

Abstract
Using infochemicals to develop a push–pull strategy in pest control is a potential way to promote sustainable crop production. Infochemicals from plant essential oils were mixed with paraffin oil for slow release in field experiments on wheat to control the population density of cereal aphids and to enhance their natural enemies. (Z)-3-Hexenol (Z3H) attracted Metopolophum dirhodum and Sitobion avenae, the predominant species on wheat in Belgium, and may be a useful infochemical for aphid control by attracting aphids away from field plots. Release of (E)-β-farnesene (EBF) or a garlic extract (GE) led to a significant decrease in the abundance of wheat aphids. The main natural enemies of cereal aphids found were lacewings (47.8%), hoverflies (39.4%), and ladybirds (12.8%). Ladybird abundance varied little before the end of the wheat-growing season. Our results suggest that these chemicals can form the basis of a “push–pull” strategy for aphid biological control, with GE and EBF acting as a pest- and beneficial-pulling stimulus and Z3H for aphid pulling.
Among aphid species, the grain aphid [Sitobion avenae (Fabricius)], bird cherry-oat aphid [Rhopalosiphum padi (L.)], and rose-grain aphid [Metopolophium dirhodum (Walker)] are considered the major pests that infest cereal crops as a result of feeding on phloem and transmitting viruses1,2, particularly on winter wheat (Triticum aestivum L. [Poaceae]) in Europe3. Aphid populations often fluctuate greatly from year to year4 and are affected by a range of biotic and abiotic factors5.
Because of the urgent need for sustainable agricultural methods and reduced reliance on pesticide use, more integrated pest management studies are focusing on the ecological effect of volatiles released by plants on herbivores and their natural enemies6,7,8,9,10,11,12,13,14. Several studies on volatiles under natural conditions have demonstrated their applicability for enhancing natural enemy abundance on strawberry (Fragaria ×ananassa)7, cotton (Gossypium spp.)12, hops (Humulus lupulus)8 and grapes (Vitis vinifera)9 and for reducing pest populations in wheat (Triticum aestivum)6,15 and barley (Hordeum vulgare)16.
While attracting natural enemies of these herbivores17, volatiles emanating from herbivore-infested plants may also stimulate plant defense against herbivores and serve as recognition cues between two or more individuals18. Dicke et al. (1987, 1990) presented the first convincing evidence that the active release of volatiles by herbivore-infested plants attracts natural enemies of the attackers19,20. Aphid behaviour is also affected by a density mechanism that is mediated by volatile compounds released at the feeding site when their density exceeds a certain threshold16. A further study revealed that these volatiles could increase the sensitivity of aphids to disturbance and promote mobility of nonsettled individuals21.
Because they are a natural emission from plants, essential oils do not pose the toxicity problems of pesticides to animals and the environment15,22. Plant semiochemicals should be considered as potential reliable infochemicals in relation to to repelling pests and attracting natural enemies of these pests. Their long-distance effects and easy production and manipulation make these molecules very good prospects for use with crops by spraying or mixing with a slow-releasing carrier to repel insect feeding or ovipositing from host plants and/or to guide them to nonhosts23.
Japanese termite (Reticulitermes speratus)24, sciarid fly [Lycoriella ingénue (Dufour)]22 and pine wood nematode (Bursaphelenchus xylophilus)25 were repelled by a garlic (Allium sativum) extract (GE), providing direct evidence that strongly aromatic crops such as garlic, can act as an olfactory camouflage by masking normal host-locating or feeding cues of insects (Perrin and Phillips, 1978). (E)-β-Farnesene (EBF), an important volatile sesquiterpene that occurs widely in both plant and animal taxa, such as aphids26 and peppermint (Mentha ×piperita L.)27, is an effective kairomone for ladybirds28,29,30, lacewings31 and hoverflies32. It is proven to be the main or only component of the aphid alarm pheromones for many pest aphids33,34,35,36,37.
Herbivore-induced volatiles (HIVs), for example, (Z)-3-hexenol (Z3H), can directly affect the physiology and behavior of herbivores38. Z3H has been demonstrated to attract Agrilus planipennis in the laboratory and field39,40 and the fruit moth Cydia molesta41. Although it has been difficult to determine whether Z3H is an attractant or a repellent, accumulating evidence suggests that Z3H is an important plant-derived infochemical that can modulate the behavior of herbivorous insects and that the release of Z3H induces defensive responses in the plants against insect pests38.
Extensive evidence implies that nearly all herbivorous insects and their natural enemies can perceive and positively respond to plant volatiles. In this investigation, the essential oils of plant volatiles (EBF, GE and Z3H) were released in a wheat field to assess their potential for managing aphid populations by reducing aphid abundance and promoting their natural enemies.
