

Abstract
Dietary structural material has been reported to improve broiler live performance and gastrointestinal tract (GIT) function. In this 50 d cage study, the effects of coarsely ground corn (CC) inclusion on broiler live performance, GIT development, apparent ileal digestibility (AID) of energy and nitrogen (N), and digesta particle size distribution and retention time were investigated. This study included 3 CC inclusions (0, 25, and 50% fine corn [FC] replaced by CC), with 6 replicate cages of 10 birds per treatment. The feed conversion ratio (FCR) at 35 and 42 d was improved (P < 0.01) as the dietary inclusion of CC increased without effect on feed intake. The 50% CC diet increased absolute and relative gizzard weight at 42 d of age as compared to diets with 0 and 25% CC (P < 0.01). Dietary CC increased absolute proventriculus weight at 28 d of age (P < 0.05). A numerically lower gizzard digesta pH (P < 0.08) was observed at 28 d but not 42 d of age, and there was no difference in proventriculus, jejunum, or ileum digesta pH at 28 or 42 d of age. The 25 and 50% CC treatments increased the digesta retention time at 30 and 45 d of age (P < 0.05 and P < 0.01, respectively). The 25 and 50% CC treatments improved AID of energy by 7.1 and 8.2%, respectively, when compared with the 0% CC treatment, and they improved AID of N by 12.2 and 12.4%, respectively (P < 0.01). The digesta particles in the jejunum exhibited a similar distribution, with a dgw (geometric mean diameter by mass) of 218, 204, and 181 μm when 0, 25, of 50% CC diets were consumed, respectively. In conclusion, birds fed pelleted and screened diets that contained 25 and 50% CC exhibited increased BW, improved FCR, and increased AID of energy and N, which was probably due to enhanced gizzard development and greater digesta retention time.
Keywords: coarse corn, gizzard, gastrointestinal tract development, digestibility, digesta retention time
INTRODUCTION
Feed particle size reduction has long remained the central paradigm of feed milling, but feed particle size has a paradoxical role in poultry nutrition. From a feed manufacturing perspective, smaller feed particle size has been found to improve mixing efficiency and uniformity, reduce ingredient segregation during further handling, and improve pelleting efficiency and quality (Behnke, 2001). However, as the particle size of the grain decreases, the cost to grind the grain increases (Behnke, 2001). From a nutritional perspective, smaller particle grind size increases relative surface area, improves digestibility by exposing ingredient substrates to digestive enzymes, and reduces selective feeding by the animal. However, birds have been found to have an instinctive preference for coarse feed particles to stimulate gizzard function, which has been suggested controls gut motility (Duke et al., 1977; Ferket and Gernat, 2006).
A fully functional gizzard has been thought to improve nutrient digestibility though enhanced gastrointestinal tract (GIT) function and greater digesta retention time (Amerah et al., 2008). The development and integrity, digestive functions, maintenance requirement, and digesta retention time of the GIT have all been reported to be influenced by enhanced gizzard activity, which has been demonstrated to respond quickly to changes in feed structural material under certain circumstances (Hetland et al., 2002, 2003; Gabriel et al., 2003, 2008; Senkoylu et al., 2009). The importance of the physical structure of the diet as a means to improve feed efficiency and live performance has become increasingly recognized, and coarser feed structure has exhibited a positive influence on nutrient digestibility and animal live performance (Nir et al., 1995; Amerah et al., 2008; Samu et al., 2010). A better understanding of the interaction between feed structure and GIT development and function has become critical to optimization of feed manufacturing strategies and broiler live performance. The objectives of the current study were to evaluate the effects of dietary inclusion of roller mill coarsely ground corn (CC) on broiler live performance, GIT development and function, apparent ileal digestibility (AID) of energy and nitrogen (N), and digesta particle size distribution and retention time.
MATERIALS AND METHODS
Feed Formulation, Manufacture, and Experiment Design
Broiler diets (Table 1) were formulated and manufactured to meet or exceed the NRC suggested minimum requirements of broilers (NRC, 1994). The same crumbled starter diet was used for all birds, while three coarsely ground corn (CC) inclusions in the grower and finisher diets were produced by replacing 0, 25, or 50% of fine corn (FC) with CC. The FC and soybean meal (SBM) were ground with a hammermill (Model 1522, Roskamp Champion, Waterloo, IA) equipped with two 2.4 mm screens, while the CC was ground with a two-pair roller mill (Model C128829, RMS, Tea, SD) with a gap setting of 0% opening (0.15 to 0.18 mm) on the top pair of rollers and 100% opening (7.16 mm) on the bottom pair of rollers. Dry ingredients were blended in a double ribbon mixer (TRDB126-0604, Hayes & Stolz Ind. Mfg. Co., Fort Worth, TX) to produce the mash diet, which was conditioned at 85°C for 45 s and pelleted with a ring die (4.4 by 35 mm) pellet mill (Model PM1112-2, California Pellet Mill Co., Crawfordsville, IN). Pellets were cooled with ambient air in a counterflow cooler (Model VK09 × 09KL, Geelen Counterflow USA Inc., Orlando, FL). The starter feed was provided as crumbles, and subsequent feeds were fed as pellets, with fines removed with a pellet screener (Model 35/7 Roto-Shaker, Sprout Bauer, Inc., Muncy, PA). Particle size distribution was determined by ASAE Standard S319.3 (ASAE Standards, 2003a), and the pellet durability index was determined by ASAE Standard S269.4 (ASAE Standards, 2003b). The particle size method included agitators (14 rubber balls, 5 plastic cleaners, and 4 bristle sieve cleaners), which were placed on sieve numbers 8 to 270 to break up agglomerations. A dispersing agent (silicon dioxide SSA-58, Gilson, Lewis Center, OH) was also added at 0.5 g/100 g sample prior to shaking to facilitate material movement through the sieves (Stark and Chewning, 2012).
