

Abstract
Purpose
Brown adipose tissue (BAT) contributes to the regulation of non-shivering thermogenesis and adiposity. Increasing BAT has recently attracted much attention as a countermeasure to obesity. Animal studies have shown that prolonged catechin treatment increases uncoupling protein 1, a thermogenic protein in BAT. On the other hand, supportable evidence in human is lacking. Thus, the purpose of this study was to examine whether BAT increases after catechin ingestion in humans.
Methods
Twenty-two healthy young women were given either a catechin-rich (540 mg/day; catechin) or placebo beverage every day for 12 weeks in a double-blind design. BAT density was measured using near-infrared time-resolved spectroscopy (NIRTRS), visceral fat area were measured using magnetic resonance imaging, extramyocellular lipids (EMCL) using proton magnetic resonance spectroscopy, and body fat mass using dual-energy X-ray absorptiometry scans.
Results
BAT density was significantly increased (18.8 %), and EMCL was decreased (17.4 %) after the 12-week ingestion. There was a significant negative correlation between the changes in BAT density and those in EMCL (r = −0.66, P < 0.05). There were no notable changes in other parameters.
Conclusions
In conclusion, prolonged ingestion of a catechin-rich beverage increases the BAT density in parallel with a decrease in EMCL.
Keywords: Near-infrared spectroscopy (NIRS), Noninvasive, Brown adipose tissue (BAT)
Background
Brown adipose tissue (BAT) produces heat during cold exposure and spontaneous overfeeding via upregulation of uncoupling protein 1 (UCP-1), whereas white adipose tissue stores excess energy as triglycerides in mammals (Cannon and Nedergaard 2004). In adult humans, metabolically active BAT has been identified in the supraclavicular and paraspinal regions in radionuclide studies using 18F-fluorodeoxyglucose positron emission tomography combined with computed tomography (FDG-PET/CT) (Cypess et al. 2009; Saito et al. 2009; van Marken Lichtenbelt et al. 2009; Virtanen et al. 2009). BAT is associated with cold-induced thermogenesis (Yoneshiro et al. 2013), glucose tolerance (Chondronikola et al. 2014; Hanssen et al. 2015; Lee et al. 2014; Matsushita et al. 2014), and lipid metabolism (Chondronikola et al. 2016) in humans as well as in animals. Therefore, an increase in the amount of BAT is expected to act as a countermeasure to obesity and obesity-related disease. However, due to the limitations of FDG-PET/CT, such as radiation exposure, it is difficult to conduct longitudinal intervention studies on the effects of an increase in the amount of BAT in humans.
Near-infrared time-resolved spectroscopy (NIRTRS) is a method for noninvasively determining total hemoglobin concentration [total-Hb], which reflects the capillaries vascularity in tissue (Hamaoka et al. 2007). Recently, the [total-Hb] under thermoneutral conditions was positively correlated with BAT activity (SUVmax) estimated by FDG-PET/CT in the supraclavicular region, which potentially contains BAT deposits (r = 0.73) (Nirengi et al. 2015). Considering the abundant vascularity of BAT compared with that of other tissues, our results suggest that [total-Hb] estimated by NIRTRS provides BAT density at the approximately 4-cm3 tissue (Nirengi et al. 2015). Quite recently, our new longitudinal study reported that the [total-Hb] increases with the increasing in the SUVmax evaluated by FDG-PET/CT during repeated thermogenic capsiate intake, which is known to increase BAT activity and mass. The amplitude of the increment of [total-Hb] (46.4 %) and SUVmax (48.8 %) is equivalent (Nirengi et al. 2016), thus confirming the validity of our measurements in longitudinal experimental setups.
Catechin is found in green tea, which has been habitually consumed in Asian countries for a long period of time (Nagao et al. 2009). Functional foods approved by US Food and Drug Administration (FDA) include those rich in catechin (Nagao et al. 2009). It has repeatedly reported that a single ingestion of green tea extract rich in catechins increases energy expenditure in humans (Dulloo et al. 2000; Gosselin and Haman 2013; Hursel and Westerterp-Plantenga 2013). Consistent with the acute thermogenic effect, daily ingestion of green tea catechins results in a small but significant reduction of body fatness (Nagao et al. 2005; Friedrich et al. 2011; Hursel et al. 2011).
Animals studies have also shown that 8 weeks of catechin intake upregulated expression of UCP-1 mRNA in BAT and decreased white adipose tissue mass compared with animals on a normal diet (Nomura et al. 2008). Choo (2003) showed in rodents an increased energy expenditure caused by catechins which was associated with an increase in protein content of BAT and that these effects were absent when the β-adrenoceptor was blocked. Thus, there is a possibility that a daily intake of catechin-rich beverages could increase BAT density in humans.
In the current study, we used NIRTRS to examine the effects of prolonged catechin-rich beverage intake on BAT density and a potential association between changes in BAT density and local and whole-body adiposity in humans.
Methods
Study design
This study was conducted from December 2013 to March 2014, the winter season in Japan. Regional average air temperature and actual daytime duration at the time of measurement were 6.1–8.3 °C and 4.1–5.5 h, respectively. The subjects were given either catechin-rich (540 mg/day; catechin group) or placebo (placebo group) beverages every day for 12 consecutive weeks in a double-blind design.
We measured following parameters for participants in both groups at baseline and after 12 weeks of intervention during the luteal phase predicted from menstrual cycle and last menstrual period: BAT density, body composition (fat mass, percent body fat, lean body mass, and bone mass), visceral fat area (VFA), subcutaneous fat area (SFA), skinfold thickness, circulatory parameters [systolic blood pressure (SBP), diastolic blood pressure (DBP), and heart rate (HR)], and intramyocellular lipid (IMCL) and extramyocellular lipid (EMCL) concentrations. In addition, BAT density also measured at weeks 4, 6, 8, and 10. The subjects were instructed to maintain their usual dietary intake and physical activity. After obtaining all variables throughout the study period, the groups were unblinded before the variables were evaluated. The study design and protocol were approved by the Institutional Review Board of Ritsumeikan University, in accordance with the ethical principles contained in the Declaration of Helsinki. Written informed consent was obtained from all participants. This trial has been registered with the University Hospital Medical Information Network UMIN000019920.
