

Abstract
Pediatric high-grade gliomas (pHGGs) are aggressive brain tumors affecting children, and outcomes have remained dismal, even with access to new multimodal therapies. In this study, we compared the miRNomes and transcriptomes of pediatric low- (pLGGs) and high-grade gliomas (pHGGs) using small RNA sequencing (smRNA-Seq) and gene expression microarray, respectively. Through integrated bioinformatics analyses and experimental validation, we identified miR-137 and miR-6500-3p as significantly downregulated in pHGGs. miR-137 or miR-6500-3p overexpression reduced cell proliferation in two pHGG cell lines, SF188 and UW479. CENPE, KIF14 and NCAPG levels were significantly higher in pHGGs than pLGGs, and were direct targets of miR-137 or miR-6500-3p. Furthermore, knockdown of CENPE, KIF14 or NCAPG combined with temozolomide treatment resulted in a combined suppressive effect on pHGG cell proliferation. In summary, our results identify novel mRNA/miRNA interactions that contribute to pediatric glioma malignancy and represent potential targets for the development of new therapeutic strategies.
Keywords: miR-137, miR-6500-3p, CENPE, KIF14, NCAPG
INTRODUCTION
Gliomas are a heterogeneous group of malignant neoplasms that are thought to arise from glial progenitor cells and astrocytes [1]. They are classified based on the following criteria: increased cellular density, nuclear atypia, mitosis, vascular proliferation and necrosis. The World Health Organization (WHO) classifies pediatric gliomas, the most common pediatric central nervous system tumors, into grades I through IV [2, 3]. Pediatric low-grade gliomas (pLGGs) (grades I and II) account for 30% of all pediatric brain tumors and have a favorable prognosis [2, 4, 5]. However, pediatric high-grade gliomas (pHGGs) (grades III and IV), which account for only 8-10% of all pediatric brain tumors, grow faster and are invasive; this hinders total resection, especially in the case of glioblastoma multiforme (GBM) [2, 5, 6]. Following maximal safe surgical resection of pHGGs, adjuvant fractionated radiotherapy is usually applied in children over three years of age. Chemotherapy (CCG-943, lomustine, vincristine and prednisone) combined with radiation therapy has been demonstrated to prolong survival compared to radiation treatment alone (48% vs. 15%, 5 year progression-free survival) [5], but overall pHGG patient survival rates remain dismal. More recently, concurrent oral administration of temozolomide (TMZ) with radiation has become the first-line treatment for newly diagnosed pHGG patients in many institutions [7]. Still, infiltrating malignant cells that survive initial treatments remain the most challenging issue.
Genetic profiling has provided insight into differences between pHGGs and adult HGGs over the last decade. PTEN alterations, loss of 10q, gain of chromosome 7, EGFR and CDKN2A amplifications and hotspot mutations in IDH1 are less common in pHGGs than in adult HGGs [8-10], while gain of 1q and loss of 1p, 4q, and 16q is more common in pHGGs [9]. PDGFRA is also amplified in pHGGs, especially in irradiation-induced tumors [8]. These findings suggest that the mechanisms associated with malignant gliomagenesis in children are different from those in adults.
MicroRNAs (miRNAs) play important roles in the modulation of tumorigenesis. A previous study showed the presence of 23 upregulated and 17 downregulated miRNAs in pediatric gliomas compared to normal brain tissue [11]. These 40 differentially expressed miRNAs regulate gene expression of tumor- and nervous system-related pathways. Miele,et al. showed that the miR-17∼92 cluster is overexpressed in pHGGs compared to adult HGGs and targets the important tumor suppressor genes, PTEN and RB1 [12].
In the present study, we analyzed the miRNomes and transcriptomes of pLGGs and pHGGs using small RNA high-throughput sequencing (smRNA-seq) and gene expression microarray analysis, respectively. We found that two miRNAs downregulated in pHGGs, miR-137 and miR-6500-3p,decreased proliferation of pHGG cells. We identified three genes, KIF14, NCAPG and CENPE that were upregulated in pHGGs and were direct miR-137 or miR-6500-3p targets. CENPE, KIF14 or NCAPG knockdown combined with TMZ treatment suppressed proliferation in pHGG cells. Our findings provide potential new targets for the development of improved therapeutic strategies.
RESULTS
Clinical characterization of the pediatric gliomas
We reviewed a total of 416 pediatric glioma cases that had been diagnosed and treated at Taipei Veterans General Hospital between 1971 and 2013(Figure S1A & S1B). About two-thirds (282/416, 67.8%) were pLGGs and about one-third (134/416, 32.2%) were pHGGs. Pilocytic astrocytomas (125/282, 44.3%) and anaplastic astrocytomas (68/134, 50.7%) were the most common tumor types among pLGGs and pHGGs, respectively. The mean age of pLGG patients was similar to that of pHGG patients (7.7 vs. 8.9 years old) (Table 1). Overall, pHGG patients (grades III & IV) had poorer overall survival than pLGG patients (grades I & II) using Kaplan-Meier survival curves (Figure 1A and S1C). The 2- and 5-year overall survival rates were 99.2% and 98.5%, 94.7% and 94.0%, 55.4% and 44.6% and 38.3% and 30.0% for grade I, II, III and IV gliomas, respectively (p < 0.05).
Table 1. Overview of the pediatric patients with LGG and HGG.
| LGG | HGG | |
|---|---|---|
| Number | 282 | 134 |
| Age | ||
| < 3 years | 55 | 22 |
| 3-10 years | 129 | 54 |
| > 10 years | 98 | 58 |
| mean | 7.7 years | 8.9 years |
| range | 1 days −18.8 years | 10 days −18.7 years |
| Gender | ||
| male | 146 | 71 |
| female | 136 | 63 |
Figure 1. Small RNA sequencing (smRNA-seq) analysis of pediatric gliomas.
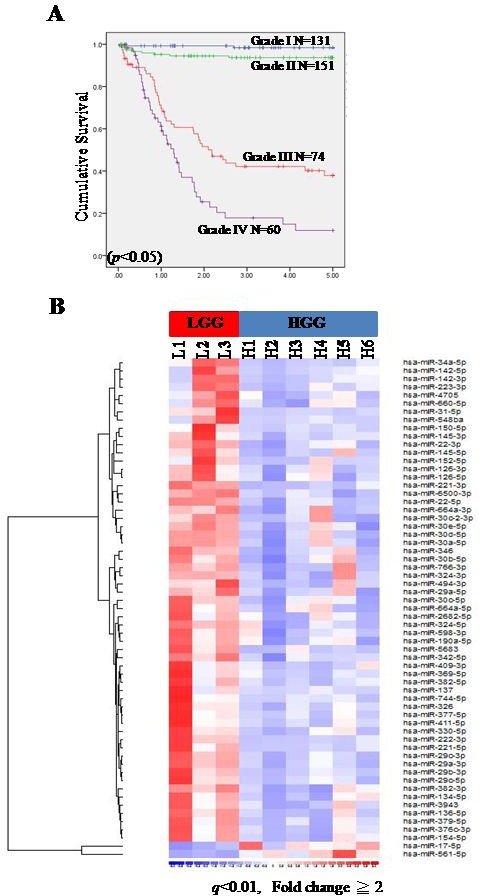
Five-year Kaplan-Meier survival curves for pediatric glioma patients at Taipei Veterans General Hospital from 1971-2013 A. Overall survival time was calculated as the time from diagnosis to death or to last follow-up. Blue line: grade I (LGG); green line: grade II (LGG); red line: grade III (HGG); purple line: grade IV (HGG). A heat map illustrating differentially expressed miRNAs between pLGGs (n = 3) and pHGGs (n = 6) B. A total of 59 miRNAs were upregulated in pLGGs and 2 were upregulated in pHGGs.
