

Abstract
Promoters are structurally and functionally diverse gene regulatory regions. The presence or absence of sequence motifs and the spacing between the motifs defines the properties of promoters. Recent alternative promoter usage analyses in Drosophila melanogaster revealed that transposable elements significantly contribute to promote diversity. In this work, we analyzed in detail one of the transposable element insertions, named FBti0019985, that has been co-opted to drive expression of CG18446, a candidate stress response gene. We analyzed strains from different natural populations and we found that besides FBti0019985, there are another eight independent transposable elements inserted in the proximal promoter region of CG18446. All nine insertions are solo-LTRs that belong to the roo family. We analyzed the sequence of the nine roo insertions and we investigated whether the different insertions were functionally equivalent by performing 5’-RACE, gene expression, and cold-stress survival experiments. We found that different insertions have different molecular and functional consequences. The exact position where the transposable elements are inserted matters, as they all showed highly conserved sequences but only two of the analyzed insertions provided alternative transcription start sites, and only the FBti0019985 insertion consistently affects CG18446 expression. The phenotypic consequences of the different insertions also vary: only FBti0019985 was associated with cold-stress tolerance. Interestingly, the only previous report of transposable elements inserting repeatedly and independently in a promoter region in D. melanogaster, were also located upstream of a stress response gene. Our results suggest that functional validation of individual structural variants is needed to resolve the complexity of insertion clusters.
Author Summary
The presence of several transposable element insertions in the promoter region of a Drosophila melanogaster gene has only been described in heat shock protein genes. In this work, we have discovered and characterized in detail several naturally occurring independent transposable element insertions in the promoter region of a cold-stress response gene in the fruitfly Drosophila melanogaster. The nine transposable element insertions described are clustered in a small 368 bp region and all belong to the same family of transposable elements: the roo family. Each individual insertion is present at relatively low population frequencies, ranging from 1% to 17%. However, the majority of strains analyzed contain one of these nine roo insertions suggesting that this region might be evolving under positive selection. Although the sequence of these insertions is highly similar, their molecular and functional consequences are different. Only one of them, FBti0019985, is associated with increased viability in nonstress and in cold-stress conditions.
Introduction
Promoters are crucial regions for the transcriptional regulation of gene expression. Recent computational and experimental advances in functional genomics techniques have allowed defining the promoter architecture to an unprecedented level. Several core promoter motifs such as the Initiator (Inr) and the Downstream core Promoter Element (DPE) have been described, and it is likely that many others remain to be discovered. The presence or absence of the core promoter motifs influences enhancer-promoter communication and thus gene regulation [1]. Promoter regions also harbour transcription factor binding motifs, which are another important component in the regulation of gene expression [2]. Besides cis-regulatory elements that influence the temporal and spatial expression patterns of genes, proximal promoters often contain alternative transcription start sites (TSSs) [1, 3]. Rather than being “biological noise” from imprecise binding of the transcription initiation machinery, genome-wide analyses of TSSs usage showed that alternative TSSs play an important role in the diversification of gene expression patterns [4–8].
Transposable elements (TEs), long proposed to play an important role in gene regulation [9, 10], have recently been found to provide at least 1,300 alternative TSSs in the Drosophila melanogaster genome [8]. TEs can also add Transcription Factor Binding Sites (TFBSs) to the promoter of genes as has been recently shown in Drosophila and humans [11–13]. As a result of adding particular sequence elements, many TEs confer their intrinsic regulatory properties to nearby genes demonstrating that they distribute cis-regulatory modules [8]. Finally, TEs inserted in promoter regions can also influence gene expression by disrupting the promoter architecture. This is the case, for example, of naturally occurring P-element insertions in the promoter of heat shock protein (hsp) genes [14].
One of the TEs identified as providing an alternative TSS by Batut et al (2013) [8], named FBti0019985, was previously reported in a screening designed to identify putatively adaptive TE insertions in D. melanogaster [15]. However, this particular TE was not further studied because its population frequency could not be accurately determined [15]. FBti0019985 is a roo solo-LTR inserted in the 5’-UTR of CG18446 gene, which is nested in the first intron of crossbronx (cbx) (Fig 1). TEs from the roo family have long been proposed to affect the expression of nearby genes by adding and distributing cis-regulatory regions [16–19]. Specifically, roo LTRs contain several TFBSs and the Inr sequence characteristic of core promoters [8, 20].
Fig 1. FBti0019985 is inserted in the first intron of cbx gene and it overlaps with CG18446 5’-UTR region.

Schematic representation of the genomic region where FBti0019985 is inserted: chromosome 2R: 9,864,510–9,875,072. FBti0019985 is shown in red. Black boxes represent exons and white boxes represent the 5’-UTRs and the 3’-UTRs. Primers used to check for the presence/absence of FBti0019985 are depicted as black arrows (FL, R, and L; see Materials and Methods).
Interestingly, CG18446 has been identified as a candidate gene for cold resistance: it is upregulated in fly strains that have been selected for increased cold resistance compared with control strains that were not subjected to cold-stress [21]. Cold resistance is an ecologically and evolutionarily relevant trait because it influences the ability of the species to adapt to different climatic conditions and thus, their geographical distribution [22, 23]. There is good evidence suggesting that D. melanogaster adapts to cold environments and a growing list of candidate genes involved in this thermotolerance phenotype is being identified [21, 24–28]. However, the molecular variants responsible for the adaptive cold-stress resistance phenotype remain elusive [29].
In this work, we further analyzed the presence/absence of FBti0019985 in four natural populations of D. melanogaster. We found that besides FBti0019985, eight other roo elements have inserted in a 368 bp region around CG18446 transcript start site. These roo elements differ in the insertion site and in their orientation. On the other hand, all elements have the same size and show high sequence conservation: all cis-regulatory elements previously described in roo LTRs are highly conserved [8, 30]. We further investigated whether these different insertions were functionally equivalent by performing 5’-RACE, gene expression, and phenotypic analyses. Our results showed that the functional consequences of the different roo insertions depend on the particular position where the element is inserted. Among the nine different roo solo-LTR insertions, only FBti0019985 is consistently associated with increased viability in nonstress and cold-stress conditions across genetic backgrounds.
Results
Besides FBti0019985, eight other roo solo-LTRs are inserted in the promoter region of CG18446
We first aimed at estimating the frequency of FBti0019985 in non-African natural D. melanogaster populations. Thus, we checked using PCR whether this insertion was present, polymorphic, or absent in 28 strains from a natural population collected in North Carolina (North America, DGRP strains [31, 32]) and in 15 strains from a natural population collected in Bari (Italy, Europe [33]) (Table 1). We obtained PCR results for 39 of the 43 strains tested: nine strains produced PCR bands consistent with FBti0019985 being present, five strains appeared as heterozygous, 13 strains showed unexpected band patterns, and 12 strains appeared as absent (Table 1) (see Material and Methods). To verify these results, we sequenced 32 of the 39 strains including all the strains that showed some evidence of presence (Table 1).
Table 1. The nine roo solo-LTR insertions analyzed in this work.