Participants
We recruited 22 healthy female college students by advertising on posters or by direct contact. Students taking any medications were excluded. The participants were randomly allocated to the catechin group or the placebo group by a third partly who did not participate in this study.
Study sample
We used a commercially available catechin-rich beverage. Each plastic (350 mL) of the catechin-rich beverage contained 540.0 mg catechin, whereas bottles of the placebo beverage contained 0 mg catechin. The bottles were masked by using opaque plastic to make them indistinguishable in appearance. The caffeine content per bottle was 80.0 and 45.5 mg in the catechin-rich and placebo beverages, respectively (Table 1). Catechin, catechin gallate, gallocatechin, gallocatechin gallate, epicatechin, epicatechin gallate, epigallocatechin, and epigallocatechin gallate, and caffeine contents in the catechin-rich beverages were similar to those used in the previous study (Nagao et al. 2009). The timing for ingestion of the test beverage was not specified.
Table 1.
Components of the test beverages
| Catechin-rich beverage | Placebo beverage | |
|---|---|---|
| Total catechin (mg) | 540 | 0 |
| Caffeine (mg) | 80 | 45.5 |
| Total energy (kJ) | 0 | 0 |
| Total protein (g) | 0 | 0 |
| Total fat (g) | 0 | 0 |
| Carbohydrate (g) | 3.9 | 0 |
| Sodium (mg) | 35 | 35 |
Outcomes
The primary endpoint was BAT density evaluated by [total-Hb] using NIRTRS. The secondary endpoint was the changes in anthropometric parameters (body composition, fat thickness, and IMCL or EMCL concentration).
Near-infrared time-resolved spectroscopy
The [total-Hb] was measured using NIRTRS (TRS-20; Hamamatsu Photonics K.K., Hamamatsu, Japan) for 5 min at 27 °C by placing the probes on the skin in the supraclavicular region potentially containing BAT deposits and, as a reference, also in the deltoid muscle region, which is separated from the BAT deposits. The distance between the emitter and detector was set at 30 mm (Nirengi et al. 2015).
The tissue was illuminated using a 200-μm core diameter optical fibre by pulsed light generated from picosecond light pulsers at 760, 800, and 830 nm with 100-ps full width at half-maximum, a 5-MHz repetition rate, and an 80-μW average power of each wavelength. The emitted photons penetrated the tissue and were reflected to a 3-mm diameter optical bundle fibre through which they were sent to a photomultiplier tube for single photon detection and a signal processing circuit for time-resolved measurement. Using the non-linear least-squares method, the digitized temporal profile data from an in vitro sample or tissue was fitted with a theoretical temporal profile derived from the analytical solution of photon diffusion theory with a semi-infinite homogeneous model in reflectance mode, convoluted with the instrumental response function so the time response of the instrument itself could be compensated, and the values for absorption coefficient and reduced scattering coefficient at 760, 800, and 830 nm were obtained. Then, the absolute concentrations of [total-Hb] were determined using a least-squares fitting method (Hamaoka et al. 2007). The NIRTRS system provided data every 10 s. The coefficient of variation for repeated measurements of [total-Hb] was 4.9 % (Nirengi et al. 2015).
Anthropometric measurements
The body mass, fat mass, percent body fat, fat-free mass, and bone mass were evaluated by a dual-energy x-ray absorptiometry scan (DXA, Lunar Prodigy; GE Healthcare, Buckinghamshire, UK). The VFA and SFA at the abdominal level of L4–L5 were estimated using 1.5-T magnetic resonance imaging (MRI) (Signa HDxt; GE Healthcare, Buckinghamshire, UK). During DXA measurements, subjects maintained a supine position. Then, a series of transaxial MRI scans of abdominal sections were acquired [field of view 420 × 420 mm, slice thickness 10 mm, echo time (TE) = 7.3 ms, repetition time (TR) = one respiration]. The images were exported and analysed by the same investigator using image analysis software (SliceOmatic 4.3; Tomovision Inc, Magog, Canada).
Subcutaneous fat thickness
B-mode ultrasonographic (SSD-3500SV; Hitachi Aloka Medical Co., Ltd, Tokyo, Japan) subcutaneous fat thickness was measured at the supraclavicular region potentially containing BAT and the deltoid muscle region which is separated from BAT deposits. During ultrasonographic measurements, subjects maintained the same posture as during the NIRTRS measurement (Nirengi et al. 2016).
IMCL and EMCL concentrations
IMCL and EMCL were measured in the vastus lateralis muscle by a 1.5-T proton magnetic resonance spectroscopy (1H-MRS) system (Signa HDxt; GE Healthcare, Buckinghamshire, UK) with an 8-channel body array coil positioned parallel to the main magnetic field. Multislice T1-weighted axial spin-echo images (TR/TE 600/7 × 8 ms, thickness 1 cm, field of view 44 cm, matrix size 512 × 512) were first acquired to guide the positioning of the volume of interest. Thereafter, single-voxel 1H-MRS measurements were made on the right vastus lateralis muscle at the midpoint between the greater trochanter and the knee cleft using the point-resolved spectroscopy sequence (TR/TE 2000/35 ms, 2 × 2 × 2 cm3, 32 acquisitions). The IMCL and EMCL concentrations were derived from the peak areas of the CH2 resonance and are expressed as ratios relative to the unsuppressed water peak area in the same voxel (Mayer et al. 2009). The peak chemical shifts of IMCL and EMCL were adjusted as 1.3 and 1.5 ppm, respectively.
Circulatory parameters
SBP, DBP, and, HR were measured using an automated sphygmomanometer (HBP-9020; Omron Corp., Kyoto, Japan) after resting for 10 min.