miR-137 and miR-6500-3p downregulation in pHGGs increased cell proliferation
To identify miRNome differences between pLGG and pHGG, the global miRNA expression patterns of three pLGGs and six pHGGs were investigated by small RNA sequencing (smRNA-Seq) [13]. We identified 61 known miRNAs as differentially expressed between pLGGs and pHGGs (p < 0.05; Reads per kilo base of exon model per million mapped reads (RPKM)≥2 in at least 2/3 pLGG and pHGG cases) (Figure 1B, Table S1). In particular, miR-137 andmiR-6500-3p were most significantly downregulated in pHGGs
pHGGs have higher mitotic activity than pLGGs [6]. miR-137 was shown to decrease tumor proliferation in various cancers [14-16], and could act as a tumor suppressor. Prior to our study, the biological function of miR-6500-3p was unknown. We investigated the roles of miR-137 and miR-6500-3p in the regulation of cell proliferation in the pHGG cell lines, SF188 and UW379. We found that miR-137 and miR-6500-3p had the lowest expression ratio between pHGGs and pLGGs (pHGG/pLGG) in 24 clinical samples (Figure 2A). Overexpression of miR-137 ormiR-6500-3p decreased cell proliferation in transfected cells (Figure 2B-2C and S2A-B). Knockdown of miR-137 in Res259 cells, a pLGG cell line, increased cell proliferation rates (Figure 2D). We also found that miR-137 or miR-6500-3p overexpression increased accumulation of the G1 phase cell fraction (Figure 2E and S2C), while miR-137 knockdown in Res259 cells resulted in a decreased G1 fraction.
Figure 2. miR-137 and miR-6500-3p suppress cell proliferation and have a combined effect with TMZ treatment.
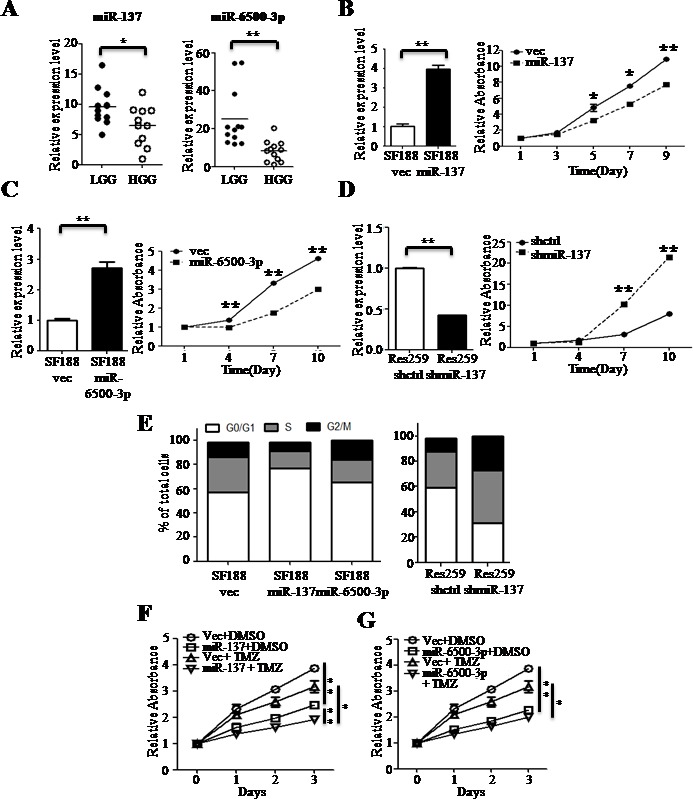
RT-qPCR confirmed downregulation of miR-137 and miR-6500-3p expression in pHGGs (n = 12) compared to pLGGs (n = 12) A. Overexpression of miR-137 B. or miR-6500-3p C. decreases cell proliferation in SF188 cells, and miR-137 knockdown D. increases cell proliferation in Res259 cells as measured by MTT assay. The effects of overexpression were evaluated by RT-qPCR. The effects of miR-137 or miR-6500-3p overexpression on SF188 cells (left panel) and of miR-137 knockdown on Res259 cells (right panel) were evaluated by flow cytometry E. SF188 cells overexpressing miR-137 F. or miR-6500-3p G. were treated with either DMSO or TMZ and cell proliferation was measured at different time points by MTT assay. Results are presented as mean±SD for duplicate samples. *p < 0.05, **p < 0.01 by t-test.
Temozolomide (TMZ) is a DNA alkylating agent that can cross the blood-brain barrier (BBB). Clinically, TMZ had been demonstrated to be active against gliomas and to prolong overall survival rates of adult patients [17]. Previous study of pediatric malignant gliomas showed improved progression-free survival in patients with low tumor levels of MGMT receiving alkylation-based chemotherapy (97 out of 109 patients) [18]. MGMT is an O6-methylguanine DNA methyltransferase that removes methyl groups and repairs DNA adducts [17]. We found that overexpression of either miR-137 or miR-6500-3p combined with TMZ treatment decreased cell proliferation rates as compared to control cells treated with TMZ and cells overexpressing miR-137 or miR-6500-3p without TMZ treatment (Figure 2F-2G and S2D-E).
Differentially expressed genes between pLGGs and pHGGs
We explored the differential molecular expression profiles of eight pLGGs and five pHGGs by microarray analysis and found that each type of glioma has unique transcriptome characteristics. A total of 360 probe sets (264 genes) were increased in pHGGs, while another 96 probe sets (76 genes) were increased in LGGs (q < 0.01, Fold change≥2, Figure 3A, Table S2). Gene Ontology analysis showed that most upregulated genes were involved in regulation of the cell cycle (57 genes) or DNA replication/repair (51 genes) (Figure 3B). In contrast, most upregulated genes in pLGGs were involved in cell-to-cell signaling/interaction (17 genes) or molecular transport (12 genes) (Figure 3C).
Figure 3. Gene expression microarray analysis of pediatric gliomas.
