kosaka, 2004; Tomioka et al., 2015). In hippocampus-related projections, long-range GABAergic neurons tend to target other GABAergic neurons in target areas (Acsady et al., 2000; Basu et al., 2016; Freund and Antal, 1988; Melzer et al., 2012; Toth et al., 1993). In addition, the types of GABAergic neurons in long-range projections in the hippocampal system seem to be heterogeneous based on their molecular markers (reviewed in (Jinno, 2009)). In neocortex, long-range projecting GABAerginc INs have been generally assumed to be nNOS-expressing Sst INs (reviewed in (Tamamaki and Tomioka, 2010)). However, long-range projecting GABAergic neurons belonging to other molecular groups, such as PV and Vip, have also been found (Jinno and Kosaka, 2004; Lee et al., 2014b; Tomioka et al., 2015)(Table 1).
GABAergic INs as sources of neuropeptides
Neuropeptides such as Sst, Vip, Cck and NPY in specific IN subtypes have been useful markers to classify and characterize IN subtypes. However, these are neuromodulators that are known to have a powerful impact on the function of neurons and the INs that express them are the main source of these peptides in the cortex. It is thought that neuropeptide release requires high-frequency firing (Baraban and Tallent, 2004; van den Pol, 2012; Zupanc, 1996), but virtually nothing is known about the conditions in which they are released from INs. Differential regulation of release between neurotransmitters and neuropeptides is possible because they are stored separately in small synaptic vesicles (SSVs) and large dense-core vesicles (LDCVs) respectively. While SSVs are densely clustered in axon terminals, LDCVs are detected in axon, soma and dendrites (Morris and Pow, 1991; Pow and Morris, 1989). Release of neuropeptides from dendrites has been reported (Castel et al., 1996; Landry et al., 2003; Simmons et al., 1995). Some of neuropetidergic neurons express autoreceptors (Freund-Mercier et al., 1994; Hurbin et al., 2002). Thus, activation of peptide receptors on the dendrites or soma provides positive feedback to dendritic peptide release, thus dendritic neuropeptide release can be self-sustaining and long- lasting (Ludwig et al., 2005; Ludwig et al., 2002). The mechanisms by which cortical INs regulate the release of neuropeptides still needs to be addressed. It has been shown however, that Vip INs release Vip, a potent vasodilator, and that single cell stimulation of Vip+ bipolar neurons is sufficient to elicit vasodilatation of nearby arterioles (Cauli et al., 2004). Interestingly, acetylcholine produces vasodilatation of cerebral arterioles (Fergus and Lee, 1997), perhaps via the nicotinic activation of Vip INs. Molnar et al., (Molnar et al., 2014) recently reported that NGFCs strongly express insulin and that local application of glucose or glibenclamide to NGFCs mimics the excitation suppressing effect of applied insulin on local microcircuits.
Conclusions
The neocortical IN classification described here is still a work in progress. However, it seems flexible enough to incorporate new discoveries. This classification was based largely on studies carried out in mouse barrel cortex. However, studies comparing various neocortical areas show largely preserved distributions of non-overlapping markers (PV, Sst, 5HT3aR and Vip) across areas (Xu et al., 2010). On the other hand we expect areal specific differences in the lower subdivisions of the hierarchical classification, since the location of dendritic and axonal processes of specific INs is expected to be circuit specific. Areal differences in inhibitory connectivity patterns have in fact been reported in neocortex (Katzel et al., 2011; Packer et al., 2013).
Similar IN types are observed in mouse and rat (Kubota, 2014), and our narrative often used data from rat studies. Interneurons have also been studied in other species beside rodents. Although similar morphological features and marker expression have been observed in primates, some differences in electrophysiological properties have been reported between rats and monkeys (Povysheva et al., 2007; Povysheva et al., 2008; Zaitsev et al., 2005; Zaitsev et al., 2009). It seems that overall, the same basic interneuron subtypes exist across most mammalian species studied (DeFelipe, 2002). However, some additional subtypes have been described in higher mammals (DeFelipe, 2002). These could result in some interneuron groups containing further subdivisions or specializations within more complex cortices. For example, primary visual cortex of primate has a more elaborate laminar organization and it seems that axonal fields of interneurons seem to match this complexity (Lund, 1988). Species differences in marker expression patterns have also been reported (Hof et al., 1999). However, it is not clear to what extent the markers define the same IN subtypes in these species as they do in rodents. Species differences in the relationships between marker expression and cell type identity may arise due to the fact that there is no clear mechanistic link between these markers and morphological or electrophysiological phenotypes. Therefore, these markers are thus far only providing convenient correlations with the cell types mentioned above. In this respect, further work on unraveling the transcriptional programs determining cell type identity from developmental studies or RNA sequencing should be of tremendous help if successful (Huang, 2014; Kepecs and Fishell, 2014; Tasic et al., 2016; Zeisel et al., 2015).
III. Interneuron circuits
Having described the main classes of INs and the key intrinsic features defining them, we will now cover the circuits they are embedded in and highlight how specific properties of INs within a given wiring configuration bring about different modes of operation to those circuits. The study of IN circuits is an active and fast moving field and in recent years a large number of studies using a large variety of approaches have provided sets of connectivity rules for specific IN subtypes.
According to Peter’s rule connectivity would be established by chance, and would be a reflection of the statistical occurrence of presynaptic axonal terminals being closely apposed to potential postsynaptic targets as dictated by the geometrical distribution of axons and dendrites (Braitenberg and Schuz, 1991). While examples that Peters’ rule (Peters and Feldman, 1976)is closely followed in the cortex exist, virtually as many exceptions have been described, even at the time that the rule was proposed (Braitenberg and Schuz, 1991). Interestingly, many of these initial exceptions involved aspiny, putative INs. For example, thalamocortical inputs to L4 preferentially make synapses onto some interneurons as compared to others (Beierlein et al., 2003; White and Rock, 1981; Xu et al., 2013), and intracortical axons of L6 corticothalamic neurons show a preference for targeting interneurons over excitatory cells (Bortone et al., 2014; McGuire et al., 1984; White and Keller, 1987). As described earlier, chandelier cells appear to be particularly specialized in terms of their postsynaptic targets, since apparently these neurons make synapses exclusively with pyramidal cells, targeting the axon initial segment of these neurons (however see (Jiang et al., 2015)). It seems that whether or not Peters’ rule applies has to be investigated in a case-by-case basis. For example, Packer et al. (Packer et al., 2013) found that PV basket cell connectivity to pyramidal neurons could be largely explained by axo-dendritic overlap, agreeing with Peters’ rule. On the other hand, Sst INs, despite having axonal plexus overlapping with dendritic domains of other Sst cells show very little interconnectivity (Adesnik et al., 2012; Dennison-Cavanagh et al., 1993; Gibson et al., 1999; Pfeffer et al., 2013), while densely connecting to other IN types and pyramidal cells (Fino and Yuste, 2011; Pfeffer et al., 2013). Finally, connectivity assessed by connection probability alone does not reflect connectivity strength. Even if INs densely contact neighboring excitatory cells and vice versa, some evidence suggests that PV INs exhibit output strengths that correlate with the amount of excitation received by the postsynaptic cell (Xue et al., 2014) or the type of postsynaptic projection neuron (Lee et al., 2014a; Lee et al., 2014d).
Given the lack of universal connectivity rules it is necessary to investigate the connectivity of each interneuron subtype with all its possible targets. There are three main methodologies available to study neuronal connectivity. First are anatomical methods that can provide information on the presence of connections and their subcellular targeting location, but lack functional information. Second there are paired recordings in living cortical slices, in which a presynaptic cell is activated while recording from putative postsynaptic cells to test for possible connections. This method provides information on the probability of connection and the strength of unitary connections. However, it suffers from the important limitation that many connections are cut during the preparation of the slices. Lastly, there are optogenetic methods in which channelrhodopsin is genetically expressed in specific types of presynaptic neurons that are then photoactivated while recording from postsynaptic neurons. Connectivity studies using this method are not affected by cut axons because these remain viable and can still be photoactivated. This method provides information on the global connectivity of a genetically defined type and does not provide independent information on probability of connection and unitary strength. Give these challenges our understanding of interneuron connectivity is quite incomplete, but it is necessary to understand the recruitment of different interneurons to the various circuit motifs described below and their role in sculpting local network dynamics. Not only there are still many details missing, but the literature contains conflicting data, likely in part due to differences in technical approaches, area and layer investigated as well as to how cell types were defined. With these caveats in mind, our current view of the connectivity of specific interneuron subtypes with pyramidal cells and other interneurons is summarized in Figure 3. Modifications to the scheme illustrated in this figure can be expected as we improve our means to identify specific interneuron subtypes and connectivity studies progress.
Figure 3. Cell-specific connectivity and subcellular domains targeted by interneuron subtypes.

(A) Main known connectivity of INs to L2/3 pyramidal cells. The two major IN subtypes targeting dendrites are Sst Martinotti cells (red) and non-Vip 5HT3aR neurogliaform cells (dark yellow). Both L2/3 and L5 Martinotti cells have been shown to connect to L2/3 PCs (Jiang et al., 2015; Kapfer et al., 2007). Since their axons target both L2/3 and L1, it is assumed that supra- and infragranular Martinotti INs are positioned to contact basal, apical and tuft dendrites. L1 and L2/3 NGFCs are known to have axonal arbors largely restricted to their own layer and thus basal and apical dendrites are expected to be targeted by L2/3 NGFCs while tuft dendrites should be contacted by L1 NGFCs (Hestrin and Armstrong, 1996; Jiang et al., 2015; Kawaguchi and Kubota, 1997; Wozny and Williams, 2011). The perisomatic region and proximal dendrites are targeted by Cck basket cells (light yellow) and PV fast-spiking basket cells (dark blue). The axon initial segment is contacted by PV chandelier cells (light blue). Although connections by Vip bipolar cells onto PC soma and proximal dendrites have been reported (Kawaguchi and Kubota, 1997; Meskenaite, 1997), the connectivity is relatively weak with low probability. (B) Main connectivity onto L5/6 pyramidal cells. L2/3 Martinotti cells only poorly contact infragranular PCs (Jiang et al., 2015; Jiang et al., 2013) thus among Martinotti cells only those in infragranular layers target L5 PCs. In contrast, L1, L2/3 and L5/6 NGFCs contact L5 PC dendrites. Perisomatic and axonal contacts are provided by infragranular basket and chandelier cells. (C) Main inhibitory connections among INs. NGFCs target all interneuron subtypes and thus their connectivity is solely restricted by the extent of their axonal arbor (Jiang et al., 2015). Martinotti cells contact all IN subtypes except other Sst INs (Pfeffer et al., 2013). Input and output connectivity of neocortical Cck basket cells with other INs has not been investigated, however it is assumed that they receive inputs from both Martinotti cells and NGFCs. PV basket cells are known to strongly connect to other PV cells. Although connections from PV cells to other INs have been reported, conflicting data exists in the literature and have thus been omitted here. Vip bipolar cells are selective for other interneurons, particularly Sst INs. L2/3 bipolar Vip INs can contact both L2/3 and L5 Sst INs. While the connectivity of infragranular Vip cells has not been investigated, it is assumed to show similar cell type selectivity than supragranular Vip INs and be restricted to deep layers given that their axons do not ascend to supragranular layers (Bayraktar et al., 2000; Pronneke et al., 2015). Only the general connectivity patterns of the most well-known cell types are illustrated in this figure. It should also be noted that that many PC subtypes exists in every layer and although some PC subtype-specific connectivity by INs has been shown, this has not been thoroughly investigated and therefore has been omitted.
Proximal versus distal inhibition
GABAergic synapses are distributed along the entire axis of principal cells and interneurons (Gulyas et al., 1999; Hioki et al., 2013; Megias et al., 2001). However, as discussed earlier, different interneuron types show subcellular compartment target biases (Bloss et al., 2016). This is particularly important in the case of pyramidal cells, which have elaborated dendritic fields differentially associated with specific excitatory input sources and intrinsic active properties. Moreover, distal regions of pyramidal cell dendrites can be separated from the soma and the action potential initiation zone in the axon initial segment by long distances. Local GABA release can affect target cells as a result of changes in membrane potential through a local IPSP that will decrease with distance according to the cell’s cable properties, as well as changes in membrane resistance created by the local conductance change (shunting inhibition) (Fishell and Rudy, 2011). Due to the electrotonic attenuation of the IPSP elicited by GABA and the local shunting effect, the inhibition will be most effective near the contact point (Koch et al., 1983; Liu, 2004). In passive dendrites, shunting will largely act locally and will be most effective at counteracting propagation of excitation if placed on its path to the soma (proximal inhibition) than if localized off the path (distal inhibition) (Koch et al., 1983; Vu and Krasne, 1992). This suggests that proximal inhibition is more likely to globally modulate the output of a cell, while distal inhibition acts more locally, such as on specific dendritic branches. However, when considering local dendritic non-linear regenerative properties such as calcium spikes through NMDA receptors or calcium channels, modeling and experimental evidence have shown that distal inhibition is more effective at preventing these non-linearities from reaching threshold than proximal inhibition due to the higher relative weight of the conductance change of inhibitory synapses placed near the sealed end of dendritic branches than those placed near the soma (Gidon and Segev, 2012; Miles et al., 1996). Given the importance of such nonlinearities for signal propagation, calcium dynamics and synaptic plasticity, distal inhibition by dendritic targeting INs can thus control the efficacy and integration of glutamatergic inputs from specific sources impinging upon specific dendritic domains. They can also control Ca2+ dependent biochemical reactions and hence influence plasticity (Spruston, 2008). For example, tuft dendrites in L1 are contacted by long-range cortico-cortical feedback inputs and high order thalamic nuclei (Petreanu et al., 2009), which have been shown to trigger such calcium events (Gambino et al., 2014; Xu et al., 2012). Dendritic Ca2+ spikes interact with backpropagating somatic spikes to increase the gain of the input/output function of the pyramidal cell and facilitate a burst-firing mode (Larkum et al., 1999). These calcium dynamics can be modulated by dendritic targeting INs, such as Sst INs. In this view, distal inhibition acts as a gate for input integration. It has been observed that even a single presynaptic dendritic targeting IN can control Ca2+ spike generation in pyramidal cells (Larkum et al., 1999), and using a fibre-optic method for recording dendritic calcium changes in vivo Murayama et al. (Murayama et al., 2009) showed that dendritic inhibition could control the slope of the stimulus-sensory response function of L5 pyramidal cells in awake and anaesthetized rats. Moreover, recordings from apical dendrites in vitro showed that activity in L5 pyramidal neurons disynaptically coupled via the Sst interneurons directly blocks the initiation of dendritic calcium spikes in neighboring pyramidal neurons. In contrast INs that target the perisomatic area of PCs, such as PV and CCK basket cells and chandelier cells have a stronger influence on regulating the spiking output of the cell, its timing and thus the firing synchrony of populations of neurons (Cobb et al., 1995; Miles et al., 1996; Royer et al., 2012). Therefore, both proximal and distal inhibitory loci are crucial in determining whether or not dendritic signaling and action potentials will be temporally coupled or occur independently.
It is generally thought that proximal and distal inhibition would exert divisive and subtractive gain changes, respectively (Atallah et al., 2012; Koch et al., 1983; Pouille et al., 2013; Vu and Krasne, 1992; Wilson et al., 2012). However, this view has also been challenged (Gidon and Segev, 2012; Lovett-Barron et al., 2012; Mehaffey et al., 2005; Mitchell and Silver, 2003). This is not so surprising since not only the sites of excitation and inhibition, but also their temporal relationship, the instantaneous membrane potential and conductance state, dendritic arbor complexity, active conductances as well as passive cable properties all come into play leading to complex cellular computations (Silver, 2010). As the same IN type can be part of many circuits, it is likely that the cellular effect of inhibition will depend on the instantaneous cellular and network states. For example, the local membrane potential and resistance of a postsynaptic cell will determine whether the dominant contribution of an inhibitory input will be a conductance change (divisive shunt) or a current flow (subtractive offset). For example, if the local membrane potential is at the reversal potential for chloride, no inhibitory current will occur and the effect of an inhibitory input will be entirely shunting. On the other hand, if the resistance of the cell is already low, the relative change of adding an inhibitory conductance will be lower than if the membrane is in a low conductance (high resistance) state (Gidon and Segev, 2012). As mentioned above, the location of inhibition relative to excitatory inputs will affect how EPSPs travel along the dendritic arbor, summate and activate voltage-gated conductances. It is therefore likely that in in vivo conditions, where spatiotemporal patterns of excitatory, inhibitory and neuromodulatory loads vary with context, different or even the same IN types will participate differently in cellular computations (El-Boustani and Sur, 2014).
Circuit motifs in the neocortex
While all these factors influence the impact a given interneuron will have on the circuits in which it is embedded, a small collection of circuit motifs involving INs have been highlighted and provide a useful perspective alluding to this contextual excitation and inhibition relationship. It is thought that cortical circuits involving INs can largely be accounted for by different iterations and interactions of such core motifs. The main “archetype circuit motifs” are feedforward inhibition, feedback inhibition and disinhibition (Figure 4). It should be stressed that these motifs coexist in probably all interneuron subtypes. However, contextual differences (connection probability, synaptic strength and network state) as well as INs intrinsic properties and subcellular targeting domains lead to specific IN subtypes exhibiting a more potent role in one circuit motif over another.
Figure 4. Circuit motifs involving INs.
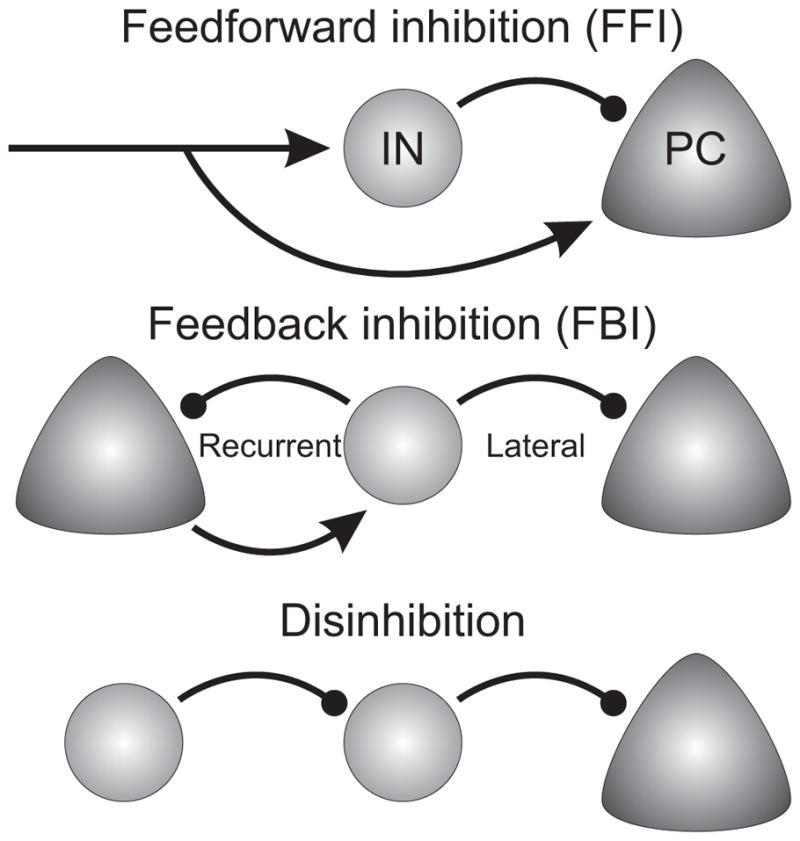
In feedforward inhibition (top), an external source makes excitatory synapses (arrows) onto both local principal cells (PC) and interneurons (IN). Interneurons in turn provide inhibitory inputs (black dot) to PCs. Feedback inhibition (middle) occurs when the source of excitation is local. INs can in turn make inhibitory synapses on the local PCs that provided the excitation (recurrent) or other neighboring PCs that did not participate in the recruitment of the IN (lateral). In disinhibition (bottom), the principal target of an IN is another interneuron, preventing it from inhibiting PCs.