Dietary diary and records of intakes
Dietary habits during the preceding month were assessed using a validated brief self-administered diet history questionnaire that contained questions about the consumption frequency of 56 foods and beverages and nine dishes that are commonly consumed in the general Japanese population. Daily intakes of energy, protein, fat, and carbohydrate were calculated at baseline and after 12 weeks (Sugawara et al. 2014).
Daily steps and activity energy expenditure were estimated using pedometers (Omron Health Counter HJ-710IT; Omron Healthcare, Kyoto, Japan), and the mean for the 7 days was evaluated.
Statistical analyses
Data are expressed as mean ± standard deviation (SD). Univariate regression analyses were used to analyse the relationship between changes in BAT density and the EMCL. A two-way analysis of variance with repeated measures was used to test the interaction (group × time) and main effect (group, time). If there was a significant interaction or main effect, the time or group differences of the variables between baseline and after 12-week were analysed using the Paired and unpaired t test, respectively. Values were considered to be statistically significant if P was <0.05. All statistical analyses were performed using SPSS version 19 (Chicago, IL, USA).
Power calculation
A sample of 22 subjects was calculated based on detecting a difference of BAT density at the 12-week follow-up between the intervention and placebo groups, with 80 % power and 5 % significance. The difference (plus SD) was based on a previous study that examined changes in BAT activity with cold exposure in healthy subjects (Yoneshiro et al. 2013). The sample size was calculated using Easy R software (Saitama Medical Center, Jichi Medical University, Saitama, Japan) (Kanda 2013).
Results
We recruited 22 healthy female college students [20.7 ± 2.0 years (mean ± SD), body mass index (BMI) 21.0 ± 1.4 kg/m2]. The subjects were given either a catechin-rich beverage (540 mg/day; catechin group; n = 11) or placebo beverage (placebo group; n = 11) every day for 12 consecutive weeks. One subject was excluded from the catechin group because of influenza. Therefore, data were analysed for 21 participants (Table 2). All anthropometric parameters and blood pressure showed no significant differences between catechin and control groups at baseline or at the end of the 12-week study period (Table 2). There were no significant changes in the physical activity levels and dietary intakes during the intervention (data not shown).
Table 2.
Changes in anthropometric parameters and blood pressure between baseline and after 12 weeks intervention in the catechin and placebo groups
| Variables | Catechin (n = 10) | Placebo (n = 11) | ||
|---|---|---|---|---|
| Baseline | 12 weeks | Baseline | 12 weeks | |
| Age (years) | 21.1 ± 2.0 | 21.1 ± 2.0 | 20.5 ± 2.1 | 20.5 ± 2.1 |
| Body weight (kg) | 55.9 ± 4.2 | 56.5 ± 4.4 | 55.7 ± 6.0 | 55.8 ± 7.8 |
| BMI (kg/m2) | 21.1 ± 1.3 | 21.2 ± 1.5 | 20.9 ± 1.6 | 20.9 ± 1.9 |
| Body fat content (%) | 24.0 ± 3.5 | 24.8 ± 2.9 | 25.8 ± 7.6 | 26.0 ± 8.7 |
| Lean body mass (kg) | 40.5 ± 3.8 | 40.6 ± 3.6 | 38.3 ± 8.2 | 38.2 ± 8.3 |
| Body fat mass (kg) | 12.8 ± 2.1 | 13.4 ± 1.8 | 14.0 ± 5.3 | 14.3 ± 6.3 |
| Bone mass (kg) | 2.6 ± 0.2 | 2.6 ± 0.3 | 2.4 ± 0.5 | 2.4 ± 0.5 |
| VFA (cm2) | 27.6 ± 7.3 | 29.3 ± 11.0 | 30.8 ± 13.2 | 34.2 ± 17.2 |
| SFA (cm2) | 117.2 ± 30.8 | 121.6 ± 29.5 | 124.1 ± 59.5 | 136.3 ± 69.0 |
| Supraclavicular subcutaneous fat thickness (cm) | 0.22 ± 0.02 | 0.23 ± 0.03 | 0.23 ± 0.2 | 0.24 ± 0.02 |
| Deltoid muscle subcutaneous fat thickness (cm) | 0.38 ± 0.06 | 0.36 ± 0.04 | 0.40 ± 0.01 | 0.41 ± 0.10 |
| SBP (mm Hg) | 117 ± 7 | 112 ± 9 | 112 ± 7 | 107 ± 8 |
| DBP (mm Hg) | 66 ± 7 | 66 ± 9 | 59 ± 6 | 61 ± 8 |
| Heart rate (bpm) | 69 ± 10 | 74 ± 7 | 67 ± 8 | 69 ± 8 |
BMI body mass index, VFA visceral fat area, SFA subcutaneous fat area, SBP systolic blood pressure, DBP diastolic blood pressure
There was a significant main group effect on the [total-Hb] in the supraclavicular region (Fig. 1a) but not in the deltoid muscle region, which is separated from the BAT depots (Fig. 1b). There was a significant increase in [total-Hb] between baseline and after 12 weeks in the catechin group (67.9 ± 20.4 vs. 80.6 ± 24.3 μM; P < 0.01), while there was no change in the placebo group (66.6 ± 24.0 vs. 67.7 ± 23.7 μM; P = 0.78). Further, the change in the [total-Hb] in the supraclavicular region during the 12-week period was significantly higher in the catechin group than in the placebo group (Fig. 1c).
Fig. 1.