Materials and Methods
Experimental design of field studies
In the experimental fields of Gembloux Agro-Bio Tech, University de Liege, Namur Province of Belgium (50 °33″ N, 4 °42″ E) in 2011, traps were set out as shown in Fig. 1. The trial consisted of four treatments in the wheat field: (1) only paraffin oil (PO) as the control, (2) (E)-β-farnesene release (EBF), (3) garlic extract release (GE), (4) (Z)-3-hexenol release (Z3H). Those extracts were provided by Prof. Frédéric Francis (Gembloux Agro-Bio-Tech., Universite de Liege). Single yellow trap sticks with the releasers were placed 20 m apart in a latin square design with 3 replicates per treatment (12 releasers and 12 traps total). Wheat (cv. Tybalt) was planted in 20-cm-apart rows at 350 seeds/m2 on 18 February 2011. No insecticides or herbicides were used in the whole experimental area.
Figure 1. Layout of traps within experimental field.
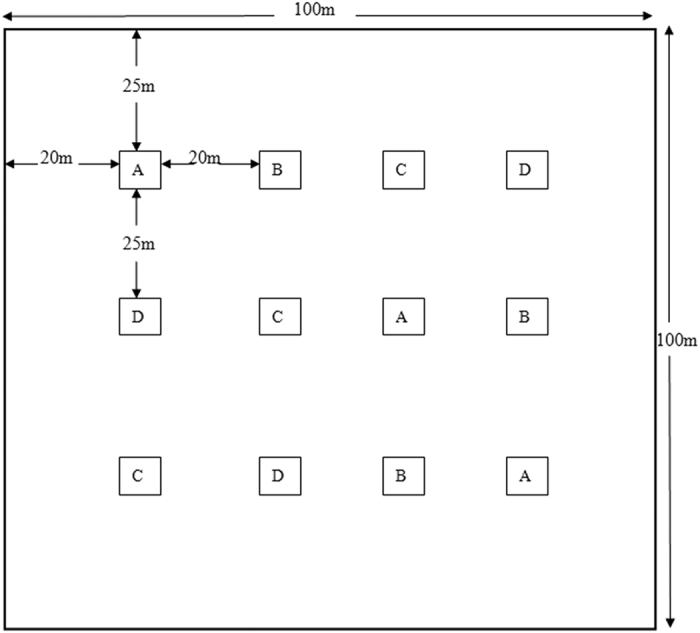
(A) Paraffin oil, (B) (E)-β-farnesene, (C) garlic extract, (D) (Z)-3-hexenol.
Assessment of insect abundance and diversity
Yellow traps (26 cm diameter 10 cm depth) that are frequently used to monitor insects in fields42 were attached to crabsticks and placed 10 cm above the surface of the wheat plants. Each trap was filled with water and a few drops detergent. Every 7 days, 100 μL of (E)-β-farnesene, garlic extract or (Z)-3-hexenol solution formulated in paraffin oil (for slow release of the infochemcial) were deposited on a 1-cm-diameter rubber septum that was placed on the top of the trap stick; 76 μg of EBF is released from the formulation over 7 days at 20 °C, 65% relative humidity and air flow of 0.5 litre/min (Dr. S. Heuskin, unpublished data). A similar release rate was applied to the other tested semiochemicals. The slow releasers were first placed in the wheat field at the jointing stage on 4 May.
Traps were emptied and reset at 7-day intervals between 11 May to 29 June. Trap contents were decanted through a 1-mm-mesh sieve and transferred to 70% ethanol in 50-mL plastic vials. In the laboratory, aphids and their natural enemies were sorted and identified to species, and the abundance of each species was recorded.
Aphid abundance in the traps was compared every 7 days to the aphid density determined by visual observation on 20 randomly selected wheat tillers.
Statistical analyses
For all parametric tests, a data sqrt (n + 1) transformation was applied to stabilize the variance. Population densities of insects were compared among the infochemical releaser tests using a one-way analysis of variance (ANOVA)43, followed by Tukey’s honestly significant difference (HSD) test.
Results
Abundance and diversity of aphids after exposure to infochemicals
M. dirhodum and S. avenae were the predominant species on wheat, and Z3H was the most attractive to these aphids. EBF and GE repelled aphids significantly within wheat fields. Trapping numbers and visual counts of aphid were consistent. M. dirhodum was far more abundant than S. avenae in observations and traps (Table 1 and Fig. 2). In addition, several wheat nontarget aphid species were recorded in traps: Cavariella aegopodii (Scopoli), Aphis fabae (Scopoli), Macrosiphum euphorbiae (Thomas), Myzus persicae (Sultzer), Rhopalosiphum maidis (Fitch), Cavariella ihedbaldi, Nasonovia ribisnigri (Mosley), Phyllaphis fagi (Linnaeus), Chaitophorus spp. and Capitophorus spp.