Table 1.
Ingredient composition and calculated analysis for broiler starter, grower, and finisher diets
| Item | Starter | Grower | Finisher |
|---|---|---|---|
| Ingredient | % | ||
| Corn | 50.51 | 54.19 | 61.20 |
| Soybean meal (SBM; 48% CP) | 33.79 | 23.22 | 16.07 |
| Distillers dried grains with solubles (DDGS) | 6.00 | 15.00 | 15.00 |
| Poultry byproduct meal | 2.00 | 1.00 | 1.00 |
| Limestone | 0.97 | 0.93 | 0.95 |
| Dicalcium phosphate, 18% P | 1.93 | 1.41 | 1.46 |
| DL-Methionine | 0.28 | 0.13 | 0.10 |
| L-Lysine | 0.12 | 0.22 | 0.28 |
| L-Threonine | 0.12 | 0.12 | 0.08 |
| Sodium chloride | 0.50 | 0.50 | 0.50 |
| Vitamin premix1 | 0.05 | 0.05 | 0.05 |
| Choline chloride, 60% | 0.10 | 0.10 | 0.10 |
| Trace mineral premix2 | 0.20 | 0.20 | 0.20 |
| Selenium premix3 | 0.10 | 0.10 | 0.10 |
| Coccidiostat4 | 0.05 | 0.05 | 0.05 |
| Poultry fat | 3.23 | 2.76 | 2.85 |
| 100.00 | 100.00 | 100.00 | |
| Calculated analysis | |||
| ME, kcal/g | 2.85 | 2.90 | 2.95 |
| Protein, % | 23.00 | 20.00 | 17.00 |
| Calcium, % | 1.00 | 0.80 | 0.80 |
| Available phosphorus, % | 0.50 | 0.40 | 0.40 |
| Total lysine, % | 1.31 | 1.13 | 0.97 |
| Total methionine + cysteine, % | 1.00 | 0.82 | 0.71 |
| Analyzed nutrients | |||
| Gross Energy, kcal/g | 3.61 | 3.71 | 3.73 |
| CP, % | 22.3 | 21.2 | 18.0 |
1The vitamin premix supplied the following per kg of feed: vitamin A, 6601 IU; cholecalciferol, 1980 IU; niacin, 55 mg; α-tocopheral, 33 mg; pantothenic acid, 11 mg; riboflavin, 6.6 mg; pyridoxine, 4 mg; menadione, 2 mg; thiamin, 2 mg; folic acid, 1.1 mg; biotin, 0.13 mg; and vitamin B12, 0.02 mg.
2The mineral premix supplied the following per kg of feed: Zn, 120 mg; Mn, 120 mg; Fe, 80 mg; Cu, 10 mg; I, 2.5 mg; Co, 1.0 mg.
3Selenium premix provided 0.2 ppm Se.
4Monensin was included at 99 mg/kg (Coban 90, Elanco Animal Health, Indianapolis, IN).
Husbandry Practices
The care of the birds in the study conformed to the Guide for Care and Use of Agricultural Animals in Research and Teaching (FASS, 2010). Ross 344 × 708 (Aviagen, Huntsville, AL) 1-d-old male broiler chicks were feather sexed, weighed, and randomly distributed among 18 cages with 10 chicks each within 3 custom-designed brooding battery cages (Alternative Design, Siloam Springs, AR) in an environmentally controlled room. Care was taken to distribute incubator position effects uniformly among all of the broiler pens. Each pen was 61 cm in width, 46 cm in length, and 61 cm in height. Each cage was randomly assigned to 1 of 3 dietary treatments (0, 25, or 50% CC), with a total of 6 replicates per treatment. The chicks were given a budget of 0.9, 2.7, and 3.6 kg starter, grower, and finisher diet, respectively. Each cage had 2 nipple drinkers and 1 feeder for ad libitum consumption of feed and water, respectively. The lighting program consisted of 23 h of light and 1 h of darkness for the 50 d experimental period. The room temperature was 35°C from 1 to 2 d, 32 to 33°C from 3 to 7 d, 30 to 32°C from 8 to 15 d, and then 22 to 30°C from 16 to 50 d.