A. Feedforward inhibition
Feedorward inhibition (FFI) is the process by which an afferent excitatory input source, in addition to contacting principal neurons, also synapses onto local inhibitory neurons, which in turn provide disynaptic inhibition to the principal cells receiving the excitatory input (Figure 4, 5). Conceptually, a FFI motif in its pure form is inherently a circuit mechanism regulating local integration by tracking the activity of incoming inputs, independently of the local network activity (Buzsaki, 1984). However, in reality this should not be taken literally, since the coexistence of other circuit and cellular mechanisms can effectively modulate the ability of FFI circuits to accomplish this task. Virtually any external excitatory input source to a neocortical area has been observed to also trigger short latency disynaptic inhibition (Toyama et al., 1974) and FFI seems ubiquitous in the central nervous system. In most, but not all, reported cases in neocortex, FFI is mediated by PV FS basket cells. The involvement of perisomatic targeting PV neurons, in combination with their intrinsic properties enabling high speed and temporal fidelity, provides unique high pass filtering properties to these feedforward inhibitory circuits, imposing coincidence detection onto postsynaptic neurons.
Figure 5. Thalamocortical FFI by PV neurons imposes coincidence detection.

A. Thalamocortical neurons synapse onto both excitatory principal cells (PC) and PV neurons. Thalamocortical connections are stronger onto PV than PC neurons. B. FFI by PV INs curtails TC mediated EPSPs on PCs leaving a narrow temporal window of opportunity for excitatory inputs to summate. Consequently, near synchronous inputs are required for efficient summation of EPSPs and to drive action potential firing on the PC. C. Weakening of PV INs FFI by short-term depression of TC synaptic inputs onto PV cells and PV INs outputs to PC (Gabernet et al., 2005). These two steps of adaptation weaken FFI more than direct feed-forward excitation of PC neurons. D. Weakening of FFI by modulation of PV IN output synapses. Both GABAB receptor-activation by NGFCs (Chittajallu et al., 2013) and muscarinic receptor-activation by acetylcholine (Kruglikov and Rudy, 2008) reduce inhibitory outputs to PCs. E. Relationship between the summation of EPSPs on PCs and their temporal difference for different strengths of FFI. As FFI is weakened, asynchronous inputs can summate more effectively (Pouille and Scanziani, 2001). F. FFI regulates the gain of PC populations. As excitatory drive increases, PV cells recruitment increases at a higher rate than the recruitment of excitatory cells. This will prevent PC population from saturation and will allow a wider dynamic range of the local PC population than if inhibition was absent (dotted line) (Pouille et al., 2009).
Thalamocortical feedforward inhibition
The best-studied feedforward inhibitory circuit in the neocortex is the one mediated by PV FS basket cell recruitment by thalamocortical neuron (TC) afferent axons in the input layer of primary sensory cortices. Although most studies dissecting this circuit were performed in the somatosensory system of rodents (Simons and Carvell, 1989), it seems that its core features are also present and are at least qualitatively similar in other sensory modalities such as vision and audition as well as in other species (Kloc and Maffei, 2014; Miller et al., 2001; Schiff and Reyes, 2012). In fact, the first convincing evidence for the existence of such a circuit came from in vivo intracellular recordings in cat V1 (Toyama et al., 1974). Crucial for thalamocortical transformation and by extension sensory processing in neocortex, this microcircuit involves L4 primary sensory thalamic afferents, L4 PCs projecting locally and to other layers, and local PV FS basket interneurons. TC axons synapse onto both PCs and neighboring PV cells (Cruikshank et al., 2007; Gabernet et al., 2005; Inoue and Imoto, 2006), but show higher connection probability onto PV INs through larger convergence and divergence (Bruno and Simons, 2002; Cruikshank et al., 2007; Inoue and Imoto, 2006; Swadlow and Gusev, 2002). Unitary thalamocortical connections onto PV cells are faster and fourfold stronger than those onto the PCs(Cruikshank et al., 2007; Gabernet et al., 2005; Inoue and Imoto, 2006), provided by high quantal amplitude with calcium permeable AMPA receptors (Hull et al., 2009) and multiple synaptic contacts forming clusters of neurotransmitter release sites (Bagnall et al., 2011). As a result of these synaptic specializations very few inputs are required to drive PV neurons, which in turn form strong, perisomatic GABAergic synapses back onto PCs resulting in a powerful disynaptic feedforward inhibition of these neurons (Figure 5). Because inhibition of PCs by PV INs is delayed by one synapse, disynaptic feedforward inhibition of PCs lags behind their monosynaptic thalamocortical excitation. However, given the synaptic and intrinsic properties of PV cells (Box 1), the delay is very short (1–2 ms), creating a limited temporal “window of opportunity” (Alonso and Swadlow, 2005; Pinto et al., 2000) for PCs to summate afferent inputs that will bring them to fire and transduce sensory signals (Gabernet et al., 2005; Wehr and Zador, 2003; Wilent and Contreras, 2005).
As a result of this circuit, thalamo-recipient principal neurons act as coincidence detectors of near synchronous thalamic input (Figure 5B) and improve the sensitivity of cortical neurons to the temporal distribution of thalamic spiking activity (Bruno and Sakmann, 2006; Bruno and Simons, 2002; Cardin et al., 2010; Pinto et al., 2000; Pinto et al., 2003), where slowly developing, asynchronous thalamic activity patterns are suppressed by FFI in the L4 network. On the other hand, stimuli producing synchronous thalamic firing will result in postsynaptic responses strong enough during the window of opportunity to drive the firing of L4 PCs. As such, thalamocortical FFI in barrel cortex participates in feature selectivity of L4 PCs by enhancing the differences in cortical responses between preferred and non-preferred stimuli (Pinto et al., 2003; Wilent and Contreras, 2005), thus making the cortex more discriminating than the thalamus. The efficient thalamic recruitment of strong FFI allows L4 neurons to encode the temporal features of sensory inputs. This mechanism is critical for representing velocity and direction selectivity of whisker deflection in the rodent barrel cortex (Pinto et al., 2003; Wilent and Contreras, 2005) and producing cortical responses that more precisely represent the timing of sensory input (Gabernet et al., 2005; Higley and Contreras, 2006).
Modulation of the strength and temporal aspects of the feedforward inhibitory circuit and the window of opportunity has also been shown to be of functional relevance (Figure 5C–E). Upon repetitive thalamic firing, stimulus adaptation takes place where TC inputs to both PV and PC neurons depress. Inputs to PV INs depress much more than those to PCs, and PV neuron synapses onto PCs are also depressing. This results in a decreased fidelity in the recruitment of PV INs and a widening of the temporal window of opportunity of excitatory cells following repetitive stimulation allowing more jitter in cortical spiking activity to take place (Gabernet et al., 2005) (Figure 5C). Interestingly, this adaptive modification of the window of opportunity has been suggested to be an important component allowing the switching from detecting the presence of a sensory stimulus to discriminating between different stimuli (Wang et al., 2010) Other examples are the neuromodulation of PV IN synapses, which are potently suppressed by the activation of presynaptic muscarinic cholinergic receptors as well as GABAB receptors (Kruglikov and Rudy, 2008) and through disinhibitory mechanisms where an inhibitory cell type will have a stronger effect onto PV INs than excitatory cells (Chittajallu et al., 2013; Xu et al., 2013) (see Disinhibition section below). Differences in the dynamics of input and output synapses of PCs and PV INs have also been suggested to mediate changes in the balance between excitation and inhibition as a function of input frequency during different brain states (Taub et al., 2013).
Feedforward inhibition has also been shown to participate in gain modulation through input normalization (Pouille et al., 2009). As excitatory drive increases, more and more inhibitory cells are concomitantly recruited as well. This inhibitory tracking of incoming excitatory drive through FFI by PV INs prevents saturation of PC ensembles by progressively increasing the excitatory current necessary to beat the inhibition and reach spiking threshold. While FFI acts homogeneously across pyramidal cells, heterogeneities in the distribution of excitatory currents in the neuronal population determined the specific set of pyramidal cells recruited. As a result of this scheme, the cortex, while being sensitive to weak stimuli also responds to stronger inputs without saturating (Figure 5F).
A circuit remarkably similar in many details to the one mediating thalamocortical communication is involved in CA3 to CA1 communication in the hippocampus (Pouille and Scanziani, 2001). PV FS basket cells mediate FFI in a qualitatively similar manner in many other cortical and subcortical circuits. This seems to be particularly true for ‘ascending pathways’ following a feedforward direction. For example, in sensory cortices, PV INs seem to be the main cell type mediating FFI of the L4 to L2/3 projection in primary somatosensory (Helmstaedter et al., 2008; House et al., 2011; Xu and Callaway, 2009) and visual (Adesnik et al., 2012) cortices and forward interareal cortico-cortical connections, such as primary to secondary visual cortices (Yang et al., 2013) as well as in the cortico-striatal pathway (Mallet et al., 2005) among others.
Non PV IN-mediated FFI
Although we highlighted a strong association of PV INs with feedforward inhibitory circuits, it should be stressed that this is not a universal phenomenon. An interesting example occurs within the olfactory system. In the piriform cortex, the input layer is L1a, which receives the lateral olfactory tract (LOT) afferents containing the axons of projecting mitral and tufted cells of the olfactory bulb. L1a lacks PV cells. Instead, LOT axons synapse on L1a interneurons, including neurogliaform and horizontal INs, that target the apical dendrites of pyramidal cells (Stokes and Isaacson, 2010; Suzuki and Bekkers, 2010, 2012). LOT axons also target the dendrites of the excitatory cells. As a result, similar to what is observed in the neocortex and hippocampus, bursts of mitral and tufted cell activity mediate a short-latency feed-forward disynaptic inhibition of the pyramidal cells, except that this inhibition occurs on the distal apical dendrite. It is interesting to note that in this way, FFI remains spatially matched with the excitation. PV FS cells in the piriform cortex are localized mainly in L3. They are recruited by recurrent excitation from the pyramidal cells. As in other structures, the PV cells in the piriform cortex target the perisomatic domain of the pyramidal cells and their inputs are depressing. However, the excitatory inputs on the pyramidal cells are facilitating. Hence, during bursts of inputs to the piriform cortex, perisomatic-targeting FS cells are recruited late and provide feedback somatic inhibition of the pyramidal cells. Thus, in the olfactory cortex, inhibition shifts from the dendrite to the soma, the opposite of what is observed in the neocortex and hippocampus.
Another example where FFI targets dendritic instead of perisomatic regions of excitatory cells has been found in S1 when studying a phenomenon called interhemispheric inhibition. Sensory evoked cortical activity is diminished if an ipsilateral sensory stimulation occurs during a certain time window before contralateral stimulation. While this phenomenon has been reported in the rodent as well as the human brain, the underlying circuit mechanisms had not been well understood. A study by Palmer et al. (Palmer et al., 2012) suggested that the circuit and cellular basis for interhemispheric inhibition in rodent somatosensory cortex largely comes from callosal fibers strongly recruiting L1 GABAergic interneurons. The activated L1 interneurons, in turn, provide GABAB-mediated inhibition to the distal dendrite of L5 pyramidal neurons thus decreasing stimulus-driven spiking activity from these L5 neurons. In contrast to the fast and temporally precise FFI circuits involving PV FS basket INs, this circuit is slow and long-lasting due to the activation of metabotropic GABAB receptors in dendrites that subsequently open GIRK channels and block voltage-gated Ca2+ channels. Thus, the distal dendritic location and cellular mechanism of L1 INs mediating interhemispheric inhibition block active dendritic integration, indirectly affecting spiking through these dendritic phenomena.
L1 interneurons are also involved in thalamocortical FFI. Higher-order thalamic nuclei show prominent fibers in L1 (Berendse and Groenewegen, 1991; Wimmer et al., 2010). In a study investigating the functional connectivity of the matrix thalamic nuclei on the medial prefrontal cortex (mPFC) (Cruikshank et al., 2012) found that axons from matrix thalamic neurons strongly excite L1 interneurons, particularly LS INs, and that these interneurons, in turn, mediate feedforward inhibition to L2/3 pyramidal neurons.
B. Feedback inhibition
In contrast to the FFI circuit motif, in which the source of excitation of INs originates from incoming external excitatory afferent axons to the local network, in feedback inhibition (FBI) the source of excitation is locally generated and interneurons synapse back to the local PC population (Figure 6). The feedback action from INs then reduces or prevents further discharges of the excitatory cells. Most IN subtypes receive inputs from multiple surrounding PCs and in turn provide inhibitory output to the excitatory cells and therefore are part of an FBI circuit motif. As was the case for FFI, the specific IN type involved in a given FBI loop will show particular recruitment patterns and provide different functional features to the local PCs. However, regardless of the IN involved, this recurrent inhibition controls the excitatory-inhibitory balance of local populations of principal cells and shapes spatial and temporal features of their activity patterns in fundamentally different ways than FFI. While the FFI is an incoming input tracking circuit mechanism and does not depend on local activity level, FBI is the opposite, i.e. a circuit mechanism tracking the local outputs that are being generated.
Figure 6. Feedback inhibition (FBI) and differential effect of PV and Sst INs-mediated inhibition.
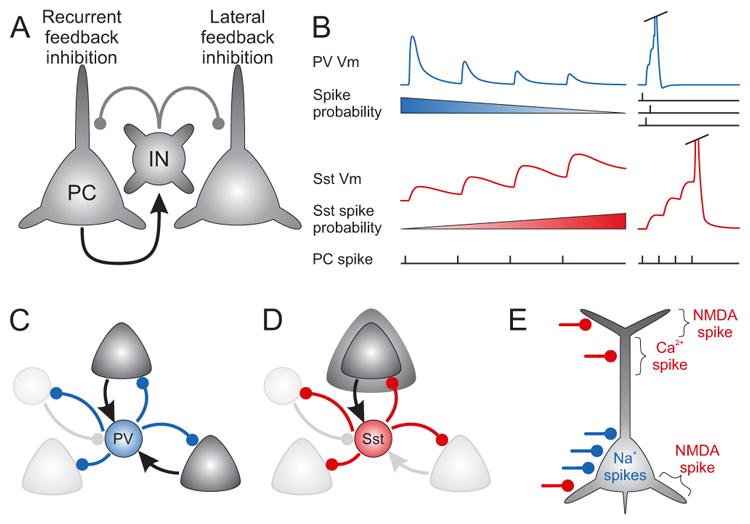
A. FBI circuit motif encompassing both recurrent and lateral feedback inhibition. B. PV and Sst interneurons differentially summate excitatory inputs from local PCs. PV neurons (top), due to their low input resistance, fast membrane time constant and depressing excitatory inputs show a decrease in spike probability upon repetitive excitation (left) and are thus synchrony detectors, requiring the near coincident action of different cells to spike (right) (Silberberg, 2008; Silberberg and Markram, 2007). In contrast, Sst INs (bottom) have a high input resistance, slow membrane time constant and facilitating excitatory inputs and therefore show an increase in spike probability upon repetitive stimulation. This makes Sst neurons sensitive to individual excitatory cell’s firing rate and bursting (Kapfer et al., 2007; Silberberg, 2008; Silberberg and Markram, 2007). C. Assembly competition and synchronization of local PCs by PV neurons. Synchronously active PCs will recruit PV neurons that will then inhibit the local population (Silberberg, 2008).. D. Assembly competition and ‘winner takes all’ circuit mechanism with Sst INs FBI (Silberberg, 2008). By following the most active PC(s), an Sst IN will prevent the activation of other neighboring cells (except other Sst INs). E. Perisomatic inhibition by PV INs regulates the timing of action potentials (Royer et al., 2012). The speed and effectiveness at which PV INs perform this task imposes short temporal windows for excitation to generate action potentials within the local population, thus favoring their synchronization (Cobb et al., 1995). In contrast, dendritic targeting bias of Sst INs regulates input integration at the dendrite and dendritic electrogenesis such as NMDA and calcium spikes that can generate burst firing (Larkum et al., 1999).
Given the divergence of interneuron connectivity, any given IN will inhibit not only PCs from which it received excitation but also others that are part of the local population. This is due to the fact that INs generally show dense local connectivity (Fino and Yuste, 2011; Packer and Yuste, 2011) and that excitatory neuron populations in many cases show sparse activity (Barth and Poulet, 2012). In addition, some cortical INs have axons that extend beyond the local area where their soma is located, which can be in a transcolumnar and/or translaminar fashion (Helmstaedter et al., 2008; Katzel et al., 2011). Thus INs can provide inhibition to neighboring populations of principal cells located at a certain distance that may not have provided excitation to that particular interneuron population, a phenomenon more generally referred to as lateral inhibition (Figure 4,6). In lateral inhibition, as we use the term here, a population of pyramidal cells receives “feedback inhibition” from interneurons that did not received excitation from this population, regardless if the PCs are within or flanking the cells driving the INs providing feedback. These different forms that FBI encompasses have been proposed to participate in important phenomena such as surround suppression (Adesnik et al., 2012), assembly selection and competition (Roux and Buzsaki, 2015), oscillatory coupling (Buzsaki and Wang, 2012), and grid field formation (Couey et al., 2013).
While it is likely that most IN subtypes participate in local feedback loops, there are qualitative and quantitative differences in the connectivity between any IN subtype and the local PC population. Not only the presence of an excitatory-inhibitory feedback loop but also its strength are important to weight its significance. There are multiple examples of recurrent inhibitory loops in cortical circuits. Here we will focus on feedback inhibitory circuits that have been described for Sst and PV INs in the neocortex. Although impairment of PV and Sst INs have been implicated in the generation of hypersynchronous activity and epilepsy (Goldberg and Coulter, 2013; Hunt et al., 2013; Ito-Ishida et al., 2015; Paz and Huguenard, 2015), there are differences in the recruitment and functional impact of these circuits that reflect the specialized properties of these two types of interneurons.