| Insertion | Fly strain | PCR results | Sequenced band | Insertion positiona |
|---|---|---|---|---|
| FBti0019985 | RAL-639 | Present | FL-R | Reference position |
| RAL-802 | Present | FL-R / L-R | Reference position | |
| RAL-810 | Present | FL-R | Reference position | |
| IV68 | Present | FL-R / L-R | Reference position | |
| roo+7 | RAL-405 | Present | FL-R | + 7 bp |
| RAL-887 | Present | FL-R | + 7 bp | |
| RAL-911 | Present | FL-R | +7 bp | |
| RAL-441b | Larger L-R | FL-R | + 7 bp | |
| Larger FL-R | ||||
| RAL-801b | Larger L-R | FL-R | + 7 bp | |
| Larger FL-R | ||||
| roo+175 | IV145 | Heterozygous | Larger FL-R | + 175 bp |
| roo+278 | RAL-502 | Smaller L-R | L-R | + 278 bp |
| No FL-R | ||||
| roo-19 | IV42 | Present | FL-R | - 19 bp inverted |
| IV127 | Present | FL-R | - 19 bp inverted | |
| roo-28 | IV40 | Heterozygous | Larger FL-R | - 28 bp inverted |
| roo-44 | RAL-195 | Only FL-R | FL-R | - 44 bp inverted |
| RAL-383 | Only FL-R | FL-R | - 44 bp inverted | |
| roo-68 | RAL-75 | Only FL-R | FL-R | - 68 bp inverted |
| RAL-716 | Only FL-R | FL-R | - 68 bp inverted | |
| IV69 | Heterozygous | Larger FL-R | - 68 bp inverted | |
| roo-90 | RAL-21 | Larger L-R | FL-R | - 90 bp |
| RAL-88 | Larger L-R | FL-R | - 90 bp | |
| RAL-177 | Larger L-R | FL-R | - 90 bp | |
| RAL-737 | Larger L-R | FL-R | - 90 bp | |
| RAL-820 | Larger L-R | FL-R / L-R | - 90 bp | |
| RAL-857 | Heterozygous | FL-R / L-R | - 90 bp | |
| IV50 | Heterozygous | Larger FL-R | - 90 bp | |
| Absent | RAL-40 | Smaller L-R | FL-R / L-R | Absent |
| RAL-371 | Absent | FL-R | Absent | |
| RAL-391 | Absent | FL-R | Absent | |
| RAL-508 | Absent | NS | Absent | |
| RAL-783 | Absent | FL-R | Absent | |
| RAL-822 | Absent | NS | Absent | |
| RAL-855 | Absent | NS | Absent | |
| RAL-908 | Absent | FL-R | Absent | |
| IV22 | Absent | FL-R | Absent | |
| IV49 | Absent | NS | Absent | |
| IV52 | Absent | NS | Absent | |
| IV72 | Absent | NS | Absent | |
| IV75 | Absent | NS | Absent | |
| No data | RAL-776 | No results | ||
| IV33 | No results | |||
| IV125 | No results | |||
| IV148 | No results |
NS, not sequenced
a "+" indicates the insertion is downstream of FBti0019985 and "-" indicates the insertion is upstream of FBti0019985
bThese strains have a 95 bp duplication upstream of the insertion
Only four of the nine strains classified as present, according to the PCR results, had the FBti0019985 insertion. For the rest of this work, we considered the position where FBti0019985 is inserted as the "reference position". The other five present strains, the five heterozygous strains, and 12 of the 13 strains that gave unexpected PCR bands contained different roo solo-LTR insertions (Table 1). Overall, besides FBti0019985, we found eight other 428 bp roo solo-LTRs inserted in eight different positions (Fig 2). Three roo insertions are located downstream of the reference position: roo+7, roo+175, and roo+278 (Fig 2). Two of the four strains carrying roo+7 have a duplication of the 95 bp region located immediately upstream of the insertion (Table 1). roo+175 element is inserted in the 5’-UTR region, and roo+278 is inserted in the first exon of CG18446 gene. Both roo+175 and roo+278 have a conserved Inr motif. If transcription starts in these insertions, flies carrying roo+175 would have a 100 bp shorter 5'-UTR, and flies carrying roo+278 would have a 35 amino acids shorter CG18446 protein. The other five roo insertions are located upstream of the reference position: roo-19, roo-28, roo-44, roo-68, roo-90 (Fig 2). Four of them, roo-19, roo-28, roo-44, and roo-68, are inserted in reverse orientation.
Fig 2. Besides FBti0019985, eight other roo solo-LTR are inserted in the proximal promoter of CG18446.

Schematic representation of the genomic region where the nine roo solo-LTRs are inserted. roo insertions are depicted as red triangles. White boxes represent CG18446 5’-UTR. Regions depicted with dotted lines are not drawn to scale. Target Site Duplications (TSDs) are shown in blue. NC, allele frequency (%) in the North American population; IT, allele frequency (%) in the Italian population; SW, allele frequency (%) in the Swedish population; Out-AF total, allele frequency (%) in all the out-of-Africa populations; ZI, allele frequency (%) in the Zambia population.
We used Tlex-2 software to further analyze the frequency of the nine roo insertions in 21 additional DGRP strains, in 26 strains from a Swedish natural population, and in 42 strains from a population collected in the ancestral range of the species, Zambia (Fig 2 and S1 Table) (see Material and Methods) [34]. Overall, we found that 67 strains, out of the 128 strains analyzed, contained one of the nine roo solo-LTR insertions. The two most common roo insertion in out-of-Africa populations are roo-90 and FBti0019985 present in 13% and 10% of the strains tested, respectively (Fig 2). Besides, some insertions are only present in the North Carolina natural population while others are specific to the Italian natural population (Fig 2). Only three of the nine insertions described in North Carolina and Italian populations are present in the Swedish population. However, we did not perform de novo discovery of TEs in this population. Thus, it could be that other private insertions are present in the Swedish population. Finally, all the nine insertions were present in the African population although most of them were present at very low frequencies (Fig 2).
In summary, we have found that besides the FBti0019985 insertion annotated in the reference genome, eight other 428 bp roo solo-LTRs are inserted nearby CG18446 TSS in natural populations of D. melanogaster (Fig 2) [35]. Each one of the strains analyzed contains a single solo-LTR roo insertion and most of the analyzed strains contain one of the nine solo-LTR roo insertions.
The nine roo solo-LTR are independent insertions that occurred at different evolutionary timepoints
We identified the Target Site Duplications (TSD) of the nine different roo insertions using data from the 26 present strains sequenced in this work (Table 1). We could identify the TSD for all roo insertions except for roo+278. We found that six of the eight TSDs identified are five nucleotides long as has been previously described for this family [36] (Fig 2). However, the TSD sequences did not match the proposed TSD consensus sequence [34, 36, 37]. We thus used all the available roo TSD sequences to build a new consensus (S1 Fig). The different roo solo-LTR insertions had different TSDs suggesting that they are independent insertions (Fig 2). Furthermore, all the roo elements located in a given insertion site have the same exact TSD and are inserted in the same orientation suggesting that each one of them is a unique insertion event (Fig 2).
To test whether these nine insertion events were the result of a burst of transposition, we constructed a phylogenetic tree. We included the nine roo insertions sequenced in this work and 115 other roo insertions present in the D. melanogaster genome (S2 Fig and S1 Text). We found that not all the newly described roo insertions clustered together suggesting that they did not insert at the same time (S2 Fig and S1 Text).
All the TEs identified in CG18446 proximal promoter region belong to the roo family. Thus, we also investigated whether roo elements annotated in the reference genome are preferentially inserted into gene proximal promoter regions as has been previously described for other TE families [38, 39]. We analyzed the 138 insertions belonging to the roo family annotated in the D. melanogaster reference genome (v5). We found 21 roo insertions located in the 1 kb region upstream of a gene or overlapping the 5’-end of a gene. Thus, only 15.2% of the roo elements in the D. melanogaster genome are located in gene promoters and/or 5’-UTRs.
In summary, TSD analyses of the nine insertions characterized in this work suggested that they are independent insertions, and confirmed the length but not the sequence previously reported as the TSD consensus for this family. Our results are not consistent with the nine roo insertions being the result of a single burst of transposition. Finally, our analyses also suggested that roo elements do not preferentially insert in 5’ gene regions.
The nine roo insertions add the same cis-regulatory sequences
We analyzed multiple sequence alignments of all the roo insertions located nearby CG18446. We identified TFBSs using the JASPAR database (see Material and Methods). We also specifically looked for conservation of the regulatory regions previously described in the roo family [8, 30], and for conserved core promoter motifs [1] (Fig 3A and S2A Table). Overall, there was very little diversity among the nine solo-LTRs (S3A Fig). The five TFBSs and the Inr sequence previously identified in the consensus sequence of roo LTRs are conserved in all the roo copies located in the proximal promoter of CG18446 [8]. Additionally, we found another four TFBSs that are also highly conserved in all the copies (Fig 3A and S3A Fig). The nine transcription factors are involved in developmental processes. Additionally, Deaf1 and Nub are also involved in immune response [40, 41]. Finally, three previously identified Matrix Associated Regions (MARs) in LTRs from the roo family are also highly conserved in the nine insertions (Fig 3A and S3B Fig) [30]. These results suggest that these roo solo-LTR insertions are introducing the same cis-regulatory regions in the CG18446 proximal promoter region. Still, the functional effect of these insertions might be different because they are located in different positions and have different orientations (Fig 2).
Fig 3. Conserved regulatory regions in the the nine roo solo-LTR insertions and in the proximal promoter region of CG18446.

(A) Location of the nine transcription factor binding sites (green boxes), the Inr motif (blue box), and regions with matrix association potential (MARs) (black boxes), in the roo solo-LTR consensus sequence. Deaf1, ara, mirr and caup TFBS have been identified in this work. (B) Location of the eight transcription factor binding sites (green boxes) and the two core promoter motifs (blue boxes) in the proximal promoter region of CG18446. Different roo insertions are depicted as red triangles. The positions of roo+175 and roo+278 are not drawn to scale.
roo insertions affect the spacing of Transcription Factor Binding Sites in the proximal promoter region of CG18446
We analyzed the proximal promoter region of CG18446 in the 30 strains sequenced in this work. We could not identify the TATA box suggesting that CG18446 has a DPE promoter [1]. We identified eight TFBSs in the proximal promoter of CG18446 (Fig 3B and S2C Table). These eight TFBSs are highly conserved in all the strains analyzed (S3C Fig). The different roo insertions characterized in this work do not disrupt any of the identified core promoter motifs or TFBSs (Fig 3B). However, they do affect the spacing between the different regulatory motifs, which might affect the protein-protein interaction at the CG18446 promoter and thus the expression level of this gene (Fig 3B) [14].
roo insertions could be recruiting the HP1a protein
Besides affecting the spacing of transcription factor binding site, another mechanism by which roo insertions could be affecting CG18446 expression is by recruiting piRNAs that would lead to heterochromatin formation [42, 43]. We mapped piRNA reads from three different available libraries to a 1.4 kb region including FBti0019985 (Fig 4A) (see Material and Methods) [44–46]. We found that most of the piRNAs mapping to the insertion were sense reads, suggesting that FBti0019985 is not acting as a target for heterochromatin assembly [42].