Data Collection
Initial pen group BW was determined at placement. Feed intake and BW by cage were recorded at 14, 28, 35, and 42 d of age. Birds were observed twice daily, and mortalities were removed and their BW was included in the feed conversion ratio (FCR) calculation. One bird per cage at 28 d and two birds per cage at 42 d of age of average BW were weighed individually, killed by cervical dislocation, and had gizzard and proventriculus excision. The surrounding fat was trimmed. Then organ contents were removed, rinsed, and blotted dry and then weighed and expressed as a percentage of BW (mg/g BW). The pH of the gizzard and proventriculus contents was measured with a portable pH meter (HACH IQ150 pH/mV/Temperature System, Loveland, CO). Viscera were excised, and gut section (duodenum, jejunum, ileum, and colon) lengths, as well as total gut length (from gizzard pylorus to distal colon) were measured. A 15 cm section of jejunum and ileum were excised, digesta content was removed, and fat was trimmed for tensile strength measurement using an Instron Tensile Tester 5542 w/ 50N load cell (Instron Corp., Norwood, MA). Bluehill 2 software was used for data collection and analysis (Instron proprietary software). The particle size distribution of jejunum digesta content was measured with a laser diffraction particle size analyzer (Model LS 13 320, Beckman Coulter, Inc., Brea, CA).
Celite was added to the finisher diet as an indigestible marker to determine apparent ileal digestibility (AID) of energy and nitrogen (N). At 50 d of age, 3 birds of average BW per cage were necropsied to collect ileal content. Ileal digesta were collected (2 cm posterior of Meckel's diverticulum to 2 cm anterior of the ileal-cecal junction) by gently squeezing the ileal contents in a manner that provided a sample sufficient for digestibility analysis. Ileal content was analyzed for moisture (Method 934.01, AOAC, 2006a), crude protein (Method 990.03, AOAC, 2006b), acid insoluble ash (Vogtmann et al., 1975), and gross energy (Merrill and Watt, 1973). Gross energy was determined using an adiabatic bomb calorimeter (Model C5003, IKA, Wilmington, NC). Apparent ileal digestibility of energy and N were calculated using the following equation:
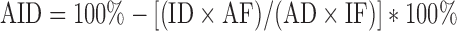 |
Where:
AID = Apparent ileal digestibility (%)
ID = Marker concentration in diet (%)
AF = Energy or N concentration in ileal digesta (%)
AD = Energy or N concentration in diet (%)
IF = Marker concentration in ileal digesta (%)
Digesta retention time was determined at 30 and 45 d of age. After a 12 h feed withdrawal period, 6 birds per treatment (a total of 18 birds) were put into individual cages, each bird was orally administered a gelatin capsule that contained 85 mg chromium oxide as an indigestible marker, and the birds were returned to ad libitum feeding. Beginning 1 h after the gelatin capsule was administrated, feces were observed every 15 min, and the time of the appearance of clear green–colored feces was recorded as the passage time.
Statistical Analysis
Results were analyzed as a one-way treatment structure using a completely randomized design of CC inclusion (0, 25, or 50%). The cage served as the experimental unit for the statistical analysis of the live performance data. All data were analyzed using GLM procedure of SAS (SAS Institute, 2012). Differences were considered significant at P < 0.05 or P < 0.01 as indicated, and the differences between means were separated by least significant difference.
RESULTS AND DISCUSSION
The geometric mean diameter by mass (dgw) and particle size distribution of fine corn (FC), course corn (CC), and mash diets prior to pelleting with 0, 25, and 50% CC replacing FC are shown in Figures 1 and 2. The dgw of FC, CC, soybean meal (SBM), and distillers dried grains with solubles (DDGS) prior to mixing was 294; 1,362; 491; and 414 μm, respectively. As the 0, 25, and 50% CC replaced FC in the diets, the dgw of mash diets prior to pelleting increased in a stepwise manner from 432 to 640 μm, respectively. The hammermill and roller mill have been two grinding devices commonly used to reduce the particle size of feed grains. Hammermill grinding creates a wider particle size distribution, while roller mill grinding creates a more compact or narrow particle size distribution (Nir et al., 1995). By blending corn ground by these two grinding devices, a biphasic distribution of particle size (as demonstrated in Figure 1) was produced, with the intention of having larger particles stimulate gizzard function and smaller particles help maintain pellet quality and improve digestion and nutrient absorption. Pellet quality of the 0, 25, and 50% CC diets as determined by the pellet durability index (PDI) was 92, 93, and 90%, respectively, with a pellet production rate of 522, 454, and 438 kg/h, respectively. As demonstrated by Angulo et al. (1996), pellet durability is inversely related to particle size, likely because smaller particles have more contact points within the pellet (Behnke, 2001). However, particle size is not the only factor that has been reported to influence pellet quality. Thomas and Poe (1996) and Briggs et al. (1999) reported that ingredient composition, steam condition, production output, and die and roller dimension greatly influence final pellet quality.
Figure 1.

The geometric mean diameter by mass (dgw) and particle size distribution of fine corn (FC) and coarse corn (CC) prior to mixing.
Figure 2.

The geometric mean diameter by mass (dgw) and particle size distribution of mash diets after mixing but prior to pelleting with 0, 25, and 50% coarse corn (CC) replacing fine corn (FC).