Differential recruitment of PV and Sst INs by local excitatory networks
The differences in synaptic dynamics as well as intrinsic properties of PV and Sst INs have interesting functional consequences. Because excitatory inputs onto PV cells are initially strong and depressing and those onto Sst cells initially weak and facilitating, trains of excitatory spikes targeting both cell types will lead to contrasting patterns of recruitment. The probability of firing of PV cells is initially high and goes down while Sst INs will show the opposite trend and will be recruited more slowly, with a delay following synaptic facilitation and EPSP summation (Kapfer et al., 2007; Pouille and Scanziani, 2004; Silberberg and Markram, 2007) (Figure 6B). Consistent with this, Pouille and Scanziani (2004) observed in simultaneous recordings from the soma and the dendrite that during a train of excitatory stimuli in the hippocampus, inhibition first targeted the soma of PCs where PV cell synapses are located and progressively shifted distally, where Sst IN synapses are located. This delayed pathway was found to be frequency-dependent and has been observed in multiple neocortical circuits (Berger et al., 2010; Kapfer et al., 2007; Silberberg and Markram, 2007). Therefore, Sst neurons will be sensitive to firing rate increases from even a small number of cells, regardless of their fine temporal relationship. In fact, it has been found that a high frequency train from even a single pyramidal cell can be sufficient to recruit Sst INs (Kapfer et al., 2007; Kwan and Dan, 2012; Silberberg and Markram, 2007). In contrast, PV neurons are unlikely to respond to the activity of a single or very few cells because the EPSP from a single PC is too small to reach threshold and the FS IN is less capable of summating consecutive EPSPs due to the their fast kinetics (Box 1). Instead, PV INs will be more sensitive to spikes distributed among different cells but occurring in a short time window. Thus, Sst INs act as burst and rate detectors, following the most active excitatory cells in the local network, while PV INs follow the different pockets of synchrony in the overall population (Kwan and Dan, 2012) (Figure 6C and D). Interestingly, these different modes of operation are also reflected in the subcellular target of these IN types. Due to their perisomatic bias, PV synaptic contacts are well suited to control the timing of individual soduim spikes while dendritic-biased Sst neuron synapses gate dendritic integration, calcium spikes and burst generation (Gentet et al., 2012; Murayama et al., 2009; Royer et al., 2012) (Figure 6E).
The disynaptic inhibition produced by Sst and PV INs can affect a substantial fraction of neighboring PCs as a result of the much higher connection probability between INs and PCs than among PCs (Fino and Yuste, 2011). The different modes of recruitment of these two IN types imply different functional consequences. The coincidence detection property of PV neurons has led to the view that they would be involved in the temporal segmentation of different cell assemblies and moment-to-moment adjustment of the inhibitory tone that is proportional to the local level of synchrony (Buzsaki and Wang, 2012). In contrast, the Sst IN-mediated disynaptic inhibition among pyramidal cells might function as a mechanism preventing runaway excitation by applying an inhibitory tone that is proportional to the magnitude of firing rates of the PC population and operates over a longer time scale. In addition, given that the most active cells are the ones dictating the inhibitory tone provided by Sst INs, this circuit can also function as a ‘winner takes all’ mechanism of assembly competition (Silberberg, 2008), whereby the pyramidal cells that fire most strongly at a given moment (e.g. following presentation of optimal stimulus or because their synapses were strengthened by previous activity) can suppress the activity of neighboring PCs.
Recently, the Sst interneuron-mediated feedback and lateral inhibition has been proposed as a candidate mechanism for surround suppression in visual cortex (Adesnik et al., 2012). Because Sst INs do not inhibit each other, but inhibit PV and PC neurons (Table I), when the size of a visual stimulus is increased the response magnitude and the number of Sst INs recruited in the superficial layers of mouse V1 continues to increase while the response of pyramidal cells and PV INs decreased due to the inhibition provided by the Sst INs.
C. Disinhibition
It has been appreciated for quite some time that neocortical GABAergic interneurons not only provide direct inhibition by targeting principal cells, but can also synapse onto other inhibitory neurons and consequently have a disinhibitory effect (Freund et al., 1983; Somogyi et al., 1983). In fact, it has been proposed that neocortical “disinhibition of cell assemblies (facilitation) could be at least as powerful as direct inhibition” (Kisvarday et al., 1993). However, early studies revealing interconnections between inhibitory interneurons were essentially based on anatomical observations, rendering the functional implications hard to assess. More recently, there has been a reappraisal of disinhibitory actions using advanced electrophysiological, imaging and molecular genetics technologies. These new studies have not only confirmed and refined the circuits drawn from earlier anatomical studies, but also helped in establishing causal links through targeted manipulations. Moreover, studies from behaving animals have started to shed light on how and when various disinhibitory circuits are in action and, importantly, what are the possible functional implications of these circuits.
Observing an increase in firing rate of excitatory cells and/or decrease spiking of inhibitory neurons can simply be due to a rerouting of excitatory drive alone without the involvement of an interneuron-to-interneuron connection. Here, disinhibition is considered as the principle by which the main function of an interneuron type in a given context is to inhibit another GABAergic type more potently than principal cells. This is an important distinction since INs are highly interconnected with many other cell types to various degrees (Table I), and therefore the mere presence of an interneuron-to-interneuron connection does not imply disinhibition, as defined above. For example, although PV basket interneurons interconnect with other PV INs and other inhibitory subtypes, their strong connectivity to principal cells makes it unclear whether there is any context in which disinhibition can be one of their primary functions. On the other hand, such connectivity scheme can result in synchronizing a highly interconnected population on the short time scale or imposing a veto as to which interneuron subtypes will provide inhibition to principal cells. Another example concerns Sst INs: since this group shows dense connectivity to other IN groups but little interconnectivity to other Sst INs, it is conceivable that in a context where the Sst IN population is highly active, they would contribute to reducing the inhibitory tone of other IN types onto pyramidal cells while promoting theirs.
Disinhibitory effect of neocortical VIP neurons
The first studies to show the existence of cortical interneurons preferentially innervating other GABAergic neurons were done in the hippocampus (Acsady et al., 1996; Gulyas et al., 1996; Hajos et al., 1996). These studies found that interneuron-selective INs included CR+ VIP-expressing cells. It was also observed that they often made contacts with putative dendritic targeting INs. Such observations were later extended to the neocortex, where CR-expressing neurons were found to often preferentially make synapses onto other non-pyramidal cells in L2/3, particularly other CR-expressing (Caputi et al., 2009; Defelipe et al., 1999; Gonchar and Burkhalter, 1999) or calbindin-expressing cells (Defelipe et al., 1999), which comprise subsets of both Sst and PV interneurons. Work investigating the postsynaptic targets of VIP cells showed that these neurons contacted the perisomatic region of PV INs (David et al., 2007; Hioki et al., 2013) as well as calbindin INs (Staiger et al., 2004). These results brought authors to suggest that as in the hippocampus, a specific neocortical interneuron subtype could be specialized in providing disinhibition (Gonchar and Burkhalter, 1999; Staiger et al., 2004).
Using slice recording from a mouse line expressing eGFP under the calretinin promoter, Caputi and colleagues (Caputi et al., 2009) found that two types of L2/3 CR-expressing cells, bursting bipolar and adapting multipolar cells, had different connectivity and synaptic short-term dynamics. Bipolar CR neurons had much higher connection probability to multipolar CR neurons than to any other neuronal type tested, seconded by other interneuronal types (including other bipolar CR cells) and pyramidal cells coming last. Combined with the insights of more recent studies (see below) and morphological associations with different markers (see Diversity section), it is interesting to note that the properties of the bipolar CR cells described by Caputi et al. (Caputi et al., 2009)resemble those of VIP neurons (Bayraktar et al., 2000), while the electrophysiological properties of multipolar CR cells are reminiscent of Sst neurons (adapting firing pattern with facilitating excitatory inputs and low interconnectivity, two IN types known to contain subsets co-expressing CR (see Table I).
More recently, thanks to the development of novel Cre mouse lines enabling the expression of ChR2 in GABAergic neuronal subpopulations (Taniguchi et al., 2011), studies using electrophysiological slice recordings and thorough cell type comparison found that VIP neurons had a striking preference to target dendritic targeting Sst interneurons in L2/3 of S1, V1, A1 and prefrontal cortices (Lee et al., 2013; Pfeffer et al., 2013; Pi et al., 2013). It therefore seems that the VIP to Sst disinhibitory connection is a general principle in the superficial layers of neocortex.
If VIP interneurons preferentially inhibit Sst cells, when and what drives VIP INs firing and what is the impact of such disinhibitory circuit? An important source of excitation of VIP INs, at least in sensory systems, is from cortico-cortical feedback projections from higher order cortices. Lee et al. (Lee et al., 2013) observed that VIP INs in barrel cortex received much stronger inputs from motor cortex than any other cell type (PC, PV, Sst and 5HT3aR non-VIP). In addition, these neurons were easily driven to spike by the stimulus, likely due in part to the large input resistance of VIP cells compared to other interneurons (Table I). These observations predicted that when M1 inputs activate VIP cells, Sst interneurons will be inhibited, and hence the dendrites of the pyramidal cells will be disinhibited (Figure 7). Consistent with this, inhibitory responses were observed on Sst neurons following stimulation of glutamatergic axons from motor cortex in S1. Moreover, inactivating the VIP neurons significantly reduced the inhibitory responses on Sst cells. In vivo observations provided evidence that this VIP-mediated disinhibitory circuit operates under physiological conditions. Whole cell recordings of different L2/3 populations of GABAergic neurons in the barrel cortex of awake mice showed that unidentified non-FS neurons were highly active during periods of whisking, in the absence of sensory stimulation (Gentet et al., 2010; Gentet et al., 2012). Conversely, Sst neurons showed the opposite effect, i.e. active during quiescence and strongly inhibited during whisker movement (Gentet et al., 2012). Lee et al., (Lee et al., 2013) found that the VIP cells were the non-FS showing elevated activity during periods of whisking, while Sst INs were anticorrelated as previously observed. Importantly, inactivation of motor cortex reduced these effects (Lee et al., 2013). These experiments strongly suggest that VIP neurons can be recruited by motor cortex, likely through a direct glutamatergic projection, and in turn inhibit Sst INs. Given the slightly facilitating synaptic dynamics of the output synapses of CR+ VIP cells, (the BCR neurons in (Caputi et al., 2009) (see Diversity section) repetitive firing of these neurons will tend to produce sustained inhibition of SST neurons.
Figure 7. VIP interneuron-mediated disinhibition.

A. VIP INs show high target selectivity for Sst INs which results in disinhibition of the dendrites of pyramidal cells when VIP INs are active (Jiang et al., 2015; Pfeffer et al., 2013; Pi et al., 2013). B. Widespread activation of this disinhibitory circuit could have a broad influence on the state of excitability of an area and is thought to be mediated by neuromodulators (see text). C. Given the narrow and vertical extent of VIP IN dendrites and axons (Pronneke et al., 2015), their recruitment by excitatory inputs showing topographic organization can mediate spatially localized sites of disinhibition, as in (Zhang et al., 2014). Such inputs are thought to originate from glutamatergic afferents from the thalamus and top-down corticocortical projections (see text).
Another recent study has shown that excitatory projections from the cingulate cortex (Cg) strongly recruit VIP INs in V1 (Zhang et al., 2014). Analogous to the M1 to S1 projection target preference for VIP cells, VIP neurons in V1 receive the strongest input from Cg when compared to other L2/3 INs and PCs. Moreover, activating or inactivating Cg axons during a visual discrimination task improved or reduced behavioral performance, respectively. This study also provided evidence that the circuitry involved was permissive for the effects of VIP IN recruitment to act in a spatially restricted manner, consistent with the narrow axonal distribution of VIP INs (see Diversity section), since Cg neurons show some form of topography in V1. The studies by Lee et al. (Lee et al., 2013) and Zhang et al. (Lee et al., 2013) demonstrate a role of VIP neurons in top-down modulation by being a major target of feedback cortico-cortical projections. This adds to the anatomical observation that L1 CR dendrites (a major proportion of which are dendrites of VIP INs) in V1 are a primary target of feedback from secondary visual area (Gonchar and Burkhalter, 2003), suggesting that this could be a common circuit motif for cortico-cortical interaction.
Subcortical neuromodulatory systems appear to provide another source of excitation to VIP INs. Studying the effect of locomotion (running) on gain modulation in V1, it was observed that the activity of VIP cells was tightly coupled to the animal’s locomotor activity (Fu et al., 2014). This is consistent with the observations in barrel cortex regarding the relation of VIP cells and motor activity (whisker movement). However, it was found that locally blocking glutamatergic neurontransmission did not affect the running-related neuronal activity of VIP cells. Instead, blocking nicotinic receptors significantly reduced the correlation of the activity of VIP INs and running. This result suggests that the basal forebrain cholinergic system has an important role in the recruitment of VIP neurons during locomotion, consistent with the observation that VIP INs, like all 5HT3aR neurons are strongly depolarized by nicotinic agonists (Lee et al., 2010) see Interneuron Diversity section, Table I). Indeed, rabies virus experiments showed that the basal forebrain is a potentially major source of inputs to V1 VIP neurons (Fu et al., 2014). Finally, Fu et al. (Fu et al., 2014)showed that the increased firing of VIP neurons during running was a broad phenomenon since it was also observed in somatosensory and auditory cortices, pointing toward a more global behavioral state change when the animal is engaged in locomotion. Reinforcement signals, particularly aversive signals, are also effective at recruiting the VIP disinhibitory circuit in both auditory and prefrontal cortices (Pi et al., 2013). This global effect is also likely to involve subcortical neuromodulatory systems.
At least in sensory cortices, VIP neurons can also be recruited by bottom-up sensory signals. Anatomical observations in somatosensory and visual cortices have shown that the sensory thalamus makes synaptic contact with VIP neurons (Hajos et al., 1996; Staiger et al., 1996). Consistent with this, monosynaptic rabies virus experiments suggested that VIP neurons in V1 receive direct inputs from LGN (Fu et al., 2014). Moreover, despite the thalamus driven synaptic currents on VIP neurons being much smaller than those on fast spiking PV interneurons, the high input resistance of VIP cells makes them significantly excitable by thalamic stimulation (Lee et al., 2010). Therefore, the circuit elements exist for VIP neurons to mediate a form of sensory driven feed-forward disinhibition. It has also been shown that VIP neurons do show sensory responses (Fu et al., 2014; Kerlin et al., 2010; Pi et al., 2013). Although these results do not imply direct thalamic recruitment, it does show that VIP cells are recruited by afferent sensory signals. Moreover, increased activity of VIP neurons during sensory stimulation is correlated with the increased gain of PCs (Pi et al., 2013), which has also been shown during natural behavior (Fu et al., 2014). These latter studies suggest that recruitment of VIP neurons by, or at least during, sensory inputs might play an important role in sensory information processing.
These studies indicate that VIP neurons could potentially act in both spatially specific and global manners, depending on the input source (Figure 7). While glutamatergic thalamocortical feedforward and cortico-cortical feedback inputs could support spatial specificity in their recruitment of VIP neurons, neuromodulatory systems seem more likely to act globally, at least during certain behavioral states. Moreover, the experiments by Fu et al. (Fu et al., 2014) showing that nicotinic receptor blockade produced only partial ablation of VIP cells’ activity suggest that another neuromodulatory system is also involved. Experiments involving a broad range of behavioral contingencies and careful circuit dissection will add to our understanding of the various actions of disinhibitory control through VIP neurons.
Other disinhibitory circuits
Evidence for other disinhibitory circuits in neocortex has also come to light in recent years. These examples suggest that disinhibition might be found more widely than what has been appreciated. Some of them also illustrate the possibility that interneuron types that are often thought to produce mainly inhibition of principal cells may in some specific cases produce predominantly disinhibition.
A Sst IN-mediated disinhibitory circuit in L4
As described in the Interneuron Diversity section, Sst INs in L4 of S1 differ substantially in morphology and electrophysiological properties from those in supra and infragranular layers (Ma et al., 2006; Xu et al., 2013). Furthermore, L4 Sst INs differ from Sst INs in other layers in their connectivity (Xu et al., 2013). In L2/3, connection probability and synaptic strength are larger for principal cells, but in L4, both, the connection probability and the synaptic strength of Sst INs are much larger on PV INs (Beierlein et al., 2003; Gibson et al., 1999; Xu et al., 2013). This suggests that activity of Sst INs in L4 will produce disinhibition of the principal cells by inhibiting FS PV INs which tightly control the activity of the principal cells. In support of this, Xu et al., (Xu et al., 2013) found that optogenetic inhibition of Sst INs in an active cortical network in slices increased firing of L2/3 pyramidal cells but it decreased the firing of L4 PCs. More work will be needed to uncover whether this disinhibitory circuit operates in vivo, but understanding the function of this circuit will require knowledge of what determines the activity of Sst INs in this layer. Xu et al. (Xu et al., 2013) suggested that the Sst INs in L4 might be activated physiologically by ACh via the well established muscarinic-mediated depolarization of Sst neurons (see Table I; Interneuron Diversity). Hence this circuit potentially contributes to the cholinergic mechanisms regulating sensory processing. In addition to ACh, high frequency local excitatory inputs also strongly recruit Sst INs due to the frequency-dependent facilitation of excitatory inputs unto these cells..
NGFC-mediated disinhibition in L4
Chittajallu et al. (Chittajallu et al., 2013) suggested a disinhibitory role of neurogliaform cells in L4 of somatosensory cortex. They observed that spiking of a L4 NGFC in slices reduces the thalamic evoked FS cell-mediated FFI onto a connected PC by a GABAB mechanism reducing GABA release from presynaptic FS PV cell terminals. However, NGFC activity did not affect monosynaptic thalamic feed forward excitation onto the same PC. The inhibition of FFI resulted in an increase in the temporal window for integration of excitatory thalamic input in spiny stellate cells (Figure 5D), as previously observed for the GABAB and cholinergic inhibition of GABA release from FS cells (Kruglikov and Rudy, 2008). These results are surprising given the notion that NGFCs produce volume release of GABA resulting in spatially unrestricted inhibition lacking target specificity, and the fact that the GABAB agonist baclofen inhibits neurotransmission of both FS and thalamocortical synapses on to the spiny stellate cells equally (Chittajallu et al., 2013; Kruglikov and Rudy, 2008). Chittajallu et al. (Chittajallu et al., 2013) speculate that a non-uniform distribution of release sites of NGFCs axons or other structural differences between thalamocortical and FS cell synapses may explain the selectivity. In any case, the results raise the possibility that in some structures NGFC-mediated inhibition can have more spatially restricted influence than what might be expected from volume transmission. In fact NGFCs have been observed to also make synaptic contacts with subcellular structures such as dendritic spines (Kawaguchi and Kubota, 1997; Tamas et al., 2003).