Table 1. Total number of aphids and their natural enemies found in yellow traps in different crop systems during 2011 growing season.
| Species | Treatments |
||||
|---|---|---|---|---|---|
| Paraffin oil | (E)-β-Farnesene | Garlic extract | (Z)-3-Hexenol | % of Totala | |
| Aphids | |||||
| Metopolophum dirhodum (Walker) | 896 | 585 | 582 | 1122 | 89.5 |
| Sitobion avenae (Fabricius) | 138 | 35 | 54 | 148 | 10.5 |
| Diversity and abundance of aphid species % | 29.0 | 17.4 | 17.9 | 35.7 | |
| Ladybirds | 12.8%b | ||||
| Harmonia axyridis Pallas | 18 | 21 | 22 | 28 | 66.8 |
| Coccinella septempunctata L. | 9 | 9 | 8 | 3 | 21.8 |
| Propylea 14-punctata L. | 3 | 3 | 1 | 0 | 5.3 |
| Harmonia 4-punctata | 2 | 1 | 0 | 0 | 2.3 |
| Calvia 14-guttata | 2 | 0 | 0 | 0 | 1.5 |
| Hippodamia variegata (Goeze) | 1 | 1 | 0 | 1 | 2.3 |
| Hoverflies | 39.4%b | ||||
| Episyrphus balteatus De Geer | 69 | 108 | 85 | 76 | 82.6 |
| Scaeva pyrastri L. | 2 | 0 | 0 | 7 | 2.2 |
| Sphaerophoria scripta L. | 12 | 16 | 9 | 8 | 11.0 |
| Melanostoma scalare Fabr. | 0 | 3 | 0 | 1 | 1.0 |
| Metasyrphus corollae Fabr. | 5 | 1 | 2 | 5 | 3.2 |
| Lacewings | 47.8%b | ||||
| Chrysoperla carnea Stephens | 95 | 128 | 152 | 121 | 100.0 |
| Total numbers of aphidophagous species | 218 | 291 | 279 | 250 | |
| Percentage of total number of aphidophagous species | 21.0 | 28.0 | 26.9 | 24.1 | |
aRelative abundance of each species by family.
bRelative occurrence of each family in aphidophagous guild.
Figure 2. Total number of aphids (mean ± SEM) found in field observations according to infochemical released.
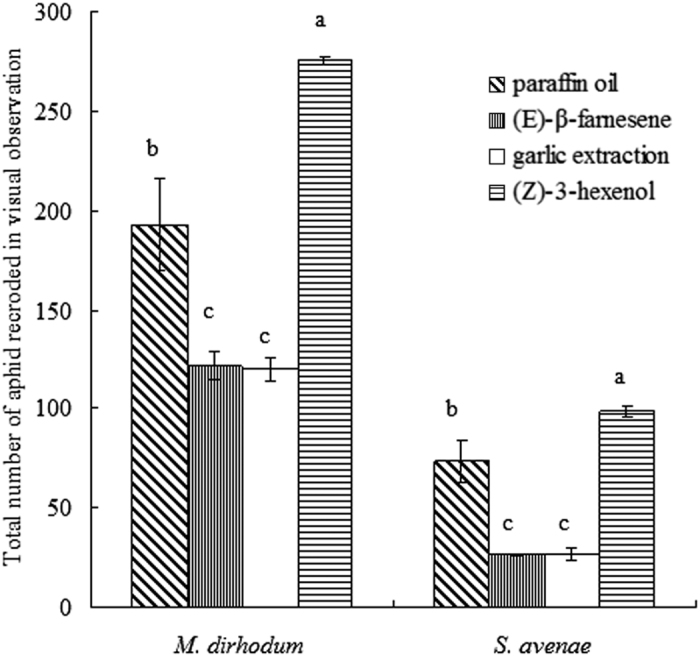
Different letters indicate a statistically significant difference between the individual treatments at P < 0.05.
According to visual observations and trapping, the population dynamics of M. dirhodum and S. avenae in each treatment followed the same trend on growing wheat, with increasing population densities that peaked on 15 June and 22 June, respectively (Fig. 3). Based on visual observations in the field, Z3H attracted mainly M. dirhodum for both the highest peak value and total during the whole observation period, whereas EBF and GE repelled aphids (Peak: F3,8 = 18.95, P < 0.01; Total: F3,8 = 34.45, P < 0.01). Similarly, significantly fewer S. avenae were found with EBF and GE releasers compared with the control PO (Peak: F3,8 = 89.30, P < 0.01; Total: F3,8 = 45.55, P < 0.01).
Figure 3. Number of aphids (mean ± SEM) trapped or counted in observations in wheat fields during different treatments in 2011.