Fig 4. Mapping of piRNA reads and HP1a reads to the FBti0019985 region.

(A) Number of piRNA reads mapped to a 1.4 kb region including FBti0019985. aLi et al (2009) piRNA library, bSatyaki et al (2014) piRNA library and cShpiz et al (2014) piRNA library. (B) Number of HP1a reads mapped to the same 1.4 kb region.
We also looked for evidence of HP1a binding to FBti0019985 using modENCODE data (see Material and Methods) [47]. HP1a is a structural chromosomal protein that mediates both gene expression and gene silencing [48]. We did find evidence of HP1a reads binding to FBti0019985 (Fig 4B). Thus, by recruiting HP1a, FBti0019985 could be affecting the expression of CG18446. The same results were obtained for the other eight roo solo-LTR insertions: most of the piRNAs mapping to the insertions were sense reads and we found evidence of HP1a binding to all of them (S3 Table). Overall, our results are suggestive but not conclusive of HP1a binding to the nine roo insertions described in this work.
To further investigate the possible functional consequences of the roo insertions, we focused on the five insertions present at higher population frequencies in out-of-Africa populations: FBti0019985, roo+7, roo-44, roo-90, and roo-68 (Fig 2).
Only FBti0019985 and roo+7 affect the transcription start site of CG18446
We investigated whether roo insertions could be providing an alternative TSS to CG18446. Batut et al (2013) [8] reported that the TSS of CG18446 is located inside FBti0019985. However, this finding was obtained using RAMPAGE and was not further validated using 5’-RACE. For this reason, we performed a 5’-RACE with the RAL-810 strain that carries FBti0019985 and with the RAL-783 strain that does not carry any of the nine roo solo-LTR insertions. As expected, we found that the TSS of CG18446 is inside the TE: the first 50 bp of the 276 bp 5’-UTR correspond to FBti0019985 (Fig 5). Additionally, flies with the insertion have also a shorter transcript, with a 201 bp 5’-UTR, that does not start in FBti0019985 (Fig 5). Most of the sequenced transcripts start in the FBti0019985 insertion (14 out of 20 transcripts analyzed). Flies without the FBti0019985 insertion only have the 201 bp 5’-UTR transcript (Fig 5).
Fig 5. FBti0019985 and roo+7 affect the transcription start site of CG18446.
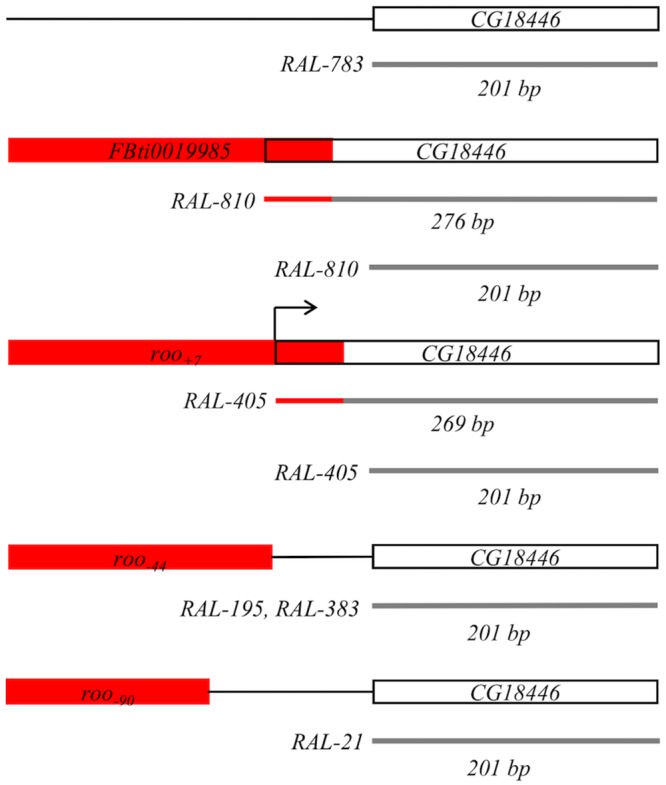
Schematic representation of the results obtained using the 5’-RACE technique. Red boxes represent different roo insertions and white boxes represent CG18446 5’-UTRs. Partial transcripts obtained by 5’-RACE are depicted as grey lines. The region of the transcript that overlaps with a roo insertion is shown as a red line. The last 50 bp of FBti0019985 and roo+7 are included in the 5’-UTR of CG18446.
We then checked whether roo+7, located only 7 bp downstream of FBti0019985, roo-90, which is the most distal insertion, and roo-44, which is inserted in reversed orientation, also provide an alternative TSS to CG18446. We found that roo+7 affects the TSS of CG18446 (Fig 5). Indeed, the TSS in roo+7 is in the same nucleotide position as in FBti0019985. Thus, CG18446 transcript in flies with roo+7 is 7 bp shorter compared with the transcript in flies with FBti0019985. Similarly to FBti0019985, most of the sequenced transcripts started in the roo+7 insertion (18 out of 22 transcripts analyzed). On the other hand, we did not find evidence of a TSS inside roo-90, which might indicate that the distance of the TE to the nearby gene affects its ability to provide an alternative TSS (Fig 5). Finally, we analyzed two different strains carrying the roo-44 insertion in the same position and we could not find evidence for a transcript with the TSS in roo-44 (Fig 5).
Overall, we found that only FBti0019985 and roo+7 insertions modify the length of CG18446 transcript. These two roo insertions are located a few nucleotides from the gene and both are inserted in 5’ to 3’ orientation.
FBti0019985 is associated with changes in embryonic CG18446 expression
We further analyzed whether different roo insertions were associated with changes in CG18446 expression in embryos, where this gene is highly expressed [49]. For FBti0019985, we analyzed the expression of CG18446 in flies with four different genetic backgrounds. In three of the four backgrounds, FBti0019985 is associated with upregulation of CG18446 (Fig 6A). This result is significant in two genetic backgrounds, RAL-810 and IV68, and marginally significant in a third background, RAL-639 (t-test p-value = 0.045, p-value = 0.005 and p-value = 0.062, respectively) (Fig 6A). On the other hand, only in one of the three genetic backgrounds analyzed for roo+7, the insertion is associated with downregulation of this gene (t-test p-value = 0.015 for RAL-405) (Fig 6B).
Fig 6. FBti0019985 is associated with changes in CG18446 expression.

Normalized CG18446 expression level relative to Act5C in embryos without roo insertion (grey) and in embryos with different roo insertions (red). (A) For FBti0019985, we compared the expression of CG18446 in flies with four different genetic backgrounds. In three backgrounds, the presence of FBti0019985 was associated with CG18446 upregulation. These results were significant in two backgrounds, RAL-810 and IV68, and marginally significant in the third background, RAL-639. (B) roo+7 was only associated with changes of expression in one of the three backgrounds analyzed: RAL-405. (C) roo-90 was also only associated with changes of expression in one of the three backgrounds analyzed: RAL-21. (D) Finally, roo-44 was not associated with changes in expression in any of the two backgrounds analyzed. Error bars represent the standard error of the mean (SEM) for the three biological replicates performed for each experiment.
We also checked the expression of CG18446 in flies with two roo solo-LTR insertions that do not provide an alternative TSS to this gene: roo-90 and roo-44. We found that roo-90 is only associated with CG18446 upregulation in one of the three backgrounds analyzed (p-value = 0.001, for RAL-21) (Fig 6C). Two different strains with the roo-44 solo-LTR insertion did not show differences in the level of expression of CG18446 compared with strains without the insertion (p-values > 0.05 in both cases) (Fig 6D).
Overall, we found that FBti0019985 is associated with CG18446 upregulation in three of the four backgrounds analyzed (Fig 6A). In the majority of strains, roo+7, roo-90, and roo-44 are not associated with changes in CG18446 expression level (Fig 6B–6D). However, we can not discard that the presence of these insertions is associated with changes in the expression of CG18446 in other developmental stages and/or in tissues not analyzed in this work.
FBti0019985 is associated with increased viability in nonstress and in cold-stress conditions
We have shown that FBti0019985 affects the transcript length and it is associated with upregulation of CG18446 in most of the genetic backgrounds analyzed (Figs 5 and 6A). Because CG18446 has been previously identified as a cold-stress candidate gene, we tested whether flies with and without FBti0019985 differed in their sensitivity to cold-stress [21]. We first compared RAL-810, which carries FBti0019985, with RAL-783, which does not carry any of the nine roo insertions (Fig 7A). We performed three biological replicates. ANOVA analyses showed that the experimental condition (nonstress or cold-stress) and the insertion genotype (presence or absence of FBti0019985) were significant (Table 2). Flies with FBti0019985 had a higher viability than flies without this insertion in both nonstress and cold-stress conditions. Furthermore, the interaction between these two factors was also significant suggesting that the effect of the insertion is larger in cold-stress conditions (Fig 7A and Table 2).
Fig 7. Flies with FBti0019985 showed increased egg-to-adult viability under nonstress and under cold-stress conditions in three different genetic backgrounds.