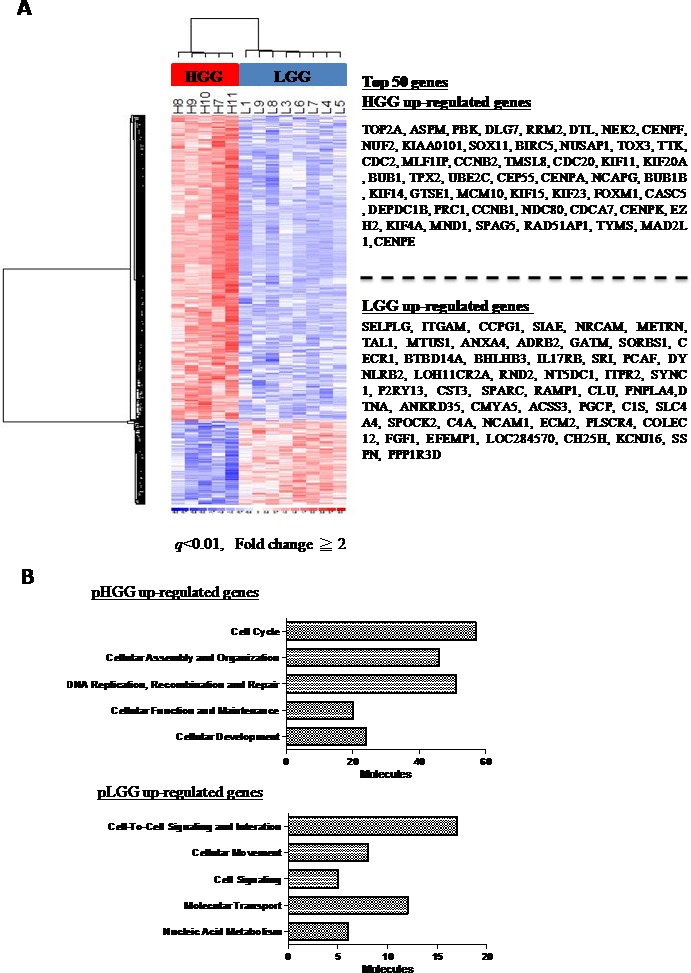
Heat map illustrating the expression of 457 probe sets (q < 0.01, fold change≥2) in five pHGGs and eight pLGGs, with the top 50 genes listed in the right panel A. 264 genes are overexpressed in pHGGs B. and 76 genes are overexpressed in pLGGs C. Genes overexpressed in pHGGs and pLGGs were subjected to Gene Ontology (GO) database searches.
Identifying the pLGG and pHGG miRNA-mRNA interactomes
We used Svmicro and Targetscan (http://www.targetscan.org/) to identify which expressed genes were direct targets of miR-137 and miR-6500-3p, and compared these results with the top 50 upregulated genes in pHGGs. We found that miR-137 and miR-6500-3p targeted six (CASC5, CENPE, DTL, NUF2, SOX11 and TOX3) and five (KIF14, GTSE1, NCAPG, MLF1IP, and SOX11) genes based on Targetscan and SVmicro prediction, respectively (Figure 4A-4B). Gene ontology showed that CASC5, CENPE, NUF2, KIF14, NCAPG and MLF1IP were cell-cycle related genes.
Figure 4. miR-137 and miR-6500-3p target CENPE, KIF14 or NCAPG, which are upregulated in pHGGs.
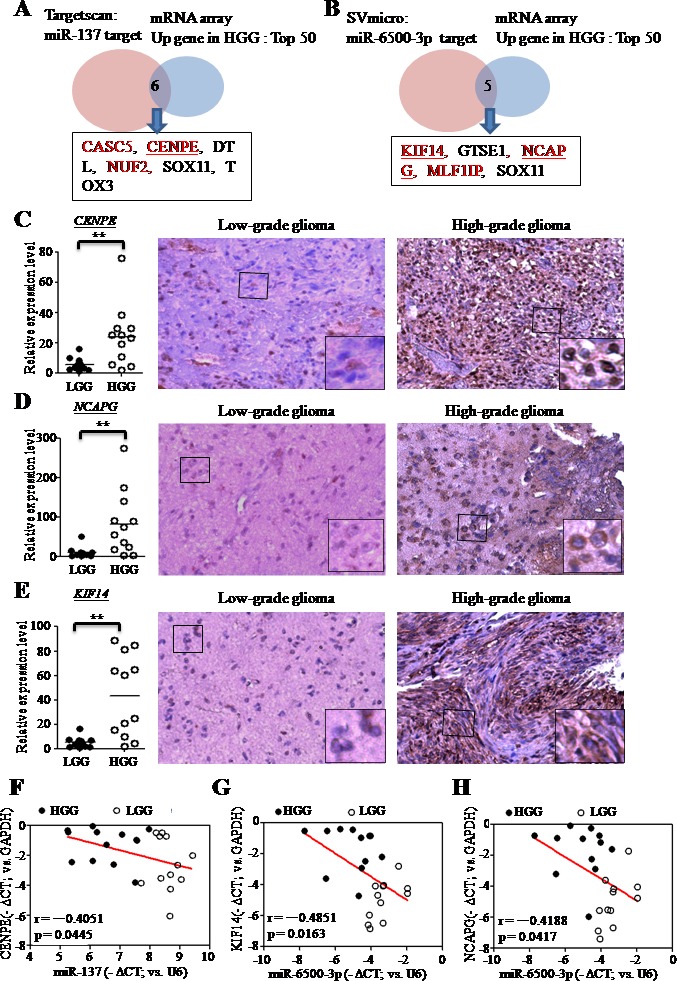
Venn diagram showing overlapping mRNAs based on microarray data (top 50 upregulated mRNAs in pHGGs) and bioinformatics prediction results (Targetscan or SVmicro). The bioinformatic analysis indicates the miR-137 A. and miR-6500-3p B. targets. RT-qPCR and IHC analyses confirmed higher mRNA and protein levels of CENPE C., KIF14 D. and NCAPG E. in pHGGs (mRNA: n = 12; protein: n = 3) than in pLGGs (mRNA: n = 12; protein: n = 3). RT-qPCR results are presented as mean±SD for duplicate samples. **p < 0.01 by t-test. Scatter plots illustrating the negative correlations between miR-137 and CENPE F., miR-6500-3p and KIF14 G., and miR-6500-3p and NCAPG H. in pediatric gliomas.
mRNA and protein expression levels of all genes were assessed using qRT-PCR and IHC staining, which showed that CENPE, KIF14 and NCAPG were potential targets of miR-137 and miR-6500-3p. CENPE (centromere-associated protein-E) is a kinesin-like centromere protein that is highly expressed during mitosis and is required for efficient and stable microtubule capture at the kinetochore [19, 20]. KIF14 (kinesin family member 14) is a kinesin superfamily protein that plays an important role in regulation of the cell cycle and mitotic progression [21]. NCAPG (non-SMC condensin I complex subunit G) is a subunit of the condensin I complex, which plays important roles in meiotic chromosome segregation [22]. mRNA and protein levels of these three genes were increased in pHGG samples (Figure 4C-4E). We re-analyzed other microarray datasets [23] and found that levels of these three genes were also higher in pHGGs compared to pLGGs and normal brain tissue (Figure S3A-D). Analysis in prognoscan (http://www.prognoscan.org/) showed that overexpression of these three genes was associated with poor prognosis (Figure 3E-3G). Correlation coefficients (Pearson's r) of clinical samples indicated negative correlations between CENPE and miR-137 expression (−0.4051), KIF14 and miR-6500-3p expression (−0.4851), and NCAPG and miR-6500-3p expression (−0.4188) (Figure 4F-4G).
We overexpressed miR-137 and miR-6500-3p in SF188 and UW479 cells and knocked-down miR-137 expression in Res259 cells, and analyzed CENPE, KIF14 and NCAPG mRNA and protein levels using qRT-PCR and immunoblotting. CENPE levels were reduced in SF188-miR-137 and UW479-miR-137 cells (Figure 5A) and increased in Res259-anti-miR-137 cells (Figure 5B). KIF14 and NCAPG levels were also decreased in SF188-miR-6500-3p and UW479-miR-6500-3p cells (Figure 5C-5D) as compared to control cells.
Figure 5. miR-137 and miR-6500-3p target directly CENPE, KIF14 or NCAPG .