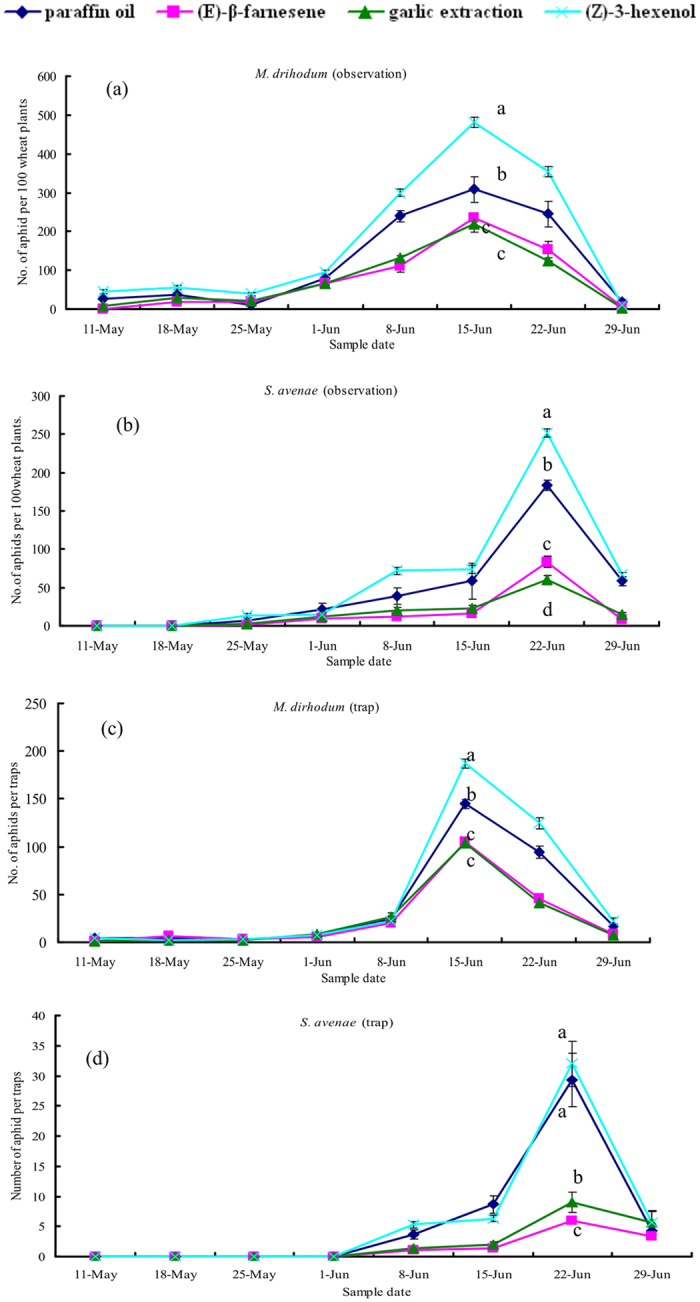
Different letters indicate a statistically significant difference between individual treatments at P < 0.05.
Consistent with the results of visual observations, both peak and total abundance of M. dirhodum in traps was higher with Z3H and lower with EBF and GE releasers during the experiment (Peak: F3,8 = 56.30, P < 0.01; Total: F3,8 = 86.27, P < 0.01). The highest abundance of S. avenae was found in traps with Z3H. EBF and GE releasers were also found to repel S. avenae as evidenced by both the peak period and the total during the experiment (Peak: F3,8 = 56.30, P < 0.01; Total: F3,8 = 86.27, P < 0.01). The consistency of the data obtained from visual observations and trapping confirmed the infochemical results for M. dirhodum and S. avenae.
Abundance and diversity of natural aphid enemies in response to the infochemical released
The main natural enemies of cereal aphids found in the trials in order of abundance were lacewings (47.8%), hoverflies (39.4%) and ladybirds (12.8%). Of the predatory species, E. balteatus, C. carnea and H. axyridis were the predominant species on wheat. On the basis of total number of aphidophagous species attracted, EBF, GE and Z3H attracted more than the control PO did (Table 1). Not all the collected hoverflies were aphidophagous species (Eristalis pertinax, Helophilus trivitatus, Cheilosia spp., Eristalis tenax, Eristalis arbustorum). The aphid predators and their diversity are presented in Table 1.
The hoverfly population density had reached its peak by 29 June (Fig. 4(a)). Before this peak, hoverfly density did not differ among the tested infochemicals. After the peak, hoverfly density in response to EBF releases was significantly higher than with Z3H (F3,8 = 4.46, P < 0.05). No significant difference in total hoverfly abundance among treatments was detected (F3,8 = 1.64, P = 0.26).
Figure 4. Number of natural enemies (mean ± SEM) trapped during the different treatments in wheat fields in 2011.