Egg-to-adult viability of strains without FBti0019985 (grey) and with the FBti0019985 insertion (red) in nonstress (control) and in cold-stress conditions. Results of the three replicates performed with (A) RAL-783 and RAL-810, (B) RAL-908 and RAL-802, and (C) IV22 and IV68. Error bars represent the SEM of the different vials analyzed in each experiment.
Table 2. ANOVA for cold-stress assays in flies with and without different roo solo-LTR insertions.
| Two-way ANOVA | |||||||
|---|---|---|---|---|---|---|---|
| Experimental condition | Insertion genotype | Experimental condition * Insertion genotype | |||||
| Insertion | Strains | P-value | Effect sizea | P-value | Effect sizea | P-value | Effect sizea |
| FBti0019985 | RAL-810 (FBti0019985) / | ≪0.001 | 0.69 | ≪0.001 | 0.13 | 0.034 | 0.04 |
| RAL-783 (roo-) | |||||||
| RAL-802 (FBti0019985) / | ≪0.001 | 0.56 | ≪0.001 | 0.57 | 0.932 | - | |
| RAL-908 (roo-) | |||||||
| IV68 (FBti0019985) / | ≪0.001 | 0.74 | 0.003 | 0.25 | 0.981 | - | |
| IV22 (roo-) | |||||||
| roo+7 | RAL-405 (roo+7) / | ≪0.001 | 0.75 | 0.001 | 0.59 | 0.497 | - |
| RAL-783 (roo-) | |||||||
| RAL-911 (roo+7) / | ≪0.001 | 0.76 | 0.530 | - | 0.220 | - | |
| RAL-783 (roo-) | |||||||
| roo-90 | RAL-21 (roo-90) / | ≪0.001 | 0.88 | 0.358 | - | 0.118 | - |
| RAL-783 (roo-) | |||||||
| RAL-820 (roo-90) / | ≪0.001 | 0.71 | 0.681 | - | 0.123 | - | |
| RAL-783 (roo-) | |||||||
| roo-44 | RAL-195 (roo-44) / | ≪0.001 | 0.79 | 0.038 | 0.31 | 0.027 | 0.35 |
| RAL-783 (roo-) | |||||||
| RAL-383 (roo-44) / | ≪0.001 | 0.95 | ≪0.001 | 0.76 | 0.991 | - | |
| RAL-783 (roo-) | |||||||
| roo-68 | RAL-75 (roo-68) / | ≪0.001 | 0.66 | 0.505 | - | 0.004 | 0.51 |
| RAL-783 (roo-) | |||||||
| RAL-716 (roo-68) / | ≪0.001 | 0.87 | 0.002 | 0.56 | 0.032 | 0.33 | |
| RAL-783 (roo-) | |||||||
aPartial eta-squared values calculated as a measure of effect size.
We repeated the experiment using flies with different genetic backgrounds: RAL-802 that carries FBti0019985 and RAL-908 that does not carry this insertion (Fig 7B). ANOVA analyses showed that the experimental condition and the insertion genotype are significant while the interaction between these two factors was not significant (Table 2). RAL-802 flies had a higher egg-to-adult viability in nonstress and in cold-stress conditions compared with flies without FBti0019985.
Finally, we tested whether flies from a different population, IV68 carrying FBti0019985 and IV22 without this particular insertion both collected in Italy, also showed significantly increased viability in nonstress and in cold-stress conditions (Fig 7C and Table 2). We found that IV68 flies had a higher viability than flies without the FBti0019985 insertion in both nonstress and cold-stress conditions (Table 2).
Overall, we found consistent results, across genetic backgrounds from two different natural populations, suggesting that flies with the FBti0019985 insertion are associated with increased viability compared to flies without this insertion in nonstress and in cold-stress conditions. In all cases, the effect of the presence of the insertion was either medium or large (Table 2). In one of the genetic backgrounds, the effect was larger under cold-stress conditions (Fig 7A) while no interaction between experimental condition and insertion genotype was found in the other two backgrounds (Fig 7B and 7C).
Other roo solo-LTR insertions in the proximal promoter of CG18446 are not consistently associated with cold-stress phenotypes
We further checked whether another four roo solo-LTR insertions described in this work are associated with cold-stress phenotypes. For each insertion, we compared the egg-to-adult viability of flies with two different genetic backgrounds with the egg-to-adult viability of RAL-783 that does not carry any of these insertions (Fig 8). In all cases, we performed ANOVA analyses to check whether the experimental conditions, insertion genotype, and/or the interaction between these two factors were significant (Table 2).
Fig 8. Other roo solo-LTR insertions are not consistently associated with cold-stress resistant phenotypes.

Egg-to-adult viability in nonstress (control) and in cold-stress conditions of the RAL-783 strain without any of the nine roo insertions (grey) and of different strains with roo insertions (red). (A) RAL-405 (roo+7), (B) RAL-911 (roo+7), (C) RAL-21 (roo-90), (D) RAL-820 (roo-90), (E) RAL-195 (roo-44), (F) RAL-383 (roo-44), (G) RAL- 75 (roo-68), and (H) RAL-716 (roo-68). Error bars represent the SEM of the different vials analyzed in each experiment.
We found that the experimental condition had a significant effect on egg-to-adult viability in all the strains tested (Table 2). On the other hand, the effect of the insertion was only significant in some of the genetic backgrounds (Table 2). Among strains that carry the roo+7 insertion, the insertion genotype had an effect only in one of the two backgrounds tested (Fig 8A and 8B and Table 2). RAL-405 flies with roo+7 insertion showed decreased viability (Fig 8A and Table 2). The presence/ absence of roo-90 did not have a significant effect on egg-to-adult viability (Fig 8C and 8D and Table 2). For roo-44, while the insertion genotype had a significant effect on the two backgrounds tested, results were not consistent. In one background, the presence of the insertion is associated with increased viability under cold-stress conditions and the interaction between the treatment and the insertion genotype is significant (Fig 8E and Table 2), while in the other background the presence of roo-44 is associated with decreased viability (Fig 8F and Table 2). Finally, the presence of roo-68 significantly affected viability in only one of the two backgrounds tested: RAL-716 flies carrying roo-68 showed decreased viability (Fig 8H and Table 2).
Overall our results suggested that the presence of roo+7, roo-90, roo-44, and roo-68 solo-LTR insertions reported in this work was not consistently associated with cold-stress phenotypes (Fig 8). These other insertions could have no phenotypic effect or could be involved in phenotypes not analyzed in this work.
Inference of selection in the region flanking the FBti0019985 insertion
We looked for evidence of positive selection in the 2 kb region flanking the FBti0019985 insertion. We analyzed the number of segregating sites (S) in this region and estimated Tajima´s D, iHS, nSL, H12 and XP-EHH (see Material and Methods). We found reduced diversity in the strains with FBti0019985: the number of segregating sites in this region is significantly smaller than the number of segregating sites found in 2 kb regions of chromosome 2R, where the FBti0019985 insertion is located (p-value = 0.015) (S4 Table). We also found that Tajima’s D was significantly negative in the 2 kb region where FBti0019985 is inserted, as expected if this region is under positive selection (p-value = 0.009) (S4 Fig and S4 Table). Finally, we also found significant values of iHS and H12 in the region flanking the FBti0019985 insertion (p-value = 0.048 and p-value = 0.023, respectively) (S5 Fig and S4 Table).
We also looked for evidence of selection taking into account not only the strains in which FBti0019985 is inserted, but all the strains that contain one of the nine roo insertions described in this work. In this case, only iHS showed a marginally significant value (p-value = 0.049) (S6 Fig).
Overall, our results suggest that the strains carrying FBti0019985 might be evolving under positive selection while the evidence for positive selection taking into account all the strains with one of the nine roo solo-LTRs, was only marginally significant.
Discussion
Besides FBti0019985, we have discovered eight other roo solo-LTR elements inserted in the 368 bp region nearby the TSS of the cold-stress response gene CG18446 (Fig 2) [21]. Each strain contained a single roo insertion and the population frequency of the different individual insertions varies from 1% to 17% (Fig 2). Full-length elements from the roo family are 8.7 kb long. Such long insertions in the proximal promoter of CG18446 located in the first intron of cbx, might be deleterious, which could explain why all the identified insertions were solo-LTR elements. In D. melanogaster, repeated insertions of TEs have only been described in the proximal promoters of a particular gene class: hsp genes [50]. The susceptibility of hsp genes to TE insertions was attributed to their peculiar chromatin architecture: constitutively decondensed chromatin and nucleosome-free regions [51, 52]. However, promoter regions of non-hsp genes with similar chromatin architecture are not targets for TE insertions suggesting that chromatin accessibility is not sufficient to explain the susceptibility of hsp genes to TE insertions [50]. From a functional point of view, the presence of TEs in the promoter regions of hsp genes has been suggested to allow a rapid gene expression response to unpredictable temperature changes [50]. Similarly, the presence of roo insertions in the promoter of CG18446 could also be enhancing the ability of this gene to respond to environmental challenges, although only one of the nine roo insertions was associated with cold-stress tolerance (see below). Interestingly, almost 100% of the insertions described in heat-shock genes are P-element insertions, and all the insertions described here are roo elements. P-elements preferentially insert in the 5' end of genes where they recognize a structural motif rather than a sequence motif [38, 39]. While 81% of P-elements insert in 5’ gene regions, our results showed that only 15.2% of the roo elements annotated in the reference genome are inserted in 5’ gene regions. Thus, with the data currently available, roo insertions do not seem to preferentially insert into 5’ gene regions although analyses of de novo insertions should shed more light on this issue.