The effects of dietary CC inclusion on feed intake, BW, and feed conversion ratio (FCR) of broilers are shown in Table 2. There were no significant treatment effects on live performance parameters at 14 d of age. The birds’ feed intake was similar among all treatments, but greater BW was obtained at 28 (P < 0.01), 35 (P < 0.01), and 42 d of age (P < 0.05) when birds were fed 25 and 50% CC. Improved FCR was observed at 35 d and 42 d for the birds fed 25 and 50% CC diets, respectively, as compared to those fed 0% CC (P < 0.01). Inconsistent results of the influence of corn particle size on broiler live performance have been reported to be due to differences in feed form (Reece et al., 1985; Chewning et al., 2012), corn dgw and particle size distribution (Reece et al., 1986; Dozier et al., 2006), and pellet quality (Parsons et al., 2006). Lott et al. (1992) observed improved broiler live performance when corn particle size decreased from 1173 to 710 μm, but Nir et al. (1994) reported improved broiler live performance when corn particle size increased from 525 to 897 μm. Reece et al. (1986) reported that both fine (679 μm) and coarse (1289 μm) corn improved BW gain and feed efficiency as compared with medium ground corn (987 μm). They also found that broilers fed a pelleted diet containing a 50% FC and 50% CC combination (908 μm) performed better than broilers fed medium ground corn (987 μm), confirming that particle size distribution is important. In the present study, a single crumbled starter (0 to 14 d) without CC was employed, and thereafter, 100% pellets (15 to 49 d) with 25 or 50% CC inclusion feed were employed. Similar feed intake and improved live performance were observed due to 25 and 50% CC when compared to 0% CC. This procedure was followed because broiler chicks may not have the gizzard capacity to fully utilize a diet containing 50% CC at an early age (Lily et al., 2011), and pellet quality could be a confounding factor that would be inversely related to CC inclusion.
Table 2.
Effect of dietary coarse corn (CC) inclusion on feed intake, BW, and feed conversion ratio (FCR) of broilers at 14, 28, 35, and 42 d of age
| CC1 | n | Feed intake | BW | FCR2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 14 d | 28 d | 35 d | 42 d | 14 d | 28 d | 35 d | 42 d | 14 d | 28 d | 35 d | 42 d | ||
| (g) | (g/g) | ||||||||||||
| 0% | 6 | 561 | 2387 | 3653 | 5257 | 474 | 1621B | 2284B | 2929b | 1.31 | 1.77 | 1.82A | 1.94A |
| 25% | 6 | 570 | 2406 | 3670 | 5350 | 477 | 1720A | 2400A | 3118a | 1.32 | 1.73 | 1.74B | 1.86B |
| 50% | 6 | 561 | 2355 | 3613 | 5154 | 469 | 1696A | 2408A | 3059a | 1.33 | 1.69 | 1.69B | 1.82B |
| SEM3 | 8 | 37 | 71 | 113 | 6 | 20 | 31 | 43 | 0.02 | 0.03 | 0.02 | 0.02 | |
| P-value | 0.58 | 0.56 | 0.81 | 0.42 | 0.53 | < 0.01 | 0.01 | 0.02 | 0.65 | 0.12 | < 0.01 | < 0.01 | |
a-bMeans within a column with different superscripts differ significantly (P < 0.05).
A-BMeans within a column with different superscripts differ significantly (P < 0.01).
1Treatments consisted of diets with 0, 25, or 50% CC inclusion.
2FCR = Feed intake per pen/total pen BW gain, including BW of mortality that occurred during each time period.
3SEM = Standard error of the means for CC inclusions effect (n = 6).
The effects of dietary CC inclusion on absolute and relative gizzard and proventriculus weight, as well as gizzard to proventriculus ratio, are shown in Table 3. The 25 and 50% CC treatments resulted in a marginal increase (P = 0.09) in absolute gizzard weight at 28 d of age as compared to the 0% CC inclusion treatment. The 25 and 50% CC treatments increased both absolute and relative gizzard weight at 42 d of age in a stepwise manner as compared to the 0% CC treatment (P < 0.01). Dietary 25 and 50% CC increased absolute proventriculus weight at 28 d of age as compared to 0% CC (P < 0.05), but no other differences were observed for absolute and relative proventriculus weight at either 28 or 42 d of age. Gizzard to proventriculus ratio was increased at 42 d of age by 25 and 50% CC as compared to 0% CC (P < 0.01). Several researchers have reported that gizzard weight increases with increased corn particle size independently of the physical form of the diet (Oliver and Jonker, 1997; Dahlke et al., 2003). Parsons et al. (2006) reported that absolute and relative gizzard weight increase as corn particle size increases (from 781; 950; 1,042; 1,109; to 2,242 μm). Healy (1992) reported increased weights of gizzard and proventriculus for broilers fed 900 μm corn compared with 300 μm corn. Apparently, birds adjusted their mechanical and enzymatic digestion according to feed structure. Furthermore, Gabriel et al. (2008) reported that dietary whole wheat inclusion increased pancreas weight by 12%, but CC had no effect in the present study.
Table 3.