Different letters indicate a statistically significant difference between individual treatments at P < 0.05.
The number of lacewings peaked in all treatments on 15 June coincident with the peak of M. dirhodum (Fig. 4(b)). The population density of lacewings in each treatment was low before 8 June. No significant difference in total lacewing abundance among treatments was detected (F3,8 = 1.25, P = 0.36).
Finally, ladybird population dynamics did not vary significantly among treatments before 22 June (Fig. 4(c)). Moreover, the ladybird population peaked in all treatments at the end of the wheat season when the aphid population declined rapidly in the field. No significant difference in the abundance of ladybirds among treatments was detected either at the peak period or for the total numbers during the experiment (Peak: F3,8 = 1.92, P = 0.21; Total: F3,8 = 0.52, P = 0.68).
Discussion
The densities of cereal aphids and their natural enemies in wheat were significantly influenced by the test infochemical releasers, mainly with EBF and GE, supporting the view that these volatiles play a significant role in the behavioural ecology of aphids and demonstrating the potential use of the volatiles in pest control. As reviewed by Kunert et al.44, several factors could contribute to the low abundance of M. dirhodum and S. avenae in the EBF-release plots. First, EBF emission may directly prevent aphid settling because wild potato (Solanum berthaultii) repels the green peach aphid (Myzus persicae) by emitting EBF45. EBF might also reduce aphid growth rate by disrupting feeding46 or by inducing wing formation and reducing aphid population size47,48. Since winged offspring leave their host plant before starting reproduction, plants that produce EBF could reduce aphid colonization49,50. Under natural conditions, plants emit infochemical as signals in response to attack by insect herbivores that recruit natural enemies of the herbivores51; thus, EBF release in plots might primarily improve the efficiency of the natural enemies in locating their prey. This hypothesis is supported by the results of our study that population densities of hoverflies were higher when EBF release was at its peak. Nevertheless, there were some exceptions to the influence of EBF on lacewings and ladybirds in our investigation. The amount of infochemical in releasers may determine the probability of predator response. Shiojiri et al.52 showed that seedlings of a cabbage variety attracted more parasitoids (Cotesia glomerata) when there were more herbivores on the plant. Further study is needed to demonstrate and clarify the mechanism for this phenomenon.
Aphids perceive the host plant and avoid nonhosts by sensing volatile cues53. Garlic plants are not hosts to cereal aphids, so a garlic extract is likely to be unsuitable for aphids. Indeed, population densities of M. dirhodum and S. avenae were significantly lower in GE-release plots than in the PO control plots. Also worth mentioning is that GE significantly attracted more lacewings than did the PO plots. Moreover, GE did not negatively influence field populations of hoverflies or ladybirds. As far as we know, this study is the first to show that GE or garlicin helps plants recruit natural enemies of aphids.
On the basis of available knowledge, wound-induced, ubiquitous (Z)-3-hexenol, a C6-alcohol synthesized in the lipoxygenase/HPL pathway, is the most important infochemical influencing herbivore repellence and attraction in tritrophic interactions38. Quiroz and Niemeyer54 found that volatiles from wheat and oat seedlings attracted winged and wingless Rhopalosiphum padi. These volatiles were identified by GC-MS, and olfactometer tests performed with each compound showed that aphids were attracted by (E)-2-hexenyl acetate, (Z)-3-hexenol, (Z)-2-hexenol and so on. Our result that the Z3H release attracted the highest population densities of M. dirhodum and S. avenae in (Fig. 2) agrees with their report on the cereal aphid R. padi54.
The push–pull strategy is a behavioral manipulation method that uses repellent/deterrent (push) and attractive/stimulant (push) stimuli to direct the movement of pest or beneficial insects for pest management55. The volatiles tested in the present study were either a repellent or attractant stimuli to aphids and either an attractant or neutral to natural enemies (beneficials), depending on the infochemical. Z3H acted as a pull stimulus to the aphids, but was neutral to beneficials; GE and EBF acted as a push stimulus for the aphids and as a pull for beneficials (EBF to hoverfly, GE to lacewings). The three infochemicals could be used to promote a push–pull strategy and have great potential for integrated pest management of wheat aphids. Recent studies have provided evidence for the potential use of synthetic volatiles as aids to enhance biological control measures in crop ecosystems13,56,57. Targeting the right volatiles for enhanced emission could lead to ecologically and economically sound ways of combating important pests. However, a remaining question surrounding the use of these materials in integrated pest management is the ecological consequences of these synthetic volatiles on predators and parasitoids in the absence of their prey. Therefore, more detailed work on ecological consequences and application rate, dose and duration under field conditions must be done before those volatiles can be developed as a semiochemical tool to replace broad-spectrum insecticides. Manipulating the behavior of natural enemies to improve biological control holds great potential for improving push–pull strategies so that they can be more widely deployed for sustainable agricultural systems in the future.