Our results showed that the different roo elements inserted in the proximal promoter of CG18446 differ in their molecular and functional effects (Table 3). We found that the two insertions that are more closely located to CG18446, FBti0019985 and roo+7, provided an alternative TSS to this gene (Fig 5 and Table 3). However, only FBti0019985 is associated with upregulation of CG18446 expression (Fig 6 and Table 3). Besides providing an alternative TSS, the effect of the FBti0019985 insertion on CG18446 expression could be due to the addition of new regulatory regions (Fig 3A), to the disruption of the spacing of pre-existing ones (Fig 3B), and/or to the recruitment of HP1a protein that could also lead to changes in the expression of CG18446 (Fig 4B). Finally, we cannot discard that polymorphisms other than the presence/absence of the FBti0019985 insertion also affect the expression of CG18446.
Table 3. Summary of the experimental results obtained in fly strains with five different roo solo-LTR insertions.
| Insertion | Orientation | Strain | 5'-RACE | CG18446 expression | Effect of the insertion in egg-to-adult viability |
|---|---|---|---|---|---|
| FBti0019985 | 5' to 3' | RAL-810 | TSS inside TE | Upregulation | Increase |
| RAL-802 | - | No differences | Increase | ||
| IV68 | - | Upregulation | Increase | ||
| RAL-639 | - | Upregulation | - | ||
| roo+7 | 5' to 3' | RAL-405 | TSS inside TE | Downregulation | Decrease |
| RAL-911 | - | No differences | No differences | ||
| RAL-887 | - | No differences | - | ||
| roo-44 | 3' to 5' | RAL-195 | TSS outside TE | No differences | Increase |
| RAL-383 | TSS outside TE | No differences | Decrease | ||
| roo-68 | 3' to 5' | RAL-75 | - | - | No differences |
| RAL-716 | - | - | Decrease | ||
| roo-90 | 5' to 3' | RAL-21 | TSS outside TE | Upregulation | No differences |
| RAL-820 | - | No differences | No differences | ||
| RAL-88 | - | No differences | - |
We found that the FBti0019985 insertion, which is associated with increased CG18446 expression, is consistently associated with increased viability in nonstress and in cold-stress conditions (Fig 7 and Table 3). Although we cannot exclude that other variants linked to FBti0019985 contribute to the increased viability phenotypes, we argue that it is unlikely that the association between the FBti0019985 insertion and increased viability in three different genetic backgrounds from two different natural populations would occur spuriously [53]. These results also suggest that CG18446 is likely to play a role in cold tolerance as was previously suggested based on cold-stress selection experiments in which this gene was found to be overexpressed [21]. However, FBti0019985 is present in only 10% of the out-of-Africa natural strains analyzed in this work. Our screening was focused on three out-of-Africa populations, thus we cannot discard that FBti0019985 is present at higher frequencies in other populations. Alternatively, it is also possible that the relatively low frequency of FBti0019985 is due to negative fitness effects of this insertion on other phenotypes. Cold-stress resistance has been associated with decreased starvation resistance [54, 55] and reduced fecundity [56, 57]. Therefore, the benefit of flies carrying FBti0019985 in cold-stress conditions might be a cost, for example, when food resources are scarce.
While FBti0019985 has a consistent cold-stress tolerance phenotype, four other roo insertions also located on the proximal promoter of CG18446 did not (Fig 8 and Table 3). The insertion that is present at higher frequencies in out-of-Africa populations is roo-90 (Fig 2). However, this insertion is not associated with changes of expression of CG18446 in embryos (Fig 6) and was not found to be associated with cold-stress tolerance phenotypes (Fig 8C and 8D and Table 3). It could be that this insertion has no phenotypic effect. Alternatively, roo-90 could be affecting a phenotype other than cold tolerance. A recent update in FlyBase revealed that CG18446 is also an ethanol-regulated gene that could contribute to ethanol sensitivity or tolerance [58]. Another possibility is that roo-90 affects cbx. As the other roo insertion described in this work and CG18446 gene, roo-90 is inserted in the first intron of cbx which has been functionally classified as a defense response to bacterium and spermatogenesis gene [59] (Fig 1). Elucidating whether roo-90 has an adaptive effect is beyond the scope of this paper.
Overall, we did not find evidence of positive selection at the DNA level in the region where the nine roo solo-LTR elements are inserted. We did find evidence of reduced diversity in this region when only the strains containing FBti0019985 were considered (S4–S6 Figs and S4 Table). Further analyses with a bigger dataset of strains is needed in order to determine whether this region shows signals of positive selection at the DNA level.
In summary, our results showed that different TE insertions in the same gene promoter region might have different molecular and functional consequences. Thus, the description of complex regions, as the one reported in this work, should be followed by functional analysis of the structural variants if we want to elucidate which ones are functionally relevant.
Materials and Methods
Fly stocks
We used inbred strains from the Drosophila Genetic Reference Panel (DGRP [31, 32]) and isofemale strains from an Italian population collected in Castellana Grotte (Bari, Italy [33]) to perform the molecular and phenotypic assays.
Analysis of presence/absence by PCR of the nine solo-LTR roo insertions
We used a PCR approach to check for presence/ absence of FBti0019985 in 28 strains from the North Carolina population and in 15 strains from Italy. The primers used were FBti0019985_FL (5’-GGCATCATAAAACCGTTGAACAC-3’), FBti0019985_L (5’-AGTCCCTTAGTGGGAGACCACAG-3’) and FBti0019985_R (5’-CGTAGGATCAGTGGGTGAAAATG-3’) (Fig 1). Primers FBti0019985_L and FBti0019985_R are expected to give a 616 bp band when the TE is present. Primers FBti0019985_FL and FBti0019985_R are expected to give a 638 bp band when the TE is absent and a 1066 bp band when the TE is present. All PCR bands giving evidence of presence and some of the PCR bands giving evidence of absence were cloned using TOPO TA Cloning Kit for Sequencing (Invitrogen) following the manufacturer’s instructions and Sanger-sequenced using M13 forward and/or M13 reverse primers to verify the results. Sequences have been deposited in GenBank under accession numbers KU672690-KU672720.
Analysis of the population frequencies of the nine roo solo-LTR insertions using Tlex2
We estimated the frequencies of the nine roo solo-LTR insertions described in this work using T-lex2 software [34]. Because T-lex2 works only for annotated TEs, we constructed eight new reference sequences including each one of the newly described roo solo-LTR insertions. The new reference sequences included 500 bp at each side of the TE and the TSD of each insertion.
We run T-lex2 in strains from three different populations: 50 strains from North Carolina (DGRP [31, 32]), 27 strains from a population collected in Stockholm, Sweden [33], and 67 strains from a population collected in Siavonga, Zambia [60]. As a control, we also run T-lex2 in the strains for which we have PCR results (S1 Table). We obtained results for 21 out of 50 DGRP strains, 26 out of 27 Swedish strains and 42 out of 67 Zambian strains. In some of the strains, T-lex2 detects more than one insertion per strain. However, PCR analyses of these strains revealed that only one insertion was present. These results suggest that T-lex2 cannot accurately estimate the frequency of insertion when they are closely located to each other. We thus discarded T-lex2 results indicating the presence of more than one insertion per strain. Other factors such as the quality of the reads and the coverage of the different strains could also be affecting T-lex2 results.
Analysis of target site motifs
Target site motifs were constructed in WebLogo (http://weblogo.berkeley.edu) using six TSDs sequences obtained in this work and 41 TSDs sequences predicted with T-lex2 software [34].
Phylogenetic analysis
For each roo solo-LTR insertion, we constructed a consensus sequence taking into account the 26 strains sequenced in this work using Sequencher 5.0 software. We aligned the nine roo insertion consensus sequences with 115 of the 137 other roo insertions present in the D. melanogaster genome using the multiple sequence aligner program MAFFT [61]. The quality sequence of the other 22 roo insertions was too low to include them in the alignment. A maximum likelihood tree was inferred using RAxML Version 8 [62] under the general time-reversible nucleotide model and a gamma distribution of evolutionary rates. We use the ETE toolkit Python framework for the analysis and visualization of trees [63].
roo insertions and CG18446 promoter sequence analysis
We looked for conservation of the Transcription Factor Binding Sites (TFBSs) previously described in the roo family [8] in all the roo solo-LTRs characterized in this work. First, we downloaded from FlyBase version r6.06 (http://flybase.org) the fasta file of FBti0019985 sequence (genome region 2R: 9,871,090–9,871,523). We also searched for TFBSs in the roo insertions and in the CG18446 promoter regions using all the available JASPAR CORE Insecta matrices (http://jaspar.genereg.net). Only those sites predicted with a relative score higher than 0.995 were considered. We identified four new TFBS in FBti0019985 sequence: Deaf1, ara, mir, and caup. We then look for conservation of the identified motifs in all the roo solo-LTR sequences described in this work. For some strains, we used the information available in http://popdrowser.uab.cat [64].