RT-qPCR and immunoblotting confirmed levels of CENPE A.-B., KIF14 C. or NCAPG D. in SF188 and UW479 cells overexpressing miR-137 A. or miR-6500-3p C.-D., and in Res259 cells with knocked-down miR-137 B. The predicted seed region duplexes formed between CENPE and miR-137 E.(left panel), KIF14 and miR-6500-3p F.(left panel), and NCAPG and miR-6500-3p G.(left panel) are shown. The seed-binding regions with base substitutions in the mutant (Mut) constructs are underlined. The relative luciferase activities of miR-137 or miR-6500-3p, when co-transfected with either the wild type or mutant 3′UTR reporter plasmids (right panels), are shown. Data are presented as mean±SD. *p < 0.05 by t-test.
We used a 3′UTR luciferase reporter assay to validate the direct targeting of CENPE, KIF14 and NCAPG by miR-137 and miR-6500-3p. The putative binding regions of miR-137 and miR-6500-3p within the 3′UTRs of CENPE, KIF14 and NCAPG are shown in Figure 5E-5G. Overexpression of miR-137 and of miR-6500-3p suppressed luciferase activity in wild type CENPE, KIF14 or NCAPG 3′UTR constructs, but this reduction was rescued when mutated CENPE, KIF14 or NCAPG 3′UTR constructs were used (Figure 5E-5G). These findings demonstrated that miR-137 was directly targeted the 3′UTRs of CENPE, and miR-6500-3p directly targeted the 3′UTRs of KIF14 and NCAPG.
CENPE, KIF14 or NCAPG knockdown reduced cell proliferation
The human pHGGs cell lines, SF188 and UW479, were stably transfected with knockdown plasmids targeting CENPE, KIF14 or NCAPG as well as a vector control and cell proliferation was measured by MTT assay. Knockdown of CENPE, KIF14 or NCAPG resulted in a lower proliferation rate compared to the vector control (Figure 6A-6C & S4A-C). CENPE, KIF14 and NCAPG are known to be involved in regulation of the cell cycle. Flow cytometry showed that the knockdown cell lines accumulated cells in the G1 phase of the cell cycle (Figure 6D and S4D). Taken together, our data suggested that CENPE, KIF14 or NCAPG knockdown caused cell cycle arrest in G1 phase and impaired cell proliferation.
Figure 6. CENPE, KIF14 or NCAPG knockdown (KD)reduces cell proliferation and has a combined anti-proliferative effect in the presence of TMZ.
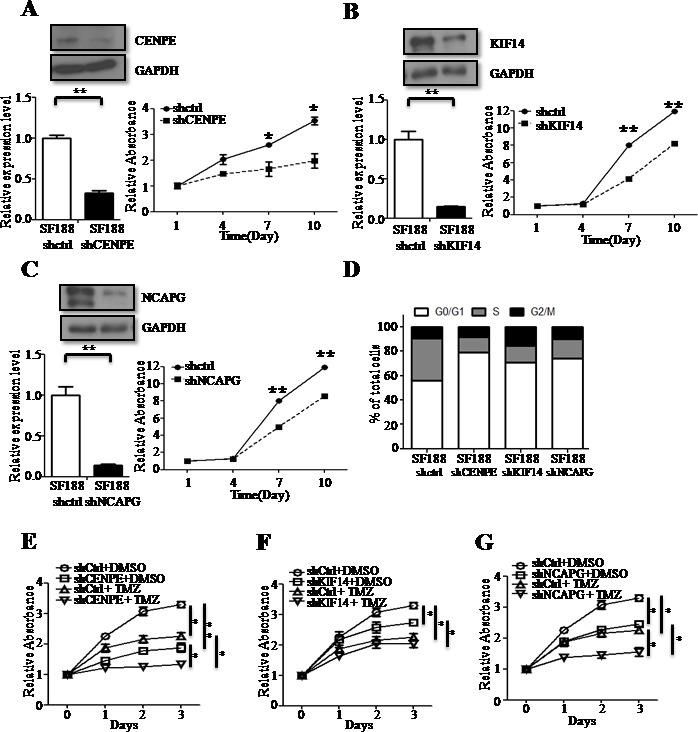
CENPE-KD A., KIF14-KD B. and NCAPG-KD C. decreased cell proliferation rate in SF188 cells asmeasured by MTT assay. KD efficacy was evaluated by RT-qPCR and immunoblotting. CENPE-KD, KIF14-KD and NCAPG-KD SF188 cells were subjected to flow cytometry D. CENPE-KD E., KIF14-KD F. or NCAPG-KD G. SF188 cells were treated with either DMSO or TMZ and cell proliferation was measured at different time points by MTT assay. Results are presented as mean±SD in duplicate samples. *p < 0.05, **p < 0.01 by t-test.
We then treated CENPE, KIF14 or NCAPG knockdown cells with TMZ and measured cell proliferation rates using MTT assay. Proliferation rates were lowest in knockdown cell lines treated with TMZ (TMZ+shCENPE, TMZ+shKIF14 or TMZ+shNCAPG), as compared to control cells (shCtrl), CENPE, KIF14, or NCAPG knockdown cells without TMZ treatment, or cells treated with TMZ alone (TMZ+shCtrl)(Figure 6E-6F and S4E-F).
DISCUSSION
Pediatric gliomas are the most common pediatric brain tumors in Taiwan and worldwide. In pediatric CNS tumors identified at Taipei Veterans General Hospital from 1975 to May 2004(patients less than 18 years of age, n = 986), the most common tumors were astrocytic, which account for 31.1% of all pediatric CNS tumors [24]. Pediatric HGGs, mainly anaplastic astrocytoma and glioblastoma multiforme, are fast growing and infiltrative malignancies that affect every location within the brain. Despite aggressive surgical resection, radiation and adjuvant chemotherapy treatments, the overall survival rate of pHGG patients is still dismal and much lower than that of pLGG patients (Figure 1A and S1C).
In the present study we showed that pLGGs and pHGGs have unique transcriptome characteristics and that many genes involved in the cell cycle are upregulated in pHGGs. Angiogenesis also plays a crucial role in tumor malignancy and progression, and various angiogenesis-related genes and pathways, especially the EGFR/IGFBP2/HIF2αpathway, are upregulated in pHGGs [25]. Our transcriptome analysis revealed upregulation of a number of angiogenesis-related genes, including FGF1, TGFB2, BMP4 and others (p < 0.05). Further validations of angiogenesis-related genes through large-scale analyses are still needed.
Dysregulated miRNAs play important roles in the development and progression of a variety of tumors. Therefore, RNA therapies, such as the application of antagomirs or chemically modified miRNAs, have been proposed as potential cancer treatments [26-29]. The present study showed that there are different miRNA signatures between pLGGs and pHGGs. We demonstrated that miR-137 and miR-6500-3p exhibit anti-tumor activity in pediatric glioma cell lines by blocking CENPE, KIF14 or NCAPG expression. miR-137 is reportedly down-regulated in various cancers, including ovarian cancers, melanoma, colorectal cancers, GBM and others. miR-137 overexpression inhibits cell proliferation, migration and angiogenesis by blocking the expression of multiple target genes [14-16, 30-33]. Previously, no study had explored the role of miR-6500-3p in cancer. We identified a number of novel mRNA/miRNA interactions, namely KIF-14/miR-6500-3p and NCAPG/miR-6500-3p, both of which show potential as novel therapeutic strategies for the treatment of pHGGs. Furthermore, we found that only two miRNAs, miR-17-5p and miR-561-5p, were upregulated in pHGGs. miR-17-5p was shown to enhance tumor metastasis and chemoresistance in various cancers, including colon cancers [34], gastric cancers [35], glioblastoma [36] and prostate cancers [37]. However, the biological function of miR-561-5p is currently unknown, and its involvement in pHGG must be elucidated in future work.