Disinhibitory circuits mediated by layer 1 interneurons
Letskus et al. (Letzkus et al., 2011) proposed the hypothesis that a disinhibitory circuit mediates associative fear learning in auditory cortex. They found that a conditioning foot shock produced nicotinic activation of a yet to be identified subtype of L1 interneuron that inhibited L2/3 PV INs resulting in disinhibition of the pyramidal neurons. Pharmacological or optogenetic block of the disinhibition abolished fear learning. Interestingly, Jiang et al. (Jiang et al., 2013) found that besides NGFCs, L1 had another large subpopulation of neurons identified as “single bouquet cells” (SBC), having a descending axon to L2/3 (see Table I). These INs preferentially formed unidirectional inhibitory connections on all subtypes of L2/3 interneurons and can thus produce disinhibition of pyramidal cells. These SBCs could the mediators of the disinhibition described by Letskus et al. (Letzkus et al., 2011). The SBCs described by Jiang et al. (Jiang et al., 2013) resemble VIP INs, raising the possibility that the fear-conditioning-related disinhibition might be another example of VIP-mediated disinhibition. This would be consistent with the observation that VIP neurons respond to aversive stimuli (Pi et al., 2013). However, other evidence suggests that SBC and VIP INs might be different cell types. While SBCs appear to be a prominent L1 interneuron type (Jiang et al., 2013), VIP INs only account for about 5% of L1 INs (see Diversity section). The output connectivity of the two cell types also differs. SBCs connect to all IN subtypes in L2/3 (but not to PCs)(Jiang et al., 2013). In addition, the connection probability of SBCs to Sst INs is the lowest among all INs. This contrast with VIP INs, which have low connectivity to all neurons including pyramidal cells but exhibit substantially stronger connectivity to Sst INs (Lee et al., 2013; Pfeffer et al., 2013; Pi et al., 2013). Although this needs to be address directly, these differences suggest that SBCs and VIP INs belong to two distinct disinhibitory circuits.
IV. GABAergic interneurons and cortical function
The ability to target, monitor and manipulate specific IN subtypes in vivo by combining modern molecular genetic tools, imaging and electrophysiological technologies have resulted in renewed efforts to understand the contribution of these cells to network dynamics, cortical computations, cognitive processes and behavior. Extracellular recordings in the anterior cingulate cortex of mice performing a reward foraging task showed that a subgroup of optogenetically-identified Sst INs with narrow spikes fired preferentially at reward approach, whereas PV INs showed an increase firing when the mouse was leaving the reward location that was correlated with stay duration in the reward zone (Kvitsiani et al., 2013). While studies such as this nicely illustrate the idea that different INs have specific functional contributions in a given context, we still lack an understanding of how their activity contributes to the computations underlying behavior. One area that has received significant attention is the function of interneurons in the generation of cortical oscillations. A large body of experimental and computer modeling work has suggested essential roles of GABAergic neurons in the generation of cortical fast oscillations (reviewed in (Allen and Monyer, 2014; Buzsaki and Wang, 2012)). Modern optogenetic methods allowing a more direct testing of the mechanisms, have resulted in strong evidence supporting the view that PV-expressing basket cells have critical roles in the generation and maintenance of gamma, theta and ripple oscillations (Cardin et al., 2009; Sohal et al., 2009; Stark et al., 2013; Stark et al., 2014). These rhythms, and the circuits underlying their generation illustrate the importance of the fast signaling properties of FS PV basket cells (Bartos et al., 2007).
Interneurons and sensory feature selectivity
Given the importance of specific cortical interneurons in sculpting excitatory cell responses, it is natural to address whether and how they are involved in the feature selectivity characteristic of excitatory cells of primary sensory cortices. There is a growing consensus that IN subtypes differentially contribute to the establishment or modulation of stimulus selectivity. Yet what their precise role might be is still actively debated. To understand this, many layers of information are required. How do different types of GABAergic neurons respond to specific sensory stimulation? How do their biophysical, morphological, and synaptic properties and connectivity contribute to their recruitment? How does the spatiotemporal profile of inhibition provided by these neurons in turn helps shape the sensory responses of excitatory cells?
In most early studies of sensory responses of cortical neurons, cell types were identified based on their spike waveform, thus these data are largely limited to putative fast-spiking neurons (Simons, 1978), based on the assumption that brief spikes belong to these neurons. Nevertheless, using spike shape analysis, genetic labeling or post hoc immunochemical identification of interneurons revealed that in mouse visual cortex, selectivity to visual stimuli such as orientation, direction, and spatial frequency are broader for FS neurons than for pyramidal cells (Atallah et al., 2012; Hofer et al., 2011; Kerlin et al., 2010; Liu et al., 2010; Runyan et al., 2010; Sohya et al., 2007). Although the data is limited, other IN groups such as Sst and VIP INs have also been reported to be less selective to sensory stimulation (Kerlin et al., 2010). It appears that for the most part cortical INs are more broadly tuned than nearby PCs, yet different INs show various degrees of sensory tuning. VIP INs, for example, tend to be more selective for orientation tuning than PV and Sst INs (Kerlin et al., 2010). It has also been reported that a few INs within a molecularly defined group show strong stimulus selectivity that is similar to that of pyramidal neurons (Ma et al., 2010; Runyan et al., 2010). For example, a subset of PV INs in superficial layers of mouse V1 responds highly selectively to visual orientation (Runyan et al., 2010). Whether these PV neurons are a subpopulation within the PV neuron group is unclear. However, the orientation selectivity of PV neurons was later shown to be inversely correlated with the extent of their dendritic arbor, suggesting that tuning of PV INs could be simply related to the spatial extent to which they can sample excitatory inputs (Runyan and Sur, 2013). If this is the case, it becomes tempting to speculate that this could in part explain the higher selectivity observed in VIP INs, which generally show narrow bipolar dendritic arbor. Similar to visual cortex, GABAergic interneurons in other sensory cortices are also broadly tuned. In somatosensory cortex, layer 4 FS PV INs show broad tuning to angular direction of whisker movement and multiwhisker receptive fields (Bruno and Simons, 2002; Lee and Simons, 2004).
What factors define tuning properties of INs and why are inhibitory neurons generally more broadly tuned? Neurons in cortex are organized in a columnar structure, such that groups of neurons within a column share similar functional properties. Similar to the excitatory neurons within a column, it is likely that interneurons also demonstrate similar sensory selectivity that is shared within a column. Rodent visual cortex, however, lacks orientation columns, thus the sum of excitatory inputs from nearby pyramidal neurons to interneurons will be broad (Hofer et al., 2011). By contrast, cat visual cortex is organized as a pinwheel-like columnar structure for orientation selectivity. FS neurons in cat visual cortex do respond more selectively to orientation stimulus reflecting similarly tuned local population (Azouz et al., 1997; Cardin et al., 2007; Martinez et al., 2005; Nowak et al., 2008). Mouse auditory cortex is tonotopically organized. PV-positive neurons in auditory cortex are tuned to sound frequency to a similar degree as nearby PV-negative neurons (Moore and Wehr, 2013). Therefore, feature selectivity would simply be a reflection of input availability within a cell’s dendritic domain (Runyan and Sur, 2013). Other factors such as intrinsic biophysical and synaptic properties of interneurons are also likely to contribute. Intracellular recording from INs, mostly FS INs, revealed that the selectivity of sensory-evoked EPSPs to FS neurons is comparable to those to excitatory neurons (Cardin et al., 2007; Hirsch et al., 2003), suggesting that they receive roughly similar inputs. However, the faster time constant of FS neurons and stronger excitatory inputs allows for larger and faster membrane potential changes, and consequently FS neurons can reach threshold more often leading to spiking activity for non-preferred stimuli and hence broader tuning (Cardin et al., 2007). Excitatory inputs to FS INs from thalamus have also been shown to be stronger (Cruikshank et al., 2007; Gabernet et al., 2005) and exhibiting more convergence (Bruno and Simons, 2002; Swadlow and Gusev, 2002) onto FS than PCs. Altogether, network architecture, dendritic morphology, connectivity, synaptic profiles and biophysical properties all seem to contribute to determining the selectivity of FS INs. More work will be required to understand the responses of other inhibitory cell types, which differ at all these levels.
What is the role of these different IN types in establishing sensory selectivity of PCs? Pharmacological blockade of GABAergic inhibition broadened orientation selectivity in visual cortex demonstrating the importance of inhibition in shaping sensory selectivity (Sillito, 1975; Tsumoto et al., 1979). In other studies, however, blockage of inhibition was found to change response magnitude yet had minor effect on tuning properties (Katzner et al., 2011; Nelson et al., 1994; Ozeki et al., 2004), suggesting sensory selectivity is inherited from thalamic inputs. Pharmacological approaches could not address the specific contribution of IN subtypes in shaping responses of excitatory neurons. Using optogenetics, several recent studies have investigated the contribution of PV and Sst neurons to stimulus selectivity in the mouse visual cortex (Atallah et al., 2012; El-Boustani and Sur, 2014; Lee et al., 2012; Wilson et al., 2012). These studies demonstrated that PV and Sst inhibitory neurons play distinct roles in the tuning of excitatory neurons producing either divisive or subtractive inhibition. However, the results from these studies do not agree on the specific role of each interneuron group. Some studies found that Sst INs sharpen orientation tuning (Wilson et al., 2012), while activation of PV INs was found to have a minor effect on tuning but changed response gain (Atallah et al., 2012; El-Boustani and Sur, 2014; Wilson et al., 2012). Others reported that PV INs sharpened tuning but not Sst INs (Lee et al., 2012). With follow-up studies, these authors agreed that most of the discrepancy was due to the protocol used to stimulate the interneurons (Lee et al., 2014c). This observation implies that the suppressive function of different interneuron groups can be flexible depending on how these neurons are engaged by visual stimulation and by brain states. Indeed, the study by El-Boustani and Sur (El-Boustani and Sur, 2014) provides compelling evidence that whether inhibition is subtractive or divisive is not a fixed property, but rather a dynamic function that depends on the response modes of INs and the nature of visual stimuli. As we discussed earlier, the activity pattern and function of interneurons can vary greatly depending on the context.
V. Perspectives
It is clear from the above account that there have been significant advances in the last ten years in the understanding of the diversity of interneurons in the neocortex in rodents. There has also been significant progress in discovering cortical circuits involving interneurons and in demonstrating that IN subtypes have specific patterns of activity vis a vis patterns of network dynamics, distinct brain states and behavioral epochs, allowing a better understanding of the functional roles of specific GABAergic INs.
Key to this progress has been the identification of non-overlapping molecular markers defining discrete IN groups together with the use of modern molecular genetic methods. This has allowed the labeling and manipulation of specific INs, permitting recording the activity of IN subtypes and perturbing their activity to assess their functional impact. However, as discussed earlier, the interneuron groups defined by the molecular markers used today are still heterogeneous. It is necessary for further progress to develop methods to study more discrete populations (subtypes of PV, Sst, VIP and non-VIP 5HT3aR INs). While it is conceivable that markers of increased specificity will be discovered, intersectional genetics, in which genetic manipulation of a cell population depends on the intersection of two genetic drivers, is a promising approach that could allow dissection of subpopulations of INs belonging to one of the presently defined molecular groups (He et al., 2016).
Developing approaches to understand IN function in species other than rodents is also a major challenge for the future. The genetic methods that have been so useful to study interneurons in rodents are more difficult to apply in other species, particularly non-human primates, but are not impossible, and developing methods that take advantage of the knowledge gained in rodents, is a challenge for the future.
Recently, Dimidschstein et al., (2016) generated recombinant adeno-associated virus (rAAV) expressing reporters under control of the Dlx5/6 gene and showed that these viruses can be used to restrict gene expression to interneurons of multiple species, including human. This study exemplifies how molecular knowledge might be applied to label, manipulate and study genetically defined neuronal types in species less amenable to traditional genetic manipulation.
Ultimately, understanding the functional role of interneurons will depend on advances in our understanding of the cortical operations mediating specific behaviors.
Acknowledgments
We thank all the members of the Rudy lab for helpful discussions during the writing of the manuscript. This work was supported by NIH grants R01S30989 and P01NS074972 to B.R.
Footnotes
The abbreviations recommended by the HGNC for the genes encoding these three molecules are Pvalb, Sst and Htr3a. We use in this review the abbreviations most commonly used in the interneuron literature.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Acsady L, Gorcs TJ, Freund TF. Different populations of vasoactive intestinal polypeptide-immunoreactive interneurons are specialized to control pyramidal cells or interneurons in the hippocampus. Neuroscience. 1996;73:317–334. doi: 10.1016/0306-4522(95)00609-5. [DOI] [PubMed] [Google Scholar]
- Acsady L, Halasy K, Freund TF. Calretinin is present in non-pyramidal cells of the rat hippocampus--III. Their inputs from the median raphe and medial septal nuclei. Neuroscience. 1993;52:829–841. doi: 10.1016/0306-4522(93)90532-k. [DOI] [PubMed] [Google Scholar]
- Acsady L, Katona I, Martinez-Guijarro FJ, Buzsaki G, Freund TF. Unusual target selectivity of perisomatic inhibitory cells in the hilar region of the rat hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2000;20:6907–6919. doi: 10.1523/JNEUROSCI.20-18-06907.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adesnik H, Bruns W, Taniguchi H, Huang ZJ, Scanziani M. A neural circuit for spatial summation in visual cortex. Nature. 2012;490:226–231. doi: 10.1038/nature11526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali AB, Thomson AM. Synaptic alpha 5 subunit-containing GABAA receptors mediate IPSPs elicited by dendrite-preferring cells in rat neocortex. Cerebral cortex. 2008;18:1260–1271. doi: 10.1093/cercor/bhm160. [DOI] [PubMed] [Google Scholar]
- Allen K, Monyer H. Interneuron control of hippocampal oscillations. Current opinion in neurobiology. 2014;31c:81–87. doi: 10.1016/j.conb.2014.08.016. [DOI] [PubMed] [Google Scholar]
- Alonso A, Kohler C. Evidence for separate projections of hippocampal pyramidal and non-pyramidal neurons to different parts of the septum in the rat brain. Neuroscience letters. 1982;31:209–214. doi: 10.1016/0304-3940(82)90021-0. [DOI] [PubMed] [Google Scholar]
- Alonso JM, Swadlow HA. Thalamocortical specificity and the synthesis of sensory cortical receptive fields. Journal of neurophysiology. 2005;94:26–32. doi: 10.1152/jn.01281.2004. [DOI] [PubMed] [Google Scholar]
- Amitai Y, Gibson JR, Beierlein M, Patrick SL, Ho AM, Connors BW, Golomb D. The spatial dimensions of electrically coupled networks of interneurons in the neocortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2002;22:4142–4152. doi: 10.1523/JNEUROSCI.22-10-04142.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arroyo S, Bennett C, Aziz D, Brown SP, Hestrin S. Prolonged disynaptic inhibition in the cortex mediated by slow, non-alpha7 nicotinic excitation of a specific subset of cortical interneurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2012;32:3859–3864. doi: 10.1523/JNEUROSCI.0115-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ascoli GA, Alonso-Nanclares L, Anderson SA, Barrionuevo G, Benavides-Piccione R, Burkhalter A, Buzsaki G, Cauli B, Defelipe J, Fairen A, et al. Petilla terminology: nomenclature of features of GABAergic interneurons of the cerebral cortex. Nature reviews Neuroscience. 2008;9:557–568. doi: 10.1038/nrn2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atallah BV, Bruns W, Carandini M, Scanziani M. Parvalbumin-expressing interneurons linearly transform cortical responses to visual stimuli. Neuron. 2012;73:159–170. doi: 10.1016/j.neuron.2011.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azouz R, Gray CM, Nowak LG, McCormick DA. Physiological properties of inhibitory interneurons in cat striate cortex. Cerebral cortex. 1997;7:534–545. doi: 10.1093/cercor/7.6.534. [DOI] [PubMed] [Google Scholar]
- Bagnall MW, Hull C, Bushong EA, Ellisman MH, Scanziani M. Multiple clusters of release sites formed by individual thalamic afferents onto cortical interneurons ensure reliable transmission. Neuron. 2011;71:180–194. doi: 10.1016/j.neuron.2011.05.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baraban SC, Tallent MK. Interneuron Diversity series: Interneuronal neuropeptides--endogenous regulators of neuronal excitability. Trends in neurosciences. 2004;27:135–142. doi: 10.1016/j.tins.2004.01.008. [DOI] [PubMed] [Google Scholar]
- Barth AL, Poulet JF. Experimental evidence for sparse firing in the neocortex. Trends in neurosciences. 2012;35:345–355. doi: 10.1016/j.tins.2012.03.008. [DOI] [PubMed] [Google Scholar]
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nature reviews Neuroscience. 2007;8:45–56. doi: 10.1038/nrn2044. [DOI] [PubMed] [Google Scholar]