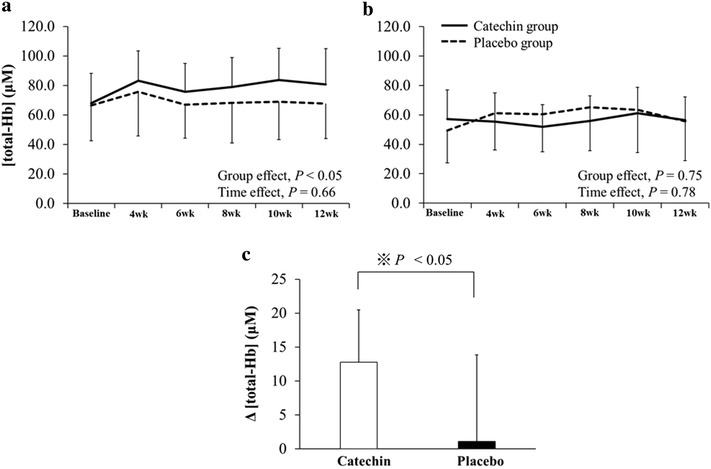
a, b Changes in total haemoglobin concentration [total-Hb] in a the supraclavicular region potentially containing brown adipose tissue and b deltoid muscle region separated from brown adipose tissue deposits. c Changes in [total-Hb] before and after the intervention
There was a significant interaction effect (time × treatment group) on the EMCL concentration, which was significantly decreased by 17.4 % catechin group. There was no significant change in EMCL concentration in the placebo group (Fig. 2a). The IMCL concentration did not change significantly in either the catechin or the placebo group (Fig. 2b).
Fig. 2.
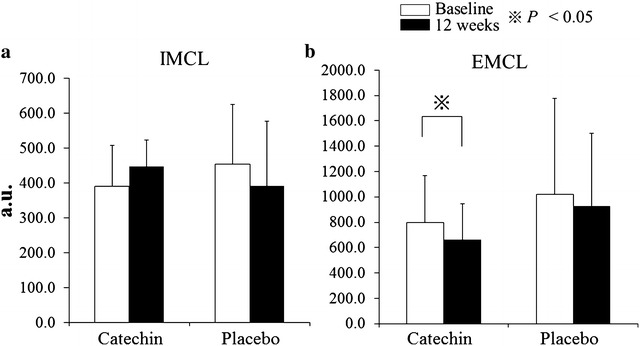
a, b Effect of 12 weeks of catechin or placebo beverage ingestion on the (a) extramyocellular lipid concentration (EMCL) and b intramyocellular lipid (IMCL) concentration. The extramyocellular lipid concentration was significantly decreased in the catechin group after 12 weeks of catechin-rich beverage ingestion (P < 0.05). a.u. arbitrary units
A negative correlation was found between the changes in BAT density and EMCL concentration (r = −0.66, P < 0.05) (Fig. 3).
Fig. 3.
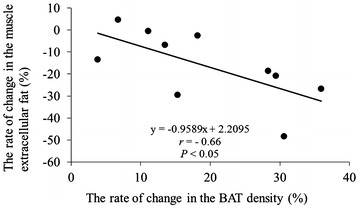
There was a significant negative correlation between the changes in brown adipose tissue (BAT) density evaluated by total haemoglobin concentration [total-Hb] and extramyocellular lipid concentration after catechin-rich beverage ingestion (r = −0.66, P < 0.05)
No apparent harmful incidents were observed in any individuals in the present study.
Discussion
To the best of our knowledge, this is the first study to examine whether a catechin-rich beverage increases BAT density in humans. In this study, the BAT density as evaluated by the [total-Hb] was increased (19 %) only in the catechin group in the supraclavicular region, whereas it did not change in the deltoid muscle of both groups after 12 weeks.
It is reported that there is a considerable intra-individual variance in BAT density, which is even true using a controlled protocol such as chronic cold exposure study (Hanssen et al. 2015; Blondin et al. 2014; van der Lans et al. 2013; Yoneshiro et al. 2013). Thus, the possibility of the type I and II error due to the intra-individual variance in BAT density could influence the results of this study.
It is known that catechins inhibit the catecholamine-degrading enzyme catechol-O-methyltransferease (COMT) in vitro, which results in the sustained effect of NE and thereby the increased lipolysis and BAT activation (Hursel and Westerterp-Plantenga 2013) Catechin- and caffeine-rich teas for control of body weight in humans (Hursel and Westerterp-Plantenga 2013). However, Lorenz et al. (2014) recently reported in humans that COMT activity is not inhibited by high doses of EGCG, indicating a negligible role of COMT in the catechin effects in vivo. Alternatively, an increase in the amount of BAT or activation of BAT is mainly regulated by the increased sympathetic outflow from the hypothalamus to the BAT. It is reported that stimulation of transient receptor potential (TRP) channels is effective for enhancement of BAT thermogenesis and upregulation of UCP-1, a key molecule of BAT thermogenesis (Morrison et al. 2014; Ono et al. 2011). This pathway has been extensively studied in other functional ingredients such as capsinoids in human as well as animals (Saito and Yoneshiro 2013). It was reported that catechin activates TRP channels in cell cultures of taste sensory cells from the gut (Kurogi et al. 2012). Therefore, it is speculated that increasing BAT density found in this study might be due to the gut TRP channel activation. Further investigation is needed to elucidate the detailed mechanism for the increase in BAT density by a catechin-rich beverage intake.
Several different strategies to increase BAT activity and overall BAT mass have been attempted by various groups (Hanssen et al. 2015; Blondin et al. 2014; van der Lans et al. 2013; Cypess et al. 2015; Yoneshiro et al. 2013). These strategies include cold acclimation (Hanssen et al. 2015; Yoneshiro et al. 2013), and repeated functional foods (Nirengi et al. 2016; Yoneshiro et al. 2013; Saito and Yoneshiro 2013) which have both been shown to increase energy expenditure through BAT activation. Acute pharmaceutical drug treatment, such as β3-adrenergic receptor (AR) agonists (Cypess et al. 2015) and bile acid (Broeders et al. 2015), have also been used to increase BAT activity. Cold exposure would be difficult to incorporate into daily life (Yoneshiro et al. 2013) and β3-AR may cause adverse side effects, such as increased blood pressure (Cypess et al. 2015). On the other hand, functional foods have no apparent side effects, unlike pharmaceuticals, and they are easy to incorporate into daily life. Therefore, we used commercially available catechin-rich beverage, green tea, which has the highest catechin concentration in Japan and is approved by the FDA. In addition, this beverage have used in many studies (Nagao et al. 2007, 2009; Hase et al. 2001). The differential caffeine content between catechin and placebo groups is a confounder. Caffeine enhances BAT activity by upregulating intracellular cyclic AMP or by sympathetically releasing NE in vitro (Dulloo et al. 2000). However, a previous study reported that the intake of caffeine (10 mg/kg of body weight), which is approximately 7- to 11-fold of the amount used in the present human study, did not increase FDG uptake in the BAT in rats (Baba et al. 2007). In addition, a tolerance develops to the effects of caffeine (250 mg) on plasma and urinary catecholamines at day 3 in humans (Robertson et al. 1981). Thus, it suggested that caffeine did not influence our results. Nevertheless, we highlight a new practical method for increasing BAT mass by using catechin-rich beverage regardless of the presence or absence of caffeine. However, the increase in BAT mass or activity in observed herein was smaller than that in previous cold exposure studies (37–58 %) (Blondin et al. 2014; van der Lans et al. 2013; Yoneshiro et al. 2013). Further studies are needed to explore functional foods, which are less effective in increasing BAT density compared with cold acclimation.