Effect of dietary coarse corn (CC) inclusion on absolute and relative weight of gizzard, proventriculus, and pancreas, and gizzard to proventriculus ratio at 28 and 42 d of age
| CC1 | n | Gizzard | Proventriculus | G/P Ratio3 | Pancreas | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 28 d | 42 d | 28 d | 42 d | 28 d | 42 d | 28 d | 42 d | 28 d | 42 d | 42 d | |||
| (g) | (mg/g BW) | (g) | (mg/g BW) | (g/g) | (g) | (mg/g BW) | |||||||
| 0% | 6 | 13.47 | 20.96C | 7.81 | 6.60C | 4.31b | 10.26 | 2.50 | 3.26 | 3.12 | 2.11B | 4.61 | 1.36 |
| 25% | 6 | 17.46 | 25.73B | 10.32 | 7.63B | 5.28a | 9.97 | 3.18 | 2.97 | 3.21 | 2.84A | 4.57 | 1.30 |
| 50% | 6 | 17.94 | 28.34A | 9.43 | 8.75A | 5.48a | 8.77 | 2.88 | 2.71 | 3.28 | 3.44A | 4.72 | 1.33 |
| SEM2 | 1.46 | 0.82 | 0.94 | 0.35 | 0.31 | 0.65 | 0.19 | 0.22 | 0.27 | 0.17 | 0.18 | 0.04 | |
| P-value | 0.09 | <0.01 | 0.19 | <0.01 | 0.04 | 0.20 | 0.08 | 0.20 | 0.91 | < 0.01 | 0.83 | 0.67 | |
a-cMeans within a column with different superscripts differ significantly (P < 0.05).
A-BMeans within a column with different superscripts differ significantly (P < 0.01).
1Treatments consisted of diets with 0, 25, or 50% CC inclusion.
2SEM = Standard error of the means for CC inclusions effect (n = 6).
3G/P Ratio = Gizzard : proventriculus.
The effect of dietary CC inclusion on digesta pH and intestinal length at 28 and 42 d of age is shown in Table 4. A numerically lower pH of gizzard digesta (P < 0.08) but not proventriculus digesta was observed at 28 d of age, and there were no effects for either at 42 d. There was also no difference in jejunum and ileum digesta pH at 28 or 42 d of age. The pH was higher for gizzard contents and lower for intestinal contents than typically reported in the literature, which may be due to differences in digesta quantity, retention time, and the chemical characteristics of water, litter, and feed in the gizzard-proventriculus region. Cages also prevented consumption of litter fiber that could have influenced gastrointestinal tract (GIT) function of these rapidly growing broilers. Several reports have demonstrated that dietary structure, such as coarse grain or fiber, decreases the pH of gizzard contents by a magnitude of between 0.2 and 1.2 units (Dahlke et al., 2003; Engberg et al., 2004; Gabriel et al., 2008; Senkoylu et al., 2009). Diet nutrient composition, water intake, feed consumption, and the tightly regulated acid–base balance of an animal could have been major confounding factors that masked any pH effect. Intestinal segment length as well as the ratio of total length to BW was not influenced by dietary CC inclusion at 42 d of age.
Table 4.
Effect of dietary coarse corn (CC) inclusion on digesta pH and intestinal length at 28 and 42 d of age
| CC1 | n | Digesta pH at 28 d | Digesta pH at 42 d | Intestinal length at 42 d | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Proventriculus | Gizzard | Jejunum | Ileum | Jejunum | Ileum | Duodenum | Jejunum | Ileum | Colon | Total length | Length/ | ||
| length | BW | ||||||||||||
| (cm) | (cm/kg) | ||||||||||||
| 0% | 6 | 5.43 | 4.67 | 5.86 | 6.17 | 6.00 | 6.32 | 15.08 | 75.54 | 79.58 | 36.33 | 185.29 | 54.49 |
| 25% | 6 | 5.21 | 4.38 | 5.76 | 6.22 | 6.13 | 6.26 | 15.96 | 74.21 | 77.75 | 39.08 | 183.88 | 52.18 |
| 50% | 6 | 5.11 | 4.10 | 5.82 | 6.29 | 6.06 | 6.36 | 15.92 | 75.46 | 79.17 | 39.51 | 186.46 | 52.70 |
| SEM2 | 0.12 | 0.16 | 0.05 | 0.07 | 0.11 | 0.17 | 0.39 | 1.11 | 1.90 | 0.56 | 3.29 | 0.92 | |
| P-value | 0.20 | 0.08 | 0.72 | 0.55 | 0.69 | 0.88 | 0.21 | 0.64 | 0.78 | 0.20 | 0.86 | 0.19 | |
1Treatments consisted of diets with 0, 25, or 50% CC inclusion.
2SEM = Standard error of the means for CC inclusions effect (n = 6).
There have been conflicting reports concerning how the relative size of intestinal segments is modified by coarse diet structure. Amerah et al. (2007) reported that all gut segments were shorter in birds fed coarsely ground grain particles than those fed finer grain particles in a mash diet, but there were no differences found in gut length when a pelleted feed form was used. Taylor and Jones (2004) found greater duodenum length with whole wheat (200 g/kg in a pelleted diet) but no difference in duodenum weight. No difference in gut segment length was observed in the present study, which may be due to a 45-d-old broiler being relatively mature and consuming screened pellets.