- Basu J, Zaremba JD, Cheung SK, Hitti FL, Zemelman BV, Losonczy A, Siegelbaum SA. Gating of hippocampal activity, plasticity, and memory by entorhinal cortex long-range inhibition. Science (New York, NY) 2016;351:aaa5694. doi: 10.1126/science.aaa5694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayraktar T, Welker E, Freund TF, Zilles K, Staiger JF. Neurons immunoreactive for vasoactive intestinal polypeptide in the rat primary somatosensory cortex: morphology and spatial relationship to barrel-related columns. The Journal of comparative neurology. 2000;420:291–304. [PubMed] [Google Scholar]
- Beierlein M, Gibson JR, Connors BW. A network of electrically coupled interneurons drives synchronized inhibition in neocortex. Nature neuroscience. 2000;3:904–910. doi: 10.1038/78809. [DOI] [PubMed] [Google Scholar]
- Beierlein M, Gibson JR, Connors BW. Two dynamically distinct inhibitory networks in layer 4 of the neocortex. Journal of neurophysiology. 2003;90:2987–3000. doi: 10.1152/jn.00283.2003. [DOI] [PubMed] [Google Scholar]
- Berendse HW, Groenewegen HJ. Restricted cortical termination fields of the midline and intralaminar thalamic nuclei in the rat. Neuroscience. 1991;42:73–102. doi: 10.1016/0306-4522(91)90151-d. [DOI] [PubMed] [Google Scholar]
- Berger TK, Silberberg G, Perin R, Markram H. Brief bursts self-inhibit and correlate the pyramidal network. PLoS biology. 2010;8 doi: 10.1371/journal.pbio.1000473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blatow M, Rozov A, Katona I, Hormuzdi SG, Meyer AH, Whittington MA, Caputi A, Monyer H. A novel network of multipolar bursting interneurons generates theta frequency oscillations in neocortex. Neuron. 2003;38:805–817. doi: 10.1016/s0896-6273(03)00300-3. [DOI] [PubMed] [Google Scholar]
- Bloss EB, Cembrowski MS, Karsh B, Colonell J, Fetter RD, Spruston N. Structured Dendritic Inhibition Supports Branch-Selective Integration in CA1 Pyramidal Cells. Neuron. 2016;89:1016–1030. doi: 10.1016/j.neuron.2016.01.029. [DOI] [PubMed] [Google Scholar]
- Bodor AL, Katona I, Nyiri G, Mackie K, Ledent C, Hajos N, Freund TF. Endocannabinoid signaling in rat somatosensory cortex: laminar differences and involvement of specific interneuron types. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:6845–6856. doi: 10.1523/JNEUROSCI.0442-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bortone DS, Olsen SR, Scanziani M. Translaminar inhibitory cells recruited by layer 6 corticothalamic neurons suppress visual cortex. Neuron. 2014;82:474–485. doi: 10.1016/j.neuron.2014.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braitenberg V, Schuz A. Peters’ Rule and White’s exceptions. In: Braitenberg V, Schuz A, editors. Cortex: Statistics and geometry of neuronal connectivity. Berlin Heidelberg Gmbh: Springer-Verlag; 1991. p. 249. [Google Scholar]
- Bruno RM, Sakmann B. Cortex is driven by weak but synchronously active thalamocortical synapses. Science (New York, NY) 2006;312:1622–1627. doi: 10.1126/science.1124593. [DOI] [PubMed] [Google Scholar]
- Bruno RM, Simons DJ. Feedforward mechanisms of excitatory and inhibitory cortical receptive fields. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2002;22:10966–10975. doi: 10.1523/JNEUROSCI.22-24-10966.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchanan KA, Blackman AV, Moreau AW, Elgar D, Costa RP, Lalanne T, Tudor Jones AA, Oyrer J, Sjostrom PJ. Target-specific expression of presynaptic NMDA receptors in neocortical microcircuits. Neuron. 2012;75:451–466. doi: 10.1016/j.neuron.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bucurenciu I, Bischofberger J, Jonas P. A small number of open Ca2+ channels trigger transmitter release at a central GABAergic synapse. Nature neuroscience. 2010;13:19–21. doi: 10.1038/nn.2461. [DOI] [PubMed] [Google Scholar]
- Bucurenciu I, Kulik A, Schwaller B, Frotscher M, Jonas P. Nanodomain coupling between Ca2+ channels and Ca2+ sensors promotes fast and efficient transmitter release at a cortical GABAergic synapse. Neuron. 2008;57:536–545. doi: 10.1016/j.neuron.2007.12.026. [DOI] [PubMed] [Google Scholar]
- Buhl DL, Harris KD, Hormuzdi SG, Monyer H, Buzsaki G. Selective impairment of hippocampal gamma oscillations in connexin-36 knock-out mouse in vivo. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2003;23:1013–1018. doi: 10.1523/JNEUROSCI.23-03-01013.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buzsaki G. Feed-forward inhibition in the hippocampal formation. Progress in neurobiology. 1984;22:131–153. doi: 10.1016/0301-0082(84)90023-6. [DOI] [PubMed] [Google Scholar]
- Buzsaki G, Wang XJ. Mechanisms of gamma oscillations. Annual review of neuroscience. 2012;35:203–225. doi: 10.1146/annurev-neuro-062111-150444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cadwell CR, Palasantza A, Jiang X, Berens P, Deng Q, Yilmaz M, Reimer J, Shen S, Bethge M, Tolias KF, et al. Electrophysiological, transcriptomic and morphologic profiling of single neurons using Patch-seq. Nat Biotechnol. 2016;34:199–203. doi: 10.1038/nbt.3445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caputi A, Melzer S, Michael M, Monyer H. The long and short of GABAergic neurons. Current opinion in neurobiology. 2013;23:179–186. doi: 10.1016/j.conb.2013.01.021. [DOI] [PubMed] [Google Scholar]
- Caputi A, Rozov A, Blatow M, Monyer H. Two calretinin-positive GABAergic cell types in layer 2/3 of the mouse neocortex provide different forms of inhibition. Cerebral cortex. 2009;19:1345–1359. doi: 10.1093/cercor/bhn175. [DOI] [PubMed] [Google Scholar]
- Cardin JA, Carlen M, Meletis K, Knoblich U, Zhang F, Deisseroth K, Tsai LH, Moore CI. Driving fast-spiking cells induces gamma rhythm and controls sensory responses. Nature. 2009;459:663–667. doi: 10.1038/nature08002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardin JA, Kumbhani RD, Contreras D, Palmer LA. Cellular mechanisms of temporal sensitivity in visual cortex neurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2010;30:3652–3662. doi: 10.1523/JNEUROSCI.5279-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardin JA, Palmer LA, Contreras D. Stimulus feature selectivity in excitatory and inhibitory neurons in primary visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2007;27:10333–10344. doi: 10.1523/JNEUROSCI.1692-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castel M, Morris J, Belenky M. Non-synaptic and dendritic exocytosis from dense-cored vesicles in the suprachiasmatic nucleus. Neuroreport. 1996;7:543–547. doi: 10.1097/00001756-199601310-00040. [DOI] [PubMed] [Google Scholar]
- Cauli B, Porter JT, Tsuzuki K, Lambolez B, Rossier J, Quenet B, Audinat E. Classification of fusiform neocortical interneurons based on unsupervised clustering. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:6144–6149. doi: 10.1073/pnas.97.11.6144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cauli B, Tong XK, Rancillac A, Serluca N, Lambolez B, Rossier J, Hamel E. Cortical GABA interneurons in neurovascular coupling: relays for subcortical vasoactive pathways. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2004;24:8940–8949. doi: 10.1523/JNEUROSCI.3065-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cauli B, Zhou X, Tricoire L, Toussay X, Staiger JF. Revisiting enigmatic cortical calretinin-expressing interneurons. Frontiers in neuroanatomy. 2014;8:52. doi: 10.3389/fnana.2014.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chittajallu R, Pelkey KA, McBain CJ. Neurogliaform cells dynamically regulate somatosensory integration via synapse-specific modulation. Nature neuroscience. 2013;16:13–15. doi: 10.1038/nn.3284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu CQ, Lur G, Morse TM, Carnevale NT, Ellis-Davies GC, Higley MJ. Compartmentalization of GABAergic inhibition by dendritic spines. Science (New York, NY) 2013;340:759–762. doi: 10.1126/science.1234274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi J, Callaway EM. Monosynaptic inputs to ErbB4-expressing inhibitory neurons in mouse primary somatosensory cortex. The Journal of comparative neurology. 2011;519:3402–3414. doi: 10.1002/cne.22680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu Z, Galarreta M, Hestrin S. Synaptic interactions of late-spiking neocortical neurons in layer 1. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2003;23:96–102. doi: 10.1523/JNEUROSCI.23-01-00096.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cobb SR, Buhl EH, Halasy K, Paulsen O, Somogyi P. Synchronization of neuronal activity in hippocampus by individual GABAergic interneurons. Nature. 1995;378:75–78. doi: 10.1038/378075a0. [DOI] [PubMed] [Google Scholar]
- Cope DW, Maccaferri G, Marton LF, Roberts JD, Cobden PM, Somogyi P. Cholecystokinin-immunopositive basket and Schaffer collateral-associated interneurones target different domains of pyramidal cells in the CA1 area of the rat hippocampus. Neuroscience. 2002;109:63–80. doi: 10.1016/s0306-4522(01)00440-7. [DOI] [PubMed] [Google Scholar]
- Couey JJ, Witoelar A, Zhang SJ, Zheng K, Ye J, Dunn B, Czajkowski R, Moser MB, Moser EI, Roudi Y, et al. Recurrent inhibitory circuitry as a mechanism for grid formation. Nature neuroscience. 2013;16:318–324. doi: 10.1038/nn.3310. [DOI] [PubMed] [Google Scholar]
- Cruikshank SJ, Ahmed OJ, Stevens TR, Patrick SL, Gonzalez AN, Elmaleh M, Connors BW. Thalamic control of layer 1 circuits in prefrontal cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2012;32:17813–17823. doi: 10.1523/JNEUROSCI.3231-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruikshank SJ, Lewis TJ, Connors BW. Synaptic basis for intense thalamocortical activation of feedforward inhibitory cells in neocortex. Nature neuroscience. 2007;10:462–468. doi: 10.1038/nn1861. [DOI] [PubMed] [Google Scholar]
- David C, Schleicher A, Zuschratter W, Staiger JF. The innervation of parvalbumin-containing interneurons by VIP-immunopositive interneurons in the primary somatosensory cortex of the adult rat. The European journal of neuroscience. 2007;25:2329–2340. doi: 10.1111/j.1460-9568.2007.05496.x. [DOI] [PubMed] [Google Scholar]
- de Lima AD, Morrison JH. Ultrastructural analysis of somatostatin-immunoreactive neurons and synapses in the temporal and occipital cortex of the macaque monkey. The Journal of comparative neurology. 1989;283:212–227. doi: 10.1002/cne.902830205. [DOI] [PubMed] [Google Scholar]
- De Marco Garcia NV, Karayannis T, Fishell G. Neuronal activity is required for the development of specific cortical interneuron subtypes. Nature. 2011;472:351–355. doi: 10.1038/nature09865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeFelipe J. Cortical interneurons: from Cajal to 2001. Progress in brain research. 2002;136:215–238. doi: 10.1016/s0079-6123(02)36019-9. [DOI] [PubMed] [Google Scholar]
- DeFelipe J, Ballesteros-Yanez I, Inda MC, Munoz A. Double-bouquet cells in the monkey and human cerebral cortex with special reference to areas 17 and 18. Progress in brain research. 2006;154:15–32. doi: 10.1016/S0079-6123(06)54002-6. [DOI] [PubMed] [Google Scholar]
- Defelipe J, Gonzalez-Albo MC, Del Rio MR, Elston GN. Distribution and patterns of connectivity of interneurons containing calbindin, calretinin, and parvalbumin in visual areas of the occipital and temporal lobes of the macaque monkey. The Journal of comparative neurology. 1999;412:515–526. doi: 10.1002/(sici)1096-9861(19990927)412:3<515::aid-cne10>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- Defelipe J, Jones EG. Cajal on the Cerebral Cortex: An anotated Translated of the Complete Writing. Oxford University Press; 1988. [Google Scholar]
- DeFelipe J, Lopez-Cruz PL, Benavides-Piccione R, Bielza C, Larranaga P, Anderson S, Burkhalter A, Cauli B, Fairen A, Feldmeyer D, et al. New insights into the classification and nomenclature of cortical GABAergic interneurons. Nature reviews Neuroscience. 2013;14:202–216. doi: 10.1038/nrn3444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennison-Cavanagh ME, Papadopoulos G, Parnavelas JG. The emergence of the cortical GABAergic neuron: with particular reference to some peptidergic subpopulations. Journal of neurocytology. 1993;22:805–814. doi: 10.1007/BF01181325. [DOI] [PubMed] [Google Scholar]
- Eggermann E, Bucurenciu I, Goswami SP, Jonas P. Nanodomain coupling between Ca(2)(+) channels and sensors of exocytosis at fast mammalian synapses. Nature reviews Neuroscience. 2012;13:7–21. doi: 10.1038/nrn3125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Boustani S, Sur M. Response-dependent dynamics of cell-specific inhibition in cortical networks in vivo. Nature communications. 2014;5:5689. doi: 10.1038/ncomms6689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erisir A, Lau D, Rudy B, Leonard CS. Function of specific K(+) channels in sustained high-frequency firing of fast-spiking neocortical interneurons. Journal of neurophysiology. 1999;82:2476–2489. doi: 10.1152/jn.1999.82.5.2476. [DOI] [PubMed] [Google Scholar]
- Fanselow EE, Richardson KA, Connors BW. Selective, state-dependent activation of somatostatin-expressing inhibitory interneurons in mouse neocortex. Journal of neurophysiology. 2008;100:2640–2652. doi: 10.1152/jn.90691.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fenno LE, Mattis J, Ramakrishnan C, Hyun M, Lee SY, He M, Tucciarone J, Selimbeyoglu A, Berndt A, Grosenick L, et al. Targeting cells with single vectors using multiple-feature Boolean logic. Nature methods. 2014;11:763–772. doi: 10.1038/nmeth.2996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferezou I, Cauli B, Hill EL, Rossier J, Hamel E, Lambolez B. 5-HT3 receptors mediate serotonergic fast synaptic excitation of neocortical vasoactive intestinal peptide/cholecystokinin interneurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2002;22:7389–7397. doi: 10.1523/JNEUROSCI.22-17-07389.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferezou I, Hill EL, Cauli B, Gibelin N, Kaneko T, Rossier J, Lambolez B. Extensive overlap of mu-opioid and nicotinic sensitivity in cortical interneurons. Cerebral cortex. 2007;17:1948–1957. doi: 10.1093/cercor/bhl104. [DOI] [PubMed] [Google Scholar]
- Fergus A, Lee KS. GABAergic regulation of cerebral microvascular tone in the rat. Journal of cerebral blood flow and metabolism: official journal of the International Society of Cerebral Blood Flow and Metabolism. 1997;17:992–1003. doi: 10.1097/00004647-199709000-00009. [DOI] [PubMed] [Google Scholar]
- Fino E, Yuste R. Dense inhibitory connectivity in neocortex. Neuron. 2011;69:1188–1203. doi: 10.1016/j.neuron.2011.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fishell G, Rudy B. Mechanisms of inhibition within the telencephalon: “where the wild things are”. Annual review of neuroscience. 2011;34:535–567. doi: 10.1146/annurev-neuro-061010-113717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freund-Mercier MJ, Stoeckel ME, Klein MJ. Oxytocin receptors on oxytocin neurones: histoautoradiographic detection in the lactating rat. The Journal of physiology. 1994;480(Pt 1):155–161. doi: 10.1113/jphysiol.1994.sp020349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freund TF. Interneuron Diversity series: Rhythm and mood in perisomatic inhibition. Trends in neurosciences. 2003;26:489–495. doi: 10.1016/S0166-2236(03)00227-3. [DOI] [PubMed] [Google Scholar]
- Freund TF, Antal M. GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature. 1988;336:170–173. doi: 10.1038/336170a0. [DOI] [PubMed] [Google Scholar]
- Freund TF, Katona I. Perisomatic inhibition. Neuron. 2007;56:33–42. doi: 10.1016/j.neuron.2007.09.012. [DOI] [PubMed] [Google Scholar]
- Freund TF, Magloczky Z, Soltesz I, Somogyi P. Synaptic connections, axonal and dendritic patterns of neurons immunoreactive for cholecystokinin in the visual cortex of the cat. Neuroscience. 1986;19:1133–1159. doi: 10.1016/0306-4522(86)90129-6. [DOI] [PubMed] [Google Scholar]
- Freund TF, Martin KA, Smith AD, Somogyi P. Glutamate decarboxylase-immunoreactive terminals of Golgi-impregnated axoaxonic cells and of presumed basket cells in synaptic contact with pyramidal neurons of the cat’s visual cortex. The Journal of comparative neurology. 1983;221:263–278. doi: 10.1002/cne.902210303. [DOI] [PubMed] [Google Scholar]
- Fricker D, Miles R. EPSP amplification and the precision of spike timing in hippocampal neurons. Neuron. 2000;28:559–569. doi: 10.1016/s0896-6273(00)00133-1. [DOI] [PubMed] [Google Scholar]
- Fu Y, Tucciarone JM, Espinosa JS, Sheng N, Darcy DP, Nicoll RA, Huang ZJ, Stryker MP. A cortical circuit for gain control by behavioral state. Cell. 2014;156:1139–1152. doi: 10.1016/j.cell.2014.01.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuzik J, Zeisel A, Mate Z, Calvigioni D, Yanagawa Y, Szabo G, Linnarsson S, Harkany T. Integration of electrophysiological recordings with single-cell RNA-seq data identifies neuronal subtypes. Nat Biotechnol. 2016;34:175–183. doi: 10.1038/nbt.3443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabernet L, Jadhav SP, Feldman DE, Carandini M, Scanziani M. Somatosensory integration controlled by dynamic thalamocortical feed-forward inhibition. Neuron. 2005;48:315–327. doi: 10.1016/j.neuron.2005.09.022. [DOI] [PubMed] [Google Scholar]
- Galarreta M, Erdelyi F, Szabo G, Hestrin S. Electrical coupling among irregular-spiking GABAergic interneurons expressing cannabinoid receptors. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2004;24:9770–9778. doi: 10.1523/JNEUROSCI.3027-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galarreta M, Hestrin S. A network of fast-spiking cells in the neocortex connected by electrical synapses. Nature. 1999;402:72–75. doi: 10.1038/47029. [DOI] [PubMed] [Google Scholar]
- Galarreta M, Hestrin S. Spike transmission and synchrony detection in networks of GABAergic interneurons. Science (New York, NY) 2001;292:2295–2299. doi: 10.1126/science.1061395. [DOI] [PubMed] [Google Scholar]
- Gambino F, Pages S, Kehayas V, Baptista D, Tatti R, Carleton A, Holtmaat A. Sensory-evoked LTP driven by dendritic plateau potentials in vivo. Nature. 2014;515:116–119. doi: 10.1038/nature13664. [DOI] [PubMed] [Google Scholar]
- Geiger JR, Melcher T, Koh DS, Sakmann B, Seeburg PH, Jonas P, Monyer H. Relative abundance of subunit mRNAs determines gating and Ca2+ permeability of AMPA receptors in principal neurons and interneurons in rat CNS. Neuron. 1995;15:193–204. doi: 10.1016/0896-6273(95)90076-4. [DOI] [PubMed] [Google Scholar]
- Gentet LJ, Avermann M, Matyas F, Staiger JF, Petersen CC. Membrane potential dynamics of GABAergic neurons in the barrel cortex of behaving mice. Neuron. 2010;65:422–435. doi: 10.1016/j.neuron.2010.01.006. [DOI] [PubMed] [Google Scholar]
- Gentet LJ, Kremer Y, Taniguchi H, Huang ZJ, Staiger JF, Petersen CC. Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex. Nature neuroscience. 2012;15:607–612. doi: 10.1038/nn.3051. [DOI] [PubMed] [Google Scholar]
- Gibson JR, Beierlein M, Connors BW. Two networks of electrically coupled inhibitory neurons in neocortex. Nature. 1999;402:75–79. doi: 10.1038/47035. [DOI] [PubMed] [Google Scholar]
- Gibson JR, Beierlein M, Connors BW. Functional properties of electrical synapses between inhibitory interneurons of neocortical layer 4. Journal of neurophysiology. 2005;93:467–480. doi: 10.1152/jn.00520.2004. [DOI] [PubMed] [Google Scholar]
- Gidon A, Segev I. Principles governing the operation of synaptic inhibition in dendrites. Neuron. 2012;75:330–341. doi: 10.1016/j.neuron.2012.05.015. [DOI] [PubMed] [Google Scholar]