Additional Information
How to cite this article: Zhou, H. et al. Use of slow-release plant infochemicals to control aphids: a first investigation in a Belgian wheat field. Sci. Rep. 6, 31552; doi: 10.1038/srep31552 (2016).
Acknowledgments
We sincerely thank Almouner A. Yattara and Poligui René-Noël (Department of Functional and Evolutionary Entomology, University of Liege, Gembloux Agro-Bio Tech) for their assistance in identifying aphids and their natural enemy species. This research was supported by the International Cooperation Project between Belgium and China (CUD\PICShandong, 2014DFG32270), Anhui Provincial Natural Science Foundation (1608085QC61), Anhui Postdoctoral Fund Project for Scientific Research and the National Natural Science Foundation of China (31371946).
Footnotes
Author Contributions J.C. and F.F. designed the experiment. H.Z. did the experiments. H.Z., Y.L. and L.C. wrote the manuscript. All authors reviewed the manuscript.
References
- Liu Y., Wang W. L., Guo G. X. & Ji X. L. Volatile emission in wheat and parasitism by Aphidius avenae after exogenous application of salivary enzymes of Sitobion avenae. Entomol Exp Appl 130, 215–221, (2009). [Google Scholar]
- Van Emden H. F. & Harrington R. Aphids as crop pests. (CAB International, 2007). [Google Scholar]
- Poehling H. M., Freier B. & Klüken A. M. In Aphids as crop pests (eds Van Emden H. F. & Harrington R.) 597–611 (CAB International, 2007). [Google Scholar]
- Kindlmann P. & Dixon A. F. G. In Aphid Biodiversity under Environmental Change (eds Kindlmann Pavel, Dixon A. F. G. & Michaud J. P.) 1–20 (Springer Netherlands, 2010). [Google Scholar]
- Leslie T. W., Van Der Werf W., Bianchi F. J. J. A. & Honěk A. Population dynamics of cereal aphids: influence of a shared predator and weather. Agric For Entomol 11, 73–82 (2009). [Google Scholar]
- Wang G. et al. Combining intercropping with semiochemical releases: optimization of alternative control of Sitobion avenae in wheat crops in China. Entomol Exp Appl 140, 189–195 (2011). [Google Scholar]
- Lee J. C. Effect of Methyl Salicylate-Based Lures on Beneficial and Pest Arthropods in Strawberry. Environ Entomol 39, 653–660 (2010). [DOI] [PubMed] [Google Scholar]
- James D. G. Further Field Evaluation Of Synthetic Herbivore-Induced Plan Volatiles As Attractants For Beneficial Insects. J Chem Ecol 31, 481–495 (2005). [DOI] [PubMed] [Google Scholar]
- James D. & Grasswitz T. Synthetic Herbivore-induced Plant Volatiles Increase Field Captures of Parasitic Wasps. BioControl 50, 871–880 (2005). [Google Scholar]
- De Boer J. G. & Dicke M. The Role of Methyl Salicylate in Prey Searching Behavior of the Predatory Mite Phytoseiulus persimilis. J Chem Ecol 30, 255–271 (2004). [DOI] [PubMed] [Google Scholar]
- Snoeren T. et al. The Herbivore-Induced Plant Volatile Methyl Salicylate Negatively Affects Attraction of the Parasitoid Diadegma semiclausum. J Chem Ecol 36, 479–489 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu H., Zhang Y., Wu K., Gao X. W. & Guo Y. Y. Field-Testing of Synthetic Herbivore-Induced Plant Volatiles as Attractants for Beneficial Insects. Environ Entomol 37, 1410–1415 (2008). [DOI] [PubMed] [Google Scholar]
- James D. G. & Price T. S. Field-Testing of Methyl Salicylate for Recruitment and Retention of Beneficial Insects in Grapes and Hops. J Chem Ecol 30, 1613–1628 (2004). [DOI] [PubMed] [Google Scholar]
- Plepys D., Ibarra F. & Löfstedt C. Volatiles from flowers of Platanthera bifolia (Orchidaceae) attractive to the silver Y moth, Autographa gamma (Lepidoptera: Noctuidae). Oikos 99, 69–74 (2002). [Google Scholar]
- Prinsloo G. et al. Test of semiochemicals and a resistant wheat variety for Russian wheat aphid management in South Africa. J Appl Entomol 131, 637–644 (2007). [Google Scholar]
- Ninkovic V., Ahmed E., Glinwood R. & Pettersson J. Effects of two types of semiochemical on population development of the bird cherry oat aphid Rhopalosiphum padi in a barley crop. Agric For Entomol 5, 27–34 (2003). [Google Scholar]