The effect of dietary CC inclusion on intestinal tensile strength at 28 and 42 d, digesta retention time at 30 and 45 d, and apparent ileal digestibility (AID) of energy and nitrogen (N) at 50 d of age are shown in Table 5. Dietary inclusion of 50% CC tended to increase (P < 0.08) the tensile strength of the ileum at 28 d of age compared to the 0% CC treatment. The 25 and 50% dietary CC inclusion increased digesta retention time at 30 and 45 d of age (P < 0.05 and P < 0.01, respectively), and there was a difference between these two CC treatments at 30 d but not at 45 d, which probably reflected increased GIT maturity. In comparison to the 0% inclusion treatment, 25 and 50% CC treatments improved AID of energy by 7.1 and 8.2%, respectively. The AID of N was less than expected but could have been negatively influenced by the inclusion of digesta from the proximal third of the ileum in the present samples, as Kluth et al. (2005) reported significantly reduced digestibility in this segment of the ileum. Nevertheless, the AID of N was improved by the presence of 25 and 50% CC by 12.2 and 12.4%, respectively, but there was no difference between the 25 and 50% CC treatments. Sacranie (2006) hypothesized that dietary inclusion of larger feed particles increased intestinal tensile strength due to increased muscular activity associated with reverse peristalsis. However, Dozier et al. (2006) reported that intestinal tensile strength was not affected by dietary CC treatments (15, 25, and 35% CC). The increased digesta retention time observed with coarse feed particles in the present study corroborated other research reports. Larger particles have been reported to be retained within the digestive tract longer than fine particles (Denbow, 2000) because coarse feed particles need to be ground to a certain critical size before they can be expelled from the gizzard (Clemens et al., 1975; Moore, 1999). A number of studies have shown improved nutrient utilization when birds are fed structural material in their diet (Preston et al., 2000; Svihus and Hetland, 2001; Rougiere et al., 2009). Parsons et al. (2006) reported coarse grinding of corn increases the efficiency of N and lysine retention in broiler diets. The improved digestibility of nutrients may arise from lower digesta pH and enhanced peptic digestion as well as increased enzyme-substrate interaction because of the greater retention time elicited by dietary CC inclusion.
Table 5.
Effect of dietary coarse corn (CC) inclusion on intestine tensile strength at 28 and 42 d, digesta retention time at 30 and 45 d, and apparent ileal digestibility of energy and nitrogen at 50 d of age
| CC1 | n | Tensile strength | Digesta retention time | Apparent ileal digestibility | |||||
|---|---|---|---|---|---|---|---|---|---|
| Jejunum | Ileum | Jejunum | Ileum | Energy | Nitrogen | ||||
| 28d | 42d | 30 d | 45 d | 50 d | |||||
| (N) | (h) | (%) | |||||||
| 0% | 6 | 2.35 | 1.90 | 2.98 | 2.43 | 1.58c | 3.54B | 54.6B | 44.0B |
| 25% | 6 | 2.10 | 2.07 | 3.18 | 2.78 | 1.75b | 4.52A | 61.7A | 56.1A |
| 50% | 6 | 2.32 | 2.69 | 3.04 | 2.65 | 1.96a | 4.32A | 62.8A | 56.4A |
| SEM2 | 0.31 | 0.19 | 0.56 | 0.39 | 0.09 | 0.27 | 4.3 | 6.5 | |
| P-value | 0.56 | 0.08 | 0.66 | 0.71 | <0.05 | <0.01 | <0.01 | <0.01 | |
a-cMeans within a column with different superscripts differ significantly (P < 0.05).
A-BMeans within a column with different superscripts differ significantly (P < 0.01).
1Treatments consisted of diets with 0, 25, or 50% CC inclusion.
2SEM = Standard error of the means for CC inclusions effect (n = 6).
The dgw and particle size distribution of jejunum digesta at 45 d are shown in Figure 3. The dgw of jejunum digesta was 218, 204, and 181 μm with 0, 25, and 50% CC inclusion, respectively. As expected, these three treatments had similar jejunum digesta particle size distribution; Clemens et al. (1975) and Moore (1999) reported that coarse feed particles needed to be ground to a certain critical size before they could exit the gizzard. Ferrando et al. (1987) suggested that the threshold particle size is between 500 and 1,500 μm before it will leave the gizzard in chickens. Hetland et al. (2002, 2003) as well as Amerah et al. (2008) reported that a majority of the particles entering the duodenum are smaller than 100 μm, even when considerable amounts of whole wheat or coarse grain particles are added to the diet.
Figure 3.

The geometric mean diameter by mass (dgw) and particle size distribution of jejunum digesta of birds at 42 d of age fed 0, 25, and 50% coarse corn (CC) diets.
Acknowledgments
This material is based on research supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, under Agreement No. 2010-1375.