- Glickfeld LL, Roberts JD, Somogyi P, Scanziani M. Interneurons hyperpolarize pyramidal cells along their entire somatodendritic axis. Nature neuroscience. 2009;12:21–23. doi: 10.1038/nn.2230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glickfeld LL, Scanziani M. Distinct timing in the activity of cannabinoid-sensitive and cannabinoid-insensitive basket cells. Nature neuroscience. 2006;9:807–815. doi: 10.1038/nn1688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg EM, Clark BD, Zagha E, Nahmani M, Erisir A, Rudy B. K+ channels at the axon initial segment dampen near-threshold excitability of neocortical fast-spiking GABAergic interneurons. Neuron. 2008;58:387–400. doi: 10.1016/j.neuron.2008.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg EM, Coulter DA. Mechanisms of epileptogenesis: a convergence on neural circuit dysfunction. Nature reviews Neuroscience. 2013;14:337–349. doi: 10.1038/nrn3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg EM, Jeong HY, Kruglikov I, Tremblay R, Lazarenko RM, Rudy B. Rapid developmental maturation of neocortical FS cell intrinsic excitability. Cerebral cortex. 2011;21:666–682. doi: 10.1093/cercor/bhq138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg EM, Watanabe S, Chang SY, Joho RH, Huang ZJ, Leonard CS, Rudy B. Specific functions of synaptically localized potassium channels in synaptic transmission at the neocortical GABAergic fast-spiking cell synapse. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:5230–5235. doi: 10.1523/JNEUROSCI.0722-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonchar Y, Burkhalter A. Connectivity of GABAergic calretinin-immunoreactive neurons in rat primary visual cortex. Cerebral cortex. 1999;9:683–696. doi: 10.1093/cercor/9.7.683. [DOI] [PubMed] [Google Scholar]
- Gonchar Y, Burkhalter A. Distinct GABAergic targets of feedforward and feedback connections between lower and higher areas of rat visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2003;23:10904–10912. doi: 10.1523/JNEUROSCI.23-34-10904.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez-Burgos G. GABA transporter GAT1: a crucial determinant of GABAB receptor activation in cortical circuits? Advances in pharmacology (San Diego, Calif) 2010;58:175–204. doi: 10.1016/S1054-3589(10)58008-6. [DOI] [PubMed] [Google Scholar]
- Gulyas AI, Hajos N, Freund TF. Interneurons containing calretinin are specialized to control other interneurons in the rat hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1996;16:3397–3411. doi: 10.1523/JNEUROSCI.16-10-03397.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulyas AI, Megias M, Emri Z, Freund TF. Total number and ratio of excitatory and inhibitory synapses converging onto single interneurons of different types in the CA1 area of the rat hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1999;19:10082–10097. doi: 10.1523/JNEUROSCI.19-22-10082.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta A, Wang Y, Markram H. Organizing principles for a diversity of GABAergic interneurons and synapses in the neocortex. Science (New York, NY) 2000;287:273–278. doi: 10.1126/science.287.5451.273. [DOI] [PubMed] [Google Scholar]
- Hajos N, Acsady L, Freund TF. Target selectivity and neurochemical characteristics of VIP-immunoreactive interneurons in the rat dentate gyrus. The European journal of neuroscience. 1996;8:1415–1431. doi: 10.1111/j.1460-9568.1996.tb01604.x. [DOI] [PubMed] [Google Scholar]
- Hefft S, Jonas P. Asynchronous GABA release generates long-lasting inhibition at a hippocampal interneuron-principal neuron synapse. Nature neuroscience. 2005;8:1319–1328. doi: 10.1038/nn1542. [DOI] [PubMed] [Google Scholar]
- Helmstaedter M, Staiger JF, Sakmann B, Feldmeyer D. Efficient recruitment of layer 2/3 interneurons by layer 4 input in single columns of rat somatosensory cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2008;28:8273–8284. doi: 10.1523/JNEUROSCI.5701-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hestrin S, Armstrong WE. Morphology and physiology of cortical neurons in layer I. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1996;16:5290–5300. doi: 10.1523/JNEUROSCI.16-17-05290.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hestrin S, Galarreta M. Electrical synapses define networks of neocortical GABAergic neurons. Trends in neurosciences. 2005;28:304–309. doi: 10.1016/j.tins.2005.04.001. [DOI] [PubMed] [Google Scholar]
- Higley MJ, Contreras D. Balanced excitation and inhibition determine spike timing during frequency adaptation. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2006;26:448–457. doi: 10.1523/JNEUROSCI.3506-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hioki H, Okamoto S, Konno M, Kameda H, Sohn J, Kuramoto E, Fujiyama F, Kaneko T. Cell type-specific inhibitory inputs to dendritic and somatic compartments of parvalbumin-expressing neocortical interneuron. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:544–555. doi: 10.1523/JNEUROSCI.2255-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirsch JA, Martinez LM, Pillai C, Alonso JM, Wang Q, Sommer FT. Functionally distinct inhibitory neurons at the first stage of visual cortical processing. Nature neuroscience. 2003;6:1300–1308. doi: 10.1038/nn1152. [DOI] [PubMed] [Google Scholar]
- Hof PR, Glezer II, Conde F, Flagg RA, Rubin MB, Nimchinsky EA, Vogt Weisenhorn DM. Cellular distribution of the calcium-binding proteins parvalbumin, calbindin, and calretinin in the neocortex of mammals: phylogenetic and developmental patterns. Journal of chemical neuroanatomy. 1999;16:77–116. doi: 10.1016/s0891-0618(98)00065-9. [DOI] [PubMed] [Google Scholar]
- Hofer SB, Ko H, Pichler B, Vogelstein J, Ros H, Zeng H, Lein E, Lesica NA, Mrsic-Flogel TD. Differential connectivity and response dynamics of excitatory and inhibitory neurons in visual cortex. Nature neuroscience. 2011;14:1045–1052. doi: 10.1038/nn.2876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- House DR, Elstrott J, Koh E, Chung J, Feldman DE. Parallel regulation of feedforward inhibition and excitation during whisker map plasticity. Neuron. 2011;72:819–831. doi: 10.1016/j.neuron.2011.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howard A, Tamas G, Soltesz I. Lighting the chandelier: new vistas for axo-axonic cells. Trends in neurosciences. 2005;28:310–316. doi: 10.1016/j.tins.2005.04.004. [DOI] [PubMed] [Google Scholar]
- Hu H, Gan J, Jonas P. Interneurons. Fast-spiking, parvalbumin(+) GABAergic interneurons: from cellular design to microcircuit function. Science (New York, NY) 2014;345:1255263. doi: 10.1126/science.1255263. [DOI] [PubMed] [Google Scholar]
- Hu H, Jonas P. A supercritical density of Na(+) channels ensures fast signaling in GABAergic interneuron axons. Nature neuroscience. 2014;17:686–693. doi: 10.1038/nn.3678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu H, Martina M, Jonas P. Dendritic mechanisms underlying rapid synaptic activation of fast-spiking hippocampal interneurons. Science (New York, NY) 2010;327:52–58. doi: 10.1126/science.1177876. [DOI] [PubMed] [Google Scholar]
- Huang ZJ. Toward a genetic dissection of cortical circuits in the mouse. Neuron. 2014;83:1284–1302. doi: 10.1016/j.neuron.2014.08.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hull C, Isaacson JS, Scanziani M. Postsynaptic mechanisms govern the differential excitation of cortical neurons by thalamic inputs. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:9127–9136. doi: 10.1523/JNEUROSCI.5971-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt RF, Girskis KM, Rubenstein JL, Alvarez-Buylla A, Baraban SC. GABA progenitors grafted into the adult epileptic brain control seizures and abnormal behavior. Nature neuroscience. 2013;16:692–697. doi: 10.1038/nn.3392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurbin A, Orcel H, Alonso G, Moos F, Rabie A. The vasopressin receptors colocalize with vasopressin in the magnocellular neurons of the rat supraoptic nucleus and are modulated by water balance. Endocrinology. 2002;143:456–466. doi: 10.1210/endo.143.2.8643. [DOI] [PubMed] [Google Scholar]
- Inoue T, Imoto K. Feedforward inhibitory connections from multiple thalamic cells to multiple regular-spiking cells in layer 4 of the somatosensory cortex. Journal of neurophysiology. 2006;96:1746–1754. doi: 10.1152/jn.00301.2006. [DOI] [PubMed] [Google Scholar]
- Ito-Ishida A, Ure K, Chen H, Swann JW, Zoghbi HY. Loss of MeCP2 in Parvalbumin-and Somatostatin-Expressing Neurons in Mice Leads to Distinct Rett Syndrome-like Phenotypes. Neuron. 2015;88:651–658. doi: 10.1016/j.neuron.2015.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang X, Shen S, Cadwell CR, Berens P, Sinz F, Ecker AS, Patel S, Tolias AS. Principles of connectivity among morphologically defined cell types in adult neocortex. Science (New York, NY) 2015;350:aac9462. doi: 10.1126/science.aac9462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang X, Wang G, Lee AJ, Stornetta RL, Zhu JJ. The organization of two new cortical interneuronal circuits. Nature neuroscience. 2013;16:210–218. doi: 10.1038/nn.3305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jinno S. Structural organization of long-range GABAergic projection system of the hippocampus. Frontiers in neuroanatomy. 2009;3:13. doi: 10.3389/neuro.05.013.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jinno S, Kosaka T. Parvalbumin is expressed in glutamatergic and GABAergic corticostriatal pathway in mice. The Journal of comparative neurology. 2004;477:188–201. doi: 10.1002/cne.20246. [DOI] [PubMed] [Google Scholar]
- Kapfer C, Glickfeld LL, Atallah BV, Scanziani M. Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex. Nature neuroscience. 2007;10:743–753. doi: 10.1038/nn1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karnani MM, Jackson J, Ayzenshtat I, Tucciarone J, Manoocheri K, Snider WG, Yuste R. Cooperative Subnetworks of Molecularly Similar Interneurons in Mouse Neocortex. Neuron. 2016;90:86–100. doi: 10.1016/j.neuron.2016.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karube F, Kubota Y, Kawaguchi Y. Axon branching and synaptic bouton phenotypes in GABAergic nonpyramidal cell subtypes. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2004;24:2853–2865. doi: 10.1523/JNEUROSCI.4814-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katzel D, Zemelman BV, Buetfering C, Wolfel M, Miesenbock G. The columnar and laminar organization of inhibitory connections to neocortical excitatory cells. Nature neuroscience. 2011;14:100–107. doi: 10.1038/nn.2687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katzner S, Busse L, Carandini M. GABAA inhibition controls response gain in visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2011;31:5931–5941. doi: 10.1523/JNEUROSCI.5753-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaguchi Y. Physiological subgroups of nonpyramidal cells with specific morphological characteristics in layer II/III of rat frontal cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1995;15:2638–2655. doi: 10.1523/JNEUROSCI.15-04-02638.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaguchi Y. Selective cholinergic modulation of cortical GABAergic cell subtypes. Journal of neurophysiology. 1997;78:1743–1747. doi: 10.1152/jn.1997.78.3.1743. [DOI] [PubMed] [Google Scholar]
- Kawaguchi Y, Kubota Y. Physiological and morphological identification of somatostatin- or vasoactive intestinal polypeptide-containing cells among GABAergic cell subtypes in rat frontal cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1996;16:2701–2715. doi: 10.1523/JNEUROSCI.16-08-02701.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaguchi Y, Kubota Y. GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cerebral cortex. 1997;7:476–486. doi: 10.1093/cercor/7.6.476. [DOI] [PubMed] [Google Scholar]
- Kawaguchi Y, Shindou T. Noradrenergic excitation and inhibition of GABAergic cell types in rat frontal cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1998;18:6963–6976. doi: 10.1523/JNEUROSCI.18-17-06963.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kepecs A, Fishell G. Interneuron cell types are fit to function. Nature. 2014;505:318–326. doi: 10.1038/nature12983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerlin AM, Andermann ML, Berezovskii VK, Reid RC. Broadly tuned response properties of diverse inhibitory neuron subtypes in mouse visual cortex. Neuron. 2010;67:858–871. doi: 10.1016/j.neuron.2010.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim U, Sanchez-Vives MV, McCormick DA. Functional dynamics of GABAergic inhibition in the thalamus. Science (New York, NY) 1997;278:130–134. doi: 10.1126/science.278.5335.130. [DOI] [PubMed] [Google Scholar]
- Kisvarday ZF. GABAergic networks of basket cells in the visual cortex. Progress in brain research. 1992;90:385–405. doi: 10.1016/s0079-6123(08)63623-7. [DOI] [PubMed] [Google Scholar]
- Kisvarday ZF, Beaulieu C, Eysel UT. Network of GABAergic large basket cells in cat visual cortex (area 18): implication for lateral disinhibition. The Journal of comparative neurology. 1993;327:398–415. doi: 10.1002/cne.903270307. [DOI] [PubMed] [Google Scholar]
- Kisvarday ZF, Gulyas A, Beroukas D, North JB, Chubb IW, Somogyi P. Synapses, axonal and dendritic patterns of GABA-immunoreactive neurons in human cerebral cortex. Brain. 1990;113(Pt 3):793–812. doi: 10.1093/brain/113.3.793. [DOI] [PubMed] [Google Scholar]
- Kisvarday ZF, Martin KA, Whitteridge D, Somogyi P. Synaptic connections of intracellularly filled clutch cells: a type of small basket cell in the visual cortex of the cat. The Journal of comparative neurology. 1985;241:111–137. doi: 10.1002/cne.902410202. [DOI] [PubMed] [Google Scholar]
- Klausberger T, Marton LF, O’Neill J, Huck JH, Dalezios Y, Fuentealba P, Suen WY, Papp E, Kaneko T, Watanabe M, et al. Complementary roles of cholecystokinin- and parvalbumin-expressing GABAergic neurons in hippocampal network oscillations. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:9782–9793. doi: 10.1523/JNEUROSCI.3269-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klausberger T, Somogyi P. Neuronal diversity and temporal dynamics: the unity of hippocampal circuit operations. Science (New York, NY) 2008;321:53–57. doi: 10.1126/science.1149381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kloc M, Maffei A. Target-specific properties of thalamocortical synapses onto layer 4 of mouse primary visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2014;34:15455–15465. doi: 10.1523/JNEUROSCI.2595-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch C, Poggio T, Torre V. Nonlinear interactions in a dendritic tree: localization, timing, and role in information processing. Proceedings of the National Academy of Sciences of the United States of America. 1983;80:2799–2802. doi: 10.1073/pnas.80.9.2799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruglikov I, Rudy B. Perisomatic GABA release and thalamocortical integration onto neocortical excitatory cells are regulated by neuromodulators. Neuron. 2008;58:911–924. doi: 10.1016/j.neuron.2008.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubota Y. Untangling GABAergic wiring in the cortical microcircuit. Current opinion in neurobiology. 2014;26:7–14. doi: 10.1016/j.conb.2013.10.003. [DOI] [PubMed] [Google Scholar]
- Kubota Y, Karube F, Nomura M, Gulledge AT, Mochizuki A, Schertel A, Kawaguchi Y. Conserved properties of dendritic trees in four cortical interneuron subtypes. Scientific reports. 2011a;1:89. doi: 10.1038/srep00089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubota Y, Kawaguchi Y. Two distinct subgroups of cholecystokinin-immunoreactive cortical interneurons. Brain research. 1997;752:175–183. doi: 10.1016/s0006-8993(96)01446-1. [DOI] [PubMed] [Google Scholar]
- Kubota Y, Shigematsu N, Karube F, Sekigawa A, Kato S, Yamaguchi N, Hirai Y, Morishima M, Kawaguchi Y. Selective coexpression of multiple chemical markers defines discrete populations of neocortical GABAergic neurons. Cerebral cortex. 2011b;21:1803–1817. doi: 10.1093/cercor/bhq252. [DOI] [PubMed] [Google Scholar]
- Kvitsiani D, Ranade S, Hangya B, Taniguchi H, Huang JZ, Kepecs A. Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature. 2013;498:363–366. doi: 10.1038/nature12176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwan AC, Dan Y. Dissection of cortical microcircuits by single-neuron stimulation in vivo. Current biology: CB. 2012;22:1459–1467. doi: 10.1016/j.cub.2012.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landry M, Vila-Porcile E, Hokfelt T, Calas A. Differential routing of coexisting neuropeptides in vasopressin neurons. The European journal of neuroscience. 2003;17:579–589. [PubMed] [Google Scholar]
- Larkum M. A cellular mechanism for cortical associations: an organizing principle for the cerebral cortex. Trends in neurosciences. 2013;36:141–151. doi: 10.1016/j.tins.2012.11.006. [DOI] [PubMed] [Google Scholar]
- Larkum ME, Zhu JJ, Sakmann B. A new cellular mechanism for coupling inputs arriving at different cortical layers. Nature. 1999;398:338–341. doi: 10.1038/18686. [DOI] [PubMed] [Google Scholar]
- Lee AT, Gee SM, Vogt D, Patel T, Rubenstein JL, Sohal VS. Pyramidal neurons in prefrontal cortex receive subtype-specific forms of excitation and inhibition. Neuron. 2014a;81:61–68. doi: 10.1016/j.neuron.2013.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee AT, Vogt D, Rubenstein JL, Sohal VS. A class of GABAergic neurons in the prefrontal cortex sends long-range projections to the nucleus accumbens and elicits acute avoidance behavior. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2014b;34:11519–11525. doi: 10.1523/JNEUROSCI.1157-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee S, Hjerling-Leffler J, Zagha E, Fishell G, Rudy B. The largest group of superficial neocortical GABAergic interneurons expresses ionotropic serotonin receptors. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2010;30:16796–16808. doi: 10.1523/JNEUROSCI.1869-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee S, Kruglikov I, Huang ZJ, Fishell G, Rudy B. A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nature neuroscience. 2013;16:1662–1670. doi: 10.1038/nn.3544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SH, Kwan AC, Dan Y. Interneuron subtypes and orientation tuning. Nature. 2014c;508:E1–2. doi: 10.1038/nature13128. [DOI] [PubMed] [Google Scholar]
- Lee SH, Kwan AC, Zhang S, Phoumthipphavong V, Flannery JG, Masmanidis SC, Taniguchi H, Huang ZJ, Zhang F, Boyden ES, et al. Activation of specific interneurons improves V1 feature selectivity and visual perception. Nature. 2012;488:379–383. doi: 10.1038/nature11312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SH, Marchionni I, Bezaire M, Varga C, Danielson N, Lovett-Barron M, Losonczy A, Soltesz I. Parvalbumin-positive basket cells differentiate among hippocampal pyramidal cells. Neuron. 2014d;82:1129–1144. doi: 10.1016/j.neuron.2014.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SH, Simons DJ. Angular tuning and velocity sensitivity in different neuron classes within layer 4 of rat barrel cortex. Journal of neurophysiology. 2004;91:223–229. doi: 10.1152/jn.00541.2003. [DOI] [PubMed] [Google Scholar]
- Letzkus JJ, Wolff SB, Meyer EM, Tovote P, Courtin J, Herry C, Luthi A. A disinhibitory microcircuit for associative fear learning in the auditory cortex. Nature. 2011;480:331–335. doi: 10.1038/nature10674. [DOI] [PubMed] [Google Scholar]