Detection of piRNA reads
We used three piRNA libraries [44–46] to map piRNA reads to a 1.4 kb region including FBti0019985 and to all the roo insertions described in this work following the methodology described in Ullastres et al (2015) [33]. Briefly, we used BWA-MEM package version 0.7.5 a-r405 [65] to align the reads and then we used SamTools and BamTools [66] to index and filter by sense/antisense reads. The total read density was obtained using R (Rstudio v0.98.507) [67].
Detection of HP1a binding sites
We used modENCODE ChIP-Seq data [47] to map HP1a reads to a 1.4 kb region including FBti0019985 and to all the roo insertions described in this work following the methodology described in Ullastres et al (2015) [33]. We aligned the reads using BWA-MEM package version 0.7.5 a-r405 [65]. The total read density was obtained using R (Rstudio v0.98.507) [67].
5’-RACE experiments
5-to-7 day-old flies were placed in a fly cage with egg-laying medium (2% agar with apple juice and a piece of fresh yeast) during 4 hours. Then, adult flies were separated and embryos were collected following the suspension method described in Schou (2013) [68]. Embryo dechorionation was done by bleach (50%) immersion. Total RNA was extracted using TRIzol Plus RNA Purification Kit (Ambion). RNA was then treated on-column with DNase I (Thermo) during purification, and then treated once more after purification. 5’-RACE was performed with FirstChoice RLM-RACE Kit and using Small-scale reaction RNA processing with RNA samples of RAL-783 (roo-), RAL-810 (FBti0019985), RAL-405 (roo+7), RAL-21 (roo-90), RAL-383 (roo-44) and RAL-195 (roo-44). The gene specific outer primer was 5’-GACACTCTTCGGTTGGTGGA-3’ and the gene specific inner primer was 5’-ACAACTGTTCTGTAGGATCGC-3’. The control primer was 5’-TAGTCCGCAGAGAAACGTCG-3’. Inner PCR products were then cloned and Sanger-sequenced as mentioned above. Sequences have been deposited in GenBank under accession numbers KU672721-KU672722.
Quantitative RT-PCR expression analysis
Embryo collection and RNA extraction was performed as described before. Reverse transcription was carried out using 500 ng of total RNA using Transcriptor First Strand cDNA Synthesis Kit (Roche). The cDNA was then used in a 1/50 dilution for qRT-PCR with SYBR green master-mix (Bio-Rad) on an iQ5 Thermal cycler. CG18446 expression was measured using specific primers (5’-GAGCAGTTGGAATCGGGTTTTAC-3’ and 5’-GTATGAATCGCAGTCCAGCCATA-3’) spanning 99 bp cDNA in the exon 1/exon 2 junction of CG18446. The primer pair efficiency was 99,1% (r2 larger than 0.99). CG18446 expression was normalized with Act5C expression levels (5’-GCGCCCTTACTCTTTCACCA-3’ and 5’-ATGTCACGGACGATTTCACG-3’).
Cold-stress resistance assays
Embryo collection was performed as mentioned above. Embryos were put into 50 ml fresh food vials. When embryos were 4–8 hour-old, they were kept at 1 C for 14 hours and then they were kept at room temperature (22–25 C). Simultaneously, control vials were always kept at room temperature (22–25 C) and never exposed to cold-stress. A total of 8–20 vials were analyzed per experiment. The same number of embryos per vial, 30 or 50, were used for all the replicates of a given experiment. Percentage viability was calculated based on the number of emerged flies to the total number of embryos placed in each vial.
Statistical significance was calculated performing two-way ANOVA using SPSS v21. We combined all the data into a full model: experimental condition (stress and nonstress), insertion genotype (presence/absence of the insertion) and interaction between these two factors. For those experiments in which more than one replicate was performed, the replicate effect was also taken into account. Because our dependent variable was a proportion, we used the arcsine transformation of the data before performing statistical analysis. We tested whether the data was normally distributed using Kolmogorov-Smirnov test. When the data was not normally distributed after the arcsine transformation, we applied the rank transformation. When the statistical test was significant, we estimated partial eta-squared values as a measure of the effect size (0.01 small effect, 0.06 medium effect, and 0.14 large effect).
Inferences of selection in the region flanking the roo solo-LTR insertions
We estimated the number of segregating sites (S), Tajima´s D, iHS, nSL and XP-EHH in the 2 kb region flanking the FBti0019985 insertion (chromosome 2R: 5758000–5760000) in 10 DGRP strains containing this insertion, in the 23 DGRP strains containing one of the roo insertions described in this work, and in the 15 strains that do not contain any insertion in the promoter region of CG18446. Note that the coordinates of FBti0019985 in the r5 of the D. melanogaster genome used by the DGRP project to generate the vcf files are 2R: 5,758,595–5,759,028. S and Tajima´s D are standard mesures of neutrality. iHS and nSL tests identify hard sweeps although they have some power to detect soft sweeps as well [69, 70]. H12 tests for positive selection on new variation and standing genetic variation within a population, that is, it searches both for soft and hard sweeps in a population [71]. Finally, XP-EHH is a statistical test of positive selection in one population that uses between populations comparisons to increase power in regions near fixation in the selected population [72].
We have used vcftools to calculate the number of segregating sites, and Tajima´s D using parameters –maf 1/(2n), where n is the sample size, and –remove-indels. We have obtained iHS, nSL, and XP-EHH using the selscan software with default parameters [73]. Finally, we have calculated H12 with ad hoc scripts. The four latter statistics require phased data. Thus, chromosome 2R of the 205 DGRP strains were phased together using ShapeIt [74].
To calculate the significance for the number of segregating sites, we resampled at random the same number of strains from the 205 DGRP strains available and calculated the distribution of segregating sites in the same 2 kb region. To calculate the significance of Tajima´s D, iHS, nSL and XP-EHH, we have used the empirical distributions of these statistics obtained from chromosome 2R.
Supporting Information
A frequency plot was built with all the TSD identified in this work, except the TSD of roo-19 and roo+7 that had four and two nucleotides instead of five, respectively, and with the 41 roo TSD motifs identified by Fiston-Lavier et al (2015) [34] (see Materials and Methods).
(TIFF)
The nine roo elements sequenced in this work are depicted in red.
(TIFF)
Single nucleotide polymorphisms are highlighted in red. (A) Alignment of the different roo insertions analyzed in this work. For RAL-502 and RAL-857 we could only sequence a partial region of the insertion and thus we only analyzed the Inr motif. (B) Alignment of the three regions with matrix association potential. (C) Alignment of the CG18446 promoter region in the different strains analyzed. Underlined sequences are from popdrowser [64]. For additional details see Fig 3 legend.
(PDF)
(PDF)
H12 was calculated on haplotypes of 40 segregating sites. All results are for the 10 strains with the FBti0019985 insertion combined with the 15 strains without any of the nine insertions, except for XP-EHH, which is calculated between the 10 strains with the FBti0019985 insertion and the 15 strains without any of the nine insertions. Horizontal dashed lines show significance levels while vertical dashed lines show the region of the insertion.
(PDF)
See legend of S5 Fig for details.
(PDF)
(A) Summary of the T-lex2 results in all populations. (B) Results for DGRP population. (C) Results for Sweden population. (D) Results for Zambia population. (E) Results for the Italian population for which PCRs were also performed. (F) Results for the DGRP strains for which PCR were also performed.
(XLSX)
(A) Transcription factor binding sites and promoter motifs, and (B) Matrix Associated regions, found in FBti0019985. (C) Transcription factor binding sites and promoter motifs found in the CG18446 promoter region.
(DOCX)
The total number of piRNA reads and of HP1a reads per nucleotide position, and the average number of piRNA reads and of HP1a reads per insertion are given.