REFERENCES
- Amerah A. M., Ravindran V., Lentle R. G., Thomas D. G. Performance and digestive tract characteristics of broilers as influenced by particle size and feed form. Proc. Aust. Poult. Sci. Symp. 2007;19:85–88. doi: 10.3382/ps.2007-00212. [DOI] [PubMed] [Google Scholar]
- Amerah A. M., Ravindran V., Lentle R. G., Thomas D. G. Influence of feed particle size on the performance, energy utilization, digestive tract development, and digesta parameters of broiler starter fed wheat- and corn-based diets. Poult. Sci. 2008;87:2320–2328. doi: 10.3382/ps.2008-00149. [DOI] [PubMed] [Google Scholar]
- Angulo E., Brufau J., Esteve-Garcia E. Effect of a sepiolite product on pellet durability in pig diets differing in particle size and in broiler starter and finisher diets. Anim. Feed Sci. Tech. 1996;63:25–34. [Google Scholar]
- AOAC. Moisture in animal feed, loss in drying at 95–100°C. AOAC Official Method 934.01; 2006a. [Google Scholar]
- AOAC. Combustion analysis (LECO) AOAC Official Method 990.03; 2006b. [Google Scholar]
- ASAE Standards. Method of determining and expressing fineness of feed materials by sieving. St. Joseph, MI: Am. Soc. Agric. Eng.; 2003a. S319.3. [Google Scholar]
- ASAE Standards. Cubes, pellets, and crumbles-definitions and methods for determining density, durability and moisture content. St. Joseph, MI: Am. Soc. Agric. Eng.; 2003b. S269.4. [Google Scholar]
- Behnke K. C. Factors influencing pellet quality. Feed Tech. 2001;5:19–22. [Google Scholar]
- Briggs J. L., Maier D. E., Watkins B. A., Behnke K. C. Effect of ingredients and processing parameters on pellet quality. Poult. Sci. 1999;78:1464–1471. doi: 10.1093/ps/78.10.1464. [DOI] [PubMed] [Google Scholar]
- Chewning C. G., Stark C. R., Brake J. Effects of particle size and feed form on broiler performance. J. Appl. Poult. Res. 2012;21:830–837. [Google Scholar]
- Clemens E. T., Stevens C. E., Southworth M. Sites of organic acid production and pattern of digesta movement in the gastrointestinal tract of geese. J. Nutr. 1975;105:1341–1350. doi: 10.1093/jn/105.10.1341. [DOI] [PubMed] [Google Scholar]
- Dahlke F., Ribeiro A. M. L., Kessler A. M., Lima A. R., Maiorka A. Effects of corn particle size and physical form of the diet on the gastrointestinal structures of broiler chickens. Br. Poult. Sci. 2003;5:61–67. [Google Scholar]
- Denbow D. M. Gastrointestinal anatomy and physiology. In: Whittow G. C., editor. Sturkie's Avian Physiology. New York, NY: Acad. Press; 2000. pp. 299–325. Pages. [Google Scholar]
- Dozier W. A. III., Behnke K., Kidd M. T., Branton S. L. Effects of the addition of roller mill ground corn to pelleted feed on pelleting parameters, broiler performance, and intestinal strength. J. Appl. Poult. Res. 2006;15:236–244. [Google Scholar]
- Duke G. E., Kuhlman W. D., Fedde M. R. Evidence for mechanoreceptors in the muscular stomach of the chicken. Poultry Sci. 1977;56:297–299. doi: 10.3382/ps.0560297. [DOI] [PubMed] [Google Scholar]
- Engberg R. M., Hedemann M. S., Steenfeldt S., Jensen B. Influence of whole wheat and xylanase on broiler performance and microbial composition and activity in the digestive tract. Poult. Sci. 2004;83:925–938. doi: 10.1093/ps/83.6.925. [DOI] [PubMed] [Google Scholar]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching. 3rd ed. Champaign, IL: Fed. Anim. Sci. Soc.; 2010. [Google Scholar]
- Ferket P. R., Gernat A. G. Factors that affect feed intake of meat birds: a review. Int. J. of Poult. Sci. 2006;5:905–911. [Google Scholar]
- Ferrando C., Vergara P., Jimenez M., Gonalons E. Study of the rate of passage of food with chromium-mordanted plant cells in chickens (Gallus gallus) Q. J. Exp. Physiol. 1987;72:252–259. doi: 10.1113/expphysiol.1987.sp003072. [DOI] [PubMed] [Google Scholar]
- Gabriel I., Mallet S., Leconte M. Differences in the digestive tract characteristics of broiler chickens fed on complete pelleted diet or whole wheat added to pelleted protein concentrate. Br. Poult. Sci. 2003;44:283–290. doi: 10.1080/0007166031000096470. [DOI] [PubMed] [Google Scholar]
- Gabriel I., Mallet S., Leconte M., Travel A., Lalles J. P. Effects of whole wheat feeding on the development of the digestive tract of broiler chickens. Anim. Feed Sci. Tech. 2008;142:144–162. [Google Scholar]
- Hetland H., Svihus B., Olaisen V. Effect of feeding whole cereals on performance, starch digestibility and duodenal particle size distribution in broiler chickens. Br. Poult. Sci. 2002;43:416–423. doi: 10.1080/00071660120103693. [DOI] [PubMed] [Google Scholar]
- Hetland H., Svihus B., Krogdahl A. Effects of oat hulls and wood shavings on digestion in broilers and layer fed diets based on whole or ground wheat. Br. Poult. Sci. 2003;44:275–282. doi: 10.1080/0007166031000124595. [DOI] [PubMed] [Google Scholar]
- Healy B. J. MS Thesis. Manhattan, KS: Kansas State University; 1992. Nutritional value of selected sorghum grain for swine and poultry and effect of particle size on performance and intestinal morphology in young pigs and broiler chicks. [Google Scholar]