- Lewis DA. Inhibitory neurons in human cortical circuits: substrate for cognitive dysfunction in schizophrenia. Current opinion in neurobiology. 2014;26:22–26. doi: 10.1016/j.conb.2013.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Bischofberger J, Jonas P. Differential gating and recruitment of P/Q-, N-, and R-type Ca2+ channels in hippocampal mossy fiber boutons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2007;27:13420–13429. doi: 10.1523/JNEUROSCI.1709-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu BH, Li P, Sun YJ, Li YT, Zhang LI, Tao HW. Intervening inhibition underlies simple-cell receptive field structure in visual cortex. Nature neuroscience. 2010;13:89–96. doi: 10.1038/nn.2443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu G. Local structural balance and functional interaction of excitatory and inhibitory synapses in hippocampal dendrites. Nature neuroscience. 2004;7:373–379. doi: 10.1038/nn1206. [DOI] [PubMed] [Google Scholar]
- Lovett-Barron M, Turi GF, Kaifosh P, Lee PH, Bolze F, Sun XH, Nicoud JF, Zemelman BV, Sternson SM, Losonczy A. Regulation of neuronal input transformations by tunable dendritic inhibition. Nature neuroscience. 2012;15:423–430. s421–423. doi: 10.1038/nn.3024. [DOI] [PubMed] [Google Scholar]
- Ludwig M, Bull PM, Tobin VA, Sabatier N, Landgraf R, Dayanithi G, Leng G. Regulation of activity-dependent dendritic vasopressin release from rat supraoptic neurones. The Journal of physiology. 2005;564:515–522. doi: 10.1113/jphysiol.2005.083931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig M, Sabatier N, Bull PM, Landgraf R, Dayanithi G, Leng G. Intracellular calcium stores regulate activity-dependent neuropeptide release from dendrites. Nature. 2002;418:85–89. doi: 10.1038/nature00822. [DOI] [PubMed] [Google Scholar]
- Lund JS. Anatomical organization of macaque monkey striate visual cortex. Annual review of neuroscience. 1988;11:253–288. doi: 10.1146/annurev.ne.11.030188.001345. [DOI] [PubMed] [Google Scholar]
- Ma WP, Liu BH, Li YT, Huang ZJ, Zhang LI, Tao HW. Visual representations by cortical somatostatin inhibitory neurons--selective but with weak and delayed responses. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2010;30:14371–14379. doi: 10.1523/JNEUROSCI.3248-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Y, Hu H, Berrebi AS, Mathers PH, Agmon A. Distinct subtypes of somatostatin-containing neocortical interneurons revealed in transgenic mice. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2006;26:5069–5082. doi: 10.1523/JNEUROSCI.0661-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallet N, Le Moine C, Charpier S, Gonon F. Feedforward inhibition of projection neurons by fast-spiking GABA interneurons in the rat striatum in vivo. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:3857–3869. doi: 10.1523/JNEUROSCI.5027-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marin O. Interneuron dysfunction in psychiatric disorders. Nature reviews Neuroscience. 2012;13:107–120. doi: 10.1038/nrn3155. [DOI] [PubMed] [Google Scholar]
- Markram H, Toledo-Rodriguez M, Wang Y, Gupta A, Silberberg G, Wu C. Interneurons of the neocortical inhibitory system. Nature reviews Neuroscience. 2004;5:793–807. doi: 10.1038/nrn1519. [DOI] [PubMed] [Google Scholar]
- Martin KA, Somogyi P, Whitteridge D. Physiological and morphological properties of identified basket cells in the cat’s visual cortex. Experimental brain research. 1983;50:193–200. doi: 10.1007/BF00239183. [DOI] [PubMed] [Google Scholar]
- Martina M, Jonas P. Functional differences in Na+ channel gating between fast-spiking interneurones and principal neurones of rat hippocampus. The Journal of physiology. 1997;505(Pt 3):593–603. doi: 10.1111/j.1469-7793.1997.593ba.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez LM, Wang Q, Reid RC, Pillai C, Alonso JM, Sommer FT, Hirsch JA. Receptive field structure varies with layer in the primary visual cortex. Nature neuroscience. 2005;8:372–379. doi: 10.1038/nn1404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massi L, Lagler M, Hartwich K, Borhegyi Z, Somogyi P, Klausberger T. Temporal dynamics of parvalbumin-expressing axo-axonic and basket cells in the rat medial prefrontal cortex in vivo. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2012;32:16496–16502. doi: 10.1523/JNEUROSCI.3475-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuire BA, Hornung JP, Gilbert CD, Wiesel TN. Patterns of synaptic input to layer 4 of cat striate cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1984;4:3021–3033. doi: 10.1523/JNEUROSCI.04-12-03021.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Megias M, Emri Z, Freund TF, Gulyas AI. Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells. Neuroscience. 2001;102:527–540. doi: 10.1016/s0306-4522(00)00496-6. [DOI] [PubMed] [Google Scholar]
- Mehaffey WH, Doiron B, Maler L, Turner RW. Deterministic multiplicative gain control with active dendrites. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:9968–9977. doi: 10.1523/JNEUROSCI.2682-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melzer S, Michael M, Caputi A, Eliava M, Fuchs EC, Whittington MA, Monyer H. Long-range-projecting GABAergic neurons modulate inhibition in hippocampus and entorhinal cortex. Science (New York, NY) 2012;335:1506–1510. doi: 10.1126/science.1217139. [DOI] [PubMed] [Google Scholar]
- Meskenaite V. Calretinin-immunoreactive local circuit neurons in area 17 of the cynomolgus monkey, Macaca fascicularis. The Journal of comparative neurology. 1997;379:113–132. [PubMed] [Google Scholar]
- Meyer HS, Schwarz D, Wimmer VC, Schmitt AC, Kerr JN, Sakmann B, Helmstaedter M. Inhibitory interneurons in a cortical column form hot zones of inhibition in layers 2 and 5A. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:16807–16812. doi: 10.1073/pnas.1113648108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles R. Variation in strength of inhibitory synapses in the CA3 region of guinea-pig hippocampus in vitro. The Journal of physiology. 1990;431:659–676. doi: 10.1113/jphysiol.1990.sp018353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles R, Toth K, Gulyas AI, Hajos N, Freund TF. Differences between somatic and dendritic inhibition in the hippocampus. Neuron. 1996;16:815–823. doi: 10.1016/s0896-6273(00)80101-4. [DOI] [PubMed] [Google Scholar]
- Miller KD, Pinto DJ, Simons DJ. Processing in layer 4 of the neocortical circuit: new insights from visual and somatosensory cortex. Current opinion in neurobiology. 2001;11:488–497. doi: 10.1016/s0959-4388(00)00239-7. [DOI] [PubMed] [Google Scholar]
- Mitchell SJ, Silver RA. Shunting inhibition modulates neuronal gain during synaptic excitation. Neuron. 2003;38:433–445. doi: 10.1016/s0896-6273(03)00200-9. [DOI] [PubMed] [Google Scholar]
- Mody I, De Koninck Y, Otis TS, Soltesz I. Bridging the cleft at GABA synapses in the brain. Trends in neurosciences. 1994;17:517–525. doi: 10.1016/0166-2236(94)90155-4. [DOI] [PubMed] [Google Scholar]
- Molnar G, Farago N, Kocsis AK, Rozsa M, Lovas S, Boldog E, Baldi R, Csajbok E, Gardi J, Puskas LG, et al. GABAergic neurogliaform cells represent local sources of insulin in the cerebral cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2014;34:1133–1137. doi: 10.1523/JNEUROSCI.4082-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore AK, Wehr M. Parvalbumin-expressing inhibitory interneurons in auditory cortex are well-tuned for frequency. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:13713–13723. doi: 10.1523/JNEUROSCI.0663-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JF, Pow DV. Widespread release of peptides in the central nervous system: quantitation of tannic acid-captured exocytoses. Anat Rec. 1991;231:437–445. doi: 10.1002/ar.1092310406. [DOI] [PubMed] [Google Scholar]
- Munoz W, Rudy B. Spatiotemporal specificity in cholinergic control of neocortical function. Current opinion in neurobiology. 2014;26:149–160. doi: 10.1016/j.conb.2014.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munoz W, Tremblay R, Rudy B. Channelrhodopsin-assisted patching: in vivo recording of genetically and morphologically identified neurons throughout the brain. Cell reports. 2014;9:2304–2316. doi: 10.1016/j.celrep.2014.11.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murayama M, Perez-Garci E, Nevian T, Bock T, Senn W, Larkum ME. Dendritic encoding of sensory stimuli controlled by deep cortical interneurons. Nature. 2009;457:1137–1141. doi: 10.1038/nature07663. [DOI] [PubMed] [Google Scholar]
- Nakajima M, Gorlich A, Heintz N. Oxytocin modulates female sociosexual behavior through a specific class of prefrontal cortical interneurons. Cell. 2014;159:295–305. doi: 10.1016/j.cell.2014.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson S, Toth L, Sheth B, Sur M. Orientation selectivity of cortical neurons during intracellular blockade of inhibition. Science (New York, NY) 1994;265:774–777. doi: 10.1126/science.8047882. [DOI] [PubMed] [Google Scholar]
- Norenberg A, Hu H, Vida I, Bartos M, Jonas P. Distinct nonuniform cable properties optimize rapid and efficient activation of fast-spiking GABAergic interneurons. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:894–899. doi: 10.1073/pnas.0910716107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowak LG, Sanchez-Vives MV, McCormick DA. Lack of orientation and direction selectivity in a subgroup of fast-spiking inhibitory interneurons: cellular and synaptic mechanisms and comparison with other electrophysiological cell types. Cerebral cortex. 2008;18:1058–1078. doi: 10.1093/cercor/bhm137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okaty BW, Miller MN, Sugino K, Hempel CM, Nelson SB. Transcriptional and electrophysiological maturation of neocortical fast-spiking GABAergic interneurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:7040–7052. doi: 10.1523/JNEUROSCI.0105-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olah S, Fule M, Komlosi G, Varga C, Baldi R, Barzo P, Tamas G. Regulation of cortical microcircuits by unitary GABA-mediated volume transmission. Nature. 2009;461:1278–1281. doi: 10.1038/nature08503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olah S, Komlosi G, Szabadics J, Varga C, Toth E, Barzo P, Tamas G. Output of neurogliaform cells to various neuron types in the human and rat cerebral cortex. Frontiers in neural circuits. 2007;1:4. doi: 10.3389/neuro.04.004.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oswald AM, Doiron B, Rinzel J, Reyes AD. Spatial profile and differential recruitment of GABAB modulate oscillatory activity in auditory cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:10321–10334. doi: 10.1523/JNEUROSCI.1703-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozeki H, Sadakane O, Akasaki T, Naito T, Shimegi S, Sato H. Relationship between excitation and inhibition underlying size tuning and contextual response modulation in the cat primary visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2004;24:1428–1438. doi: 10.1523/JNEUROSCI.3852-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Packer AM, McConnell DJ, Fino E, Yuste R. Axo-dendritic overlap and laminar projection can explain interneuron connectivity to pyramidal cells. Cerebral cortex. 2013;23:2790–2802. doi: 10.1093/cercor/bhs210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Packer AM, Yuste R. Dense, unspecific connectivity of neocortical parvalbumin-positive interneurons: a canonical microcircuit for inhibition? The Journal of neuroscience: the official journal of the Society for Neuroscience. 2011;31:13260–13271. doi: 10.1523/JNEUROSCI.3131-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer LM, Schulz JM, Murphy SC, Ledergerber D, Murayama M, Larkum ME. The cellular basis of GABA(B)-mediated interhemispheric inhibition. Science (New York, NY) 2012;335:989–993. doi: 10.1126/science.1217276. [DOI] [PubMed] [Google Scholar]
- Paz JT, Huguenard JR. Microcircuits and their interactions in epilepsy: is the focus out of focus? Nature neuroscience. 2015;18:351–359. doi: 10.1038/nn.3950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters A. The axon terminals of vasoactive intestinal polypeptide (VIP)-containing bipolar cells in rat visual cortex. Journal of neurocytology. 1990;19:672–685. doi: 10.1007/BF01188036. [DOI] [PubMed] [Google Scholar]
- Peters A, Feldman ML. The projection of the lateral geniculate nucleus to area 17 of the rat cerebral cortex. I. General description. Journal of neurocytology. 1976;5:63–84. doi: 10.1007/BF01176183. [DOI] [PubMed] [Google Scholar]
- Petreanu L, Mao T, Sternson SM, Svoboda K. The subcellular organization of neocortical excitatory connections. Nature. 2009;457:1142–1145. doi: 10.1038/nature07709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeffer CK, Xue M, He M, Huang ZJ, Scanziani M. Inhibition of inhibition in visual cortex: the logic of connections between molecularly distinct interneurons. Nature neuroscience. 2013;16:1068–1076. doi: 10.1038/nn.3446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pi HJ, Hangya B, Kvitsiani D, Sanders JI, Huang ZJ, Kepecs A. Cortical interneurons that specialize in disinhibitory control. Nature. 2013;503:521–524. doi: 10.1038/nature12676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinto DJ, Brumberg JC, Simons DJ. Circuit dynamics and coding strategies in rodent somatosensory cortex. Journal of neurophysiology. 2000;83:1158–1166. doi: 10.1152/jn.2000.83.3.1158. [DOI] [PubMed] [Google Scholar]
- Pinto DJ, Hartings JA, Brumberg JC, Simons DJ. Cortical damping: analysis of thalamocortical response transformations in rodent barrel cortex. Cerebral cortex. 2003;13:33–44. doi: 10.1093/cercor/13.1.33. [DOI] [PubMed] [Google Scholar]
- Porter JT, Cauli B, Staiger JF, Lambolez B, Rossier J, Audinat E. Properties of bipolar VIPergic interneurons and their excitation by pyramidal neurons in the rat neocortex. The European journal of neuroscience. 1998;10:3617–3628. doi: 10.1046/j.1460-9568.1998.00367.x. [DOI] [PubMed] [Google Scholar]
- Pouille F, Marin-Burgin A, Adesnik H, Atallah BV, Scanziani M. Input normalization by global feedforward inhibition expands cortical dynamic range. Nature neuroscience. 2009;12:1577–1585. doi: 10.1038/nn.2441. [DOI] [PubMed] [Google Scholar]
- Pouille F, Scanziani M. Enforcement of temporal fidelity in pyramidal cells by somatic feed-forward inhibition. Science (New York, NY) 2001;293:1159–1163. doi: 10.1126/science.1060342. [DOI] [PubMed] [Google Scholar]
- Pouille F, Scanziani M. Routing of spike series by dynamic circuits in the hippocampus. Nature. 2004;429:717–723. doi: 10.1038/nature02615. [DOI] [PubMed] [Google Scholar]
- Pouille F, Watkinson O, Scanziani M, Trevelyan AJ. The contribution of synaptic location to inhibitory gain control in pyramidal cells. Physiological reports. 2013;1:e00067. doi: 10.1002/phy2.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Povysheva NV, Zaitsev AV, Kroner S, Krimer OA, Rotaru DC, Gonzalez-Burgos G, Lewis DA, Krimer LS. Electrophysiological differences between neurogliaform cells from monkey and rat prefrontal cortex. Journal of neurophysiology. 2007;97:1030–1039. doi: 10.1152/jn.00794.2006. [DOI] [PubMed] [Google Scholar]
- Povysheva NV, Zaitsev AV, Rotaru DC, Gonzalez-Burgos G, Lewis DA, Krimer LS. Parvalbumin-positive basket interneurons in monkey and rat prefrontal cortex. Journal of neurophysiology. 2008;100:2348–2360. doi: 10.1152/jn.90396.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pow DV, Morris JF. Dendrites of hypothalamic magnocellular neurons release neurohypophysial peptides by exocytosis. Neuroscience. 1989;32:435–439. doi: 10.1016/0306-4522(89)90091-2. [DOI] [PubMed] [Google Scholar]
- Price CJ, Cauli B, Kovacs ER, Kulik A, Lambolez B, Shigemoto R, Capogna M. Neurogliaform neurons form a novel inhibitory network in the hippocampal CA1 area. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:6775–6786. doi: 10.1523/JNEUROSCI.1135-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price CJ, Scott R, Rusakov DA, Capogna M. GABA(B) receptor modulation of feedforward inhibition through hippocampal neurogliaform cells. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2008;28:6974–6982. doi: 10.1523/JNEUROSCI.4673-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pronneke A, Scheuer B, Wagener RJ, Mock M, Witte M, Staiger JF. Characterizing VIP Neurons in the Barrel Cortex of VIPcre/tdTomato Mice Reveals Layer-Specific Differences. Cerebral cortex. 2015;25:4854–4868. doi: 10.1093/cercor/bhv202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reyes A, Lujan R, Rozov A, Burnashev N, Somogyi P, Sakmann B. Target-cell-specific facilitation and depression in neocortical circuits. Nature neuroscience. 1998;1:279–285. doi: 10.1038/1092. [DOI] [PubMed] [Google Scholar]
- Rossignol E, Kruglikov I, van den Maagdenberg AM, Rudy B, Fishell G. CaV 2.1 ablation in cortical interneurons selectively impairs fast-spiking basket cells and causes generalized seizures. Annals of neurology. 2013;74:209–222. doi: 10.1002/ana.23913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roux L, Buzsaki G. Tasks for inhibitory interneurons in intact brain circuits. Neuropharmacology. 2015;88:10–23. doi: 10.1016/j.neuropharm.2014.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Royer S, Zemelman BV, Losonczy A, Kim J, Chance F, Magee JC, Buzsaki G. Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nature neuroscience. 2012;15:769–775. doi: 10.1038/nn.3077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozov A, Jerecic J, Sakmann B, Burnashev N. AMPA receptor channels with long-lasting desensitization in bipolar interneurons contribute to synaptic depression in a novel feedback circuit in layer 2/3 of rat neocortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2001;21:8062–8071. doi: 10.1523/JNEUROSCI.21-20-08062.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudy B, Chow A, Lau D, Amarillo Y, Ozaita A, Saganich M, Moreno H, Nadal MS, Hernandez-Pineda R, Hernandez-Cruz A, et al. Contributions of Kv3 channels to neuronal excitability. Annals of the New York Academy of Sciences. 1999;868:304–343. doi: 10.1111/j.1749-6632.1999.tb11295.x. [DOI] [PubMed] [Google Scholar]
- Rudy B, McBain CJ. Kv3 channels: voltage-gated K+ channels designed for high-frequency repetitive firing. Trends in neurosciences. 2001;24:517–526. doi: 10.1016/s0166-2236(00)01892-0. [DOI] [PubMed] [Google Scholar]
- Runyan CA, Schummers J, Van Wart A, Kuhlman SJ, Wilson NR, Huang ZJ, Sur M. Response features of parvalbumin-expressing interneurons suggest precise roles for subtypes of inhibition in visual cortex. Neuron. 2010;67:847–857. doi: 10.1016/j.neuron.2010.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Runyan CA, Sur M. Response selectivity is correlated to dendritic structure in parvalbumin-expressing inhibitory neurons in visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:11724–11733. doi: 10.1523/JNEUROSCI.2196-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scanziani M. GABA spillover activates postsynaptic GABA(B) receptors to control rhythmic hippocampal activity. Neuron. 2000;25:673–681. doi: 10.1016/s0896-6273(00)81069-7. [DOI] [PubMed] [Google Scholar]