(XLSX)
(DOCX)
(TXT)
Acknowledgments
We thank all members of the González lab for comments on the manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Ministerio de Economía y Competitividad (MINECO: http://www.idi.mineco.gob.es) BFU-2011-24397 and RYC-2010-07306 to JG, and BES-2012-052999 to AU, Ministerio de Economía y Competitividad and Fondo Europeo de Desarrollo Regional (MINECO/FEDER:http://www.idi.mineco.gob.es) BFU2014-57779-P, the European Commission (https://ec.europa.eu) FP7-PEOPLE-2011-CIG-293860, the Secretaria d’Universitats i Recerca del Departament d’Economia i Coneixement de la Generalitat de Catalunya (http://agaur.gencat.cat) 2014-SGR-201 to JG, and by the Agence Nationale de la Recherche (http://www.agence-nationale-recherche.fr) ANR-14-CE02-0003 to MARdC. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Juven-Gershon T, Kadonaga JT. Regulation of gene expression via the core promoter and the basal transcriptional machinery. Developmental biology. 2010;339(2):225–9. 10.1016/j.ydbio.2009.08.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Haberle V, Lenhard B. Promoter architectures and developmental gene regulation. Semin Cell Dev Biol. 2016. [DOI] [PubMed] [Google Scholar]
- 3.Hoskins RA, Landolin JM, Brown JB, Sandler JE, Takahashi H, Lassmann T, et al. Genome-wide analysis of promoter architecture in Drosophila melanogaster. Genome Res. 2011;21(2):182–92. 10.1101/gr.112466.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Carninci P, Sandelin A, Lenhard B, Katayama S, Shimokawa K, Ponjavic J, et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat Genet. 2006;38(6):626–35. [DOI] [PubMed] [Google Scholar]
- 5.Kawaji H, Frith MC, Katayama S, Sandelin A, Kai C, Kawai J, et al. Dynamic usage of transcription start sites within core promoters. Genome Biol. 2006;7(12):R118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Consortium Fantom, Suzuki H, Forrest AR, van Nimwegen E, Daub CO, Balwierz PJ, et al. The transcriptional network that controls growth arrest and differentiation in a human myeloid leukemia cell line. Nat Genet. 2009;41(5):553–62. 10.1038/ng.375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rach EA, Winter DR, Benjamin AM, Corcoran DL, Ni T, Zhu J, et al. Transcription initiation patterns indicate divergent strategies for gene regulation at the chromatin level. PLoS genetics. 2011;7(1):e1001274 10.1371/journal.pgen.1001274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Batut P, Dobin A, Plessy C, Carninci P, Gingeras TR. High-fidelity promoter profiling reveals widespread alternative promoter usage and transposon-driven developmental gene expression. Genome Res. 2013;23(1):169–80. 10.1101/gr.139618.112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.McClintock B. Intranuclear systems controlling gene action and mutation. Brookhaven Symp Biol. 1956;February(8):58–74. [PubMed] [Google Scholar]
- 10.Britten RJ, Davidson EH. Gene regulation for higher cells: a theory. Science. 1969;165(3891):349–57. [DOI] [PubMed] [Google Scholar]
- 11.Guio L, Barron MG, Gonzalez J. The transposable element Bari-Jheh mediates oxidative stress response in Drosophila. Molecular ecology. 2014;23(8):2020–30. 10.1111/mec.12711 [DOI] [PubMed] [Google Scholar]
- 12.Guio L, Gonzalez J. The dominance effect of the adaptive transposable element insertion Bari-Jheh depends on the genetic background. Genome biology and evolution. 2015;7(5):1260–6. 10.1093/gbe/evv071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chuong EB, Elde NC, Feschotte C. Regulatory evolution of innate immunity through co-option of endogenous retroviruses. Science. 2016;351(6277):1083–7. 10.1126/science.aad5497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lerman DN, Feder ME. Naturally occurring transposable elements disrupt hsp70 promoter function in Drosophila melanogaster. Molecular biology and evolution. 2005;22(3):776–83. [DOI] [PubMed] [Google Scholar]
- 15.Gonzalez J, Lenkov K, Lipatov M, Macpherson JM, Petrov DA. High rate of recent transposable element-induced adaptation in Drosophila melanogaster. PLoS biology. 2008;6(10):e251 10.1371/journal.pbio.0060251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bronner G, Taubert H, Jackle H. Mesoderm-specific B104 expression in the Drosophila embryo is mediated by internal cis-acting elements of the transposon. Chromosoma. 1995;103(10):669–75. [DOI] [PubMed] [Google Scholar]
- 17.Scherer G, Telford J, Baldari C, Pirrotta V. Isolation of cloned genes differentially expressed at early and late stages of Drosophila embryonic development. Developmental biology. 1981;86(2):438–47. [DOI] [PubMed] [Google Scholar]
- 18.Scherer G, Tschudi C, Perera J, Delius H, Pirrotta V. B104, a new dispersed repeated gene family in Drosophila melanogaster and its analogies with retroviruses. J Mol Biol. 1982;157(3):435–51. [DOI] [PubMed] [Google Scholar]
- 19.Meyerowitz EM, Hogness DS. Molecular organization of a Drosophila puff site that responds to ecdysone. Cell. 1982;28(1):165–76. [DOI] [PubMed] [Google Scholar]
- 20.FitzGerald PC, Sturgill D, Shyakhtenko A, Oliver B, Vinson C. Comparative genomics of Drosophila and human core promoters. Genome Biol. 2006;7(7):R53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Telonis-Scott M, Hallas R, McKechnie SW, Wee CW, Hoffmann AA. Selection for cold resistance alters gene transcript levels in Drosophila melanogaster. Journal of insect physiology. 2009;55(6):549–55. 10.1016/j.jinsphys.2009.01.010 [DOI] [PubMed] [Google Scholar]
- 22.Hoffmann AA, Scott M, Partridge L, Hallas R. Overwintering in Drosophila melanogaster: outdoor field cage experiments on clinal and laboratory selected populations help to elucidate traits under selection. J Evol Biol. 2003;16(4):614–23. [DOI] [PubMed] [Google Scholar]
- 23.Kellermann V, Loeschcke V, Hoffmann AA, Kristensen TN, Flojgaard C, David JR, et al. Phylogenetic constraints in key functional traits behind species' climate niches: patterns of desiccation and cold resistance across 95 Drosophila species. Evolution. 2012;66(11):3377–89. 10.1111/j.1558-5646.2012.01685.x [DOI] [PubMed] [Google Scholar]
- 24.Goto SG. Expression of Drosophila homologue of senescence marker protein-30 during cold acclimation. Journal of insect physiology. 2000;46(7):1111–20. [DOI] [PubMed] [Google Scholar]
- 25.Goto SG. A novel gene that is up-regulated during recovery from cold shock in Drosophila melanogaster. Gene. 2001;270(1–2):259–64. [DOI] [PubMed] [Google Scholar]
- 26.Greenberg AJ, Moran JR, Coyne JA, Wu CI. Ecological adaptation during incipient speciation revealed by precise gene replacement. Science. 2003;302(5651):1754–7. [DOI] [PubMed] [Google Scholar]
- 27.Qin W, Neal SJ, Robertson RM, Westwood JT, Walker VK. Cold hardening and transcriptional change in Drosophila melanogaster. Insect Mol Biol. 2005;14(6):607–13. [DOI] [PubMed] [Google Scholar]
- 28.Morgan TJ, Mackay TF. Quantitative trait loci for thermotolerance phenotypes in Drosophila melanogaster. Heredity (Edinb). 2006;96(3):232–42. [DOI] [PubMed] [Google Scholar]
- 29.Hoffmann AA, Blacket MJ, McKechnie SW, Rako L, Schiffer M, Rane RV, et al. A proline repeat polymorphism of the Frost gene of Drosophila melanogaster showing clinal variation but not associated with cold resistance. Insect Mol Biol. 2012;21(4):437–45. 10.1111/j.1365-2583.2012.01149.x [DOI] [PubMed] [Google Scholar]
- 30.Mamillapalli A, Pathak RU, Garapati HS, Mishra RK. Transposable element 'roo' attaches to nuclear matrix of the Drosophila melanogaster. Journal of insect science. 2013;13:111 10.1673/031.013.11101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huang W, Massouras A, Inoue Y, Peiffer J, Ramia M, Tarone AM, et al. Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 2014;24(7):1193–208. 10.1101/gr.171546.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mackay TF, Richards S, Stone EA, Barbadilla A, Ayroles JF, Zhu D, et al. The Drosophila melanogaster Genetic Reference Panel. Nature. 2012;482(7384):173–8. 10.1038/nature10811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ullastres A, Petit N, Gonzalez J. Exploring the Phenotypic Space and the Evolutionary History of a Natural Mutation in Drosophila melanogaster. Molecular biology and evolution. 2015;32(7):1800–14. 10.1093/molbev/msv061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fiston-Lavier AS, Barron MG, Petrov DA, Gonzalez J. T-lex2: genotyping, frequency estimation and re-annotation of transposable elements using single or pooled next-generation sequencing data. Nucleic Acids Res. 2015;43(4):e22 10.1093/nar/gku1250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Attrill H, Falls K, Goodman JL, Millburn GH, Antonazzo G, Rey AJ, et al. FlyBase: establishing a Gene Group resource for Drosophila melanogaster. Nucleic Acids Res. 2016;44(D1):D786–92. 10.1093/nar/gkv1046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bernstein M, Lersh RA, Subrahmanyan L, Cline TW. Transposon Insertions Causing Constitutive Sex-Lethal Activity in Drosophila melanogaster Affect Sxl Sex-Specific Transcript Splicing. Genetics. 1995;139:631–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Linheiro RS, Bergman CM. Whole genome resequencing reveals natural target site preferences of transposable elements in Drosophila melanogaster. PLoS One. 2012;7(2):e30008 10.1371/journal.pone.0030008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Spradling AC, Stern DM, Kiss I, Roote J, Laverty T, Rubin GM. Gene disruptions using P transposable elements: an integral component of the Drosophila genome project. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(24):10824–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liao GC, Rehm EJ, Rubin GM. Insertion site preferences of the P transposable element in Drosophila melanogaster. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(7):3347–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Reed DE, Huang XM, Wohlschlegel JA, Levine MS, Senger K. DEAF-1 regulates immunity gene expression in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(24):8351–6. 