Previous studies have reported that ingesting a catechin-rich beverage decreased VFA and SFA in obese (BMI 24–30 kg/m2) humans (Nagao et al. 2007) and decreased the waist circumference of patients with type 2 diabetes mellitus (Nagao et al. 2009). Thus, these studies show that catechin decreased body fat. Meanwhile, some studies have reported that there was no significant decrease in body fat after catechin intake (Fukino et al. 2005; Leenen et al. 1992). Hase et al. (2001) reported that subjects with BMI ≥ 25 showed a decrease in body fat, but subjects with BMI < 25 showed no changes in body fat during 12 weeks of catechin ingestion. Although the amount of ingestion and the study period were similar to previous studies (Nagao et al. 2007, 2009), the decrease in body fat was lower in subjects with lower body fatness than in those with higher body fatness (Leenen et al. 1992). Subjects in our study had low BMI (average 21.0 ± 1.4 kg/m2), which is one possible reason why we observed no changes in the whole-body fat parameters despite the increase in BAT density.
In this study, 12 weeks of catechin-rich beverage ingestion led to a significant decrease in EMCL. EMCL has been related to a decrease in insulin sensitivity (Hausman et al. 2014) and an increase in arterial stiffness (Hasegawa et al. 2015), whereas the role of IMCL is as an energy source, such as during acute exercise (Rico-Sanz et al. 2000). Dulloo et al. (1999) reported that treatment with green tea extract resulted in a significant increase in 24-h energy expenditure and a significant decrease in the 24-h respiratory quotient. An increase in energy expenditure may be associated with an increase in β-oxidation in liver (Murase et al. 2002) and muscle (Murase et al. 2006) after catechin ingestion. The molecular mechanism of lipid oxidation from catechin ingestion may, at least in part, be activation of liver kinase B1/AMP-activated protein kinase in various tissues (Murase et al. 2009). Therefore, we considered that an increase in muscle β-oxidation induced by catechin intake may, at least in part, account for the decrease in EMCL. We also speculated in a recent study that in normal cells, catechin might selectively affect the activity of sirtuin 3 (Tao et al. 2015), which regulates mitochondrial fatty acid oxidation (Hirschey et al. 2010). In this study, there was a significant negative correlation between the changes in BAT density and the changes in EMCL. Although there might be a coincident relationship between changes in BAT density and EMCL after 12 weeks intervention and a common pathway might simultaneously activate the two factors, the possibility cannot be ruled out that EMCL decreased due to an unidentified hormonal factor released from the BAT deposits. To our knowledge, this is the first study in which the decrease in EMCL was associated by catechin ingestion. Further studies are required to understand why EMCL showed a decreased response.
There were several limitations in this study. NIRTRS, which is an indirect measurement of BAT density, cannot be used to evaluate the responsiveness of BAT activity to an acute metabolic change such as an acute cold exposure in nature because NIRTRS does not reflect the blood flow, but the blood volume. According to our previous study (Nirengi et al. 2015), the magnitude of the increase in blood volume detectable by NIRTRS is presumably much smaller than that of blood flow velocity during highly metabolic activity. It remains a challenge for future research to confirm validation using 18FDG-PET/CT experiments and monitor energy expenditure such as cold-induced thermogenesis. In this study, we conducted an experiment with a double-blind design. Although some subjects, who may have previously consumed a catechin-rich beverage similar to the one used in this study, might identify the type of beverage based on its taste, we do not believe that knowing its type influenced the physiological consequences. Nonetheless, a study using taste-blind capsules is needed to eliminate completely a placebo effect. Only young female college students were included in this study. Therefore, the effect of catechin-rich beverage ingestion on obese or elderly individuals with a lower BAT content remains to be elucidated.
Conclusions
In conclusion, our results indicate that repeated ingestion of a catechin-rich beverage increases BAT density evaluated by NIRTRS method with a concomitant decrease in EMCL. Although the degree of increasing in BAT mass was smaller than previous cold exposure studies, the result of this study widens the choice of methods for increasing BAT activity/mass. Daily ingestion of a catechin-rich beverage may be useful as an easier and more convenient treatment than chronic cold exposure once the effectiveness for the increase in BAT. 18FDG-PET/CT experiments is required to extensively support the conclusion of the study.
Authors’ contributions
SN and TH designed the research. SN, SA, TH, YK, KE performed the literature search and analyzed data. SN, SA, TY, NS, MS and TH reviewed the articles for inclusion. SN wrote the manuscript. TY, NS, MS, and TH interpreted data and critically reviewed the manuscript. All authors revised the manuscript and approved the final version. All authors read and approved the final manuscript.