- Turlings T. C. J., Tumlinson J. H. & Lewis W. J. Exploitation of Herbivore-Induced Plant Odors by Host-Seeking Parasitic Wasps. Science 250, 1251–1253 (1990). [DOI] [PubMed] [Google Scholar]
- Howard R. W. & Blomquist G. J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu Rev Entomol 50, 371–393 (2005). [DOI] [PubMed] [Google Scholar]
- Dicke M. & Sabelis M. W. How Plants Obtain Predatory Mites as Bodyguards. Netherlands Journal of Zoology 38, 148–165 (1987). [Google Scholar]
- Dicke M., Sabelis M. W., Takabayashi J., Bruin J. & Posthumus M. A. Plant strategies of manipulating predatorprey interactions through allelochemicals: Prospects for application in pest control. J Chem Ecol 16, 3091–3118 (1990). [DOI] [PubMed] [Google Scholar]
- Pettersson J., Quiroz A., Stephansson D. & Niemeyer H. Odour communication of Rhopalosiphum padi on grasses. ENTOMOL EXP APPL 76, 325–328 (1995). [Google Scholar]
- Park I. I. K. et al. Fumigant activity of plant essential oils and components from horseradish (Armoracia rusticana), anise (Pimpinella anisum) and garlic (Allium sativum) oils against Lycoriella ingenua (Diptera: Sciaridae). Pest Manag Sci 62, 723–728 (2006). [DOI] [PubMed] [Google Scholar]
- Pickett J. A., Wadhams L. J. & Woodcock C. M. New approaches to the development of semiochemicals for insect control. 333–345 (Hague, 1991). [Google Scholar]
- Park I.-K. & Shin S.-C. Fumigant Activity of Plant Essential Oils and Components from Garlic (Allium sativum) and Clove Bud (Eugenia caryophyllata) Oils against the Japanese Termite (Reticulitermes speratus Kolbe). Journal of Agricultural and Food Chemistry 53, 4388–4392 (2005). [DOI] [PubMed] [Google Scholar]
- Park I.-K. et al. Nematicidal activity of plant essential oils and components from garlic (Allium sativum) and cinnamon (Cinnamomum verum) oils against the pine wood nematode (Bursaphelenchus xylophilus). Nematology 7, 767–774 (2005). [Google Scholar]
- Francis F., Vandermoten S., Verheggen F., Lognay G. & Haubruge E. Is the (E)-β-farnesene only volatile terpenoid in aphids? J Appl Entomol 129, 6–11 (2005). [Google Scholar]
- Crock J., Wildung M. & Croteau R. Isolation and bacterial expression of a sesquiterpene synthase cDNA clone from peppermint (Mentha x piperita, L.) that produces the aphid alarm pheromone (E)-beta-farnesene. Proc. Natl. Acad. Sci. USA 94, 12833–12838 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis F., Lognay G. & Haubruge E. Olfactory Responses to Aphid and Host Plant Volatile Releases: (E)-β-Farnesene an Effective Kairomone for the Predator Adalia bipunctata. J Chem Ecol 30, 741–755 (2004). [DOI] [PubMed] [Google Scholar]
- Verheggen F. et al. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis pallas, to sesquiterpene semiochemicals. J Chem Ecol 33, 2148–2155 (2007). [DOI] [PubMed] [Google Scholar]
- Cui L.-L. et al. The functional significance of E-β-Farnesene: Does it influence the populations of aphid natural enemies in the fields? Biol Control 60, 108–112 (2012). [Google Scholar]
- Zhu J., Cossé A. A., Obrycki J. J., Boo K. S. & Baker T. C. Olfactory Reactions of the Twelve-Spotted Lady Beetle, Coleomegilla maculata and the Green Lacewing, Chrysoperla carnea to Semiochemicals Released from Their Prey and Host Plant: Electroantennogram and Behavioral Responses. J Chem Ecol 25, 1163–1177 (1999). [Google Scholar]
- Almohamad R., Verheggen F. J., Francis F. & Haubruge E. Predatory hoverflies select their oviposition site according to aphid host plant and aphid species. Entomol Exp Appl 125, 13–21 (2007). [Google Scholar]
- Wohlers P. Effect of alarm pheromone (E)-β-farnesene on aphid behaviour during flight and after landing on plants. Zeitschrift für Angewandte Entomologie 93, 102–108 (1982). [Google Scholar]
- Yu X. et al. (E)-β-Farnesene synthase genes affect aphid (Myzus persicae) infestation in tobacco (Nicotiana tabacum). Functional & Integrative Genomics. 1–7 (2011). [DOI] [PubMed] [Google Scholar]
- Edwards L. J., Siddall J. B., Dunham L. L., Uden P. & Kislow C. J. Trans-[beta]-farnesene, Alarm Pheromone of the Green Peach Aphid, Myzus persicae (Sulzer). Nature 241, 126–127 (1973).4121143 [Google Scholar]