In this study, we demonstrated that knockdown of CENPE, KIF14 and NCAPG expression inhibited cell proliferation in pHGGs. Previous studies suggested that CENPE acts as a tumor suppressor gene or oncogene in various cancers. CENPE overexpression was correlated with cyclin B1 expression and is related to poor prognosis in breast cancers [38]. Conversely, Liu, et al. indicated that CENPE is downregulated in human hepatocellular carcinoma (HCC) tissues [39]. Unlike CENPE, KIF14 acts as an oncogene in various cancers, including lung cancers, ovarian cancers, breast cancers and adult gliomas [40-45]. Previous reports showed that KIF14 knockdown results in cell apoptosis and reduced colony formation [40, 41]. KIF14 overexpression has also been associated with a poor outcome in lung cancers, ovarian cancers, breast cancers and adult gliomas [40-43]. NCAPG in cancers has been less well studied, but it is overexpressed in melanomas [46] and downregulated in out-of-niche primary tumor cells from multiple myelomas and acute myeloid leukemias [47].
This study demonstrated that CENPE, KIF14 or NCAPG knockdown combined with TMZ treatment had a combined suppressive effect on cell proliferation. The CENPE inhibitor, GSK923295, has entered phase I clinical trials and has been found to have only mild side effects in one-third of patients [48]. GSK923295 has been demonstrated to inhibit CENPE motor activity and to cause growth arrest in vitro and in vivo [49]. Recently, pulsed high-intensity focused ultrasound (HIFU) was demonstrated to temporally disrupt the BBB. A combination of pulsed HIFU and atherosclerotic plaque-specific peptide-1 (AP-1)-conjugated liposomes that contain doxorubicin and that specifically bind to the interleukin 4 receptor (IL-4R) worked to elevate the tumor-to-brain drug ratio in a xenografted GBM model [50-52]. A combination of new therapies, such as GSK923295 and pulsed HIFU, could enhance the effect of TMZ and provide improved pHGG cytoreduction strategies.
In summary, integrated transcriptome analysis demonstrated that cell cycle-related genes, specifically CENPE, KIF14 and NCAPG, are upregulated in pHGGs and contribute to cell proliferation. Our study found that CENPE expression is inhibited by miR-137, while KIF14 and NCAPG expression are inhibited by miR-6500-3p, and an anti-tumor effect was achieved by combining TMZ with either miR137 or miR-6500-3p. The unique transcriptomes and miRNomes of pHGGs and pLGGs analyzed in this study provide insight into the mechanisms of pediatric gliomagenesis and could aid in the development of new anti-tumor therapeutic strategies.
MATERIALS AND METHODS
Biological samples
This study was approved by the Institutional Ethics Committee/Institutional Review Board of Taipei Veterans General Hospital (VGH-TPE). The parents/legal guardians of patients in this study provided informed consent (VGHIRB No.: 2013-07-016A, 2013-01-020B&2014-05-006C). Fresh tumor tissues were removed during surgery and then snap-frozen, followed by storage in liquid nitrogen until RNA extraction. Overall survival time was calculated as the time from diagnosis or surgery until death or last follow-up appointment. Differences in survival times were assessed using the log-rank test.
Cell cultures and plasmids
Pediatric glioma cell lines used in this study were kindly provided by Pediatric Oncology, the Institute of Cancer Research, Sutton, United Kingdom [53], and included Res259 (pediatric astrocytoma, Grade II), UW479 (pediatric anaplastic astrocytoma, Grade III) and SF188 (pediatric glioblastoma multiforme, Grade IV). Glioma cells and embryonic kidney 293T cells were cultured in Dulbecco's Modified Eagle Medium-F12 (DMEM-F12) and Dulbecco's Modified Eagle Medium (DMEM)(GIBCO, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS) (GIBCO) in 5% CO2 at 37°C, respectively.
shCENPE, shKIF14, shNCAPG and a control plasmid were obtained from the RNAi consortium at Academia Sinica, Taiwan. For miRNA expression using a plasmid, human miR-137 and miR-6500-3p were PCR-amplified from genomic DNA (Table S3) and cloned into the pLenti4 expression vector (Invitrogen). miRZip-137, which allows for stable expression of anti-microRNA-137(a lentivirus-based plasmid), was purchased from System Biosciences (SBI). All 3′UTR reporter plasmids were PCR-amplified from human genomic DNA (Table S3) and cloned into the pGL3-Basic reporter plasmid (Promega).
Microarray and computational analyses
The AffymetrixTM HG-U133 Plus 2.0 whole genome array was applied as described previously [54]. Principal component analysis (PCA) was performed using Partek Genomics Suite software (http://www.partek.com). A Gene Ontology database search was performed using Ingenuity Pathway Analysis (http://www.ingenuity.com/products/ipa).
Small RNA sequencing (smRNA-Seq) and data analysis
Total RNA sample preparation, sequencing and data analysis were performed as described previously [55]. Briefly, total RNA was collected and small RNA fractions were sequenced using an Illumina HiSeq2000 sequencer (Illumina, San Diego, CA USA) according to manufacturer's instructions. Raw sequencing reads in fastq format were subjected to our in-house bioinformatics pipelines for miRNA profiling and discovery [13].
Quantitative real-time reverse transcription-PCR (qRT-PCR)
Total RNA was isolated from fresh-frozen tumor samples and cells using the DNeasy Blood & Tissue and RNAeasy Kits, respectively (Qiagen, GmbH, Germany). Next, 1 μg of total RNA was used to perform reverse transcription (RT) with a RevertAid™ Reverse transcriptase kit (Cat. K1622; Fermentas, Glen Burnie, Maryland, USA). For qPCR, paired-primers were used to detect specific genes and miRNAs (Table S3).
Immunoblotting
Knockdown or miRNA expression plasmids were introduced into pediatric glioma cell lines using lentiviruses. After 48 h, cells were harvested and lysed using NET lysis buffer containing protease and phosphatase inhibitors. Immunoblotting was carried out using anti-CENPE (1:1000, OriGene Technologies), anti-KIF14 (1:5000, Sigma-Aldrich) and anti-NCAPG (1:5000, Sigma-Aldrich) antibodies, followed by visualization with horseradish peroxidase-conjugated secondary antibodies and an enhanced chemiluminescence detection system (Millipore).
Proliferation assay
Pediatric glioma cells were seeded in 24-well plates at 2×103 cells per well. After 1, 4, 7 and 10 days, cells were treated with 1 ml of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) in phosphate buffered saline (PBS) for 2 h. Purple formazan was solubilized in0.5 ml isopropanol containing 0.1% sodium dodecyl sulfate (SDS). One-hundred μl of solution was transferred into a 96-well plate and optical density was measured by a spectrophotometer at 570 nm.
Cell cycle analysis
5×105 cells were fixed in cold 70% alcohol overnight at −20°C, and then treated with 0.1% TritonX-100 containing 0.2 mg/ml RNase for 1.5 hat 37°C. Cells were then washed with PBS, stained with propidium iodide (PI) for 30 min and analyzed on a FACSCanto (BD Pharmingen). Results were processed using FlowJo.