10.1073/pnas.0802921105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dantoft W, Davis MM, Lindvall JM, Tang X, Uvell H, Junell A, et al. The Oct1 homolog Nubbin is a repressor of NF-kappaB-dependent immune gene expression that increases the tolerance to gut microbiota. BMC biology. 2013;11:99 10.1186/1741-7007-11-99 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sentmanat MF, Elgin SC. Ectopic assembly of heterochromatin in Drosophila melanogaster triggered by transposable elements. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(35):14104–9. 10.1073/pnas.1207036109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lee YC. The Role of piRNA-Mediated Epigenetic Silencing in the Population Dynamics of Transposable Elements in Drosophila melanogaster. PLoS genetics. 2015;11(6):e1005269 10.1371/journal.pgen.1005269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li C, Vagin VV, Lee S, Xu J, Ma S, Xi H, et al. Collapse of germline piRNAs in the absence of Argonaute3 reveals somatic piRNAs in flies. Cell. 2009;137(3):509–21. 10.1016/j.cell.2009.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Satyaki PR, Cuykendall TN, Wei KH, Brideau NJ, Kwak H, Aruna S, et al. The Hmr and Lhr hybrid incompatibility genes suppress a broad range of heterochromatic repeats. PLoS genetics. 2014;10(3):e1004240 10.1371/journal.pgen.1004240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shpiz S, Ryazansky S, Olovnikov I, Abramov Y, Kalmykova A. Euchromatic transposon insertions trigger production of novel Pi- and endo-siRNAs at the target sites in the drosophila germline. PLoS genetics. 2014;10(2):e1004138 10.1371/journal.pgen.1004138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kharchenko PV, Alekseyenko AA, Schwartz YB, Minoda A, Riddle NC, Ernst J, et al. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. Nature. 2011;471(7339):480–5. 10.1038/nature09725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Eissenberg JC, Elgin SC. HP1a: a structural chromosomal protein regulating transcription. Trends Genet. 2014;30(3):103–10. 10.1016/j.tig.2014.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.dos Santos G, Schroeder AJ, Goodman JL, Strelets VB, Crosby MA, Thurmond J, et al. FlyBase: introduction of the Drosophila melanogaster Release 6 reference genome assembly and large-scale migration of genome annotations. Nucleic Acids Res. 2015;43(Database issue):D690–7. 10.1093/nar/gku1099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Walser JC, Chen B, Feder ME. Heat-shock promoters: targets for evolution by P transposable elements in Drosophila. PLoS genetics. 2006;2(10):e165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lerman DN, Michalak P, Helin AB, Bettencourt BR, Feder ME. Modification of Heat-Shock Gene Expression in Drosophila melanogaster Populations via Transposable Elements. Molecular biology and evolution. 2003;20(1):135–44. [DOI] [PubMed] [Google Scholar]
- 52.Shilova VY, Garbuz DG, Myasyankina EN, Chen B, Evgen'ev MB, Feder ME, et al. Remarkable site specificity of local transposition into the Hsp70 promoter of Drosophila melanogaster. Genetics. 2006;173(2):809–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gruber JD, Genissel A, Macdonald SJ, Long AD. How repeatable are associations between polymorphisms in achaete-scute and bristle number variation in Drosophila? Genetics. 2007;175(4):1987–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kenny MC, Wilton A, Ballard JWO. Seasonal trade-off between starvation resistance and cold resistance in temperate wild-caught Drosophila simulans. Australian Journal of Entomology. 2008;47(1):20–3. [Google Scholar]
- 55.Hoffmann AA, Hallas R, Anderson AR, Telonis-Scott M. Evidence for a robust sex-specific trade-off between cold resistance and starvation resistance in Drosophila melanogaster. J Evol Biol. 2005;18(4):804–10. [DOI] [PubMed] [Google Scholar]
- 56.Watson MJO, Hoffmann AA. Acclimation, cross-generation effects, and the response to selection for increased cold resistance in Drosophila. Evolution. 1996;50(3):1182–92. [DOI] [PubMed] [Google Scholar]
- 57.Marshall KE, Sinclair BJ. Repeated stress exposure results in a survival-reproduction trade-off in Drosophila melanogaster. Proc Biol Sci. 2010;277(1683):963–9. 10.1098/rspb.2009.1807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kong EC, Allouche L, Chapot PA, Vranizan K, Moore MS, Heberlein U, et al. Ethanol-regulated genes that contribute to ethanol sensitivity and rapid tolerance in Drosophila. Alcohol Clin Exp Res. 2010;34(2):302–16. 10.1111/j.1530-0277.2009.01093.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ayres JS, Freitag N, Schneider DS. Identification of Drosophila Mutants Altering Defense of and Endurance to Listeria monocytogenes Infection. Genetics. 2008;178:1807–15. 10.1534/genetics.107.083782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lack JB, Cardeno CM, Crepeau MW, Taylor W, Corbett-Detig RB, Stevens KA, et al. The Drosophila genome nexus: a population genomic resource of 623 Drosophila melanogaster genomes, including 197 from a single ancestral range population. Genetics. 2015;199(4):1229–41. 10.1534/genetics.115.174664 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular biology and evolution. 2013;30(4):772–80. 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–3. 10.1093/bioinformatics/btu033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Huerta-Cepas J, Serra F, Bork P. ETE 3: Reconstruction, Analysis, and Visualization of Phylogenomic Data. Molecular biology and evolution. 2016;33(6):1635–8. 10.1093/molbev/msw046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ramia M, Librado P, Casillas S, Rozas J, Barbadilla A. PopDrowser: the Population Drosophila Browser. Bioinformatics. 2012;28(4):595–6. 10.1093/bioinformatics/btr691 [DOI] [PubMed] [Google Scholar]
- 65.Li H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. bioRxiv 13033997v2. 2013.
- 66.Barnett DW, Garrison EK, Quinlan AR, Stromberg MP, Marth GT. BamTools: a C++ API and toolkit for analyzing and managing BAM files. Bioinformatics. 2011;27(12):1691–2. 10.1093/bioinformatics/btr174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.RStudioTeam. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA: URL http://www.rstudio.com/. 2015. [Google Scholar]
- 68.Schou MF. Fast egg collection method greatly improves randomness of egg sampling in Drosophila melanogaster. Fly (Austin). 2013;7(1):44–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Voight BF, Kudaravalli S, Wen X, Pritchard JK. A map of recent positive selection in the human genome. PLoS biology. 2006;4(3):e72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ferrer-Admetlla A, Liang M, Korneliussen T, Nielsen R. On detecting incomplete soft or hard selective sweeps using haplotype structure. Molecular biology and evolution. 2014;31(5):1275–91. 10.1093/molbev/msu077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Garud NR, Messer PW, Buzbas EO, Petrov DA. Recent selective sweeps in North American Drosophila melanogaster show signatures of soft sweeps. PLoS genetics. 2015;11(2):e1005004 10.1371/journal.pgen.1005004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sabeti PC, Varilly P, Fry B, Lohmueller J, Hostetter E, Cotsapas C, et al. Genome-wide detection and characterization of positive selection in human populations. Nature. 2007;449(7164):913–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Szpiech ZA, Hernandez RD. selscan: an efficient multithreaded program to perform EHH-based scans for positive selection. Molecular biology and evolution. 2014;31(10):2824–7. 10.1093/molbev/msu211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Delaneau O, Howie B, Cox AJ, Zagury JF, Marchini J. Haplotype estimation using sequencing reads. Am J Hum Genet. 2013;93(4):687–96. 10.1016/j.ajhg.2013.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A frequency plot was built with all the TSD identified in this work, except the TSD of roo-19 and roo+7 that had four and two nucleotides instead of five, respectively, and with the 41 roo TSD motifs identified by Fiston-Lavier et al (2015) [34] (see Materials and Methods).
(TIFF)
The nine roo elements sequenced in this work are depicted in red.
(TIFF)
Single nucleotide polymorphisms are highlighted in red. (A) Alignment of the different roo insertions analyzed in this work. For RAL-502 and RAL-857 we could only sequence a partial region of the insertion and thus we only analyzed the Inr motif. (B) Alignment of the three regions with matrix association potential. (C) Alignment of the CG18446 promoter region in the different strains analyzed. Underlined sequences are from popdrowser [64]. For additional details see Fig 3 legend.
(PDF)
(PDF)
H12 was calculated on haplotypes of 40 segregating sites. All results are for the 10 strains with the FBti0019985 insertion combined with the 15 strains without any of the nine insertions, except for XP-EHH, which is calculated between the 10 strains with the FBti0019985 insertion and the 15 strains without any of the nine insertions. Horizontal dashed lines show significance levels while vertical dashed lines show the region of the insertion.
(PDF)
See legend of S5 Fig for details.
(PDF)
(A) Summary of the T-lex2 results in all populations. (B) Results for DGRP population. (C) Results for Sweden population. (D) Results for Zambia population. (E) Results for the Italian population for which PCRs were also performed. (F) Results for the DGRP strains for which PCR were also performed.
(XLSX)
(A) Transcription factor binding sites and promoter motifs, and (B) Matrix Associated regions, found in FBti0019985. (C) Transcription factor binding sites and promoter motifs found in the CG18446 promoter region.
(DOCX)
The total number of piRNA reads and of HP1a reads per nucleotide position, and the average number of piRNA reads and of HP1a reads per insertion are given.
(XLSX)
(DOCX)
(TXT)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.