Acknowledgements
This work was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (15H03100 and 16K16572). The authors wish to acknowledge the volunteers who participated in this study.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- AMP
adenosine monophosphate
- BAT
brown adipose tissue
- BMI
body mass index
- DBP
diastolic blood pressure
- DXA
dual-energy X-ray
- TE
echo time
- EMCL
extramyocellular lipid
- FDG-PET/CT
18F-fluorodeoxyglucose positron emission tomography combined with computed tomography
- HR
heart rate
- IMCL
intramyocellular lipid
- MRI
magnetic resonance imaging
- NA
noradrenaline
- NIRTRS
near-infrared time-resolved spectroscopy
- 1H-MRS
proton magnetic resonance spectroscopy
- TR
repetition time
- SFA
subcutaneous fat area
- SBP
systolic blood pressure
- SNS
sympathetic nervous system
- [total-Hb]
total haemoglobin concentration
- TRP
transient receptor potential channels
- UCP-1
uncoupling protein 1
- VFA
visceral fat area
Footnotes
Shinsuke Nirengi, Shiho Amagasa, Toshiyuki Homma, Takeshi Yoneshiro, Saori Matsumiya, Yuko Kurosawa, Naoki Sakane, Kumiko Ebi, Masayuki Saito and Takafumi Hamaoka contributed equally to this work
Contributor Information
Shinsuke Nirengi, Email: snirengi@kyotolan.hosp.go.jp.
Shiho Amagasa, Email: shiho.ama@gmail.com.
Toshiyuki Homma, Email: t-homma@ic.daito.ac.jp.
Takeshi Yoneshiro, Email: yoneshiro@med.hokudai.ac.jp.
Saori Matsumiya, Email: saorim@mukogawa-u.ac.jp.
Yuko Kurosawa, Email: yuko.kurosawa.2011@gmail.com.
Naoki Sakane, Email: nsakane@kyotolan.hosp.go.jp.
Kumiko Ebi, Email: ab@fc.ritsumei.ac.jp.
Masayuki Saito, Email: ms-consa@krf.biglobe.ne.jp.
Takafumi Hamaoka, Phone: +81-3-3351-6141, Email: kyp02504@nifty.com.
References
- Baba S, Tatsumi M, Ishimori T, Lilien DL, Engles JM, Wahl RL. Effect of nicotine and ephedrine on the accumulation of 18F-FDG in brown adipose tissue. J Nucl Med. 2007;48:981–986. doi: 10.2967/jnumed.106.039065. [DOI] [PubMed] [Google Scholar]
- Blondin DP, Labbé SM, Tingelstad HC, Noll C, Kunach M, Phoenix S, Guérin B, Turcotte EE, Carpentier AC, Richard D, Haman F. Increased brown adipose tissue oxidative capacity in cold-acclimated humans. J Clin Endocrinol Metab. 2014;99:E438–E446. doi: 10.1210/jc.2013-3901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broeders EP, Nascimento EB, Havekes B, Brans B, Roumans KH, Tailleux A, Schaart G, Kouach M, Charton J, Deprez B, Bouvy ND, Mottaghy F, Staels B, van Marken Lichtenbelt WD, Schrauwen P. The bile acid chenodeoxycholic acid increases human brown adipose tissue activity. Cell Metab. 2015;22:418–426. doi: 10.1016/j.cmet.2015.07.002. [DOI] [PubMed] [Google Scholar]
- Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004;84:277–359. doi: 10.1152/physrev.00015.2003. [DOI] [PubMed] [Google Scholar]
- Chondronikola M, Volpi E, Børsheim E, Porter C, Annamalai P, Enerbäck S, Lidell ME, Saraf MK, Labbe SM, Hurren NM, Yfanti C, Chao T, Andersen CR, Cesani F, Hawkins H, Sidossis LS. Brown adipose tissue improves whole-body glucose homeostasis and insulin sensitivity in humans. Diabetes. 2014;63:4089–4099. doi: 10.2337/db14-0746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chondronikola M, Volpi E, Børsheim E, Porter C, Saraf MK, Annamalai P, Yfanti C, Chao T, Wong D, Shinoda K, Labbė SM, Hurren NM, Cesani F, Kajimura S, Sidossis LS. Brown adipose tissue activation is linked to distinct systemic effects on lipid metabolism in humans. Cell Metab. 2016;23:1200–1206. doi: 10.1016/j.cmet.2016.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choo JJ. Green tea reduces body fat accretion caused by high-fat diet in rats through beta-adrenoceptor activation of thermogenesis in brown adipose tissue. J Nutr Biochem. 2003;14:671–676. doi: 10.1016/j.jnutbio.2003.08.005. [DOI] [PubMed] [Google Scholar]
- Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng YH, Doria A, Kolodny GM, Kahn CR. Identification and importance of brown adipose tissue in adult humans. N Engl J Med. 2009;360:1509–1517. doi: 10.1056/NEJMoa0810780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cypess AM, Weiner LS, Roberts-Toler C, Franquet Elía E, Kessler SH, Kahn PA, English J, Chatman K, Trauger SA, Doria A, Kolodny GM. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab. 2015;21:33–38. doi: 10.1016/j.cmet.2014.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulloo AG, Duret C, Rohrer D, Girardier L, Mensi N, Fathi M, Chantre P, Vandermander J. Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am J Clin Nutr. 1999;70:1040–1045. doi: 10.1093/ajcn/70.6.1040. [DOI] [PubMed] [Google Scholar]
- Dulloo AG, Seydoux J, Girardier L, Chantre P, Vandermander J. Green tea and thermogenesis: interactions between catechin-polyphenols, caffeine and sympathetic activity. Int J Obes Relat Metab Disord. 2000;24:252–258. doi: 10.1038/sj.ijo.0801101. [DOI] [PubMed] [Google Scholar]
- Friedrich M, Petzke KJ, Raederstorff D, Wolfram S, Klaus S. Acute effects of epigallocatechin gallate from green tea on oxidation and tissue incorporation of dietary lipids in mice fed a high-fat diet. Int J Obes (Lond) 2011;36:735–743. doi: 10.1038/ijo.2011.136. [DOI] [PubMed] [Google Scholar]