In vitro drug sensitivity assay
Pediatric glioma cells were seeded at 5×104 cells per well in DMEM-F12 supplemented with 10% FBS into a 24-well plate. After 24 h, medium was removed and new medium containing 250 μM temozolomide (CAS_No. 85622931; Sigma) was added to the test group (day 0). After drug treatment for various times (24, 48, 72h), cells were incubated with 500μl of 0.1 mg/ml MTT in PBS for 2 h. Purple formazan was then solubilized using 1 ml of isopropanol containing 0.1% sodium dodecyl sulfate (SDS) and the optical density was measured by spectrophotometer at 570 nm.
Reporter assay
The microRNA expression plasmids (500 ng), the reporter plasmid (100 ng), and the renilla luciferase plasmid (10 ng) were co-transfected into 1×105 HEK293T cells using the TurboFect transfection reagent (Thermo Scientific). Luciferase activity was measured using a luminometer (model LB593; Berthod, Bad Wildbad, Germany). All assays were performed in duplicate and transfection experiments were repeated at least three times. Firefly luciferase activity was normalized against Renilla luciferase activity.
Immunohistochemistry
IHC sample preparation and staining were performed as described previously [54]. Antibodies used were anti-CENPE (1:500, Sigma-Aldrich), anti-KIF14 (1:200, Sigma-Aldrich) and anti-NCAPG (1:50, Sigma-Aldrich).
Statistical analysis
Two-tailed Student's t-testswere used to assess the significance of mean differences. Differences were considered significant at a p≤0.05. The relationships of microRNAs and their target genes were evaluated with Pearson's correlation coefficients r.
SUPPLEMENTARY MATERIAL FIGURES AND TABLES
Acknowledgments
The authors would like to dedicate this paper to the memory of Dr. Hsei-Wei Wang, who passed away during the period of this research. Dr. Wang originated the rational design of the pediatric glioma genomic study; without his long-term devotion to pediatric brain tumor research, this paper could not have been completed.We also thank the Clinical Research Core Laboratory of Taipei Veterans General Hospital for providing experimental space and facilities.
Footnotes
CONFLICTS OF INTEREST
The authors declare no competing financial or non-financial interests.
GRANT SUPPORT
The array services were provided by the Microarray & Gene Expression Analysis Core Facility of the National Yang-Ming University Genome Research Center, which is supported by the National Research Program for Genomic Medicine, National Science Council (NSC).
This work was supported by the Ministry of Science and Technology [MOST; 103-2911-I-010-506, 102-2314-B-010-045, 102-2314-B-075-033, 103-2314-B-075-035 and 102-2314-B-038-060-MY3], Veterans General Hospitals University System of Taiwan (VGHUST) Joint Research Program, Tsou's Foundation [VGHUST103-G7-2-3], National Health Research Institutes [NHRI-EX103-10254SI], Taipei City Hospital [10401-62-014], National Science Council [102-2911-I-010-503, 101-2320-B-010-059-MY3], National Yang-Ming University [Ministry of Education, Aim for the Top University Plan], Veterans General Hospital Grant [V102-C-126], Taipei Medical University Hospital Grant [No.104 TMU-TMUH-11] and Health and Welfare Surcharge of Tobacco Products [MOHW105-TDU-B-212-134-001]. This work was also supported in part by the UST-UCSD International Center for Excellence in Advanced Bioengineering sponsored by the Taiwan MOST I-RiCE Program [103-2911-I-009-101].
Editorial Note
This paper has been accepted based in part on peerreview conducted by another journal and the authors’ response and revisions as well as expedited peer-review in Oncotarget.
REFERENCES
- 1.Mischel PS, Cloughesy TF, Nelson SF. DNA-microarray analysis of brain cancer: molecular classification for therapy. Nature reviews Neuroscience. 2004;5:782–792. doi: 10.1038/nrn1518. [DOI] [PubMed] [Google Scholar]
- 2.Minturn JE, Fisher MJ. Gliomas in children. Current treatment options in neurology. 2013;15:316–327. doi: 10.1007/s11940-013-0225-x. [DOI] [PubMed] [Google Scholar]
- 3.Figarella-Branger D, Colin C, Coulibaly B, Quilichini B, Maues De Paula A, Fernandez C, Bouvier C. [Histological and molecular classification of gliomas] Revue neurologique. 2008;164:505–515. doi: 10.1016/j.neurol.2008.03.011. [DOI] [PubMed] [Google Scholar]
- 4.Sievert AJ, Fisher MJ. Pediatric low-grade gliomas. Journal of child neurology. 2009;24:1397–1408. doi: 10.1177/0883073809342005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hargrave D. Paediatric high and low grade glioma: the impact of tumour biology on current and future therapy. British journal of neurosurgery. 2009;23:351–363. doi: 10.1080/02688690903158809. [DOI] [PubMed] [Google Scholar]
- 6.Fangusaro J. Pediatric high grade glioma: a review and update on tumor clinical characteristics and biology. Frontiers in oncology. 2012;2:105. doi: 10.3389/fonc.2012.00105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.MacDonald TJ, Aguilera D, Kramm CM. Treatment of high-grade glioma in children and adolescents. Neuro-oncology. 2011;13:1049–1058. doi: 10.1093/neuonc/nor092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pollack IF, Hamilton RL, James CD, Finkelstein SD, Burnham J, Yates AJ, Holmes EJ, Zhou T, Finlay JL, Children's Oncology G Rarity of PTEN deletions and EGFR amplification in malignant gliomas of childhood: results from the Children's Cancer Group 945 cohort. Journal of neurosurgery. 2006;105:418–424. doi: 10.3171/ped.2006.105.5.418. [DOI] [PubMed] [Google Scholar]
- 9.Paugh BS, Qu C, Jones C, Liu Z, Adamowicz-Brice M, Zhang J, Bax DA, Coyle B, Barrow J, Hargrave D, Lowe J, Gajjar A, Zhao W, et al. Integrated molecular genetic profiling of pediatric high-grade gliomas reveals key differences with the adult disease. Journal of clinical oncology. 2010;28:3061–3068. doi: 10.1200/JCO.2009.26.7252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rood BR, MacDonald TJ. Pediatric high-grade glioma: molecular genetic clues for innovative therapeutic approaches. J Neurooncol. 2005;75:267–272. doi: 10.1007/s11060-005-6749-5. [DOI] [PubMed] [Google Scholar]