- Kluth H., Mehlhorn K., Rodehutscord M. Studies on the intestine section to be sampled in broiler studies on precaecal amino acid digestibility. Arch. Anim. Nutr. 2005;59:271–279. doi: 10.1080/17450390500217058. [DOI] [PubMed] [Google Scholar]
- Lily K. G. S., Gehring C. K., Beaman K. R., Turk P. J., Sperow M., Moritz J. S. Examining the relationships between pellet quality, broiler performance, and bird sex. J. Appl. Poult. Res. 2011;20:231–239. [Google Scholar]
- Lott B. D., Day E. J., Deaton J. W., May J. D. The effect of temperature, dietary energy level, and corn particle size on broiler performance. Poult. Sci. 1992;71:618–624. doi: 10.3382/ps.0710618. [DOI] [PubMed] [Google Scholar]
- Merrill A. L., Watt B. K. Agriculture Handbook, No.74. Washington, DC: U.S. Government Printing Office; 1973. Energy Value of Foods-Basis and Derivation. [Google Scholar]
- Moore S. J. Food breakdown in an avian herbivore: Who needs teeth? Austr. J. Zool. 1999;47:625–632. [Google Scholar]
- Nir I., Hillel R., Shefet G., Nitsan Z. Effect of grain particle size on performance. II. Grain texture interactions. Poult. Sci. 1994;73:781–79. doi: 10.3382/ps.0730781. [DOI] [PubMed] [Google Scholar]
- Nir I., Hillel R., Ptichi I., Shefet G. Effect of particle size on performance. III. Grinding pelleting interactions. Poult. Sci. 1995;74:771–783. doi: 10.3382/ps.0740771. [DOI] [PubMed] [Google Scholar]
- NRC. Nutrient Requirements of Poultry. 9th rev. ed. Washington, DC: Natl. Acad. Press; 1994. [Google Scholar]
- Oliver M. D., Jonker A. Effect of choice feeding on the performance of broilers. Br. Poult. Sci. 1997;35:571–576. doi: 10.1080/00071669708418038. [DOI] [PubMed] [Google Scholar]
- Parsons A. S., Buchanan N., Blemings K., Wilson M., Moritz J. Effect of corn particle size and pellet texture on broiler performance in the growing phase. J. Appl. Poult. Res. 2006;15:245–255. [Google Scholar]
- Preston C. M., McCracken K. J., McAllister A. Effect of diet form and enzyme supplementation on growth. Br. Poult. Sci. 2000;41:324–331. doi: 10.1080/713654933. [DOI] [PubMed] [Google Scholar]
- Reece F. N., Lott B. D., Deaton J. W. The effects of feed form, grinding method, energy level, and gender on broiler performance in a moderate environment. Poult. Sci. 1985;64:1834–1839. doi: 10.3382/ps.0631906. [DOI] [PubMed] [Google Scholar]
- Reece F. N., Lott B. D., Deaton J. W. The effects of hammermill screen size on ground corn particle size, pellet size, pellet durability, and broiler performance. Poult. Sci. 1986;65:1257–1261. [Google Scholar]
- Rougiere N., Gomez J., Mignon-Grasteau S., Carre B. Effects of diet particle size on digestive parameters in D+ and D− genetic chicken lines selected for divergent digestion efficiency. Poult. Sci. 2009;88:1206–1215. doi: 10.3382/ps.2008-00408. [DOI] [PubMed] [Google Scholar]
- SAS Institute Inc. SAS/STAT 9.1 User's Guide. Cary, NC: SAS Institute; 2012. [Google Scholar]
- Sacranie A. MS thesis. Biddeford, ME: Univ. of New England; 2006. Dietary and age regulation of antiperistalsis in broiler chickens. [Google Scholar]
- Samu P., Matti N., Palvi P. Digestibility and energy value of cereal-based diets in relation to digesta viscosity and retention time in turkeys and chickens at different ages estimated with different markers. Arch. Anim. Nutr. 2010;64:238–253. doi: 10.1080/17450391003625029. [DOI] [PubMed] [Google Scholar]
- Senkoylu N., Samli H. E., Akyurek H., Okur A. A., Kanter M. Effects of whole wheat with or without xylanase supplementation on performance of layers and digestive organ development. Ital. J. Anim. Sci. 2009;8:155–163. [Google Scholar]
- Stark C.R., Chewning C.G. The effect of sieve agitators and dispersing agent on the method of determining and expressing fineness of feed materials by sieving. Anim. Prod. Sci. 2012;52:69–72. [Google Scholar]
- Svihus B., Hetland H. Ileal starch digestibility in growing broiler chickens fed on a wheat based diet is improved by mashing feeding, dilution with cellulose or whole wheat inclusion. Br. Poult. Sci. 2001;42:633–637. doi: 10.1080/00071660120088461. [DOI] [PubMed] [Google Scholar]
- Taylor R. D., Jones G. P. D. The incorporation of whole grain into pelleted broiler chicken diets. II. Gastrointestinal and digesta characteristics. Br. Poult. Sci. 2004;45:237–246. doi: 10.1080/00071660410001715849. [DOI] [PubMed] [Google Scholar]
- Thomas M., Van der Poe A. F. B. Physical quality of pelleted animal feed. I. Criteria for pellet quality. Anim. Feed Sci. Tech. 1996;61:89–112. [Google Scholar]
- Vogtmann H., Frirter P., Prabuck A. L. A new method of determining metabolizability of energy and digestibility of fatty acids in broiler diets. Br. Poult. Sci. 1975;16:531–534. doi: 10.1080/00071667508416222. [DOI] [PubMed] [Google Scholar]