- Scanziani M, Gahwiler BH, Charpak S. Target cell-specific modulation of transmitter release at terminals from a single axon. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:12004–12009. doi: 10.1073/pnas.95.20.12004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiff ML, Reyes AD. Characterization of thalamocortical responses of regular-spiking and fast-spiking neurons of the mouse auditory cortex in vitro and in silico. Journal of neurophysiology. 2012;107:1476–1488. doi: 10.1152/jn.00208.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silberberg G. Polysynaptic subcircuits in the neocortex: spatial and temporal diversity. Current opinion in neurobiology. 2008;18:332–337. doi: 10.1016/j.conb.2008.08.009. [DOI] [PubMed] [Google Scholar]
- Silberberg G, Markram H. Disynaptic inhibition between neocortical pyramidal cells mediated by Martinotti cells. Neuron. 2007;53:735–746. doi: 10.1016/j.neuron.2007.02.012. [DOI] [PubMed] [Google Scholar]
- Sillito AM. The contribution of inhibitory mechanisms to the receptive field properties of neurones in the striate cortex of the cat. The Journal of physiology. 1975;250:305–329. doi: 10.1113/jphysiol.1975.sp011056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver RA. Neuronal arithmetic. Nature reviews Neuroscience. 2010;11:474–489. doi: 10.1038/nrn2864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmons ML, Terman GW, Gibbs SM, Chavkin C. L-type calcium channels mediate dynorphin neuropeptide release from dendrites but not axons of hippocampal granule cells. Neuron. 1995;14:1265–1272. doi: 10.1016/0896-6273(95)90273-2. [DOI] [PubMed] [Google Scholar]
- Simon A, Olah S, Molnar G, Szabadics J, Tamas G. Gap-junctional coupling between neurogliaform cells and various interneuron types in the neocortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:6278–6285. doi: 10.1523/JNEUROSCI.1431-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simons DJ. Response properties of vibrissa units in rat SI somatosensory neocortex. Journal of neurophysiology. 1978;41:798–820. doi: 10.1152/jn.1978.41.3.798. [DOI] [PubMed] [Google Scholar]
- Simons DJ, Carvell GE. Thalamocortical response transformation in the rat vibrissa/barrel system. Journal of neurophysiology. 1989;61:311–330. doi: 10.1152/jn.1989.61.2.311. [DOI] [PubMed] [Google Scholar]
- Sohal VS, Zhang F, Yizhar O, Deisseroth K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature. 2009;459:698–702. doi: 10.1038/nature07991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohya K, Kameyama K, Yanagawa Y, Obata K, Tsumoto T. GABAergic neurons are less selective to stimulus orientation than excitatory neurons in layer II/III of visual cortex, as revealed by in vivo functional Ca2+ imaging in transgenic mice. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2007;27:2145–2149. doi: 10.1523/JNEUROSCI.4641-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somogyi J, Baude A, Omori Y, Shimizu H, El Mestikawy S, Fukaya M, Shigemoto R, Watanabe M, Somogyi P. GABAergic basket cells expressing cholecystokinin contain vesicular glutamate transporter type 3 (VGLUT3) in their synaptic terminals in hippocampus and isocortex of the rat. The European journal of neuroscience. 2004;19:552–569. doi: 10.1111/j.0953-816x.2003.03091.x. [DOI] [PubMed] [Google Scholar]
- Somogyi P, Kisvarday ZF, Martin KA, Whitteridge D. Synaptic connections of morphologically identified and physiologically characterized large basket cells in the striate cortex of cat. Neuroscience. 1983;10:261–294. doi: 10.1016/0306-4522(83)90133-1. [DOI] [PubMed] [Google Scholar]
- Somogyi P, Klausberger T. Defined types of cortical interneurone structure space and spike timing in the hippocampus. The Journal of physiology. 2005;562:9–26. doi: 10.1113/jphysiol.2004.078915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somogyi P, Tamas G, Lujan R, Buhl EH. Salient features of synaptic organisation in the cerebral cortex. Brain Res Brain Res Rev. 1998;26:113–135. doi: 10.1016/s0165-0173(97)00061-1. [DOI] [PubMed] [Google Scholar]
- Spruston N. Pyramidal neurons: dendritic structure and synaptic integration. Nature reviews Neuroscience. 2008;9:206–221. doi: 10.1038/nrn2286. [DOI] [PubMed] [Google Scholar]
- Staiger JF, Masanneck C, Schleicher A, Zuschratter W. Calbindin-containing interneurons are a target for VIP-immunoreactive synapses in rat primary somatosensory cortex. The Journal of comparative neurology. 2004;468:179–189. doi: 10.1002/cne.10953. [DOI] [PubMed] [Google Scholar]
- Staiger JF, Zilles K, Freund TF. Distribution of GABAergic elements postsynaptic to ventroposteromedial thalamic projections in layer IV of rat barrel cortex. The European journal of neuroscience. 1996;8:2273–2285. doi: 10.1111/j.1460-9568.1996.tb01191.x. [DOI] [PubMed] [Google Scholar]
- Stark E, Eichler R, Roux L, Fujisawa S, Rotstein HG, Buzsaki G. Inhibition-induced theta resonance in cortical circuits. Neuron. 2013;80:1263–1276. doi: 10.1016/j.neuron.2013.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stark E, Roux L, Eichler R, Senzai Y, Royer S, Buzsaki G. Pyramidal cell-interneuron interactions underlie hippocampal ripple oscillations. Neuron. 2014;83:467–480. doi: 10.1016/j.neuron.2014.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepanyants A, Martinez LM, Ferecsko AS, Kisvarday ZF. The fractions of short- and long-range connections in the visual cortex. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:3555–3560. doi: 10.1073/pnas.0810390106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stokes CC, Isaacson JS. From dendrite to soma: dynamic routing of inhibition by complementary interneuron microcircuits in olfactory cortex. Neuron. 2010;67:452–465. doi: 10.1016/j.neuron.2010.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki N, Bekkers JM. Distinctive classes of GABAergic interneurons provide layer-specific phasic inhibition in the anterior piriform cortex. Cerebral cortex. 2010;20:2971–2984. doi: 10.1093/cercor/bhq046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki N, Bekkers JM. Microcircuits mediating feedforward and feedback synaptic inhibition in the piriform cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2012;32:919–931. doi: 10.1523/JNEUROSCI.4112-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swadlow HA, Gusev AG. Receptive-field construction in cortical inhibitory interneurons. Nature neuroscience. 2002;5:403–404. doi: 10.1038/nn847. [DOI] [PubMed] [Google Scholar]
- Sylwestrak EL, Ghosh A. Elfn1 regulates target-specific release probability at CA1-interneuron synapses. Science (New York, NY) 2012;338:536–540. doi: 10.1126/science.1222482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabadics J, Tamas G, Soltesz I. Different transmitter transients underlie presynaptic cell type specificity of GABAA, slow and GABAA, fast. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:14831–14836. doi: 10.1073/pnas.0707204104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabadics J, Varga C, Molnar G, Olah S, Barzo P, Tamas G. Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits. Science (New York, NY) 2006;311:233–235. doi: 10.1126/science.1121325. [DOI] [PubMed] [Google Scholar]
- Taki K, Kaneko T, Mizuno N. A group of cortical interneurons expressing mu-opioid receptor-like immunoreactivity: a double immunofluorescence study in the rat cerebral cortex. Neuroscience. 2000;98:221–231. doi: 10.1016/s0306-4522(00)00124-x. [DOI] [PubMed] [Google Scholar]
- Tamamaki N, Tomioka R. Long-Range GABAergic Connections Distributed throughout the Neocortex and their Possible Function. Frontiers in neuroscience. 2010;4:202. doi: 10.3389/fnins.2010.00202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamas G, Lorincz A, Simon A, Szabadics J. Identified sources and targets of slow inhibition in the neocortex. Science (New York, NY) 2003;299:1902–1905. doi: 10.1126/science.1082053. [DOI] [PubMed] [Google Scholar]
- Tan Z, Hu H, Huang ZJ, Agmon A. Robust but delayed thalamocortical activation of dendritic-targeting inhibitory interneurons. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:2187–2192. doi: 10.1073/pnas.0710628105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taniguchi H, He M, Wu P, Kim S, Paik R, Sugino K, Kvitsiani D, Fu Y, Lu J, Lin Y, et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron. 2011;71:995–1013. doi: 10.1016/j.neuron.2011.07.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taniguchi H, Lu J, Huang ZJ. The spatial and temporal origin of chandelier cells in mouse neocortex. Science (New York, NY) 2013;339:70–74. doi: 10.1126/science.1227622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tasic B, Menon V, Nguyen TN, Kim TK, Jarsky T, Yao Z, Levi B, Gray LT, Sorensen SA, Dolbeare T, et al. Adult mouse cortical cell taxonomy revealed by single cell transcriptomics. Nature neuroscience. 2016;19:335–346. doi: 10.1038/nn.4216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taub AH, Katz Y, Lampl I. Cortical balance of excitation and inhibition is regulated by the rate of synaptic activity. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:14359–14368. doi: 10.1523/JNEUROSCI.1748-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomson AM. Presynaptic frequency- and pattern-dependent filtering. Journal of computational neuroscience. 2003;15:159–202. doi: 10.1023/a:1025812808362. [DOI] [PubMed] [Google Scholar]
- Thomson AM, Destexhe A. Dual intracellular recordings and computational models of slow inhibitory postsynaptic potentials in rat neocortical and hippocampal slices. Neuroscience. 1999;92:1193–1215. doi: 10.1016/s0306-4522(99)00021-4. [DOI] [PubMed] [Google Scholar]
- Thomson AM, Lamy C. Functional maps of neocortical local circuitry. Frontiers in neuroscience. 2007;1:19–42. doi: 10.3389/neuro.01.1.1.002.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomson AM, West DC, Hahn J, Deuchars J. Single axon IPSPs elicited in pyramidal cells by three classes of interneurones in slices of rat neocortex. The Journal of physiology. 1996;496(Pt 1):81–102. doi: 10.1113/jphysiol.1996.sp021667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomioka R, Sakimura K, Yanagawa Y. Corticofugal GABAergic projection neurons in the mouse frontal cortex. Frontiers in neuroanatomy. 2015;9:133. doi: 10.3389/fnana.2015.00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth K, Borhegyi Z, Freund TF. Postsynaptic targets of GABAergic hippocampal neurons in the medial septum-diagonal band of broca complex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1993;13:3712–3724. doi: 10.1523/JNEUROSCI.13-09-03712.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toyama K, Matsunami K, Ono T, Tokashiki S. An intracellular study of neuronal organization in the visual cortex. Experimental brain research. 1974;21:45–66. doi: 10.1007/BF00234257. [DOI] [PubMed] [Google Scholar]
- Tsumoto T, Eckart W, Creutzfeldt OD. Modification of orientation sensitivity of cat visual cortex neurons by removal of GABA-mediated inhibition. Experimental brain research. 1979;34:351–363. doi: 10.1007/BF00235678. [DOI] [PubMed] [Google Scholar]
- Urban-Ciecko J, Fanselow EE, Barth AL. Neocortical somatostatin neurons reversibly silence excitatory transmission via GABAb receptors. Current biology: CB. 2015;25:722–731. doi: 10.1016/j.cub.2015.01.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Pol AN. Neuropeptide transmission in brain circuits. Neuron. 2012;76:98–115. doi: 10.1016/j.neuron.2012.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varga C, Oijala M, Lish J, Szabo GG, Bezaire M, Marchionni I, Golshani P, Soltesz I. Functional fission of parvalbumin interneuron classes during fast network events. Elife. 2014;3 doi: 10.7554/eLife.04006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vu ET, Krasne FB. Evidence for a computational distinction between proximal and distal neuronal inhibition. Science (New York, NY) 1992;255:1710–1712. doi: 10.1126/science.1553559. [DOI] [PubMed] [Google Scholar]
- Wang Q, Webber RM, Stanley GB. Thalamic synchrony and the adaptive gating of information flow to cortex. Nature neuroscience. 2010;13:1534–1541. doi: 10.1038/nn.2670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Hooks BM, Sun QQ. Thorough GABAergic innervation of the entire axon initial segment revealed by an optogenetic ‘laserspritzer’. The Journal of physiology. 2014;592:4257–4276. doi: 10.1113/jphysiol.2014.275719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Gupta A, Toledo-Rodriguez M, Wu CZ, Markram H. Anatomical, physiological, molecular and circuit properties of nest basket cells in the developing somatosensory cortex. Cerebral cortex. 2002;12:395–410. doi: 10.1093/cercor/12.4.395. [DOI] [PubMed] [Google Scholar]
- Wang Y, Toledo-Rodriguez M, Gupta A, Wu C, Silberberg G, Luo J, Markram H. Anatomical, physiological and molecular properties of Martinotti cells in the somatosensory cortex of the juvenile rat. The Journal of physiology. 2004;561:65–90. doi: 10.1113/jphysiol.2004.073353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wehr M, Zador AM. Balanced inhibition underlies tuning and sharpens spike timing in auditory cortex. Nature. 2003;426:442–446. doi: 10.1038/nature02116. [DOI] [PubMed] [Google Scholar]
- White EL, Keller A. Intrinsic circuitry involving the local axon collaterals of corticothalamic projection cells in mouse SmI cortex. The Journal of comparative neurology. 1987;262:13–26. doi: 10.1002/cne.902620103. [DOI] [PubMed] [Google Scholar]
- White EL, Rock MP. A comparison of thalamocortical and other synaptic inputs to dendrites of two non-spiny neurons in a single barrel of mouse SmI cortex. The Journal of comparative neurology. 1981;195:265–277. doi: 10.1002/cne.901950207. [DOI] [PubMed] [Google Scholar]
- Wilent WB, Contreras D. Dynamics of excitation and inhibition underlying stimulus selectivity in rat somatosensory cortex. Nature neuroscience. 2005;8:1364–1370. doi: 10.1038/nn1545. [DOI] [PubMed] [Google Scholar]
- Wilson NR, Runyan CA, Wang FL, Sur M. Division and subtraction by distinct cortical inhibitory networks in vivo. Nature. 2012;488:343–348. doi: 10.1038/nature11347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer VC, Bruno RM, de Kock CP, Kuner T, Sakmann B. Dimensions of a projection column and architecture of VPM and POm axons in rat vibrissal cortex. Cerebral cortex. 2010;20:2265–2276. doi: 10.1093/cercor/bhq068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodruff A, Xu Q, Anderson SA, Yuste R. Depolarizing effect of neocortical chandelier neurons. Frontiers in neural circuits. 2009;3:15. doi: 10.3389/neuro.04.015.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodruff AR, McGarry LM, Vogels TP, Inan M, Anderson SA, Yuste R. State-dependent function of neocortical chandelier cells. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2011;31:17872–17886. doi: 10.1523/JNEUROSCI.3894-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wozny C, Williams SR. Specificity of synaptic connectivity between layer 1 inhibitory interneurons and layer 2/3 pyramidal neurons in the rat neocortex. Cerebral cortex. 2011;21:1818–1826. doi: 10.1093/cercor/bhq257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu H, Jeong HY, Tremblay R, Rudy B. Neocortical somatostatin-expressing GABAergic interneurons disinhibit the thalamorecipient layer 4. Neuron. 2013;77:155–167. doi: 10.1016/j.neuron.2012.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu NL, Harnett MT, Williams SR, Huber D, O’Connor DH, Svoboda K, Magee JC. Nonlinear dendritic integration of sensory and motor input during an active sensing task. Nature. 2012;492:247–251. doi: 10.1038/nature11601. [DOI] [PubMed] [Google Scholar]
- Xu X, Callaway EM. Laminar specificity of functional input to distinct types of inhibitory cortical neurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:70–85. doi: 10.1523/JNEUROSCI.4104-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu X, Roby KD, Callaway EM. Mouse cortical inhibitory neuron type that coexpresses somatostatin and calretinin. The Journal of comparative neurology. 2006;499:144–160. doi: 10.1002/cne.21101. [DOI] [PubMed] [Google Scholar]
- Xu X, Roby KD, Callaway EM. Immunochemical characterization of inhibitory mouse cortical neurons: three chemically distinct classes of inhibitory cells. The Journal of comparative neurology. 2010;518:389–404. doi: 10.1002/cne.22229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue M, Atallah BV, Scanziani M. Equalizing excitation-inhibition ratios across visual cortical neurons. Nature. 2014;511:596–600. doi: 10.1038/nature13321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W, Carrasquillo Y, Hooks BM, Nerbonne JM, Burkhalter A. Distinct balance of excitation and inhibition in an interareal feedforward and feedback circuit of mouse visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:17373–17384. doi: 10.1523/JNEUROSCI.2515-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaitsev AV, Gonzalez-Burgos G, Povysheva NV, Kroner S, Lewis DA, Krimer LS. Localization of calcium-binding proteins in physiologically and morphologically characterized interneurons of monkey dorsolateral prefrontal cortex. Cerebral cortex. 2005;15:1178–1186. doi: 10.1093/cercor/bhh218. [DOI] [PubMed] [Google Scholar]
- Zaitsev AV, Povysheva NV, Gonzalez-Burgos G, Rotaru D, Fish KN, Krimer LS, Lewis DA. Interneuron diversity in layers 2–3 of monkey prefrontal cortex. Cerebral cortex. 2009;19:1597–1615. doi: 10.1093/cercor/bhn198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaitsev AV, Povysheva NV, Lewis DA, Krimer LS. P/Q-type, but not N-type, calcium channels mediate GABA release from fast-spiking interneurons to pyramidal cells in rat prefrontal cortex. Journal of neurophysiology. 2007;97:3567–3573. doi: 10.1152/jn.01293.2006. [DOI] [PubMed] [Google Scholar]
- Zeisel A, Munoz-Manchado AB, Codeluppi S, Lonnerberg P, La Manno G, Jureus A, Marques S, Munguba H, He L, Betsholtz C, et al. Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science (New York, NY) 2015;347:1138–1142. doi: 10.1126/science.aaa1934. [DOI] [PubMed] [Google Scholar]
- Zhang S, Xu M, Kamigaki T, Hoang Do JP, Chang WC, Jenvay S, Miyamichi K, Luo L, Dan Y. Selective attention. Long-range and local circuits for top-down modulation of visual cortex processing. Science (New York, NY) 2014;345:660–665. doi: 10.1126/science.1254126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou FM, Hablitz JJ. Morphological properties of intracellularly labeled layer I neurons in rat neocortex. The Journal of comparative neurology. 1996;376:198–213. doi: 10.1002/(SICI)1096-9861(19961209)376:2<198::AID-CNE3>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- Zhu Y, Stornetta RL, Zhu JJ. Chandelier cells control excessive cortical excitation: characteristics of whisker-evoked synaptic responses of layer 2/3 nonpyramidal and pyramidal neurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2004;24:5101–5108. doi: 10.1523/JNEUROSCI.0544-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zupanc GK. Peptidergic transmission: from morphological correlates to functional implications. Micron (Oxford, England: 1993) 1996;27:35–91. doi: 10.1016/0968-4328(95)00028-3. [DOI] [PubMed] [Google Scholar]