- Fukino Y, Shimbo M, Aoki N, Okubo T, Iso H. Randomized controlled trial for an effect of green tea consumption on insulin resistance and inflammation markers. J Nutr Sci Vitaminol. 2005;51:335–342. doi: 10.3177/jnsv.51.335. [DOI] [PubMed] [Google Scholar]
- Gosselin C, Haman F. Effects of green tea extracts on non-shivering thermogenesis during mild cold exposure in young men. Br J Nutr. 2013;110:282–288. doi: 10.1017/S0007114512005089. [DOI] [PubMed] [Google Scholar]
- Hamaoka T, McCully KK, Quaresima V, Yamamoto K, Chance B. Near-infrared spectroscopy/imaging for monitoring muscle oxygenation and oxidative metabolism in healthy and diseased humans. J Biomed Opt. 2007;12:062105. doi: 10.1117/1.2805437. [DOI] [PubMed] [Google Scholar]
- Hanssen MJ, Hoeks J, Brans B, van der Lans AA, Schaart G, van den Driessche JJ, Jörgensen JA, Boekschoten MV, Hesselink MK, Havekes B, Kersten S, Mottaghy FM, van Marken Lichtenbelt WD, Schrauwen P. Short-term cold acclimation improves insulin sensitivity in patients with type 2 diabetes mellitus. Nat Med. 2015;21:863–865. doi: 10.1038/nm.3891. [DOI] [PubMed] [Google Scholar]
- Hase T, Komine Y, Meguro S, Takeda Y, Takahashi H, Matsui Y, Inaoka S, Katsuragi Y, Tokimitsu I, Shimasaki H, Itakura H. Anti-obesity effect of tea catechins in humans. J Oleo Sci. 2001;50:599–605. doi: 10.5650/jos.50.599. [DOI] [Google Scholar]
- Hasegawa N, Kurihara T, Sato K, Homma T, Fujie S, Fujita S, Sanada K, Hamaoka T, Iemitsu M. Intramyocellular and extramyocellular lipids are associated with arterial stiffness. Am J Hypertens. 2015;28:1473–1479. doi: 10.1093/ajh/hpv041. [DOI] [PubMed] [Google Scholar]
- Hausman GJ, Basu U, Du M, Fernyhough-Culver M, Dodson M. Intermuscular and intramuscular adipose tissues: bad vs. good adipose tissues. Adipocyte. 2014;3:241–254. doi: 10.4161/adip.28546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirschey MD, Shimazu T, Goetzman E, Jing E, Schwer B, Lombard DB, Grueter CA, Harris C, Biddinger S, Ilkayeva OR, Stevens RD, Li Y, Saha AK, Ruderman NB, Bain JR, Newgard CB, Farese RV, Jr, Alt FW, Kahn CR, Verdin E. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature. 2010;464:121–125. doi: 10.1038/nature08778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursel R, Westerterp-Plantenga MS. Catechin- and caffeine-rich teas for control of body weight in humans. Am J Clin Nutr. 2013;98:1682S–1693S. doi: 10.3945/ajcn.113.058396. [DOI] [PubMed] [Google Scholar]
- Hursel R, Viechtbauer W, Dulloo AG, Tremblay A, Tappy L, Rumpler W, Westerterp-Plantenga MS. The effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: a meta-analysis. Obes Rev. 2011;12:e573–e581. doi: 10.1111/j.1467-789X.2011.00862.x. [DOI] [PubMed] [Google Scholar]
- Kanda Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013;48:452–458. doi: 10.1038/bmt.2012.244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurogi M, Miyashita M, Emoto Y, Kubo Y, Saitoh O. Green tea polyphenol epigallocatechin gallate activates TRPA1 in an intestinal enteroendocrine cell line, STC-1. Chem Senses. 2012;37:167–177. doi: 10.1093/chemse/bjr087. [DOI] [PubMed] [Google Scholar]
- Lee P, Smith S, Linderman J, Courville AB, Brychta RJ, Dieckmann W, Werner CD, Chen KY, Celi FS. Temperature-acclimated brown adipose tissue modulates insulin sensitivity in humans. Diabetes. 2014;63:3686–3698. doi: 10.2337/db14-0513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leenen R, van der Kooy K, Deurenberg P, Seidell JC, Weststrate JA, Schouten FJ, Hautvast JG. Visceral fat accumulation in obese subjects: relation to energy expenditure and response to weight loss. Am J Physiol. 1992;263:E913–E919. doi: 10.1152/ajpendo.1992.263.5.E913. [DOI] [PubMed] [Google Scholar]
- Lorenz M, Paul F, Moobed M, Baumann G, Zimmermann BF, Stangl K, Stangl V. The activity of catechol-O-methyltransferase (COMT) is not impaired by high doses of epigallocatechin-3-gallate (EGCG) in vivo. Eur J Pharmacol. 2014;740:645–651. doi: 10.1016/j.ejphar.2014.06.014. [DOI] [PubMed] [Google Scholar]
- Matsushita M, Yoneshiro T, Aita S, Kameya T, Sugie H, Saito M. Impact of brown adipose tissue on body fatness and glucose metabolism in healthy humans. Int J Obes (Lond) 2014;38:812–817. doi: 10.1038/ijo.2013.206. [DOI] [PubMed] [Google Scholar]
- Mayer LE, Klein DA, Black E, Attia E, Shen W, Mao X, Shungu DC, Punyanita M, Gallagher D, Wang J, Heymsfield SB, Hirsch J, Ginsberg HN, Walsh BT. Adipose tissue distribution after weight restoration and weight maintenance in women with anorexia nervosa. Am J Clin Nutr. 2009;90:1132–1137. doi: 10.3945/ajcn.2009.27820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison SF, Madden CJ, Tupone D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metab. 2014;19:741–756. doi: 10.1016/j.cmet.2014.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murase T, Nagasawa A, Suzuki J, Hase T, Tokimitsu I. Beneficial effects of tea catechins on diet-induced obesity: stimulation of lipid catabolism in the liver. Int J Obes Relat Metab Disord. 2002;26:1459–1464. doi: 10.1038/sj.ijo.0802141. [DOI] [PubMed] [Google Scholar]