- Pickett J. A. & Griffiths D. C. Composition of aphid alarm pheromones. J Chem Ecol 6, 349–360 (1980). [Google Scholar]
- Vandermoten S., Mescher M. C., Francis F., Haubruge E. & Verheggen F. J. Aphid alarm pheromone: An overview of current knowledge on biosynthesis and functions. Insect Biochemistry and Molecular Biology 42, 155–163 (2012). [DOI] [PubMed] [Google Scholar]
- Wei J. N. & Kang L. Roles of (Z)-3-hexenol in plant-insect interactions. Plant Signaling & Behavior 6, 369–371 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant G. G., Ryall K. L., Lyons D. B. & Abou-Zaid M. M. Differential response of male and female emerald ash borers (Col., Buprestidae) to (Z)-3-hexenol and manuka oil. J Appl Entomol 134, 26–33 (2010). [Google Scholar]
- Crook D. J. et al. Laboratory and Field Response of the Emerald Ash Borer (Coleoptera: Buprestidae), to Selected Regions of the Electromagnetic Spectrum. J Econ Entomol 102, 2160–2169 (2009). [DOI] [PubMed] [Google Scholar]
- Dorn S. et al. Response of female Cydia molesta (Lepidoptera: Tortricidae) to plant derived volatiles. Bull Entomol Res 93, 335–342 (2003). [DOI] [PubMed] [Google Scholar]
- Laubertie E. A., Wratten S. D. & Sedcole J. R. The role of odour and visual cues in the pan-trap catching of hoverflies (Diptera: Syrphidae). Ann Appl Biol 148, 173–178 (2006). [Google Scholar]
- SAS. SAS User’s Guide, Version 8.02. SAS Institute. Cary, NC, USA, (2001). [Google Scholar]
- Kunert G., Reinhold C. & Gershenzon J. Constitutive emission of the aphid alarm pheromone, (E)-β-farnesene, from plants does not serve as a direct defense against aphids. BMC Ecology 10, 23 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson R. W. & Pickett J. A. Wild potato repels aphids by release of aphid alarm pheromone. Nature 302, 608–609 (1983). [Google Scholar]
- Pickett J. A., Wadhams L. J. & Woodcock C. M. The chemical ecology of aphids. Ann. Rev. Entomol. 37, 67–90 (1992). [Google Scholar]
- Fan J. et al. Orco mediates olfactory behaviors and winged morph differentiation induced by alarm pheromone in the grain aphid, Sitobion avenae. Insect Biochemistry and Molecular Biology. 16–24 (2015). [DOI] [PubMed] [Google Scholar]
- Jiang S. S., Deng Q., Fan J., Sun J. R. & Chen J. L. Behavioral responses of Sitobion avenae (Hemiptera: Aphididae) to E-β-farnesene. Acta Entomologica Sinica 58, 776–782 (2015). [Google Scholar]
- Hatano E., Kunert G. & Weisser W. W. Aphid wing induction and ecological costs of alarm pheromone emission under field conditions. Plos One 5, e11188 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunert G., Otto S., Röse U. S., Gershenzon J. & Weisser W. W. Alarm pheromone mediates production of winged dispersal morphs in aphids. Ecology Letters 8, 596–603 (2005). [Google Scholar]
- Verheggen F. J., Haubruge E. & Mescher M. C. Chapter Nine - Alarm Pheromones-Chemical Signaling in Response to Danger. Vol. Volume 83 (Academic Press, 2010). [DOI] [PubMed] [Google Scholar]
- Shiojiri K. et al. Herbivore-Specific, Density-Dependent Induction of Plant Volatiles: Honest or “Cry Wolf” Signals? PLoS ONE 5, e12161 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickett J. A. & Glinwood R. In Aphids as crop pests (eds Van Emden H. F. & Harrington R.) 235–260 (CABI, 2007). [Google Scholar]
- Quiroz A. & Niemeyer H. M. Olfactometer-assessed responses of aphid Rhopalosiphum padi to wheat and oat volatiles. J Chem Ecol 24, 113–124 (1998). [Google Scholar]
- Cook S. M., Khan Z. R. & Pickett J. A. The use of push-pull strategies in integrated pest management. Annu Rev Entomol. 375–400 (2007). [DOI] [PubMed] [Google Scholar]
- James D. G. Synthetic Herbivore-Induced Plant Volatiles as Field Attractants for Beneficial Insects. Environ Entomol 32, 977–982 (2003). [DOI] [PubMed] [Google Scholar]
- Sabelis M. W. et al. Behavioural responses of predatory and herbivorous arthropods to induced plant volatiles: from evolutionary ecology to agricultural applications. (APS Press, 1999). [Google Scholar]