- 11.Liu F, Xiong Y, Zhao Y, Tao L, Zhang Z, Zhang H, Liu Y, Feng G, Li B, He L, Ma J, Qin S, Yang Y. Identification of aberrant microRNA expression pattern in pediatric gliomas by microarray. Diagnostic pathology. 2013;8:158. doi: 10.1186/1746-1596-8-158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Miele E, Buttarelli FR, Arcella A, Begalli F, Garg N, Silvano M, Po A, Baldi C, Carissimo G, Antonelli M, Spinelli GP, Capalbo C, Donofrio V, et al. High-throughput microRNA profiling of pediatric high-grade gliomas. Neuro Oncol. 2014;16:228–240. doi: 10.1093/neuonc/not215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cheng WC, Chung IF, Tsai CF, Huang TS, Chen CY, Wang SC, Chang TY, Sun HJ, Chao JY, Cheng CC, Wu CW, Wang HW. YM500v2: a small RNA sequencing (smRNA-seq) database for human cancer miRNome research. Nucleic acids research. 2015;43(Database issue):D862–867. doi: 10.1093/nar/gku1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hao S, Luo C, Abukiwan A, Wang G, He J, Huang L, Weber CE, Lv N, Xiao X, Eichmuller SB, He D. miR-137 inhibits proliferation of melanoma cells by targeting PAK2. Experimental dermatology. 2015 doi: 10.1111/exd.12812. [DOI] [PubMed] [Google Scholar]
- 15.Sun J, Zheng G, Gu Z, Guo Z. MiR-137 inhibits proliferation and angiogenesis of human glioblastoma cells by targeting EZH2. Journal of neuro-oncology. 2015;122:481–489. doi: 10.1007/s11060-015-1753-x. [DOI] [PubMed] [Google Scholar]
- 16.Zhang L, Li Z, Gai F, Wang Y. MicroRNA-137 suppresses tumor growth in epithelial ovarian cancer in vitro and in vivo. Molecular medicine reports. 2015;12:3107–3114. doi: 10.3892/mmr.2015.3756. [DOI] [PubMed] [Google Scholar]
- 17.Barone G, Maurizi P, Tamburrini G, Riccardi R. Role of temozolomide in pediatric brain tumors. Childs Nerv Syst. 2006;22:652–661. doi: 10.1007/s00381-006-0081-z. [DOI] [PubMed] [Google Scholar]
- 18.Pollack IF, Hamilton RL, Sobol RW, Burnham J, Yates AJ, Holmes EJ, Zhou T, Finlay JL. O6-methylguanine-DNA methyltransferase expression strongly correlates with outcome in childhood malignant gliomas: results from the CCG-945 Cohort. Journal of clinical oncology. 2006;24:3431–3437. doi: 10.1200/JCO.2006.05.7265. [DOI] [PubMed] [Google Scholar]
- 19.Tanudji M, Shoemaker J, L'Italien L, Russell L, Chin G, Schebye XM. Gene silencing of CENP-E by small interfering RNA in HeLa cells leads to missegregation of chromosomes after a mitotic delay. Molecular biology of the cell. 2004;15:3771–3781. doi: 10.1091/mbc.E03-07-0482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wood KW, Chua P, Sutton D, Jackson JR. Centromere-associated protein E: a motor that puts the brakes on the mitotic checkpoint. Clinical cancer research. 2008;14:7588–7592. doi: 10.1158/1078-0432.CCR-07-4443. [DOI] [PubMed] [Google Scholar]
- 21.Gruneberg U, Neef R, Li X, Chan EH, Chalamalasetty RB, Nigg EA, Barr FA. KIF14 and citron kinase act together to promote efficient cytokinesis. The Journal of cell biology. 2006;172:363–372. doi: 10.1083/jcb.200511061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hirano T. Condensins: organizing and segregating the genome. Current biology : CB. 2005;15:R265–275. doi: 10.1016/j.cub.2005.03.037. [DOI] [PubMed] [Google Scholar]
- 23.Gump JM, Donson AM, Birks DK, Amani VM, Rao KK, Griesinger AM, Kleinschmidt-DeMasters BK, Johnston JM, Anderson RC, Rosenfeld A, Handler M, Gore L, Foreman N, et al. Identification of targets for rational pharmacological therapy in childhood craniopharyngioma. Acta neuropathologica communications. 2015;3:30. doi: 10.1186/s40478-015-0211-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wong TT, Ho DM, Chang KP, Yen SH, Guo WY, Chang FC, Liang ML, Pan HC, Chung WY. Primary pediatric brain tumors: statistics of Taipei VGH, Taiwan (1975-2004) Cancer. 2005;104:2156–2167. doi: 10.1002/cncr.21430. [DOI] [PubMed] [Google Scholar]
- 25.Khatua S, Peterson KM, Brown KM, Lawlor C, Santi MR, LaFleur B, Dressman D, Stephan DA, MacDonald TJ. Overexpression of the EGFR/FKBP12/HIF-2alpha pathway identified in childhood astrocytomas by angiogenesis gene profiling. Cancer research. 2003;63:1865–1870. [PubMed] [Google Scholar]
- 26.Kitade Y, Akao Y. MicroRNAs and their therapeutic potential for human diseases: microRNAs, miR-143 and -145, function as anti-oncomirs and the application of chemically modified miR-143 as an anti-cancer drug. J Pharmacol Sci. 2010;114:276–280. doi: 10.1254/jphs.10r12fm. [DOI] [PubMed] [Google Scholar]
- 27.Mercatelli N, Coppola V, Bonci D, Miele F, Costantini A, Guadagnoli M, Bonanno E, Muto G, Frajese GV, De Maria R, Spagnoli LG, Farace MG, Ciafrè SA. The Inhibition of the Highly Expressed Mir-221 and Mir-222 Impairs the Growth of Prostate Carcinoma Xenografts in Mice. PloS one. 2008;3:e4029. doi: 10.1371/journal.pone.0004029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bertoli G, Cava C, Castiglioni I. MicroRNAs: New Biomarkers for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Breast Cancer. Theranostics. 2015;5:1122–1143. doi: 10.7150/thno.11543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kaboli PJ, Rahmat A, Ismail P, Ling KH. MicroRNA-based therapy and breast cancer: A comprehensive review of novel therapeutic strategies from diagnosis to treatment. Pharmacological research. 2015;97:104–121. doi: 10.1016/j.phrs.2015.04.015. [DOI] [PubMed] [Google Scholar]
- 30.Smith AR, Marquez RT, Tsao WC, Pathak S, Roy A, Ping J, Wilkerson B, Lan L, Meng W, Neufeld KL, Sun XF, Xu L. Tumor suppressive microRNA-137 negatively regulates Musashi-1 and colorectal cancer progression. Oncotarget. 2015;6:12558–12573. doi: 10.18632/oncotarget.3726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yang Y, Li F, Saha MN, Abdi J, Qiu L, Chang H. miR-137 and miR-197 Induce Apoptosis and Suppress Tumorigenicity by Targeting MCL-1 in Multiple Myeloma. Clin Cancer Res. 2015;21:2399–2411. doi: 10.1158/1078-0432.CCR-14-1437. [DOI] [PubMed] [Google Scholar]
- 32.Bier A, Giladi N, Kronfeld N, Lee HK, Cazacu S, Finniss S, Xiang C, Poisson L, deCarvalho AC, Slavin S, Jacoby E, Yalon M, Toren A, et al. MicroRNA-137 is downregulated in glioblastoma and inhibits the stemness of glioma stem cells by targeting RTVP-1. Oncotarget. 2013;4:665–676. doi: 10.18632/oncotarget.928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu LL, Lu SX, Li M, Li LZ, Fu J, Hu W, Yang YZ, Luo RZ, Zhang CZ, Yun JP. FoxD3-regulated microRNA-137 suppresses tumour growth and metastasis in human hepatocellular carcinoma by targeting AKT2. Oncotarget. 2014;5:5113–5124. doi: 10.18632/oncotarget.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fang L, Li H, Wang L, Hu J, Jin T, Wang J, Yang BB. MicroRNA-17-5p promotes chemotherapeutic drug resistance and tumour metastasis of colorectal cancer by repressing PTEN expression. Oncotarget. 2014;5:2974–2987. doi: 10.18632/oncotarget.1614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wu Q, Luo G, Yang Z, Zhu F, An Y, Shi Y, Fan D. miR-17-5p promotes proliferation by targeting SOCS6 in gastric cancer cells. FEBS letters. 2014;588:2055–2062. doi: 10.1016/j.febslet.2014.04.036. [DOI] [PubMed] [Google Scholar]