- Murase T, Haramizu S, Shimotoyodome A, Tokimitsu I. Reduction of diet-induced obesity by a combination of tea-catechin intake and regular swimming. Int J Obes (Lond) 2006;30:561–568. doi: 10.1038/sj.ijo.0803135. [DOI] [PubMed] [Google Scholar]
- Murase T, Misawa K, Haramizu S, Hase T. Catechin-induced activation of the LKB1/AMP-activated protein kinase pathway. Biochem Pharmacol. 2009;78:78–84. doi: 10.1016/j.bcp.2009.03.021. [DOI] [PubMed] [Google Scholar]
- Nagao T, Komine Y, Soga S, Meguro S, Hase T, Tanaka Y, Tokimitsu I. Ingestion of a tea rich in catechins leads to a reduction in body fat and malondialdehyde-modified LDL in men. Am J Clin Nutr. 2005;81:122–129. doi: 10.1093/ajcn/81.1.122. [DOI] [PubMed] [Google Scholar]
- Nagao T, Hase T, Tokimitsu I. A green tea extract high in catechins reduces body fat and cardiovascular risks in humans. Obesity (Silver Spring) 2007;15:1473–1483. doi: 10.1038/oby.2007.176. [DOI] [PubMed] [Google Scholar]
- Nagao T, Meguro S, Hase T, Otsuka K, Komikado M, Tokimitsu I, Yamamoto T, Yamamoto K. A catechin-rich beverage improves obesity and blood glucose control in patients with type 2 diabetes. Obesity (Silver Spring) 2009;17:310–317. doi: 10.1038/oby.2008.505. [DOI] [PubMed] [Google Scholar]
- Nirengi S, Yoneshiro T, Sugie H, Saito M, Hamaoka T. Human brown adipose tissue assessed by simple noninvasive near-infrared time-resolved spectroscopy. Obesity (Silver Spring) 2015;23:973–980. doi: 10.1002/oby.21012. [DOI] [PubMed] [Google Scholar]
- Nirengi S, Homma T, Inoue N, Sato H, Yoneshiro T, Matsushita M, Kameya T, Sugie H, Tsuzaki K, Saito M, Sakane N, Kurosawa Y, Hamaoka T. Assessment of human brown adipose tissue density during daily ingestion of thermogenic capsinoids using near-infrared time-resolved spectroscopy. J Biomed Opt. 2016;21:91305. doi: 10.1117/1.JBO.21.9.091305. [DOI] [PubMed] [Google Scholar]
- Nomura S, Ichinose T, Jinde M, Kawashima Y, Tachiyashiki K, Imaizumi K. Tea catechins enhance the mRNA expression of uncoupling protein 1 in rat brown adipose tissue. J Nutr Biochem. 2008;19:840–847. doi: 10.1016/j.jnutbio.2007.11.005. [DOI] [PubMed] [Google Scholar]
- Ono K, Tsukamoto-Yasui M, Hara-Kimura Y, Inoue N, Nogusa Y, Okabe Y, Nagashima K, Kato F. Intragastric administration of capsiate, a transient receptor potential channel agonist, triggers thermogenic sympathetic responses. J Appl Physiology. 2011;110:789–798. doi: 10.1152/japplphysiol.00128.2010. [DOI] [PubMed] [Google Scholar]
- Rico-Sanz J, Moosavi M, Thomas EL, McCarthy J, Coutts GA, Saeed N, Bell JD. In vivo evaluation of the effects of continuous exercise on skeletal muscle triglycerides in trained humans. Lipid. 2000;35:1313–1318. doi: 10.1007/s11745-000-0647-2. [DOI] [PubMed] [Google Scholar]
- Robertson D, Wade D, Workman R, Woosley RL, Oates JA. Tolerance to the humoral and hemodynamic effects of caffeine in man. J Clin Invest. 1981;67:1111–1117. doi: 10.1172/JCI110124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito M, Yoneshiro T. Capsinoids and related food ingredients activating brown fat thermogenesis and reducing body fat in humans. Curr Opin Lipidol. 2013;24:71–77. doi: 10.1097/MOL.0b013e32835a4f40. [DOI] [PubMed] [Google Scholar]
- Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, Nio-Kobayashi J, Iwanaga T, Miyagawa M, Kameya T, Nakada K, Kawai Y, Tsujisaki M. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes. 2009;58:1526–1531. doi: 10.2337/db09-0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugawara N, Yasui-Furukori N, Sato Y, Saito M, Furukori H, Nakagami T, Ishioka M, Kaneko S. Dietary patterns are associated with obesity in Japanese patients with schizophrenia. BMC Psychiatry. 2014;14:184. doi: 10.1186/1471-244X-14-184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tao L, Park JY, Lambert JD. Differential prooxidative effects of the green tea polyphenol, (-)-epigallocatechin-3-gallate, in normal and oral cancer cells are related to differences in sirtuin 3 signaling. Mol Nutr Food Res. 2015;59:203–211. doi: 10.1002/mnfr.201400485. [DOI] [PubMed] [Google Scholar]
- van der Lans AA, Hoeks J, Brans B, Vijgen GH, Visser MG, Vosselman MJ, Hansen J, Jörgensen JA, Wu J, Mottaghy FM, Schrauwen P, van Marken Lichtenbelt WD. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J Clin Invest. 2013;123:3395–3403. doi: 10.1172/JCI68993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, Schrauwen P, Teule GJ. Cold-activated brown adipose tissue in healthy men. N Engl J Med. 2009;360:1500–1508. doi: 10.1056/NEJMoa0808718. [DOI] [PubMed] [Google Scholar]
- Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, Taittonen M, Laine J, Savisto NJ, Enerbäck S, Nuutila P. Functional brown adipose tissue in healthy adults. N Engl J Med. 2009;360:1518–1525. doi: 10.1056/NEJMoa0808949. [DOI] [PubMed] [Google Scholar]
- Yoneshiro T, Aita S, Matsushita M, Kayahara T, Kameya T, Kawai Y, Iwanaga T, Saito M. Recruited brown adipose tissue as an antiobesity agent in humans. J Clin Invest. 2013;123:3404–3408. doi: 10.1172/JCI67803. [DOI] [PMC free article] [PubMed] [Google Scholar]