- 36.Li H, Yang BB. Stress response of glioblastoma cells mediated by miR-17-5p targeting PTEN and the passenger strand miR-17-3p targeting MDM2. Oncotarget. 2012;3:1653–1668. doi: 10.18632/oncotarget.810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yang X, Du WW, Li H, Liu F, Khorshidi A, Rutnam ZJ, Yang BB. Both mature miR-17-5p and passenger strand miR-17-3p target TIMP3 and induce prostate tumor growth and invasion. Nucleic acids research. 2013;41:9688–9704. doi: 10.1093/nar/gkt680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Agarwal R, Gonzalez-Angulo AM, Myhre S, Carey M, Lee JS, Overgaard J, Alsner J, Stemke-Hale K, Lluch A, Neve RM, Kuo WL, Sorlie T, Sahin A, et al. Integrative analysis of cyclin protein levels identifies cyclin b1 as a classifier and predictor of outcomes in breast cancer. Clin Cancer Res. 2009;15:3654–3662. doi: 10.1158/1078-0432.CCR-08-3293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liu Z, Ling K, Wu X, Cao J, Liu B, Li S, Si Q, Cai Y, Yan C, Zhang Y, Weng Y. Reduced expression of cenp-e in human hepatocellular carcinoma. Journal of experimental & clinical cancer research. 2009;28:156. doi: 10.1186/1756-9966-28-156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Corson TW, Gallie BL. KIF14 mRNA expression is a predictor of grade and outcome in breast cancer. International journal of cancer Journal international du cancer. 2006;119:1088–1094. doi: 10.1002/ijc.21954. [DOI] [PubMed] [Google Scholar]
- 41.Corson TW, Zhu CQ, Lau SK, Shepherd FA, Tsao MS, Gallie BL. KIF14 messenger RNA expression is independently prognostic for outcome in lung cancer. Clin Cancer Res. 2007;13:3229–3234. doi: 10.1158/1078-0432.CCR-07-0393. [DOI] [PubMed] [Google Scholar]
- 42.Theriault BL, Pajovic S, Bernardini MQ, Shaw PA, Gallie BL. Kinesin family member 14: an independent prognostic marker and potential therapeutic target for ovarian cancer. International journal of cancer Journal international du cancer. 2012;130:1844–1854. doi: 10.1002/ijc.26189. [DOI] [PubMed] [Google Scholar]
- 43.Wang Q, Wang L, Li D, Deng J, Zhao Z, He S, Zhang Y, Tu Y. Kinesin family member 14 is a candidate prognostic marker for outcome of glioma patients. Cancer epidemiology. 2013;37:79–84. doi: 10.1016/j.canep.2012.08.011. [DOI] [PubMed] [Google Scholar]
- 44.Theriault BL, Cybulska P, Shaw PA, Gallie BL, Bernardini MQ. The role of KIF14 in patient-derived primary cultures of high-grade serous ovarian cancer cells. Journal of ovarian research. 2014;7:123. doi: 10.1186/s13048-014-0123-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Singel SM, Cornelius C, Zaganjor E, Batten K, Sarode VR, Buckley DL, Peng Y, John GB, Li HC, Sadeghi N, Wright WE, Lum L, Corson TW, et al. KIF14 promotes AKT phosphorylation and contributes to chemoresistance in triple-negative breast cancer. Neoplasia. 2014;16:247–256. doi: 10.1016/j.neo.2014.03.008. 256 e242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ryu B, Kim DS, Deluca AM, Alani RM. Comprehensive expression profiling of tumor cell lines identifies molecular signatures of melanoma progression. PloS one. 2007;2:e594. doi: 10.1371/journal.pone.0000594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cohen Y, Gutwein O, Garach-Jehoshua O, Bar-Haim A, Kornberg A. The proliferation arrest of primary tumor cells out-of-niche is associated with widespread downregulation of mitotic and transcriptional genes. Hematology. 2014;19:286–292. doi: 10.1179/1607845413Y.0000000125. [DOI] [PubMed] [Google Scholar]
- 48.Chung V, Heath EI, Schelman WR, Johnson BM, Kirby LC, Lynch KM, Botbyl JD, Lampkin TA, Holen KD. First-time-in-human study of GSK923295, a novel antimitotic inhibitor of centromere-associated protein E (CENP-E), in patients with refractory cancer. Cancer chemotherapy and pharmacology. 2012;69:733–741. doi: 10.1007/s00280-011-1756-z. [DOI] [PubMed] [Google Scholar]
- 49.Wood KW, Lad L, Luo L, Qian X, Knight SD, Nevins N, Brejc K, Sutton D, Gilmartin AG, Chua PR, Desai R, Schauer SP, McNulty DE, et al. Antitumor activity of an allosteric inhibitor of centromere-associated protein-E. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:5839–5844. doi: 10.1073/pnas.0915068107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yang FY, Wong TT, Teng MC, Liu RS, Lu M, Liang HF, Wei MC. Focused ultrasound and interleukin-4 receptor-targeted liposomal doxorubicin for enhanced targeted drug delivery and antitumor effect in glioblastoma multiforme. Journal of controlled release. 2012;160:652–658. doi: 10.1016/j.jconrel.2012.02.023. [DOI] [PubMed] [Google Scholar]
- 51.Yang FY, Teng MC, Lu M, Liang HF, Lee YR, Yen CC, Liang ML, Wong TT. Treating glioblastoma multiforme with selective high-dose liposomal doxorubicin chemotherapy induced by repeated focused ultrasound. International journal of nanomedicine. 2012;7:965–974. doi: 10.2147/IJN.S29229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yang FY, Wang HE, Liu RS, Teng MC, Li JJ, Lu M, Wei MC, Wong TT. Pharmacokinetic analysis of 111 in-labeled liposomal Doxorubicin in murine glioblastoma after blood-brain barrier disruption by focused ultrasound. PloS one. 2012;7:e45468. doi: 10.1371/journal.pone.0045468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bax DA, Little SE, Gaspar N, Perryman L, Marshall L, Viana-Pereira M, Jones TA, Williams RD, Grigoriadis A, Vassal G, Workman P, Sheer D, Reis RM, et al. Molecular and phenotypic characterisation of paediatric glioma cell lines as models for preclinical drug development. PloS one. 2009;4:e5209. doi: 10.1371/journal.pone.0005209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ho DM, Shih CC, Liang ML, Tsai CY, Hsieh TH, Tsai CH, Lin SC, Chang TY, Chao ME, Wang HW, Wong TT. Integrated genomics has identified a new AT/RT-like yet INI1-positive brain tumor subtype among primary pediatric embryonal tumors. BMC medical genomics. 2015;8:32. doi: 10.1186/s12920-015-0103-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hsieh TH, Chien CL, Lee YH, Lin CI, Hsieh JY, Chao ME, Liu DJ, Chu SS, Chen W, Lin SC, Ho DM, Liu RS, Lin CH, et al. Downregulation of SUN2, a novel tumor suppressor, mediates miR-221/222-induced malignancy in central nervous system embryonal tumors. Carcinogenesis. 2014;35:2164–2174. doi: 10.1093/carcin/bgu105. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.